
Abstract
Significance:
Plant chloroplasts generate reactive oxygen species (ROS) during photosynthesis, especially under stresses. The sulfhydryl groups of protein cysteine residues are susceptible to redox modifications, which regulate protein structure and function, and thus different signaling and metabolic processes. The ROS-governed protein thiol redox switches play important roles in chloroplasts.
Recent Advances:
Various high-throughput thiol redox proteomic approaches have been developed, and they have enabled the improved understanding of redox regulatory mechanisms in chloroplasts. For example, the thioredoxin-modulated antioxidant enzymes help to maintain cellular ROS homeostasis. The light- and dark-dependent redox regulation of photosynthetic electron transport, the Calvin/Benson cycle, and starch biosynthesis ensures metabolic coordination and efficient energy utilization. In addition, redox cascades link the light with the dynamic changes of metabolites in nitrate and sulfur assimilation, shikimate pathway, and biosynthesis of fatty acid hormone as well as purine, pyrimidine, and thiamine. Importantly, redox regulation of tetrapyrrole and chlorophyll biosynthesis is critical to balance the photodynamic tetrapyrrole intermediates and prevent oxidative damage. Moreover, redox regulation of diverse elongation factors, chaperones, and kinases plays an important role in the modulation of gene expression, protein conformation, and posttranslational modification that contribute to photosystem II (PSII) repair, state transition, and signaling in chloroplasts.
Critical Issues:
This review focuses on recent advances in plant thiol redox proteomics and redox protein networks toward understanding plant chloroplast signaling, metabolism, and stress responses.
Future Directions:
Using redox proteomics integrated with biochemical and molecular genetic approaches, detailed studies of cysteine residues, their redox states, cross talk with other modifications, and the functional implications will yield a holistic understanding of chloroplast stress responses.
Introduction
Plants are constantly challenged by abiotic and biotic stresses which lead to the accumulation of reactive molecular species (RMS), including reactive oxygen species (ROS) and reactive nitrogen species (RNS). RMS work as signal molecules and/or oxidizing agents that can affect various cellular components (e.g., nucleic acids, proteins, and lipids) by altering their structure and function (186).
Protein cysteine (Cys) residues are among the most RMS-susceptible sites (Fig. 1) (5, 186). Most Cys thiol oxidation takes place through nonenzymatic reaction by ROS/RNS (176). Exposure of the redox-sensitive Cys thiols to ROS leads to reversible sulfenic acid (-SOH) formation. The highly reactive -SOH is stabilized by forming a disulfide bond (S-S) with a nearby thiol or a mixed disulfide bond with reduced glutathione (GSH) (5, 186). In addition, RNS-mediated S-nitrosylation occurs through a covalent attachment of nitric oxide (NO) to a Cys residue to form S-nitrosothiol (Fig. 1) (36, 48, 65). These reversible redox posttranslational modifications (PTMs) allow Cys to act as a regulatory switch that can alter enzyme activity, conformational integrity, and protein stability in response to cellular redox changes (5, 186).

In the presence of high levels of ROS, some Cys thiols can be irreversibly oxidized to sulfinic acid (-SO2H) and sulfonic acid (-SO3H) through S-sulfinylation and S-sulfonylation, respectively (5, 186) (Fig. 1). In general, -SO2H and -SO3H are irreversible oxidative PTMs (Ox-PTMs), with the exception of the -SO2H moiety of 2-Cys peroxiredoxin (2-Cys Prx), which can be reduced by sulfiredoxin (96). Overall, the irreversible Cys PTMs deactivate protein functions and play a role in protein turnover (5, 186).
Chloroplasts are important organelles for photosynthesis and biosynthesis of diverse metabolites in plants. The light-driven photosynthetic reactions (light reactions) contribute to ROS production, especially under stress conditions. Chloroplasts generate singlet oxygen (1O2) in photosystem II (PSII), superoxide anion radical (O2 •−) at photosystem I (PSI), hydrogen peroxide (H2O2) after O2 •− dismutation (catalyzed by Fe/Zn-superoxide dismutase [Fe/Zn-SOD] and Cu/Zn-SOD), and the highly reactive hydroxyl radical (•OH) derived from H2O2 through the Fenton reaction (Fig. 2) (171).
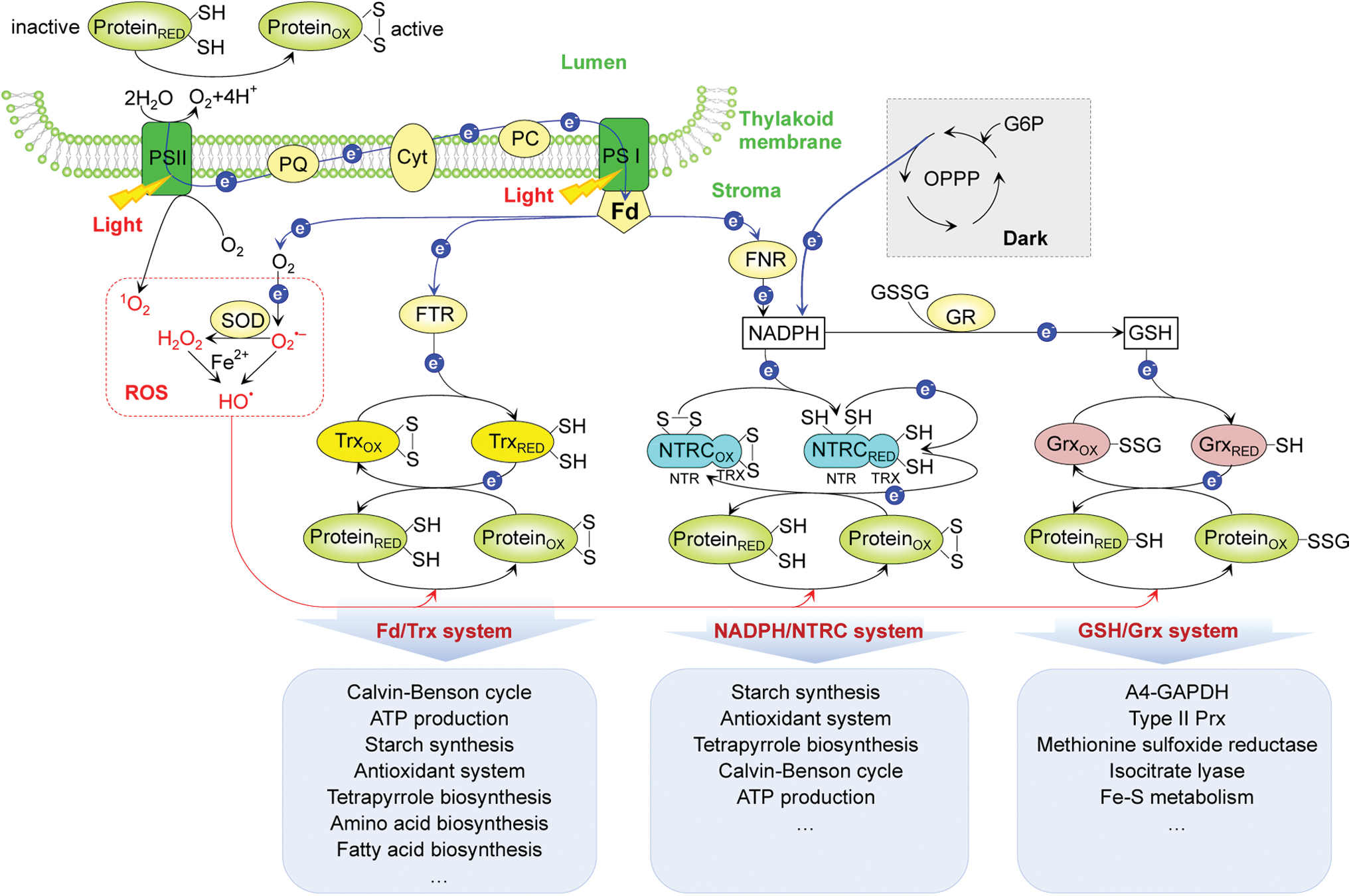
The redox states of proteins in the chloroplast stroma are mainly regulated by three thiol redox systems (Fig. 2). (i) Ferredoxin/thioredoxin (Fd/Trx) system: Fd is reduced by the photosynthetic electron transfer chain at PSI, and then the reduced Fd transfers electrons to ferredoxin/thioredoxin reductase (FTR), which subsequently reduces disulfide bonds present in the active sites of oxidized Trxs (f, m, x, y, and z) (147). Reduced Trxs are able to reduce regulatory disulfides in their target enzymes via dithiol/disulfide exchange. The Fd/Trx system is associated with diverse metabolic pathways, such as the Calvin/Benson cycle (CBC) (78, 146), ATP production (172), starch synthesis (15, 89, 104, 156), the antioxidant system (38), tetrapyrrole biosynthesis (136), amino acid biosynthesis (14, 47, 49, 59, 192), and fatty acid biosynthesis (144) (Fig. 2).
(ii) NADPH/NADPH thioredoxin reductase C (NTRC) system: the bifunctional enzyme NTRC contains both an NTR module and a Trx module. The NTR module contains a flavin adenine dinucleotide (FAD) cofactor and a redox active disulfide (127, 148). The reducing equivalents from NADPH generated either via ferredoxin-NADP+ reductase (FNR) in the light or the oxidative pentose phosphate pathway in the dark are transferred via FAD to an internal disulfide bridge in the NTR module. Following electron transfer, the reduced Cys of the NTR module serves to reduce the Trx module, which in turn reduces target proteins (31). The NTRC system has been implicated in the redox control of starch synthesis (83, 104), antioxidant metabolism (30), and tetrapyrrole biosynthesis (136) (Fig. 2).
Recently, Nikkanen et al. (121) proposed that the NTRC and Fd/Trx systems cross talk and play overlapping roles in the activation of fructose-1,6-bisphosphatase (FBPase), phosphoribulokinase (PRK), and ATP synthase. However, Yoshida and Hisabori (181) observed distinct target selectivity of the NTRC and Trx systems. For example, NTRC is inefficient in reducing FBPase, sedoheptulose-1,7-bisphosphatase (SBPase), and NADP-dependent malate dehydrogenase (NADP-MDH). These proteins are well-known Trx target proteins. The discrepancy between these two studies (121, 181) deserves attention.
(iii) Reduced glutathione/glutaredoxin (GSH/Grx) system: oxidized glutathione (GSSG) is reduced to GSH by glutathione reductase with NADPH (103). The generated GSH can then reduce Grxs (e.g., GrxC5, GrxS12, GrxS14, and GrxS16). The reduced Grxs catalyze the reduction of glutathionylated proteins, such as glyceraldehyde-3-phosphate dehydrogenase (GAPDH), type II Prx, methionine sulfoxide reductase, isocitrate lyase, and Fe-S metabolic proteins (Fig. 2) (42, 52, 66, 143, 185). These three thiol-reducing systems are crucial in regulating redox homeostasis in the stroma. However, our knowledge of the protein targets regulated by the three systems is far from complete.
More than 30 proteins in the thylakoid lumen were found to contain regulatory disulfide bridges (58, 69). Interestingly, the activity regulation of some redox-responsive lumen proteins is opposite to that of the stromal proteins. For example, violaxanthin de-epoxidase (VDE), a typical luminal enzyme converting violaxanthin to zeaxanthin, was inactivated when reduced in the dark and reactivated when oxidized in the light (150). The VDE and other luminal redox proteins (e.g., PsaN, PsbO, immunophilin FKBP13, and plastidic type I signal peptidase) were reported to be targets of Trx-like proteins (27, 58, 69, 70, 112). Thylakoid membrane-localized Trx-like proteins (e.g., high chlorophyll fluorescence 164 [HCF164] and lumen thiol oxidoreductase 1 [LTO1]) have soluble lumen-exposed Trx-like domains (70, 112). For example, PsaN (a PSI protein facing the lumen) was identified as a luminal target of HCF164, and PsbO (a luminal PSII subunit) is a target of LTO1 action, which produces a disulfide bond in PsbO during the biogenesis of PSII (70, 112). These data suggest that HCF164 and LTO1 function as important redox regulators in the thylakoid lumen. However, other luminal targets of HCF164 and LTO1 remain to be discovered.
Current proteomic technologies provide a high-throughput platform for investigating thiol redox proteins in plants, and have revealed dynamic regulations in chloroplasts under various metabolic and environmental conditions. Since RNS-mediated protein S-nitrosylation in plants has been recently reviewed (36, 48, 65), here we summarize the development of plant redox proteomic approaches for studying ROS-induced thiol PTMs, and discuss stress-responsive thiol redox signaling and metabolic pathways in chloroplasts to provide novel insights toward understanding stress-responsive redox regulation.
Quantitative Thiol Redox Proteomic Approaches
Most ROS-induced reversible Ox-PTMs of protein thiols (e.g., S-S, -SOH, and S-glutathionylation) have been identified using indirect thiol-labeling and comparative proteomic approaches (Fig. 3). First, the free thiols are irreversibly blocked by various alkylation reagents such as iodoacetamide (IAM) (7, 13, 192), N-ethylmaleimide (50, 93, 94, 117, 168), or 3-(N-maleimido-propionyl) biocytin (64). Then, the reversibly oxidized thiols are reduced by dithiothreitol (DTT) (7, 50, 93, 94, 117, 168) or tris-2-carboxyethyl phosphine (TCEP) (13, 64, 192). The newly generated free thiols are immediately labeled with various reagents for mass spectrometry (MS) analysis.
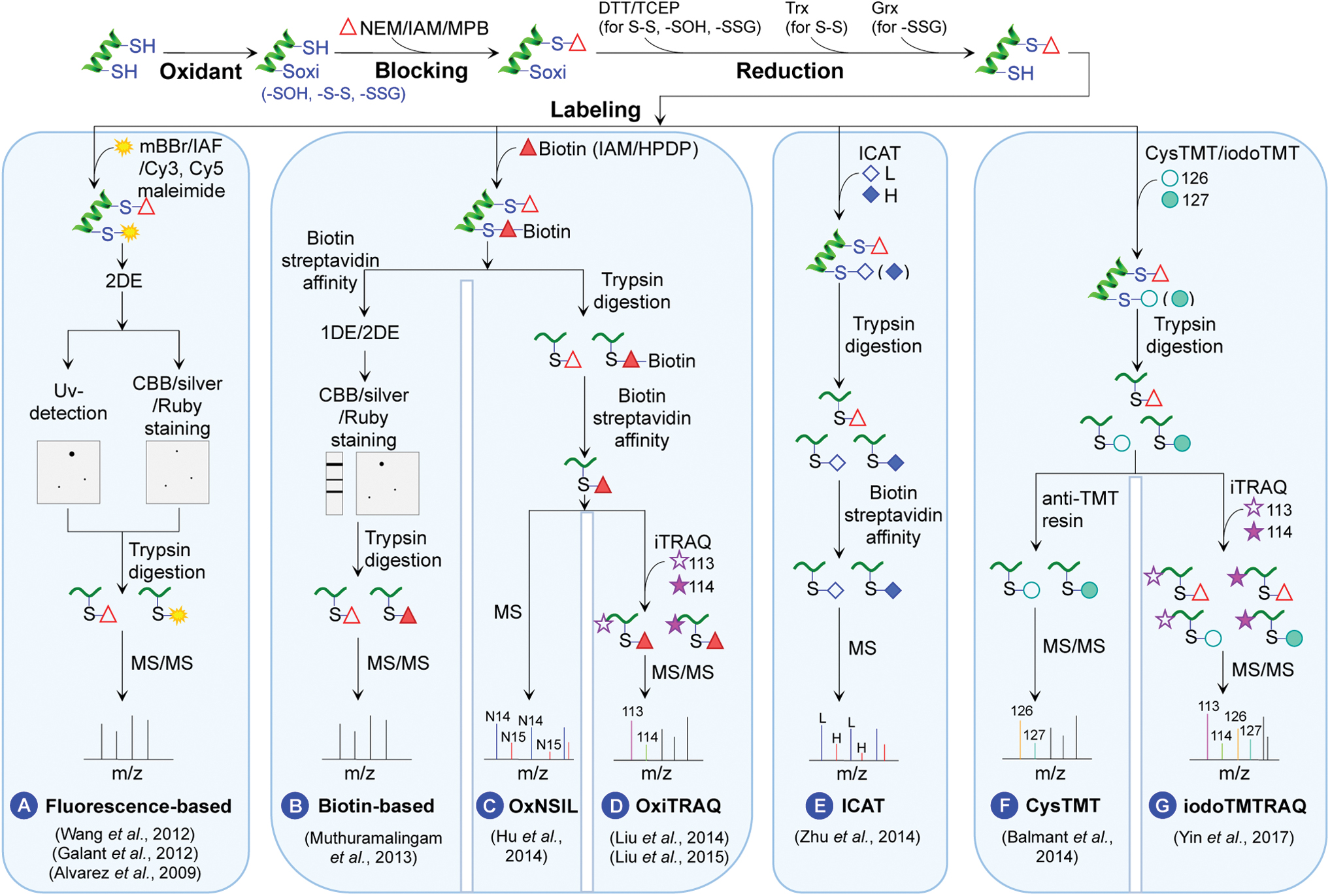
Several labeling methods have been developed to increase the sensitivity and efficiency of MS identification (Fig. 3). In fluorescence-based methods, the DTT/TCEP-reduced thiols are labeled with monobromobimane (7), iodoacetamidofluorescein (50, 168), or Cy3/Cy5 maleimide (192). The labeled proteins are separated by two-dimensional electrophoresis (2DE), visualized under ultraviolet, and identified by MS/MS (Fig. 3A). In the biotin-based method, the thiols are labeled by biotin-conjugated alkylating agents for affinity enrichment, for example, biotin-IAM or N-[6-(biotinamido) hexyl]-3′-(2′-pyridyldithio) propionamide. The enriched proteins are separated by one-dimensional electrophoresis (1DE) or 2DE followed by MS/MS identification (Fig. 3B).
Although the gel-based approach may identify proteins with changes in their redox state and quantify their overall redox levels, it turns out to be difficult to map specific redox-modified Cys residues (7, 50, 117, 168, 192). To overcome this deficiency, a 15N stable isotope metabolic labeling (OxNSIL)-based thiol redox proteomic method was developed to identify and quantify the reduced and reversibly oxidized Cys thiols by integrating biotin-tagged alkylating reagents with nitrogen isotope-based metabolic labeling (64). In OxNSIL, the proteins from different samples are labeled with either N14 or N15 during seedling growth. Comparison of light versus heavy N incorporation into the Cys-containing peptides allows for quantification of Cys redox changes (Fig. 3C) (64).
Another method OxiTRAQ was developed to label the Cys-containing peptides enriched via biotin tags with isobaric tag for relative and absolute quantification (iTRAQ) (Fig. 3D) (93, 94). The iTRAQ reagents contain eight reporter ions of 113, 114, 115, 116, 117, 118, 119, and 121 Da, allowing multiplexing eight samples in a single experiment. The peak area ratios of the iTRAQ reporter ions reflect the relative abundances of the Cys peptide redox states (Fig. 3D). However, the Cys redox quantification depends on successful enrichment of Cys peptides because the iTRAQ reagents label the peptide N-termini not the Cys residues.
To directly measure Cys redox states, several Cys labeling proteomic methods were developed, such as isotope-coded affinity tag (ICAT) (192), Cys tandem mass tag (cysTMT) (13), and iodoacetyl tandem mass tag (iodoTMT) (126, 179). In the ICAT method, the Cys thiols are labeled and enriched directly by the biotin-containing and thiol reactive ICAT reagents (heavy and light tags). Cys redox quantification can be performed by determining the ratios of heavy/light isotope-labeled peptide peaks in full scan spectra (MS1). Identification of the peptides and Cys sites is based on the product ion spectra (MS2) (Fig. 3E) (192). The biotinylated ICAT reagents allow enrichment of low abundant Cys peptides/proteins through avidin affinity chromatography. However, the duplex ICAT reagents limit its throughput (192).
Recently, six-plex cysTMT and iodoTMT reagents have shown multiplexing utility, and the use of anti-TMT antibody resin can enhance the depth of thiol redox proteomics (Fig. 3F) (13, 126, 179). The advantage of the iodoTMT reagents lies in the irreversible and efficient labeling of Cys residues, compared with the reversible labeling with the cysTMT reagents.
When studying Cys redox modifications, it is important to consider changes in total protein level. Recently, we developed a multiplex integrative method iodoTMTRAQ, which combines iodoTMT and iTRAQ quantification in a single experiment. It allows for the determination of bona fide redox-modified Cys in proteins (quantified by iodoTMT) after protein amount correction (quantified by iTRAQ) (Fig. 3G) (126, 179).
These indirect thiol-labeling methods overcome false positives arising from thiol/disulfide exchanges occurring during the protein extraction process. However, the reducing agents (e.g., DTT and TCEP) reduce all the reversible Ox-PTMs, and different types of Ox-PTMs cannot be distinguished. To overcome this limitation, specific reducing agents can be used. For example, arsenite can be used to reduce sulfenylated proteins, and Trx and Grx for the reduction of disulfide bonds and S-glutathionylated proteins, respectively (88, 163, 178). Moreover, ascorbate (AsA)/Cu+, Trx, and Grx can be sequentially used to reduce S-nitrosylation, disulfide bond, and S-glutathionylation, respectively. Each reduction reaction can be followed by differential thiol-based tag labeling, which will enable quantification of the specific reversible Ox-PTMs in a single experiment.
It should be noted that indirect proteomic methods cannot directly distinguish reduced and irreversible modifications (i.e., -SO2H and -SO3H). Aryl-nitroso probes for -SO2H and poly-arginine-coated nanodiamonds for selective enrichment of proteins containing -SO3H offer new avenues to study -SO2H and -SO3H PTMs (33, 95).
Recently, an activated thiol Sepharose-based proteomic approach was developed to enrich H2O2-sensitive thiol peptides for quantification of low-abundant redox proteins (174). In addition, a hybrid SILAC-iodoTMT expanded the multiplex redox proteomic capability through in vivo metabolic labeling and determination of cysteine redox state using reporter ion quantification (165). Moreover, oxSWATH was used to combine the label-free data-independent SWATH-MS acquisition method with a differential alkylation pipeline to discover and quantify relative thiol redox changes at the whole-proteome scale (9). To date, these newly developed thiol reactive probes and redox proteomic methods have not been utilized in plant redox proteomics.
Direct thiol-labeling methods use specific thiol probes to directly label Cys in different redox states and do not involve blocking the free Cys thiols during protein extraction. Three specific probes [dimedone (3), DYn-2 (4), and YAP1C probe (170)] have been applied to detect in vivo -SOH-modified proteins in H2O2-treated Arabidopsis thaliana (At) (Supplementary Fig. S1A–C). Also, 35S-radiolabeled Cys, biotinylated glutathione (GSH-biotin/GSSG-biotin) (42, 185), and biotinylated glutathione ethyl ester have been used for detecting plant S-glutathionylated proteins (66) (Supplementary Fig. S1D, E).
In addition, diagonal 2DE was used to analyze the Arabidopsis chloroplast thiol/disulfide redox proteome (156) based on differential migration of proteins in the gel under nonreducing and reducing conditions (Supplementary Fig. S1F). Plant Trx-interactome studies have been well studied and reviewed (28, 90, 91, 108, 191). To identify Trx target proteins, a trapping mutant approach was used (118). The normal Trx has a CxxC active motif, which can reduce the disulfide-bonded Cys residues in target proteins. When the C-terminal Cys of the CxxC motif is mutated, heterodisulfide bridges formed between the Trx and target proteins are stabilized, allowing enrichment of the Trx target proteins for subsequent MS/MS identification (Supplementary Fig. S1G).
Using this Trx-trapping mutant approach, more than 400 putative Trx target proteins have been identified in plants, among which ∼160 Trx target proteins are localized in the chloroplast stroma (14, 15, 113), thylakoid membranes (16), and lumen (58). These Trx target proteins are involved in central pathways for assimilation and storage of carbon, sulfur, and nitrogen, and protein translation and folding (14 –16, 58, 90, 113, 156).
Recently, some exciting in vivo probes have emerged to determine the redox states of living cells, such as a series of genetically encoded fluorescent sensors for H2O2 and redox states of NADH, NADPH, and thiols (186, 193), as well as various chemical probes for profiling transient protein S-sulfenic acid modification sites (17, 177). Improvement of these in vivo methods and further application to plants will contribute to our understanding of plant redox biology.
Stress-Responsive Thiol Redox Proteomics in Chloroplasts
Using the aforementioned thiol redox proteomic approaches, a total of 1285 redox-sensitive proteins were identified in A. thaliana (3, 4, 7, 42, 64, 66, 93, 94, 117, 168, 170, 179), tomato (Solanum lycopersicum) (13), soybean (Glycine max) (50), Brassica napus (192), and wheat (Triticum aestivum) (52), after exposure to various stresses, such as dehydration (52), NaHCO3 treatment (179), oxidants including H2O2 (3, 4, 64, 66, 93, 117, 168, 170), methylviologen (MV) (117), GSSG (42), and ozone (O3) (50), phytohormones including methyl jasmonate (MeJA) (7, 192), abscisic acid (ABA) (192), and salicylate (SA) (94), as well as the bacterial flagellin peptide flg22 (94) and the pathogen Pseudomonas Syringae pv tomato (Pst) (13) (Fig. 4A, B and Table 1).
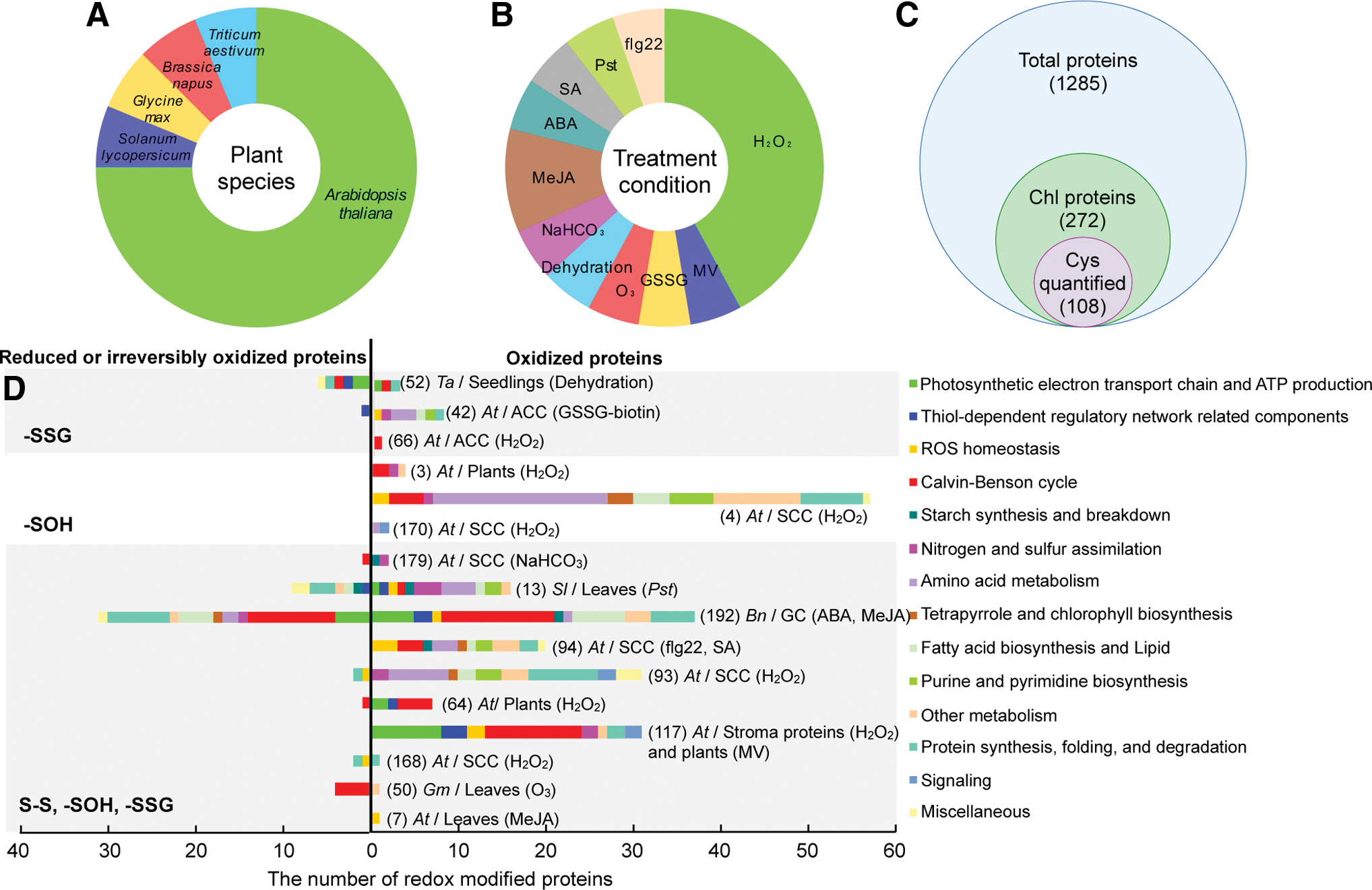
Summary of Publications on Plant Stress-Responsive Redox Proteomics
Numberical list of redox-responsive proteins by proteomic approaches.
Numberical list of redox-responsive proteins localized to chloroplast.
“Y” and “N” represent the approach is able or unable to quantify the ratio of reduced/oxidized at a specific cysteine site, respectively.
List of original research articles cited in this article. Please refer to the References section for details.
ABA, abscisic acid; BIAM, N-(biotinoyl)-N′-(iodoacetyl)-ethylenediamine; Bio-GEE, biotinylated glutathione ethyl ester; biotin-HPDP, N-[6-(biotinamido)hexyl]-3′-(2′-pyridyldithio) propionamide; Cys, cysteine; cysTMT, Cys tandem mass tag; GSH, reduced glutathione; GSSG, oxidized glutathione; GSSG-biotin, biotinylated glutathione; H2O2, hydrogen peroxide; IAF, 5-iodoacetamidofluorescein; ICAT, isotope-coded affinity tag; IDs, identified proteins; iTRAQ, isobaric tag for relative and absolute quantification; mBBr, monobromobimane; MeJA, methyl jasmonate; MV, methylviologen; OxNSIL, 15N stable isotope metabolic labeling; prf3, tomato Pst-susceptible genotype; Pst, Pseudomonas Syringae pv tomato; PtoR, tomato Pst-resistant genotype; SA, salicylate; -SOH, sulfenic acid; S-S, disulfide bond.
Among the 1285 redox-responsive proteins, 272 proteins were predicted to localize in chloroplasts (Supplementary Table S1), including 258 proteins solely in chloroplasts and 14 proteins targeted to both chloroplasts and other organelles. Among these proteins, 108 have the identified redox-responsive Cys sites (Fig. 4C). These proteins are mainly involved in thiol-dependent processes, such as ROS homeostasis, photosynthesis, metabolite biosynthesis, gene expression, protein turnover, and signal transduction in the chloroplasts (Fig. 4D and Supplementary Table S2).
Thiol Redox Regulation of ROS Scavenging Systems in Chloroplasts
Environmental stresses trigger redox changes in chloroplast antioxidant enzymes, such as stroma ascorbate peroxidase (sAPX), Prx, monodehydroascorbate reductase (MDHAR), dehydroascorbate reductase (DHAR), and glutathione-S-transferase (GST) (Fig. 5B).
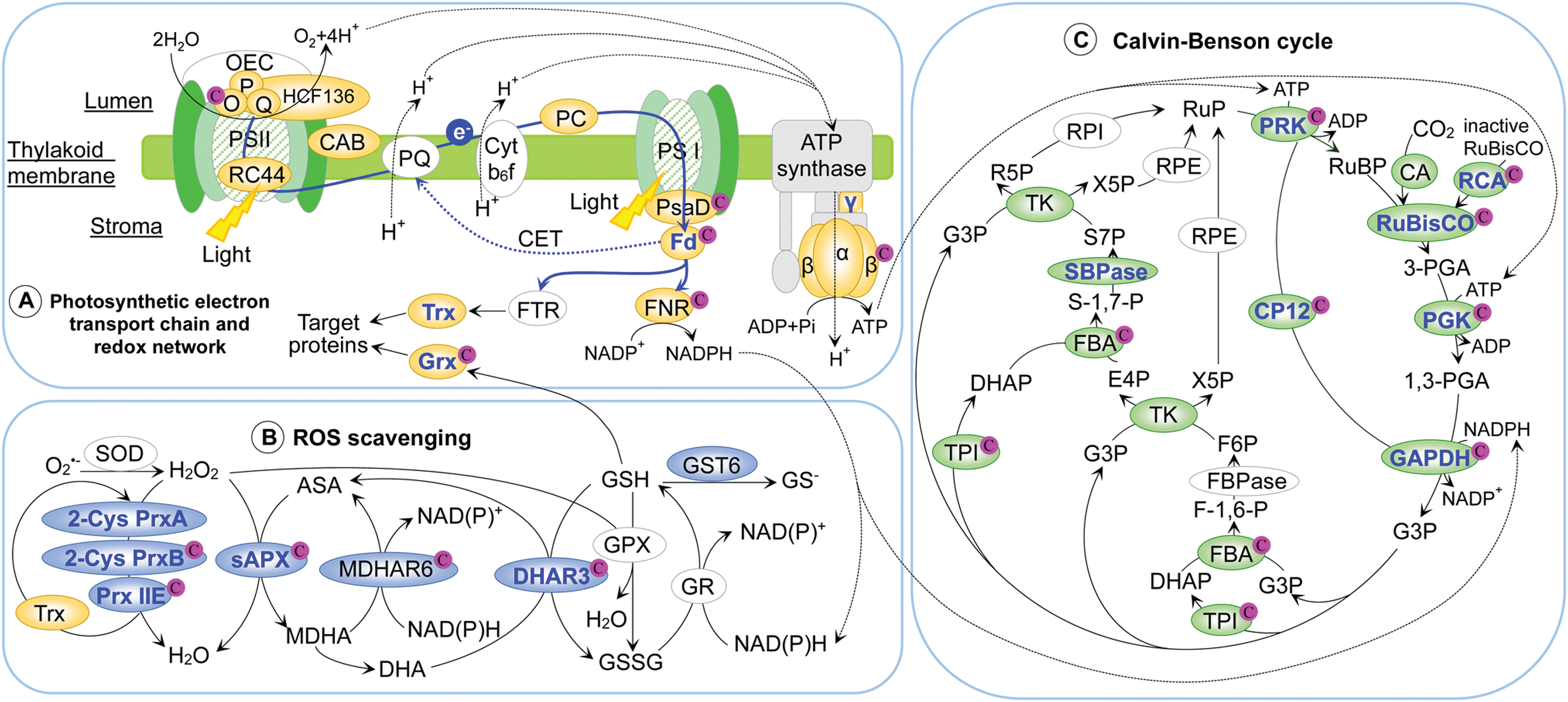
sAPX plays an important role in scavenging H2O2 generated by the photosystems (39). Redox proteomics has shown that sAPX was oxidized in ABA-treated B. napus (192), and sulfenylated in H2O2-treated Arabidopsis (4). Oxidation of Cys26 in a tobacco sAPX disrupted its binding of AsA, and led to enzymatic inactivation (72, 73). In response to moderate H2O2 and GSH, Cys26 and Cys126 in the tobacco sAPX were glutathionylated, likely to protect it against oxidative damage (72). Cys126 is homologous to a redox-sensitive Cys202 in B. napus sAPX (Supplementary Fig. S2A), indicating that this Cys glutathionylation may be a conserved mechanism for protecting the enzyme against irreversible oxidation.
Prx also contributes to chloroplast redox regulation. Arabidopsis chloroplasts contain two typical 2-Cys Prxs (i.e., PrxA and PrxB) and atypical monomeric Prx Q and Prx IIE (80). Redox proteomic studies have shown that the 2-Cys PrxA is oxidized upon treatment with H2O2, MV, or GSSG, while 2-Cys PrxB is oxidized by SA treatment (42, 94, 117). Prx reduces peroxide substrates and forms an intermolecular or intramolecular disulfide bridge with another resolving thiol, causing inactivation of Prx peroxidase activities (132). The inactive Prx can be restored after reacting with thiol-oxidoreductases (e.g., Trx, Grx, and cyclophilins), ensuring its cyclic utilization for peroxide reduction (38). Furthermore, transient accumulation of H2O2 may initiate ROS-related signaling to induce expression of Prxs, which may lead to reactivation of the Prx pathway (72).
NTRC can efficiently reduce 2-Cys Prxs, which are regulated by Trxs (128, 180). It has been proposed that the Fd/Trx and NTRC redox systems are linked through the redox balance of 2-Cys Prxs (128, 180). Thus, the thiol-based 2-Cys Prx was viewed as a redox switch crucial for chloroplast functions, such as GSH (32) (see the Amino Acid Biosynthesis in Chloroplasts section) and chlorophyll biosynthesis (137) (see the Redox-Modulated Tetrapyrrole and Chlorophyll Biosynthesis section).
Using the biotin switch method, an AtPrxIIE was found to be S-nitrosylated in NO-flushed leaves (92) and during the hypersensitive response (141). Moreover, upon in vitro S-nitrosoglutathione and diethylamine-NONOate treatments, the H2O2-reducing activity of AtPrxIIE was inhibited because of its Cys121 nitrosylation (142). Interestingly, this Cys121 was also oxidized after SA treatment (94). These results suggest that Cys121 is an important regulatory site of AtPrxIIE (94, 142).
DHAR and MDHAR function to regenerate AsA, critical for redox homeostasis in chloroplasts. A chloroplastic AtDHAR3 forms an intramolecular disulfide bond between Cys25 and Cys28 (41). Redox proteomics has revealed that Cys25 and Cys28 showed decreased oxidation in chloroplasts under H2O2 stress (41, 93). In contrast, a plastid-localized AtMDHAR6 was oxidized in response to H2O2 (3), and Cys69 of a tomato MDHAR was also oxidized in response to Pst infection (13). The activity of the chloroplastic MDHAR is regulated by Trx, but its thiol-mediated mechanism is not clear (156).
GST and glyoxylase (Glo) are detoxifiers of electrophilic compounds and methylglyoxal, respectively. Redox proteomics showed that a chloroplastic AtGST6 was oxidized in response to MeJA (7), and an Arabidopsis chloroplastic Glo I was oxidized in response to flg22 (94). A recent study found that a reversible intramolecular disulfide between Cys65-Cys110 in GST tau 23 (a Grx substrate) plays an important role in protecting Arabidopsis from oxidative stress (159). Although Glo I activity is known to be redox modified in human cells (22), its redox regulation in plants remains to be investigated.
It is noteworthy that these redox-responsive ROS scavenging enzymes, sAPX (100), Prx [including 2-Cys Prx (14, 81, 99, 113, 135, 143, 156), Prx IIE (99, 100, 143), and Prx Q (99, 135, 143)], DHAR (99, 100), MDHAR (156), and GST (99, 100) were identified as Trx targets in redox proteomic studies. Trxs modulate the redox status and activities of various antioxidant enzymes for directly scavenging ROS, contributing to ROS homeostasis and oxidative stress-linked signaling pathways (166). Further experiments are required to determine how Trxs modulate the redox states of these enzymes and their specific functions in chloroplasts under different biotic and abiotic stresses.
Thiol Redox Regulation of Photosynthesis and Starch Metabolism
Redox modulation of photosynthetic electron transport and ATP production
Redox proteomics has revealed redox proteins in photosystems (PSII and PSI), soluble electron carriers, and ATP synthase in response to stresses (Fig. 5A). Two subunits of Trx-regulated PSII oxygen-evolving complex (OEC; PsbO and PsbP) were oxidized in Arabidopsis and B. napus under stress conditions (e.g., H2O2, MV, or MeJA) (16, 58, 64, 99, 117, 192). PsbO contains two conserved Cys residues that form a disulfide bridge (e.g., Cys28 and Cys51 in spinach [Spinacia oleracea], Cys29 and Cys52 in Arabidopsis PsbO1) (116, 119), which is critical for PSII assembly and activity (19, 173). PsbO degradation was induced after reduction of the disulfide bond (140). Therefore, the redox sensitivity of the Cys residues and the stoichiometry of oxidized versus reduced PsbO determine the functionality of PsbO (16, 58, 64, 99, 117, 192). This represents an interesting redox regulatory mechanism underlying PSII biogenesis and degradation.
In addition, a PSII 44 kDa reaction center protein and HCF136 protein, involved in PSII reaction center assembly, were also redox modified in A. thaliana and B. napus in response to H2O2, ABA, or MeJA treatments (117, 192). Clearly, redox modulation affects the OEC activity and PSII functions under environmental stresses (85). Proteomics revealed that chlorophyll a/b binding protein (52, 192), plastocyanin (117), and PsaD (64) are oxidized by treatments of MeJA, MV, or H2O2. The stress-induced oxidization of these proteins may affect light harvesting and electron transport activity.
In chloroplasts, FNR transfers electrons from Fd to NADP+ to produce NADPH for carbon fixation, nitrogen metabolism, and biosynthesis of lipids and chlorophylls (167). FNR was also oxidized in response to H2O2, MV, or MeJA (117, 192), and it has two Cys residues essential for its activity (6). The plant-type [2Fe-2S] Fd has four cysteinyl ligands coordinating the 2Fe-2S cluster (68). Two of them in Arabidopsis, Cys44 and Cys47, were oxidized by H2O2 (64), but reduced or irreversibly oxidized in leaves of a tomato Pst-susceptible genotype after Pst infection (Supplementary Fig. S2B) (13). In the absence of functional studies, it is not clear how the Fd redox changes affect the assembly of the [2Fe-2S] cluster, PSI electron transport, and plant stress responses.
The activity of chloroplast ATP synthase is mainly regulated by the redox state of its central axis γ-subunit (63). Trx-mediated reduction of a disulfide bond between Cys199 and Cys205 in the γ-subunit induces a conformational change to activate the ATP synthase (172). Interestingly, the γ-subunit is reduced in B. napus guard cells in response to ABA, providing energy currency for stomatal closure (192). Furthermore, all of the subunits of the rotor (α-, β-, γ-, δ-, and ɛ-subunits) in the ATP synthase are potential Trx targets based on independent Trx interactomic studies in Arabidopsis (156), spinach (15, 16), potato (135), Chlamydomonas (81), and the cyanobacterium (Synechocystis sp. PCC 6803) (84, 102, 188). Among the rotor subunits, α-, β-, and γ-subunits were also redox modified in dehydration-treated wheat, Pst-treated tomato, H2O2-treated Arabidopsis, and ABA-treated B. napus (13, 52, 117, 192). However, the redox modification sites and their regulatory roles in these subunits are not known (63).
It should be noted that NTRC may also be involved in redox regulation of the ATP synthase (29, 121). Importantly, when there is no electrochemical proton gradient across the membrane for ATP synthesis, the ATP synthase will switch from ATP synthesis to ATP hydrolysis. Therefore, redox regulation of the chloroplast ATP synthase complex is crucial for prevention of this wasteful reverse reaction, which ensures efficient ATP synthesis and avoids futile ATP hydrolysis (63).
Redox-modified enzymes in the CBC
Thiol-mediated light/dark redox modulation of CBC enzymes has been well documented (25). Light-dependent activation of PRK, GAPDH, FBPase, and SBPase is regulated by the Fd/Trx system (78, 146). The other CBC enzymes are also targets of Trx, suggesting that the Trx-mediated redox modulation is very important for CBC (56, 78, 90). Mounting evidence indicates that besides disulfide bond formation, glutathionylation, sulfenylation, and nitrosylation also play important roles in regulating the CBC enzyme activities. For example, all the CBC enzymes of Chlamydomonas (78), and fructose-1,6-bisphosphate aldolase (FBA) (66) and GAPDH in Arabidopsis (187) can be glutathionylated. In addition, all the CBC enzymes in Chlamydomonas and other plants were found to be nitrosylated (1, 2, 52, 71, 87, 92, 123, 134, 141, 157).
Redox proteomic studies have revealed that most of the CBC enzymes, as well as CBC-related Calvin cycle protein (CP12) and NADP/NAD-MDH, were redox modified under various stresses (Fig. 5C) (3, 4, 13, 50, 52, 64, 94, 117, 192). Among them, thiol-based activation of ribulose bisphosphate carboxylase (RubisCO), RubisCO activase (RCA), phosphoglycerate kinase (PGK), GAPDH, CP12, PRK, SBPase, and NADP-MDH via Fd, FTR, and Trx has been documented at the biochemical and/or structural levels. Here are some detailed studies.
The RubisCO activity is well known to be modulated by the redox environment (110). For example, most of the Cys residues in RubisCO large subunits (LSU; Cys84, Cys172, Cys192, Cys247, Cys284, Cys427, and Cys459) (3, 52, 64, 117, 192) and small subunits (SSU; Cys96, Cys132, Cys145, and Cys167) (64, 94, 117, 192) are oxidized by different stresses. Specifically, Cys459 in LSU is nitrosylated, and Cys172, Cys247, and Cys427 undergo both nitrosylation and glutathionylation (71, 87, 92, 185).
To determine the functional significance of the Cys residues, site-directed mutagenesis of conserved Cys in LSU of Chlamydomonas revealed that Cys84 plays a predominant structural rather than regulatory role (51). In contrast, oxidation of Cys449 and Cys459 leads to RubisCO inactivation, and reduction restores its activity, supporting regulatory roles of these two Cys residues (110). Interestingly, Cys247 and Cys284 were found to be oxidized in proteomic studies (192), but their redox changes appear to be unrelated to RubisCO activity under different redox environments (51), highlighting the importance of functional validation through mutagenesis and other means. Nonetheless, many Cys residues in RubisCO are redox responsive, but how certain Cys redox states and modifications affect RubisCO conformation and activity requires further experimental testing.
Light activation of RubisCO requires RCA (189), which is also oxidized in response to H2O2 (64) and MeJA (192). The H2O2-oxidized Cys175 in Arabidopsis RCA was found to be nitrosylated (64, 78), and Cys392 and Cys411 can form a disulfide bond, which is reduced by Trx f in the light (190). The oxidized RCA has a decreased affinity for ATP, while reduction by Trx f alleviates this inhibition (131). After Trx f activation, RCA dissociates inhibitory sugar phosphates from the RubisCO active site, thereby restoring its activity (189). This is a good example showing a mechanistic connection between Cys modifications and regulation of RubisCO activity.
Proteomic results have also shown redox regulation of chloroplastic PGK under various stresses (3, 4, 13, 50, 192). PGKs from diatom (Phaeodactylum tricornutum) (23), Synechocystis (161), and Chlamydomonas (111) have different Cys sites, and two of them form a regulatory disulfide bond. These disulfide bonds are at different locations [e.g., Cys58 and Cys95 in P. tricornutum (23), Cys314 and Cys340 in Synechocystis (161), and Cys227 and Cys361 in Chlamydomonas (111)]. The PGK activity is inhibited upon disulfide formation and restored by Trx (23, 111, 161). Although PGK functions are conserved among different photosynthetic species, the diversity in redox regulation is intriguing (111).
Photosynthetic GAPDH is encoded by two genes (gapA and gapB) in plants. GAPA and GAPB proteins are both redox regulated (4, 50, 117, 192). GAPA and GAPB are almost identical, except that GAPB contains a pair of light/dark- and Trx-regulated Cys residues in its C-terminal extension (CTE) of ∼30 amino acids (154). The activity of AB-GAPDH primarily depends on the redox state of the two Cys residues in the CTE of GAPB (e.g., Cys349 and Cys358 in spinach), which are specifically regulated by Trx f (101, 153). GAPA monomers form stable homotetramers A4-GAPDH. In the absence of CTE, A4-GAPDH is not directly regulated by Trx per se, but affected by redox through its interaction with Trx-regulated CP12 and PRK (160).
CP12 has a C-terminus homologous to the CTE of GAPB. Upon oxidation, CP12 assists the formation of a GAPDH/CP12/PRK ternary complex, which strongly decreases all of the enzyme activities (155). When CP12 is reduced by Trx, it dissociates from its binding sites on GAPDH and PRK, and the enzyme activities can be rapidly recovered (53, 101). In Arabidopsis, Cys68 and Cys77 in AtCP12-1 are oxidized after SA treatment (94), possibly promoting GAPDH/CP12/PRK complex formation and decrease of the enzyme activities (155). In this context, CP12 itself acts as a specific redox regulator. In H2O2-treated Arabidopsis or ABA-treated B. napus plants, PRK is oxidized (64, 117, 192), and Cys295 in Arabidopsis was found to be sensitive to oxidation (64). The stress-induced redox perturbation of CP12 and PRK regulates the dynamics of ternary complex formation, and thereby modulates GAPDH activity (94).
Other CBC enzymes, including SBPase (117, 192), carbonic anhydrase (52, 117), triose phosphate isomerase (4, 64, 192), FBA (50, 66, 94, 117, 192), and transketolase (117, 192), are redox modified in response to various stresses.
In addition, NADP-MDH uses NADPH to produce malate that is transported out of the chloroplast, allowing for adjusting the NADP to NADPH ratio inside the chloroplasts under highly reducing conditions (145). This step needs to be strictly controlled because NADPH supply is critical for CBC. In chloroplasts, NADP-MDH is activated by reduced Trx in the light, but inactivated in the dark (12). An Arabidopsis NADP-MDH was sulfenylated in response to H2O2 (Fig. 6A) (3, 4). Interestingly, an NAD-dependent malate dehydrogenase (NAD-MDH) was also found to be oxidized in B. napus in response to ABA and MeJA (192). However, the NAD-MDH may not be redox modulated as DTT did not significantly affect its activity (18). Further experiments are needed to resolve the redox regulation of NAD-MDH.

Taken together, all the CBC enzymes and CBC-associated CP12 and NADP-MDH undergo multiple redox PTMs. Among them, several CBC enzymes (e.g., RubisCO, RCA, PGK, GAPB, FBPase, SBPase, and PRK) and CP12 are inactivated by oxidation at night and activated by Trx reduction in the light. Such light- and dark-dependent redox regulation helps to avoid a futile cycle between starch synthesis and starch utilization (86). Therefore, redox regulation of the CBC and associated enzymes constitutes a fine-tuning mechanism to modulate photosynthetic pathways responsible for carbon assimilation, and thus for plants to adapt to varying environmental conditions, especially during biotic and abiotic stresses.
Starch metabolism is redox regulated to cope with stresses
Starch biosynthesis and degradation in chloroplasts are required for energy supply and photosynthate cycling (Fig. 6B). During starch biosynthesis, ADP glucose pyrophosphorylase (AGPase) catalyzes the rate-limiting step, converting glucose 1-phosphate to ADP-glucose, a building block for starch synthesis (54). Proteomic studies have identified AGPase as a target of Trx (15, 89, 104, 156), Grx (143) and NTRC (83, 104), and it was oxidized in B. napus guard cells after MeJA treatment (192). The activity of AGPase is redox modulated through a disulfide bridge between two SSU of the AGPase heterotetrameric enzyme complex (60). The redox regulation was found to be crucial for starch synthesis in potato tubers (158) and in the leaves of potato, pea, and Arabidopsis (60). Moreover, an Arabidopsis mutant of the AGPase with a serine substituting Cys81 cannot form the disulfide bridge and is permanently activated, leading to more starch in the leaves (57).
In addition, the redox state of a granule-bound starch synthase (GBSS) was found to be modified in response to Pst infection (13). An OsGBSS forms a disulfide bond between Cys337 and Cys529, which plays an important role in facilitating starch biosynthesis (107).
Since transitory starch is a major product of leaf photosynthesis through the CBC, which is known to be strictly redox regulated, redox regulation of starch biosynthesis is obviously important to ensure coordination between photosynthesis and starch biosynthesis, which may link starch metabolism to the light-dependent redox states of CBC enzymes (151).
Starch breakdown is induced by glucan phosphorylation, which is catalyzed by α-glucan water dikinase (GWD) and phosphoglucan water dikinase (PWD) (139). The activity of GWD and its binding capacity for starch granules are modulated by Trx-mediated reduction of an intramolecular disulfide bond between two Cys residues in a conserved regulatory sequence (C1004FATC1008) (106). PWD shares sequence similarity with GWD, and both of them are subjected to sulfenylation upon H2O2 treatment (35). Moreover, PWD is oxidized in Arabidopsis seedlings treated with SA (94), which is known to cause cellular redox changes (61, 115). Oxidation of GWD and PWD under stresses is expected to inhibit carbon and energy supply from starch degradation for plant growth and development. It thus plays a regulatory role in balancing plant growth and defense responses. However, experimental evidence linking redox modifications of the starch degradation enzymes to the aforementioned regulatory function is lacking.
Redox Regulation of the Biosynthesis of Diverse Metabolites in Chloroplasts
During plant development and stress response, chloroplasts are important compartments for synthesizing a variety of metabolites, including amino acids, specific lipids and hormones, chlorophylls, purines, and pyrimidines, as well as various secondary metabolites (Fig. 6C–P). Here we discuss redox regulation of the biosynthesis of these metabolites.
Amino acid biosynthesis in chloroplasts
In chloroplasts, ammonia is assimilated in the glutamine synthetase (GS)/glutamate synthetase (GOGAT) cycle (Fig. 6C). The chloroplastic GS2 was found to be redox modified in response to H2O2, Pst, or ABA (3, 13, 117, 192), and Fd-dependent GOGAT was oxidized in response to H2O2 (93, 117, 143). GS2 is a Trx target (81, 99, 100, 113, 143, 175) and is activated by Trx m (49). Also, Fd-dependent GOGAT is a target of Grx C4 in poplar (143), and chloroplastic GOGATs in spinach and soybean are activated by DTT and Trx, but not GSH, indicating that gluthathionylation may not play a role (86). Redox activation of GS2 and GOGAT may help to alleviate the toxicity of ammonium ion built-up (by converting to glutamine and glutamate) during light-dependent processes of nitrate assimilation and photorespiration (86).
In chloroplasts, Cys synthesis is catalyzed by a hetero-oligomeric cysteine synthase complex containing O-acetylserine (thiol) lyase (OASB) and serine O-acetyl transferase 1 (SAT1) (Fig. 6D). OASB is a potential Trx/Grx target (14, 143), which is oxidized in Pst-treated tomato (13) and sulfenylated in H2O2-treated Arabidopsis (4). Although direct redox regulation of SAT1 has not been described, a Trx-target cyclophilin 20-3 (CYP20-3) can bind SAT1 and trigger formation of the cysteine synthase complex, leading to accumulation of Cys and GSH in response to wounding (44, 125).
The rate-limiting step of GSH synthesis is catalyzed by glutamate-cysteine ligase (GCL) (Fig. 6E). GCL can be activated by forming intramolecular disulfide bridges, for example, Cys178-Cys398 and Cys341-Cys356 in Brassica juncea (55), and Cys186-Cys406 and Cys349-Cys364 in Arabidopsis (62). In addition, OxiTRAQ proteomics revealed that Cys349 in AtGCL was oxidized in response to H2O2 (93). Interestingly, GCL interacts with 2-Cys Prx (32), but the role of the interaction is not known. The thiol redox regulation of these enzymes involved in sulfur assimilation provides a post-translational mechanism for modulating cellular redox homeostasis afforded by thiol metabolites and GSH in plants.
Aromatic amino acids (tryptophan, phenylalanine, and tyrosine) are produced from chorismate in the shikimate pathway in chloroplasts (Fig. 6I). Redox proteomics revealed that three enzymes in the shikimate pathway (i.e., 3-deoxy-D-arabinoheptulosonate 7-phosphate synthase [DAHPS], 5-enolpyruvylshikimate-3-phosphate synthase, and chorismate synthase) and two enzymes in the postchorismate pathway (i.e., arogenate dehydratase 2 and anthranilate synthase 2) were oxidized in response to various stresses (4, 13, 93, 94).
DAHPS catalyzes the first committed step in the shikimate pathway, converting phosphoenolpyruvate and D-erythrose 4-phosphate to chorismate (98). Its activity is inhibited by formation of a disulfide bond (e.g., Cys139-Cys494 in Arabidopsis and Cys87-Cys440 in Mycobacterium tuberculosis), which prevents metal ligand binding to the Cys sites for activation (Supplementary Fig. S2C) (20). An earlier study showed that AtDAHPS1 could be activated by the chloroplastic Fd/Trx system (47). Since the shikimate pathway is one of the major routes of carbon flow in plants, redox regulation of its enzymes (especially Fd/Trx activation of the DAHPS) links PSI with carbon flow into the shikimate pathway. Thus, the biosynthesis of the aromatic amino acids and aromatic secondary metabolites may be redox regulated by the Fd/Trx system.
Branched-chain amino acids (valine, leucine, and isoleucine) synthesized in chloroplasts are precursors for secondary metabolite synthesis (Fig. 6G) (21). Several enzymes (e.g., acetolactate synthase, 2-isopropylmalate synthase 1, isopropylmalate isomerase, branched-chain-amino-acid aminotransferase 3, and 3-isopropylmalate dehydratase [IPMDH]) in the branched-chain amino acid biosynthesis are oxidized under various stress conditions (4, 13, 42, 93, 94, 192). IPMDH1 is involved in methionine chain-elongation of glucosinolate biosynthesis, and IPMDH2 and IPMDH3 are mainly responsible for leucine biosynthesis (59). AtIPMDH1 activity is regulated by Trx-mediated redox regulation at Cys232 and Cys390 (59, 192). In addition, IPMDH1 from S. lycopersicum-resistant (PtoR) genotype and B. napus guard cells, and IPMDH2 from Arabidopsis suspension cells are oxidized in response to Pst, MeJA, or H2O2 (4, 13, 192).
Furthermore, ten chloroplastic enzymes involved in the biosynthesis of other amino acids (e.g., lysine, threonine, histidine, arginine, aspartate, and asparagine) (Fig. 6F, H, and Supplementary Table S2) (4, 13, 42, 93, 94, 170, 192), and several methyltransferases (4, 94) and aminotransferases (4) implicated in amino acid metabolism are also redox responsive. Some of them are potential Trx targets through Trx-interactomic analyses, for example, DapF (81), threonine synthase (16, 81), argininosuccinate synthase (81), serine hydroxymethyltransferase 3 (15), and aspartate aminotransferase (15). These results imply that the redox environment regulates amino acid metabolism and secondary metabolism in chloroplasts. However, the types of Ox-PTMs and redox-regulated Cys sites in most of these enzymes remain to be investigated.
Redox-regulated fatty acid and lipid biosynthesis
Fatty acid biosynthesis in plastids is essential to provide acyl chains for the synthesis of various acyl lipids involved in membrane dynamics, signaling, carbon storage, and energy (Fig. 6K). Acetyl-CoA carboxylase (ACC) catalyzes the first committed step of de novo fatty acid synthesis in chloroplasts, and it is activated by reduced Trx, DTT, or light (144). ACC is a multienzyme complex consisting of a biotin carboxylase subunit (BCS), a biotin carboxyl carrier protein (BCCP) and carboxyltransferases with two nuclear-encoded α-subunits and two chloroplast-encoded β-subunits (144). BCS in Arabidopsis suspension cells (42) and BCCP in Chlamydomonas (105) were both subjected to S-glutathionylation. BCCP was also reduced or irreversibly oxidized in response to Pst and ABA, respectively (13, 192). Also, the carboxyltransferase was inactivated when Cys267 of the α-subunit and Cys442 of the β-subunit formed an intermolecular disulfide bridge (76). All these results indicate that redox modulation of ACC activity may be a major determinant of fatty acid synthesis and lipid metabolism in chloroplasts.
In addition, three enzymes in the fatty acid synthase complex (including a 3-ketoacyl-[acyl carrier protein] synthase I [KAS I], a KAS II, and an NADH enoyl-acyl carrier protein reductase) are redox modified in response to stresses (4, 13, 192). Furthermore, several desaturases are oxidized in response to H2O2 (4, 93). Redox regulation of fatty acid desaturases is critical for production of unsaturated fatty acids and adjustment of membrane fluidity in the course of stress acclimation (164). These data indicate that a redox cascade is likely to link light and fatty acid synthesis, resulting in coordination of fatty acid synthesis with photosynthesis (144).
Plastid lipid-associated proteins (aka fibrillin/CDSP34 proteins) important for the structural stabilization of thylakoid membrane were found to be Trx regulated and oxidized in B. napus guard cells in response to ABA and MeJA (Fig. 6L) (58, 192). The significance of redox-regulated thylakoid membrane stability in guard cell signaling and stomatal movement is not known (77). In addition, essential enzymes for jasmonic acid biosynthesis (allene oxide cyclase 4 and its close homologue early response to dehydration protein) were redox modified in B. napus guard cells (Fig. 6M) (192). Redox regulation of guard cell hormone biosynthesis and signaling is an exciting direction for future research.
Redox-modulated tetrapyrrole and chlorophyll biosynthesis
Tetrapyrrole biosynthesis is critical for producing a variety of important macromolecules, including chlorophyll, heme, and phytochromobilin, and it involves more than 25 enzymes (Fig. 6J) (136). Several of these, such as glutamate-1-semialdehyde-2,1-aminomutase (GSAT), aminolevulinic acid dehydratase (ALAD), uroporphyrinogen III decarboxylase, protogen oxidase I, and Mg chelatase (MgCh), were identified as potential Trx targets using Trx-affinity chromatography MS (14, 99). Five other enzymes, glutamyl-tRNA reductase (GluTR), magnesium-protoporphyrin IX monomethyl ester (MgPMME) cyclase, MgProto methyltransferase, light-dependent protochlorophyllide oxidoreductase, and subunit CHLI of MgCh, are redox modulated by NTRC (24, 136, 137). Interestingly, recent data support coordinated roles of Trx m and NTRC in the redox control of Mg-protoporphyrin IX methyltransferase stability and activity (34).
Redox proteomics revealed that GSAT, ALAD, MgCh, ferrochelatase 1 (FC1), and MgPMME cyclase are redox modified in response to various stresses (4, 93, 94, 192). Among these enzymes, GSAT has three highly conserved Cys residues for Trx-mediated reduction (26). Formation of disulfide bridges between interacting GluTR and GSAT may be important for stabilizing the multimeric complex GluTR2GSAT2 (136), and the disulfide bonds can be reduced by NTRC or Trx (14, 137). In addition, CHLI, one MgCh subunit, is activated by Trx m, Trx f, and NTRC (97, 130). Although FC activity was inhibited by thiol blocking reagents (152), the active Cys sites and Ox-PTMs in FC1 are unknown. The MgPMME cyclase activity was stimulated by NTRC and 2-Cys Prx (137).
Upon plant transition from dark to light, tetrapyrrole and chlorophyll biosynthesis regulated by light-dependent Trx and NTRC meets the chlorophyll demand for fast activation of photosynthesis. Under stress conditions, excess excitation energy of chlorophylls and accumulation of light-absorbing tetrapyrrole intermediates generate ROS, leading to redox perturbation in chloroplasts (11). Thus, redox regulation of the enzymes involved in tetrapyrrole and chlorophyll biosynthesis contributes to balancing the photodynamic tetrapyrrole intermediates and preventing the risk of oxidative damage. Although tetrapyrrole and chlorophyll production is redox modulated by Trx and NTRC, the extent of coordination between Trx and NTRC in tetrapyrrole metabolism under stress conditions is not known.
Redox-modulated biosynthesis of purine, pyrimidine, and thiamine
All ten steps of purine biosynthesis and the first three steps of pyrimidine biosynthesis take place in plastids (194). Five enzymes in purine biosynthesis and two enzymes for pyrimidine biosynthesis are oxidized in Arabidopsis and S. lycopersicum in response to various stresses (Fig. 6O and P) (4, 13, 42, 93, 94). However, little is known about their redox mechanisms. Thiamine (vitamin B1) is an essential cofactor with strong antioxidant potential that protects plants from oxidative damage (162). Two enzymes (thiamine requiring 1 and Thi1) involved in thiamine biosynthesis are redox modified in Arabidopsis, B. napus, and G. max in response to stresses (Fig. 6N) (4, 50, 192). Thi1 with conserved Cys residues is a potential target of chloroplastic Trx (14, 81). How Trx regulates Thi1 under normal plant growth and stress conditions is an interesting question.
Redox Regulation of Chloroplast Gene Expression and Protein Fate
Semiautonomous plastids/chloroplasts have their own genome with ∼120 genes (183). The expression of photosynthetic genes is partly redox regulated to modulate photosynthetic electron transport, plastoquinone pool, and ROS homeostasis (182). For example, in cyanobacterium, de novo synthesis of D1 protein was suppressed by H2O2 (122). ROS also decreased the level of PsbA messenger RNA (mRNA)-polysome complex (122). A catalase/peroxidase double knockout mutant with elevated ROS levels displayed a much lower rate of protein synthesis than wild type (122). Interestingly, this inhibition could be alleviated by reduced elongation factor G (EF-G) (75).
Redox proteomic results have corroborated the above findings. For instance, the eukaryotic translation initiation factor 3G1 was oxidized in response to H2O2 and MeJA treatments (93, 192), and several other elongation factors (e.g., EF-Tu and AtRABE1b) are also redox modified (4, 52, 93). In line with this, several translation elongation factors, including EF-Tu (14, 15, 81, 84, 89, 102, 126a, 143) and EF-G (14, 89, 126a), were identified as Trx targets through Trx-interactomic analysis.
Biochemical analysis of the Synechocystis translation system revealed that EF-G and EF-Tu were targets of ROS inactivation via formation of an intramolecular disulfide bond between Cys105 and Cys242 in EF-G (74), and oxidation of Cys82 in EF-Tu (184). Moreover, oxidation of EF-G (46, 75) and EF-Tu (67, 184) under strong light inhibited de novo synthesis of proteins (e.g., D1) required for PSII repair, resulting in photoinhibition. Therefore, the redox status of EF-G and EF-Tu might act as a regulator for modulating PSII repair, which would contribute to redox homeostasis by preventing excessive oxidative stress caused by photosynthetic electron transport under strong light (46, 67, 75, 184). However, the ROS-regulated oxidation of chloroplast elongation factors CpEF-G and CpEF-Tu in higher plants, similar to those in Synechocystis in terms of structure and function, is yet to be demonstrated (43).
Several chloroplastic ribosomal proteins (4, 52, 93, 192), a ribosome recycling factor (93), and translation-related proteins (93, 94, 192) were found to be redox-modified in chloroplasts to cope with stresses. Some of these proteins have been identified as potential Trx targets (14, 15).
Different CYPs (117, 168, 192), chaperones (e.g., CPN60α and CPN60β) (4, 42, 117, 192), and heat shock proteins (HSPs, e.g., cpHSP70-1, cpHSP70-2, HSP 93-V, and HSP101) (4, 93) involved in protein folding and processing are redox modified in chloroplasts in response to various stresses. Redox proteomics has revealed that the CYP20-3 is redox modified in response to H2O2 and MeJA (117, 168, 192). The peptidylprolyl cis-trans isomerase activity of CYP20-3 is fully inactivated in its oxidized form due to formation of a disulfide bridge (79). Its activity can be recovered by Trx m (114). The redox regulation of peptidylprolyl cis-trans isomerase activity may be vital to CYP20-3-mediated protein/protein interaction (79). In chloroplasts, Cys129 and Cys171 of CYP20-3 are required for the regeneration of 2-Cys PrxA and 2-Cys PrxB, which in turn regulate their target proteins (79). Thus, redox regulation plays an important role in maintaining normal protein conformation and function in chloroplasts under stress conditions (169).
In addition, several chloroplastic aminopeptidases and proteases (e.g., ClpC, ClpB, Zn oligopeptidase A, and FtsH) are potential Trx or Grx targets. Some of these proteins are reduced or irreversibly oxidized in Arabidopsis and B. napus in response to Pst, MeJA, or ABA (13, 192). As to many of the above proteins, their detailed redox regulatory mechanisms and functions still need to be investigated.
Cross Talk Between Chloroplast Protein Redox Modification and De/Phosphorylation
Reversible protein de/phosphorylation catalyzed by protein phosphatases/kinases is crucial for mediating various signal transduction events and enzymatic activities. The cross talk between the phosphoPTMs and Ox-PTMs of chloroplastic proteins is poorly understood. Redox proteomics has indicated that several kinases are redox modified in response to H2O2 (93, 117, 170).
The Arabidopsis STN7 and Chlamydomonas Stt7 are orthologous thylakoid light-harvesting antenna complex II (LHCII) kinases required for state transitions (37, 124). STN7/Stt7 contains a transmembrane region that separates its catalytic kinase domain on the stromal side from its N-terminus in the thylakoid lumen with two conserved Cys residues in both plants and green algae (124). Also, there are two conserved Cys residues located in the stroma kinase catalytic domain in land plants (e.g., Cys187 and Cys191 in Arabidopsis STN7), but they are not conserved in unicellular green algae (149). Arabidopsis STN7 was found to be oxidized in response to H2O2 (117).
STN7 was inhibited by reduction of a disulfide bridge possibly by Trx (10, 138). Recently, overexpression of a Trx m in tobacco chloroplasts was found to inactivate STN7 through reduction (8). The Trx-mediated STN7 inactivation could be explained by two working hypotheses: (i) Trx directly breaks the stromal disulfide bridge of STN7, interfering with ATP-binding and leading to its inactivation in land plants (133, 138); and (ii) the luminal disulfide bridge could be reduced through a trans-thylakoid redox pathway (via the Trx-like proteins HCF164 and LTO1 described in the Introduction section) for delivery of reductants from the stroma to the lumen (40, 70, 82, 112). This hypothesis is supported by a recent finding that a stromal region of the Cytb6f complex is responsible for the activation of the Stt7 kinase (45). However, extensive experimental support for this scenario is lacking.
In addition, NTRC is also involved in redistribution of excitation energy between PSII and PSI (120). However, the NTRC-mediated excitation energy redistribution is unlikely to alter the STN7 activity, because phosphorylation of the principal STN7 substrate LHCII protein in the light was the same in the Arabidopsis wild-type, NTRC overexpression lines, and ntrc mutants (120). Although LHCII phosphorylation has been shown to be thiol sensitive, no light-dependent changes in STN7 thiol redox state were observed (149). The enigma of the light and Trx regulation of STN7 may lie in the special thylakoid localization of STN7 and Trx-like proteins.
Chloroplast-localized nucleoside diphosphate kinase 2 (NDPK2) is a positive signal transducer of phytochrome-mediated plant light signaling. AtNDPK2 interacts with two oxidative stress-activated mitogen-activated protein kinases (MAPKs) (AtMPK3 and AtMPK6) to regulate the cellular redox state, which enhances tolerance to multiple stresses (e.g., cold, salt, and H2O2) (109). Redox proteomics revealed that NDPK2 is oxidized in Arabidopsis in response to MeJA (117) and H2O2 (93). In addition, Cys219 of the AtNDPK2 was identified as a redox-sensitive site (93). How MAPKs act in redox regulation and how redox affects autophosphorylation and substrate phosphorylation activities remain to be explored.
Conclusion and Perspectives
Thiol-based redox switches are of fundamental significance in regulating cellular molecular processes. Over the past decades, thiol-redox proteomic studies have provided a wealth of information and insights toward understanding redox-regulated gene expression and protein turnover, ROS homeostasis, photosynthetic electron transport and ATP production, assimilation of carbon and ammonia, and the biosynthesis of different metabolites (e.g., fatty acids, lipids, tetrapyrroles, purines, pyrimidines, and thiamine) in the chloroplasts to cope with different stress conditions.
Since most of these redox-responsive proteins have been identified in the reference plant Arabidopsis, translating the knowledge of redox regulatory networks to crop species will be an important and exciting task in the future. Also, the types of oxidants causing specific Cys Ox-PTMs are often unclear, and thus, major breakthroughs may be expected as to how different oxidants (e.g., ROS) are sensed, trigger redox PTMs, and how the antioxidant systems are activated.
To date, no redox proteomic approaches/methods are perfect. The gel-based and sulfenylation labeling-based redox methods are deficient in mapping the Ox-PTMs and quantifying the reduced Cys/oxidized Cys stoichiometry of chloroplast proteins. Although the gel-free liquid chromatographic-MS/MS-based redox proteomic methods can identify the Cys sites and quantify their redox states, they cannot discriminate between reduced and irreversibly oxidized Cys in chloroplasts. Therefore, development of specific redox proteomic approaches for sequential thiol-based reduction and differential labeling of reduced Cys and irreversibly oxidized Cys (e.g., -SO2H and -SO3H) in plants will enhance the depth and coverage of redox proteomes.
New in vivo accurate probes for Cys redox states and multiplex labeling together with enrichment strategies for different PTMs (e.g., phosphorylation, acylation, and ubiquitination) need to be developed. They will allow determination of the stoichiometry and dynamics of Cys Ox-PTMs and the cross talk between multiple PTMs in chloroplasts.
Currently, most target proteins of Trx, Grx, and NTRC identified by proteomics need to be validated using orthogonal approaches and methods. In addition, the cooperative redox modulation of the targets by the three redox systems in chloroplasts should be investigated in due course. Besides, the novel genetically encoded sensors for NADPH, AsA, Trx, Grx, and RMS for determining their subcellular localization and redox states will contribute to a comprehensive view of redox homeostasis in chloroplasts and other compartments.
Importantly, the functional significance of the identified redox Cys sites of chloroplast proteins needs to be evaluated using complementary approaches and means. For example, phenotypic and biochemical analyses of overexpression and loss-of-function mutants of specific redox proteins and/or Cys sites will facilitate understanding their respective functions in different biological processes. In addition, protein crystal structure will also contribute to elucidating the modes of action of Cys redox regulations in the chloroplasts.
In summary, integration of thiol-redox proteomic results with findings from other large-scale PTM “omics” (e.g., phosphoproteomics), as well as targeted functional characterization, will generate a comprehensive regulatory network underlying chloroplast functions and stress responses. In addition, putting the identified redox proteins and conserved/alternative Cys sites in the context of plastid/chloroplast development and evolution from cyanobacteria, diatoms, and algae to land plants will be an exciting direction for future studies.
Footnotes
Acknowledgments
The authors thank Prof. Jean-David Rochaix from the University of Geneva, as well as Dr. Joseph M. Collins and Hope Hersh from the University of Florida Genetics Institute, for critical reading and editing of the article.
Funding Information
The project was supported by grants from the Foundation of Shanghai Science and Technology Committee, China (No. 17391900600) to S.D., the Fund of Shanghai Engineering Research Center of Plant Germplasm Resources (No. 17DZ2252700), and the Fundamental Research Funds for the Central Universities (Nos. 2572017ET01 and 2572017EA05) to S.D. and Y.L.
Supplementary Material
Supplementary Figure S1
Supplementary Figure S2
Supplementary Table S1
Supplementary Table S2
Abbreviations Used
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.