
Abstract
Aims:
Polymyxin B (PMB) is known to require reactive oxygen species (ROS) for its bactericidal activity, but the mechanism of PMB resistance in various Pseudomonas aeruginosa strains has been poorly understood. This study examined the role of nitrate respiration (NR) of some P. aeruginosa strains in the PMB resistance.
Results:
We observed that the minimum inhibitory concentration (MIC) value of PMB against P. aeruginosa PA14 was eightfold reduced (from 2.0 to 0.25 μg/mL) by agitation, but not against P. aeruginosa PAO1 (from 2.0 to 1.0 μg/mL). Transcriptomic and phenotypic analyses using both strains and their NR mutants revealed that the higher NR in PAO1 than in PA14 accounted for the higher MIC value (i.e., PMB resistance) of PAO1, which was sufficient to compromise the antibacterial activity of PMB in Drosophila infections. We also confirmed the contribution of the NR to the PMB resistance is independent of the major catalase (KatA), suggesting that the NR might affect the ROS generation rather than the ROS disintegration. Furthermore, this PMB resistance was relatively common among clinical P. aeruginosa isolates and correlated with higher NR in those strains.
Innovation and Conclusion:
These results suggest P. aeruginosa strains could display intrinsic resistance to antibiotics in clinical settings and that NR is a crucial factor in the intrinsic antibiotic resistance, and also provide an insight into another key target for successful antibiotic treatment of P. aeruginosa infections. Antioxid. Redox Signal. 34, 442–451.
Introduction
P
PMB has in vitro bactericidal activity against almost all Gram-negative bacteria at relatively low concentrations, which involves its interaction with the lipid A of lipopolysaccharide (LPS) (17, 22, 32). PMB changes the packing order of LPS and increases the outer membrane (OM) permeability to various molecules inducing PMB itself (i.e., self-promoted uptake). Although the detailed mechanism of the OM permeability increase remains unveiled (11), it is obvious that the OM-disorganizing activity of PMB is independent of the bactericidal activity of PMB that is associated with the functions in the cytoplasmic membrane (CM) such as active transport and respiratory electron transfer (36, 38). PMB also interacts with the acidic phospholipids exposed on the CM (5) and, at high concentrations, induces ion-permeable pores in the OM and depolarizes the CM. But the bactericidal effect of PMB was evident even at lower concentrations and thus deemed independent of the CM depolarization (11).
Such complicated action of PMB on the Gram-negative bacteria is likely associated with the lower incidence of PMB-resistant bacteria (15). Nevertheless, there are PA clinical isolates that have acquired PMB resistance (3, 31, 37). The main reason for the resistance is decreased PMB binding due to the reduced anionicity of LPS, which involves modifications in the core region and/or lipid A (15). For example, the mutations in the two-component systems, PhoPQ or PmrAB result in the constitutive expression of the LPS modification (arn) operon encoding the proteins necessary for the aminoarabinosylation of the lipid A (30, 32, 37). This is the most common mechanism of the acquired PMB resistance observed in many Gram-negative bacteria. Besides the reduced LPS binding directed by LPS modification, other species-specific maneuvers of PMB resistance have been known; for example, the expression of multidrug efflux pumps or OM proteins such as OprH that can pump out PMB (42) and the presence of capsular polysaccharides that can bind to PMB by ionic interactions (7), both of which prevent PMB from reaching the bacterial target, resulting in the reduction of the effective concentration. Despite these advances in the understanding of the acquired resistance of PA to PMB, little has been known about the intrinsic resistance determinants to PMB in PA strains, considering that PA is quintessentially notorious for its intrinsic resistance to various antibacterials (33, 35).
Innovation
This is the first report to show that nitrate respiration promotes the intrinsic antibiotic resistance of the opportunistic human pathogen, Pseudomonas aeruginosa. These results also suggest that various P. aeruginosa strains could display differential nitrate respiration activity based on their own physiological adaptation in clinical settings and thus provide insight into another key factor to target for successful antibiotic treatment of P. aeruginosa infections.
In this study, we discovered the intrinsic PMB resistance of the most widely used PA strain, PAO1, which is not observed in another PA strain, PA14. The difference in intrinsic PMB resistance is directed by higher nitrate respiration (NR) even in PAO1 under aeration/agitation condition, which is sufficient to trounce the antibacterial efficacy of PMB in experimental infections. The higher NR activity in PAO1 that directs the higher expression of the major catalase, KatA, but the NR-independent contribution of KatA was suggested, in that NR could prevent the reactive oxygen species (ROS) generation by partially bypassing oxygen respiration, which might be observed at a considerable frequency in clinical PA isolates.
Results
PAO1, but not PA14, displays intrinsic PMB resistance
In our previous study, we observed that a widely used P. aeruginosa (PA) strain, PAO1, displays higher resistance to H2O2 and PMB than another PA strain, PA14 (19). Despite this observation, we have confirmed that both strains have the same minimum inhibitory concentration (MIC) value of 2.0 μg/mL as assessed by the conventional microdilution assay according to the Clinical & Laboratory Standard Institute (CLSI) guideline (10) (Table 1). In the previous study, we used different media (LBN; LB media containing 15 mM nitrate) for either surface cultures on the plates or submerged cultures under shaking (i.e., vigorous aeration/agitation). Therefore, we exploited the same culture conditions for MIC determination to verify the differential PMB resistance of the two strains. As shown in Table 1, the less MIC values were observed in the shaking condition than in the conventional assay condition for the tested PA strains, including PA14 and PAO1 as well as the PMB-resistant clinical isolates (P26 and P80) (24). This confirmed the quantitative difference in antibacterial activity of PMB between both culture conditions. Duetz et al. (14) reported that agitation conditions affect oxygen transfer rate (OTR) of the culture media: the OTR of nutrient broth was increased from 12.5 to 20.7 mmol O2 L−1·h−1 just by 75° angled agitation. Moreover, surface plate cultures are most likely more oxic than submerged broth cultures. Thus, our previous culture conditions might have ensured higher OTR than the conventional microdilution assay condition. Since PMB involves ROS generation for its bactericidal activity (12), it is most likely that the alteration in the oxygen utilization of PA strains might affect their resistance to PMB.
Minimum Inhibitory Concentrations (μg/ml) of Polymyxin B Against Pseudomonas aeruginosa Strains Under Different Conditions a
MIC values were obtained using both LBN broth and MH either under shaking (5-mL test tube) or conventional (96-well microtiter plate) conditions.
Clinical isolates of colistin-resistant PA strains.
LBN, LB supplemented with 15 mM KNO3; MH, Müller–Hinton II; MIC, minimum inhibitory concentration; PA, Pseudomonas aeruginosa.
It is noted that, however, even under the shaking condition, PAO1 displayed higher MIC values than PA14 did in LBN but not in Müller-Hinton II broth (Table 1), suggesting that PAO1, but not PA14, showed some resistance to PMB in LBN only under agitation condition as in the surface culture conditions as our previous study (19). This “operational” or “intrinsic” PMB resistance that PAO1 displays clearly differs from the resistome-mediated acquired resistance of P26 and P80, because the resistance was conditionally observed (i.e., in LBN only under shaking condition in vitro).
The intrinsic PMB resistance of PAO1 involves NR
In an attempt to elucidate the physiological clues associated with the intrinsic PMB resistance of PAO1, we compared the transcriptomes of PAO1 and PA14 in LBN broth under shaking condition (Supplementary Table S1), which identified the genes differentially regulated in both strains. Most of the upregulated genes by more than twofolds in PAO1 belong to the genes involved in NR even in the normal aerobic culture condition. This observation is in a good agreement with our previous study that the aerobic NR triggers the expression of katAp2, which is dependent on Anr, the master regulator of the NR in PA (9, 39, 44). To verify the transcriptome results, the promoter activities of the selected genes (nirS for nitrite reductase and fhp for flavohemoprotein) were examined in both PAO1 and PA14 by using lacZ transcriptional fusion (Fig. 1A). The higher transcription activities were observed in PAO1 than in PA14, indicating that PAO1 has higher NR activity than PA14. We also measured the mRNA levels of the nirS gene in the PMB-resistant strains (P26 and P80) (Fig. 1B). P26 and P80 have a mutation in pmrA and phoQ, respectively, which induce the overexpression of the pmrH gene required for the PMB resistance (24). The similar nirS mRNA levels of P26 and P80 as that of PA14 indicate that the acquired PMB resistance in P26 and P80 is not associated with higher NR and that the mechanism of the intrinsic PMB resistance of PAO1 might differ from those of the acquired PMB resistance. Besides the gene expression results, the higher NR activity in PAO1 than in PA14 was also confirmed by the higher steady-state level of nitrite (NO2−) (Fig. 1C), in that nitrite, a stable intermediate in NR is one of the tractable biomarkers of NR and/or nitric oxide production (27).

To verify whether NR is indeed necessary for the intrinsic PMB resistance of PAO1, we exploited the anr mutants of PAO1 and PA14, which are completely defective in NR (43) and measured their MIC values. As shown in Figure 2, unlike the PA14 anr mutant, whose susceptibility to PMB was no less than that of the isogenic wild type, the PAO1 anr mutant showed lower MIC value than the wild type (i.e., 0.25 μg/mL vs. 2.0 μg/mL) under LBN-agitation condition, similar to the MIC values of PA14 and the PA14 anr mutant (i.e., 0.25 μg/mL). All these results led us to the conclusion that the intrinsic PMB resistance of PAO1 might involve the Anr-mediated upregulation of the NR genes and the subsequently increased NR activity under LBN-agitation condition.

NR-associated PMB resistance is independent of KatA
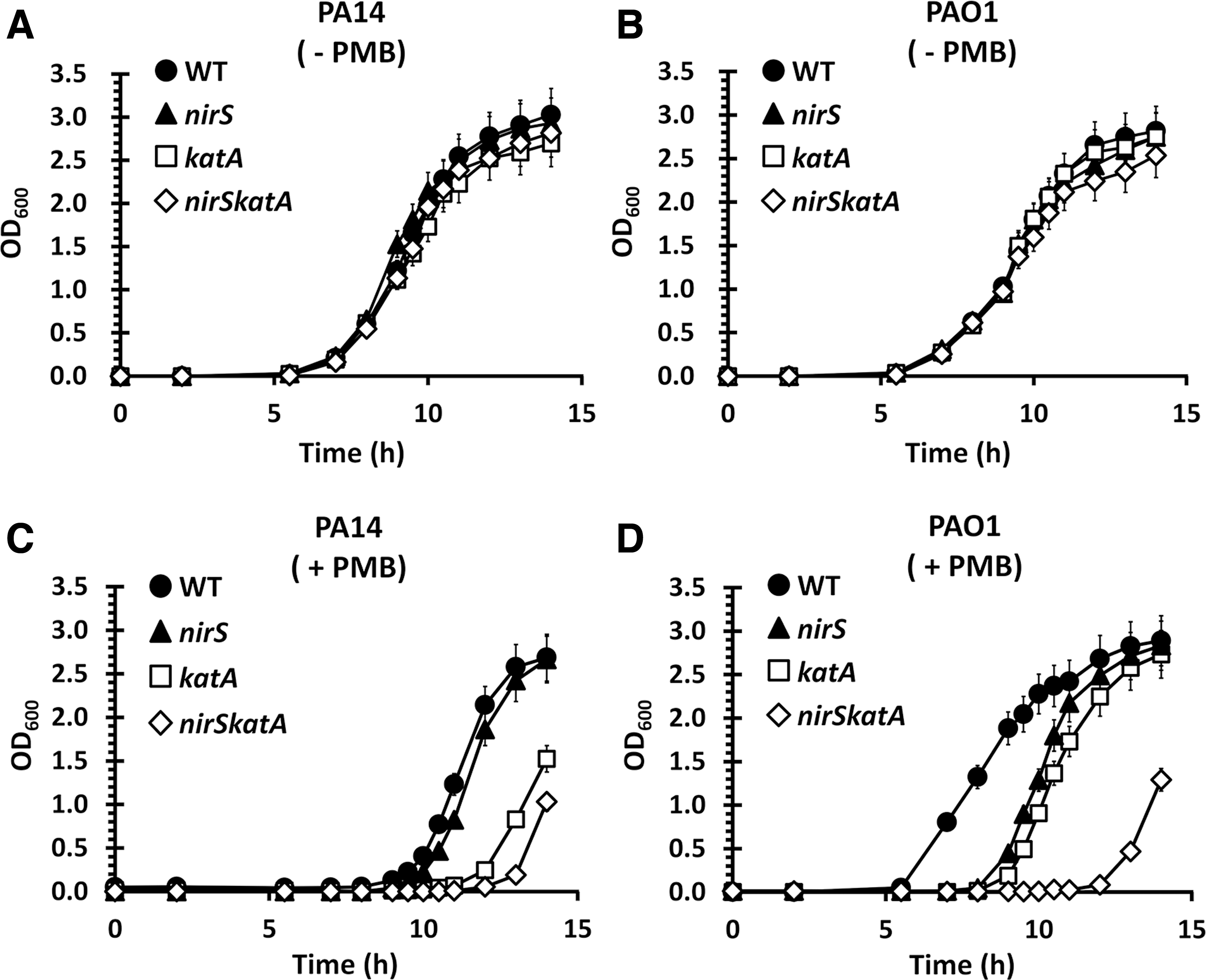
KatA, the major catalase of PA, is preponderantly responsible for detoxifying H2O2 and its expression is upregulated under NR conditions (9). More importantly, our recent observation that the KatA expression is higher in PAO1 than in PA14 (19) has prompted us to investigate whether the NR-mediated upregulation of KatA might be necessary for the intrinsic PMB resistance of PAO1. We first generated the nirSkatA double mutants of PAO1 and PA14 to address the genetic interaction between NR and KatA on the PMB resistance. The created mutants were tested for their ability to grow in the prolonged exposure to PMB (0.125 μg/mL) in LBN broth as described previously (19). As shown in Figure 3, no difference was observed between the wild type and the nirS mutant as well as between the katA and the nirSkatA mutants in the PA14 background. However, as expected from Figure 2, the PAO1 nirS mutant showed similar susceptibility as the wild type PA14 or the PA14 nirS mutant (Fig. 3C). Interestingly, the PAO1 nirS mutant showed similar PMB susceptibility as the PAO1 katA mutant, whereas the PAO1 nirSkatA mutant was even more susceptible to PMB than the PAO1 nirS mutant (Fig. 3D). This result indicates that the NR-mediated intrinsic PMB resistance of PAO1 is independent of (i.e., not hypostatic to) the KatA function, although it is also probable that reactive nitrogen intermediates such as nitric oxide generated during NR might induce KatA expression, the KatA-independent contribution of NR is evident, most likely affecting different step(s) of the PMB toxicity.
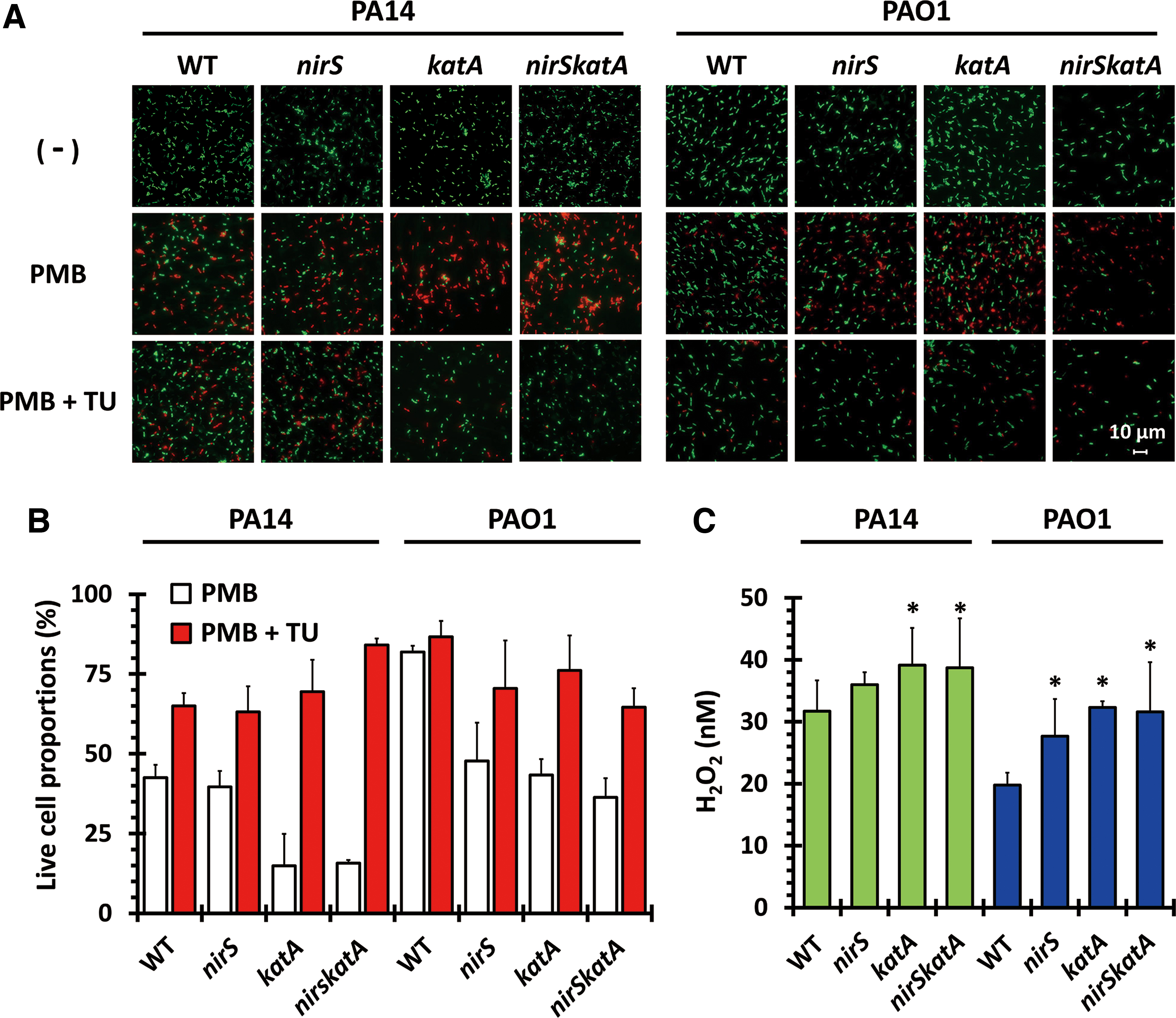
To gain a mechanistic insight into the NR-mediated PMB resistance, we investigated whether or not the PMB susceptibility of the nirS mutant could be recovered by an ROS quenching chemical, thiourea (TU) (18). The nirS, katA, and nirSkatA mutants were pulse-treated with PMB (2.0 μg/mL for 1 h) followed by TU treatment (5 mM), then the surviving cell proportions were counted after fluorescent staining of the live and dead cells (Fig. 4A, B). Clearly, based on the proportions of dead cells, it is notable that all the nirS and/or katA mutants survived the PMB treatment in the presence of TU. This result suggests that NR could reduce the accumulation of ROS presumably in part by bypassing oxygen respiration, the main source of ROS generation. Considering that PMB treatment can induce ROS generation (12, 26, 36), we were prompted to address whether the PMB treatment could generate ROS or just require ROS without affecting the amount of ROS. To this end, the steady-state level of H2O2 was measured in those mutants upon PMB treatment by using H2O2-specific dye, Amplex Red. As shown in Figure 4C, more ROS accumulation (31.7 nM of H2O2) was observed in PA14 than in PAO1 (19.76 nM H2O2). Most importantly, the H2O2 level in the PAO1 nirS mutant was increased to 27.68 nM, indicating the possibility that higher NR activity in PAO1 effectively mitigates the ROS accumulation. Taken together, higher KatA and NR activities are deemed required for the intrinsic PMB resistance in PAO1. This dismantling of the contributions of KatA and NR to the PMB resistance suggests that NR could prevent ROS generation, whereas KatA detoxifies generated ROS.
Intrinsic PMB resistance compromises the PMB treatment in infection conditions
To observe the significance of this NR-mediated intrinsic PMB resistance of PAO1 in infection conditions, we have investigated the antibacterial efficacy of PMB using the Drosophila systemic infection model. Flies were infected with both strains and fed with PMB after infection to see if the intrinsic PMB resistance can compromise the antibacterial activity of PMB in the infection condition. As shown in Figure 5A, PMB treatment (2 mg/mL per vial) was able to rescue the flies from the mortal infection caused by PA14. In contrast, however, the PAO1-infected flies were not rescued by PMB treatment (Fig. 5B). More importantly, the involvement of NR in the PMB resistance of PAO1 was also confirmed, in that the nirS mutation compromised the PMB resistance in PAO1. It is also noted that the mortality of the PA14 nirS mutant was no less than that of the isogenic wild type, irrespective of PMB treatment. This result underlines that the NR-mediated intrinsic PMB resistance of PAO1 is sufficient to overcome the antibacterial efficacy of PMB in infection conditions.

NR correlates with the intrinsic PMB resistance in PA clinical strains
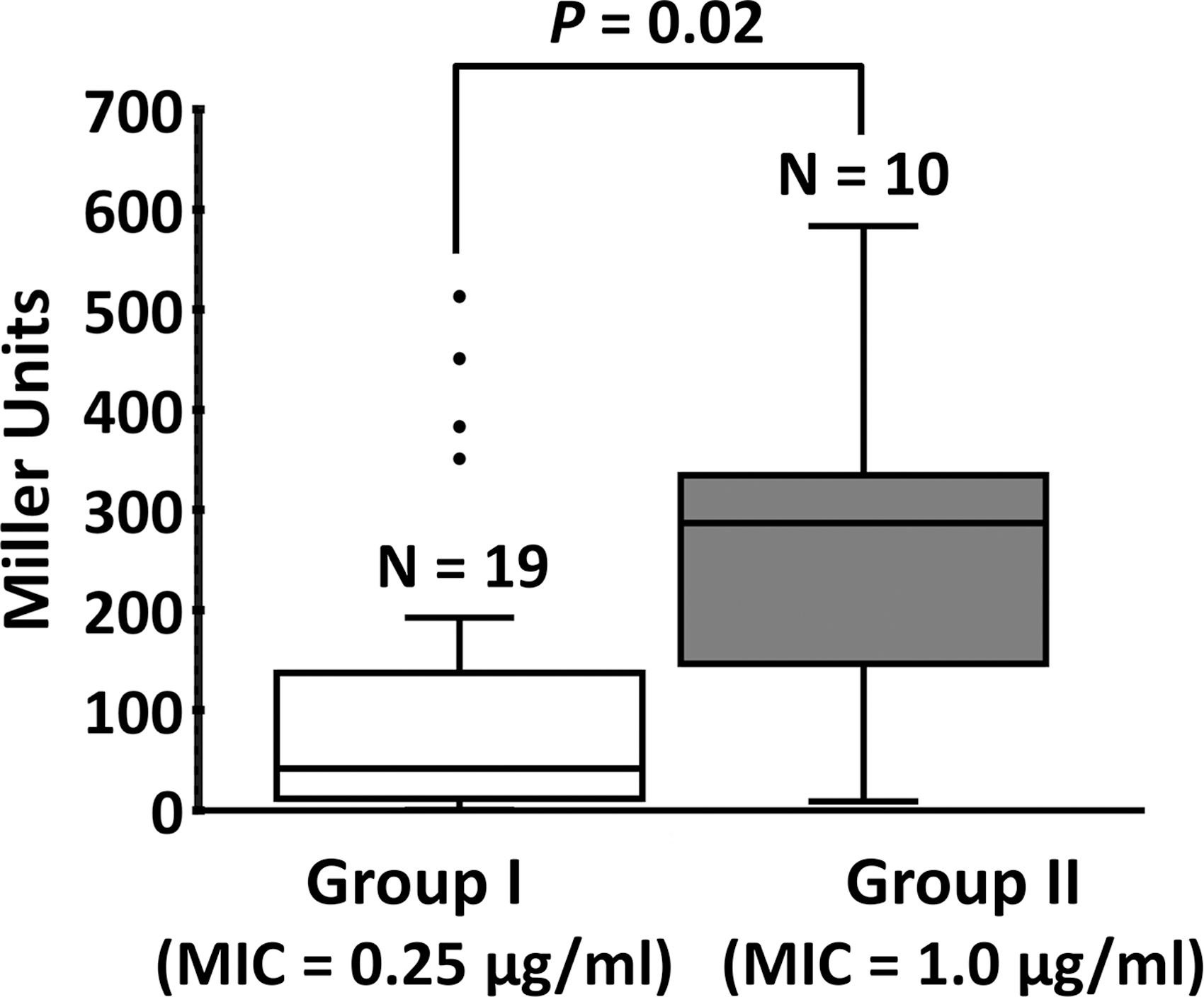
The aforementioned results are supporting that the higher NR activity in PAO1 is an important determinant for the intrinsic PMB resistance, which is not observed in PA14. We have wondered how common this strain-specific intrinsic resistance is in clinical PA isolates. To this end, the MIC values were measured for the PA isolates in LBN under both shaking and conventional assay conditions. Based on the differences between MIC values measured in both conditions, the tested 29 strains were divided into 3 groups (I to III) (Supplementary Table S2): 19 group I strains, including PA14, showed no PMB resistance, whereas 10 group II strains, including PAO1, showed intrinsic PMB resistance. The group III strains (P26 and P80) exhibit acquired PMB resistance.
To test whether the group I and II strains might differ in NR activity, the nirS-lacZ fusion construct was introduced into the clinical PA strains and β-galactosidase assay was performed as in Figure 1A (Fig. 6). It was conspicuous that most (15 out of 19) of the group I strains showed relatively lower nirS promoter activities, whereas the 10 group II strains showed higher nirS promoter activities. A few outliers existed especially among the group I strains, presumably due to other contributing factors, including differential expression of KatA (19) and/or the resistance determinants such as efflux pumps and membrane modification (13, 29). Nevertheless, the statistically significant correlation between the nirS promoter activity (i.e., NR) and the intrinsic PMB resistance in clinical PA strains suggests that the NR-mediated PMB resistance might be a relatively common trait of PA strains in the clinical settings.
Discussion
This is the first study to highlight the function of NR in PMB resistance, initially based on the comparison using two widely used P. aeruginosa (PA) laboratory strains, PAO1 and PA14. It becomes more and more important to reincarnate the old drugs such as PMB into the modern antibacterial regimen in the era of antimicrobial resistance, which has been facilitated by the recent advancement in the knowledge about the antibacterial and resistance mechanisms of the old drugs. This study provides an important insight into another layer of the molecular mechanisms of antibiotic resistance to PMB in PA strains and a key consideration for treatment options against PA infections, given that PMB is currently one of the last-resort antibiotics for the treatment of infections caused by the multidrug-resistant Gram-negative bacteria.
The bactericidal mechanism of PMB is generally attributed to its ability to bind to the OM lipid A. The classical explanation is that PMB-lipid A binding results in membrane dysfunction due to membrane lysis and/or component exchange at the OM and the CM through its hydrophobic parts such as the N-terminal fatty acyl chain and the amino acid residues such as Phe and Leu. An alternative mechanism that has been recently suggested is the involvement of highly toxic ROS that can be generated through aberrant electron transfers in the PMB-perturbed membrane. Superoxide anion (O2 −) can be formed upon entry of PMB into and cross OM and CM (36). O2 − is subjected to enzymatic conversion to hydrogen peroxide (H2O2), which leads to the formation of the most toxic ROS, hydroxyl radical (•OH) through Fe2+ oxidation to Fe3+, which reverts back to Fe2+ by absorbing an electron from another molecule of H2O2 (Fenton reaction) or from O2 − (Harber–Weiss reaction).
Despite some disputes still going on (21, 28), ROS generation is regarded as a common killing mechanism for the bactericidal antibiotics (21, 26). It is yet to be ascertained that ROS generation is the sole or at least the primary contributor to the antibacterial activity of PMB. However, the data in this study validated that PAO1, but not PA14, has intrinsic resistance to PMB directed by higher NR activity. Therefore, it is certain that ROS generation can be regarded as an important contributor to the antibacterial mechanism of PMB in this context. More importantly, this intrinsic resistance of PAO1 is sufficient to abrogate the antibacterial activity of PMB in Drosophila systemic infections and some clinical PA isolates have higher NR activity, displaying higher MIC in vitro. Thus, it needs to pay more attention to use PMB for the treatment of multidrug-resistant PA infections.
Yoon et al. (41) previously showed that PA respires on nitrate during chronic lung infections and tends to form robust biofilms during anaerobic growth. This implies that PMB resistance might be triggered for some PA strains under the NR and/or biofilm growth conditions, which needs to be further investigated in the relevant infection conditions. Recently, the PMB resistome of PA14 was characterized using its nonredundant transposon mutant library, which identified the genes involved in the intrinsic layer of resistance in PA14 (16). Although they might have provided a comprehensive snapshot of the PMB resistance mechanisms in PA strains, we were able to identify another layer of the PMB resistance that was more evidently observed in the PA strains with higher NR activity as PAO1. Since the higher NR activity is deemed required for successful chronic infections by PA, targeting the physiological determinants to trigger NR would be combined with PMB treatment to ensure the antibacterial efficacy of PMB to treat chronic infections. Because the intrinsic resistance and the associated mechanisms involving NR diminishes the effectiveness of PMB and potentially other related antibiotics, the successful inhibition of NR can be considered as an augmentation therapy. More importantly, the fact that the enzymes in NR are not found in human and livestock prompts us to design specific inhibitors targeting these enzymes for the successful treatment of the chronic PA infections.
Materials and Methods
Bacterial strains and culture conditions
All bacterial strains and plasmids used in this study are listed in Table 2. Strains of P. aeruginosa (PA) were grown at 37°C using Luria–Bertani (LB) broth, LB containing 15 mM KNO3 (LBN) or on 2% Bacto-agar (Difco) LB plates for 18 h. Bacterial cultures that had been grown to the late stationary growth phase (OD600 of 5.0) were inoculated into LB broth with an inoculum of 1.6 × 107 colony forming units (cfu)/mL and then grown to the late-logarithmic growth phase (OD600 of 0.7) before being used for experiments.
Strains Used in This Study
The deletion constructs were generated for both PA14 and PAO1.
The promoter fusion constructs were generated using either PA14 or PAO1 chromosome.
CbR, carbenicillin- and ampicillin-resistant; RifR, rifampicin-resistant; SmR, streptomycin-resistant; TpR, trimethoprim-resistant.
Oligonucleotide primers
The DNA oligonucleotide primers used for gene deletion, expression, and detection in this study are listed in Supplementary Table S3.
Generation of deletion mutants
All the in-frame deletion mutants were created using pEX18T. Oligonucleotide primers were designed using the PAO1 genome sequence. anr, nirS, and nirSkatA mutants were created by SOEing (splicing by overlap extension) polymerase chain reaction (PCR) as described previously (23), using four oligonucleotide primers listed in Supplementary Table S3. The resulting PCR products were cloned into pEX18T and then introduced into both PAO1 and PA14 chromosomes. In general, >30% of the coding regions were deleted for all the deletions, which were verified by PCR.
Measurement of MICs
The MICs of PMB were determined against 31 PA strains using the standard microbiological procedures according to the CLSI guideline, as per document no. (M7-A10) and the 5-mL test tube assay for vigorous agitation. In brief, bacterial cells that had been grown to OD600 of 0.7 were diluted to 5.0 × 105 cfu/mL and mixed with the equal volume of 30 mM KNO3-amended LB containing serially diluted PMB (from 16 μg/mL to 125 ng/mL) or Müller–Hinton II broth with PMB (from 16 μg/mL to 125 ng/mL). Then, the mixtures were grown at 37°C for 18 h either with 96-well microtiter plates in a shaking incubator (HANBEAK, Korea) (200 rpm) or with 5-mL test tubes in a rotator (Glas-col®) (80 rpm). The MIC values were recorded as the lowest concentration at which no signs of growth were observed based on the OD600 value of <0.05 after 18-h incubation.
Transcriptome analysis
Affymetrix PAO1 GeneChip was used for microarray analysis. RNA was isolated from the late logarithmic-growth phase (OD600 of 0.7) cultures in LBN broth of either PAO1 or PA14. Trizol (Invitrogen) was used and the samples were treated according to the manufacturers' recommendation. The eluted RNA was treated with DNase I for 1 h at 37°C. After enzyme inactivation at 65°C for 1 h, further treatment of RNA samples was done as described by the manufacturer (Affymetrix) with minor modifications as suggested elsewhere (40). cDNA synthesis, fragmentation, and labeling and hybridization, and washing were performed according to the manufacturer's instruction. Three independent experiments were performed using independent RNA preparations. The data were analyzed using Affymetrix microarray suite software suite (MAS) (version 5.0) with the default parameters. Gene expression changes were calculated by comparisons of the signal averages from PAO1 and PA14 cultures. The statistical significance of the gene expression changes was analyzed according to the Mann–Whitney U test with a cutoff p-value of 0.05 (8).
Construction of lacZ fusions and β-galactosidase assay
The transcriptional fusion of nirS were created by SOEing PCR using four oligonucleotide primers (Supplementary Table S3). In brief, the nirS promoter regions from both PAO1 and PA14 were prepared using the primer pair (nirS-lacZ-F and nirS-lacZ-UR) and the cognate chromosomes. The partial lacZ coding region was amplified using the primers pair (nirS-lacZ-DF and pQF50-R1) and pQF50. Then, the PCR products were fused by SOEing PCR using nirS-lacZ-F and pQF50-R1 and the amplified products were cloned into pQF50. The fhp transcriptional fusions were created as described elsewhere (2) with the primers listed in Supplementary Table S3. The lacZ fusion constructs were then used to transform the cognate PA strain by electroporation and the pQF50-nirS for PAO1 was used to transform the clinical PA strains except for PA14. LacZ (β-galactosidase) activity was determined as previously described (20). The results are presented as the means with standard deviations and were analyzed by unpaired t-tests using GraphPad Prism 5.0 statistical software. For box and whisker plotting, unpaired t-tests were performed to analyze one-tailed p-value with confidence intervals of 95%. A p-value of <0.05 was considered statistically significant.
Real-time PCR
RNA was extracted from the late-logarithmic growth phase (OD600 of 0.7) cultures with RNeasy Mini Kit (QIAGEN, Germany) according to the manufacturer's instruction. The cDNA samples that had been synthesized from the RNA samples (1.0 μg/mL) using ReverTra Ace qPCR RT kit (Toyobo, Japan) were subjected to qPCR with Thunderbird SYBR qPCR Mix (Toyobo) for StepOnePlus qPCR system (Applied Biosystems). The cDNA synthesis and qPCR primers are listed in the Supplementary Table S3.
Measurement of NO2
−
accumulation
The steady-state level of NO2 − during the growth in LBN was measured using the modified Griess reagent (Sigma). In brief, the cells that had been grown up to the OD600 of 3.0 were centrifuged at 8000 g for 5 min and the supernatants were collected. The aliquot (100 μL) from the supernatants at the indicated time points were mixed with the equal volume of modified Griess reagent and incubated at 25°C for 15 min. The NO2 − concentrations were calculated from the absorbance of the supernatants that had been measured in 96-well microtiter plates at 540 nm. The measurements were repeated three times and the results were presented as the means with standard deviations.
PMB-mediated growth inhibition
Bacterial cells that had been grown to OD600 of 0.7 were diluted to 5.0 × 105 cfu/mL in LBN broth with 125 ng/mL of PMB, then incubated in a rotator (Glas-col) under shaking (80 rpm) for 15 h at 37°C. Aliquots (100 μL) from the cultures were collected at the indicated time points for OD600 measurement.
Microscopic observation of cell death
Live/Dead-BacLight® (Thermo-Fischer) was used to observe the cell death. In brief, cells that had been grown to OD600 of 0.5 were pulse-treated with 2.0 μg/mL of PMB in LBN broth with or without 5 mM TU (Sigma) in the rotator under shaking (80 rpm) for 1 h at 37°C. After incubation, 1 mL of each culture was washed with PBS three times and resuspended in the equal volume of PBS and 3 μL of the dye was added to the resuspension, followed by incubation for 15 min in the dark. The stained cells were mounted on a 2% agarose pad for fluorescence observation using a fluorescent microscopy (Zeiss, Germany). Two single channel (green: viable cells, red: dead cells) pictures of three different random locations were selected and saved for further analysis. The selected pictures were then converted into 8-bit images using ImageJ (version 1.52k) for cell counting. Particle analyzer (default) and cell-counter (add-on) were used to count the numbers of viable and dead cells. The counting results were presented as the means with standard deviations of the three independent experiments.
Measurement of H2O2 accumulation
H2O2 accumulation was measured using the Amplex® Red Hydrogen Peroxide assay kit (Thermo-Fischer). In brief, aliquots (50 μL) of the cells that had been grown for the indicated time points in the rotator (Glas-col) under shaking (80 rpm) with LBN containing 125 ng/mL of PMB were mixed with the equal volume of Amplex Red reagent, followed by incubation for 30 min. The fluorescence was measured using 96-well microtiter plates at 580 nm by excitation at 550 nm. H2O2 concentrations were calculated by using a standard curve. The measurement was repeated three times and the results were presented as the means with standard deviations. The results were analyzed by unpaired t-test with GraphPad Prism 5.0 statistical software. A p-value of <0.05 was considered statistically significant.
Determination of antibacterial efficacy in vivo
To determine antibacterial efficacy of PMB in vivo, Drosophila systemic infection was performed as previously described (20). PA cells that had been grown to OD600 of 3.0 were diluted into 10 mM MgSO4 (up to 107 cfu/mL). Drosophila melanogaster strain Oregon R was grown at 25°C with 5 mL of corn meal-dextrose medium (0.93% agar, 6.24% dry yeast, 4.08% corn meal, 8.62% dextrose, 0.1% methyl paraben, and 0.45% [v/v] propionic acid). For infection, 4- to 5-day-old adult female flies were infected by pricking at the dorsal thorax with a 10 μm needle (Ted Pella) that had been dipped in the bacterial suspension in 10 mM MgSO4. Infected flies were transferred to a new medium overlaid with PMB at the final concentration of 2 mg/mL. Survival rate of infected flies were monitored for up to 54 h postinfection. Flies that died within 12 h were excluded in mortality determination. Mortality assay was repeated at least three times. Log-rank tests were used to compare the survival rate difference between the groups. A p-value of <0.05 was considered to be significant.
Authors' Contributions
Y.-H.C. conceived and designed the research. B.-o.K, H.-J.J., and I.-Y.C. designed and performed the experiments, and collected and analyzed the experimental data. H.-W.B. and E.S.K. provided reagents. B.-o.K., H.-J.J., and Y.-H.C. wrote the article. All authors reviewed the article.
Footnotes
Author Disclosure Statement
No competing interests exist.
Funding Information
This study was supported by the National Research Foundation of KOREA (NRF) Grant (NRF-2017R1A2B3005239).
Supplementary Material
Supplementary Table S1
Supplementary Table S2
Supplementary Table S3
Abbreviations Used
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.