
Abstract
Aims:
Diabetic nephropathy (DN) is characterized by microalbuminuria, mainly associated with pathological and morphological alterations of podocyte. New drug targeting podocyte injury is a promising approach for treating DN. The present study is aimed at developing new drug targeting podocyte injury for treating DN.
Results:
In this study, germacrone ameliorated kidney damage and inhibited podocyte apoptosis in a DN mouse model. Based on RNA-seq, mmu_mmu_circRNA_0000309, located in host gene vascular endothelial zinc finger 1 (Vezf1), showed a sharp decline in DN mice and a remarkable recovery in germacrone-challenged DN mice. mmu_circRNA_0000309 silence or miR-188-3p mimics abrogated the antiapoptosis and anti-injury effects of germacrone through aggravating mitochondria damage, and elevating reactive oxygen species and ferroptosis-related protein levels. Mechanistically, mmu_circRNA_0000309 competitively sponged miR-188-3p, and subsequently promoted glutathione peroxidase 4 (GPX4) expression, thereby inactivating ferroptosis-dependent mitochondrial damage and podocyte apoptosis. In addition, GPX4 overexpression neutralized mmu_circRNA_0000309 silence-mediated mitochondria damage and ferroptosis in germacrone-exposed MPC5 cells.
Innovation:
We describe the novel effect and mechanism of germacrone on treating DN, which is linked to ferroptosis for the first time.
Conclusion:
mmu_circRNA_0000309 silence mediates drug resistance to germacrone in DN mice. mmu_circRNA_0000309 sponges miR-188-3p, and subsequently upregulates GPX4 expression, inactivating ferroptosis-dependent mitochondrial function and podocyte apoptosis. Possibly germacrone-based treatment for DN can be further motivated by regulating mmu_circRNA_0000309/miR-188-3p/GPX4 signaling axis. Antioxid. Redox Signal. 36, 740–759.
Color images are available online.

Introduction
Diabetes is a common chronic metabolic disease (44). Diabetic nephropathy (DN) is the prevalent complication both in type I and II diabetes, which is recognized as the dominated cause of end-stage renal disease (4, 42). The most common characteristic of DN is microalbuminuria, which may be associated with pathological and morphological alterations of podocyte (3, 5, 24). Current research shows that the damage, detachment, epithelial/mesenchymal transdifferentiation, and apoptosis of podocyte are the main causes for DN (28, 40). Although many researchers are committed to developing new drug targeting podocyte injury for DN, the underlying reasons of aberrant podocyte damage have not been elucidated, and need to be further investigated.
Results
Germacrone treatment relieves DN situation
As shown in Figure 1A, the blood glucose concentration of db/db mice exceeded 16.7 mM at 12 weeks, which was significantly higher than the control C57BL/KsJ group, indicating that the type II diabetes mouse models were successfully constructed (Fig. 1A). Of note, 13-week db/db mice were accompanied by obvious DN characteristics, which displayed a significant increase of 24-h proteinuria, blood urea nitrogen (BUN), serum creatinine (Scr), and blood uric acid (BUA), and a declined urine creatinine (Ucr) compared with the control mice (Fig. 1B–F).
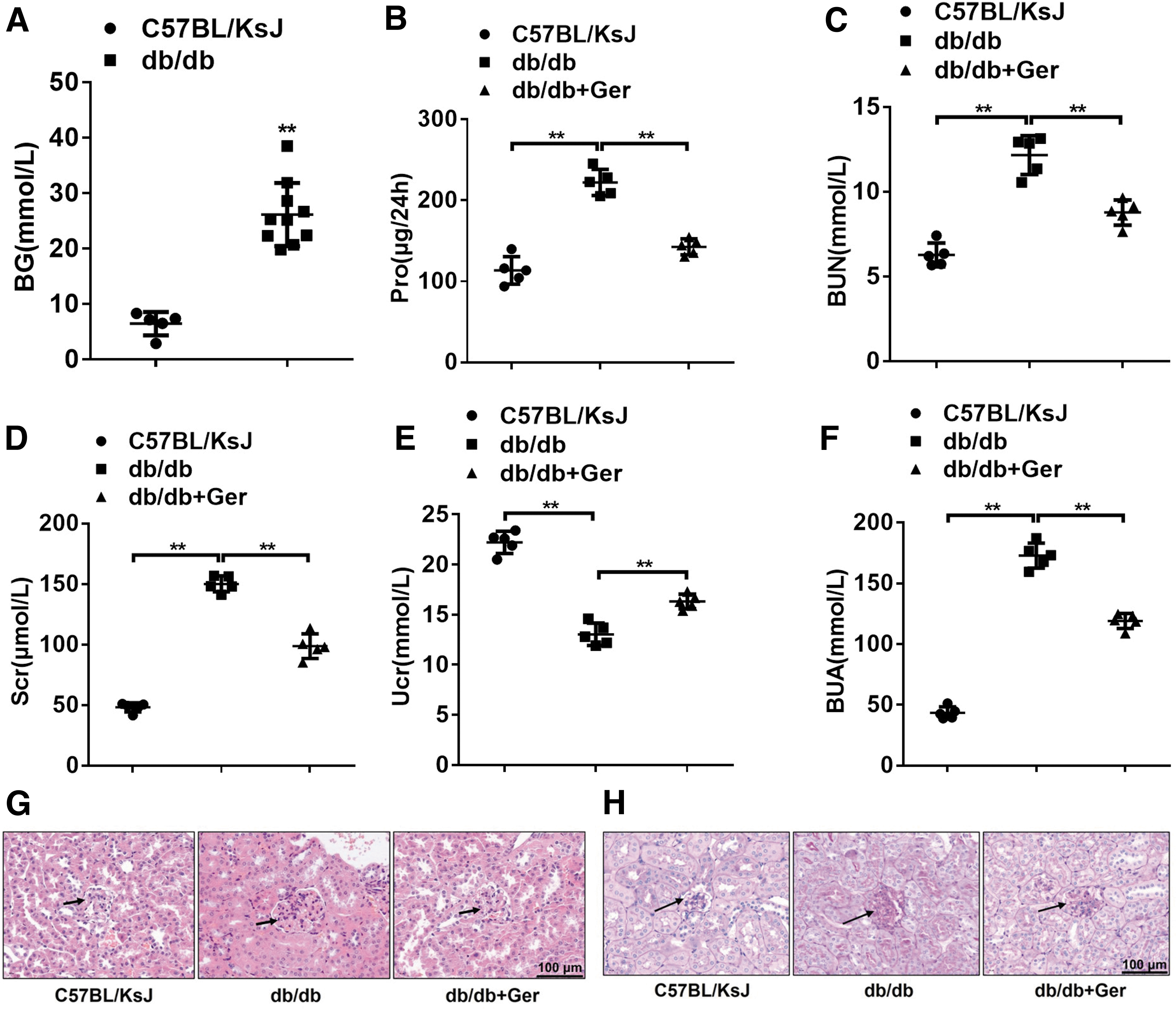
However, with the treatment of germacrone, the kidney function indicators of 24-h proteinuria, BUN, Scr, and BUA, all tended toward normalization (Fig. 1B–F), which suggested that germacrone could relieve DN symptoms in db/db mice. In the kidney tissues, glomerular hypertrophy and the basement membrane became thickened and contained fluid vacuoles in DN db/db mice (Fig. 1G, H). In germacrone-challenged mice, the glomeruli and basement membrane significantly recovered compared with the model mice (Fig. 1G, H). These phenomenal changes indicate that germacrone has a certain curative effect on DN.
Mmu_circRNA_0000309 is lowly expressed in podocytes of DN mice
Podocytes, as terminally differentiated glomerular epithelial cells, exert a predominant role in the glomerular filtration barrier. Podocyte variant forms, such as apoptosis and loss, can cause glomerular filtration membrane damage, thereby inducing unfavorable DN. Podocin and nephrin are highly expressed in podocytes as the podocyte foot markers (6, 33). Thus, podocytes were isolated from db/db mice and the purity was authenticated by cell immunofluorescence (Fig. 2A), podocin and nephrin in db/db mice were sickly expressed, which were notably rescued in germacrone-exposed mice, showing more podocin- and nephrin-positive cell populations (Fig. 2B, C).
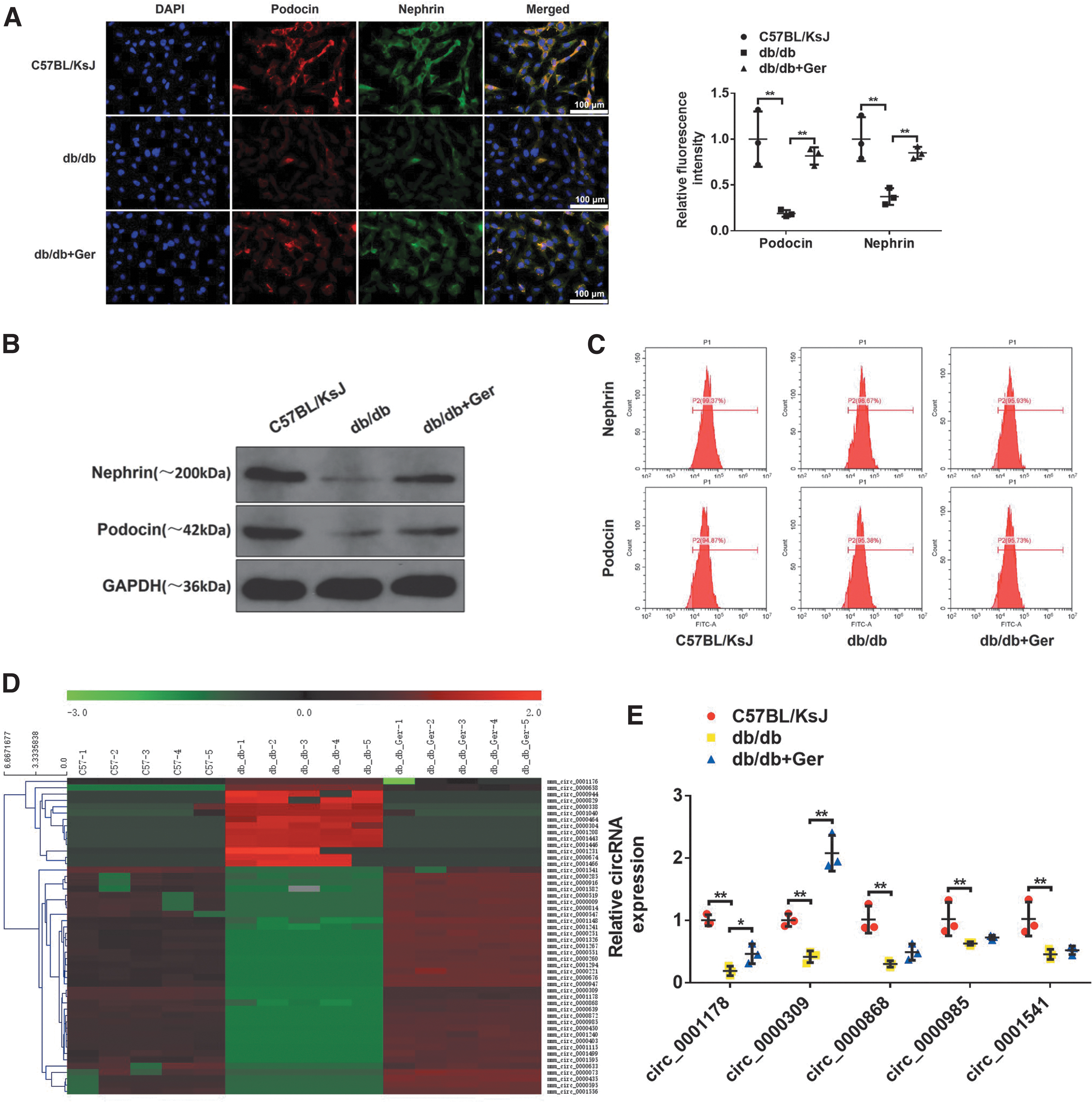
Next, we tried to screen the differential expression circular RNAs (circRNAs) in podocytes obtained from the normal control group, DN db/db mice, and germacrone-treated db/db mice using circRNA microarray analysis. The results of microarray data were presented in the heat map (Fig. 2D). Among the differential expression circRNAs, several remarkably downregulated circRNAs were observed under three conditions, including mmu_circRNA_0001178, mmu_circRNA_0000309, mmu_circRNA_0000868, mmu_circRNA_0000985, and mmu_circRNA_0001541 (
Innovation
Curcuma species play a protective effect on diverse kidney diseases, including diabetic nephropathy (DN). Germacrone is the principal bioactive component of Rhizoma Curcuma, which is discovered to exert a leading impact on many diseases, including antiinflammatory and antiapoptotic functions. Here, we describe the novel effect and mechanism of germacrone on treating DN, which is linked to ferroptosis for the first time.
Curcuma species play a protective effect on diverse kidney diseases, including diabetic nephropathy (DN) (22, 45). Germacrone is the principal bioactive component of Rhizoma Curcuma, which is discovered to exert a leading impact on many diseases, including anti-inflammatory and antiapoptotic functions (1, 32). It is shown that germacrone can lower rheumatoid arthritis and inflammatory infiltration via adjusting the balance of T helper type 1/T helper type 2 and inactivating the NF-κB signaling (55).
Germacrone treatment distinctly increases the expression of anti-inflammatory factor IL-10 (2), and exerts a remarkable protective effect on acute cerebral ischemia/reperfusion in rats through antioxidative and antiapoptotic activities (56), which is similar to the result that Curcuma oil (Curcuma, another extract, the active ingredient is germacrone) can diminish oxidative stress and against apoptosis in transient focal ischemia in rat brain (41). Thus, germacrone possesses anti-inflammatory, antiapoptotic, and antioxidative effects. Of note, there is evidence that oxidative stress, inflammation, and podocyte apoptosis play a pivotal role in the progression of DN (36).
Germacrone may be a good candidate agent for decreasing cisplatin-induced nephrotoxicity via inhibiting organic cation transporter 2 function (46). Possibly germacrone-mediated antioxidation and antiapoptotic actions may facilitate the improvement of DN, which should be explored further.
Ferroptosis, characterized by the accumulation of membrane lipid peroxidation products, results by the lack of activity of the glutathione peroxidase (GPX). Ferroptosis is an iron-dependent nonapoptotic cell death form (7, 48). Various reports demonstrate that ferroptosis is involved in lots of pathological processes of kidney diseases (26). Hu et al. have sufficiently described the role mechanism of ferroptosis on acute kidney injury (21). It is demonstrated that targeting iron-based statins can inhibit abnormal mitochondrial permeability to protect renal tubular cell death (30). Thus, the ferroptosis process possibly monitors disease progression in renal-related diseases.
Circular RNA (circRNA) is one of the endogenous noncoding RNAs that have a circular structure (12, 39). CircRNAs have emerged as a vital moderator in multiple diseases, including inflammation, liver function, cardiovascular disease, and DN progression (25, 58). Mmu_circRNA_15698 exacerbates the extracellular matrix of mesangial cells in DN mice through the miR-185/TGF-β1 axis, accompanied by the accumulation of collagen and fibronectin, resulting in kidney fibrosis (20). Mmu_circ_0000491 and mmu_circ-AKT3 are found to possess the disease acceleration function in DN (37, 50). It is reported that circRNAs play a role in an iron-dependent manner, for example, hsa_circ_IL4R promotes the occurrence of hepatocellular carcinoma through inhibiting ferroptosis via the miR-541-3p/GPX4 axis (59). However, whether ferroptosis implicates circRNA-regulated DN progression remains unclear.
In the present study, we investigated that a novel identified circRNA, mmu_circRNA_0000309, located in the host gene vascular endothelial zinc finger 1 (Vezf1) knockdown inhibited the protective effect of germacrone on DN via hindering podocyte apoptosis. Specifically, germacrone combined with exogenous mmu_circRNA_0000309 facilitated the treatment of DN through repressing ferroptosis-dependent mitochondrial damage and podocyte apoptosis by regulating the miR-188-3p/GPX4 axis, which offered distinctive insights to the management of the DN disease.
Reverse transcription-polymerase chain reaction (RT-PCR) results also confirmed that these candidate circRNAs were lower in db/db mice compared with K57BL/KsJ group, but only mmu_circRNA_0000309 and mmu_circRNA_0001178 expressions were rescued in germacrone-challenged db/db mice (Fig. 2E). However, the changes of cirRNA_0000309 are particularly dramatic. Thus, mmu_cirRNA_0000309 was chosen to perform the subsequent experiments. This indicates that mmu_circRNA_0000309 declines in DN mice and it may play an important role in germacrone-regulated DN progression.
mmu_circRNA_0000309 silence abolishes the effect of germacrone on high-glucose-induced podocyte damage
Once mouse podocyte cells (MPC5) were induced in low serum medium, mature podocytes were enlarged and rounded, with many long protrusions extended from the cell body. Under the stimulation of high glucose (HG), the morphology of MPC5 was notably changed, with fewer and shorter protrusions, resulting in cell apoptosis. However, most of the cells' morphology returned to star-shaped polydendrum after germacrone treatment (Fig. 3A). It had found that HG exposure led to a decrease in the population of MPC5; we further evaluated the antiapoptotic effect of germacrone by flow cytometry analysis.

As expected, HG-induced apoptotic cells and apoptosis-related proteins including cleaved poly-ADP-ribose polymerase (PARP) and caspase 3 were evidently abrogated by the pretreatment with germacrone (Fig. 3B, C, and Supplementary Fig. S1). Cell immunofluorescence staining results also confirmed that podocyte foot markers, including podocin and nephrin, were downregulated with HG exposure, while germacrone exposure inhibited these events (Fig. 3D, E).
To explore the role of mmu_circRNA_0000309 during germacrone-mediated improvement on DN, mmu_circRNA_0000309 shRNAs were synthesized. The knockdown efficiency of mmu_circRNA_0000309 shRNAs was verified by quantitative reverse transcription-polymerase chain reaction (qRT-PCR), and mmu_circRNA_0000309 shRNA-1 (the strongest effect) was chosen to perform the subsequent experiments (Supplementary Fig. S2). Interestingly, the antiapoptotic effects and the elevation effects on podocin and nephrin expressions afforded by germacrone were notably abrogated by mmu_circRNA_0000309 shRNA transfection (Fig. 3B–E). Collectively, these data suggest that germacrone attenuates HG-induced apoptosis in podocytes possibly through regulating mmu_circRNA_0000309 expression.
mmu_circRNA_0000309 knockdown prevents the effect of germacrone on DN
Next, we tried to exhaustively prove the connections between mmu_circRNA_0000309 and germacrone in a murine DN model. First, germacrone recovered the concentration of blood glucose in DN mice, while mmu_circRNA_0000309 shRNA injection prevented the protective role of germacrone (Fig. 4A). In addition, germacrone-mediated decrease of 24-h proteinuria, BUN, Scr, and BUA, and the increase of Ucr were all rescued by mmu_circRNA_0000309 knockdown in db/db mice (Fig. 4B–F).

Hematoxylin/eosin (HE) and periodic acid-Schiff (PAS) staining results exhibited that glomerular hypertrophy and the thickened basement membrane in germacrone-challenged DN mice were also reversed in the presence of mmu_circRNA_0000309 shRNA (Fig. 4G, H), which indicated that mmu_circRNA_0000309 knockdown can make germacrone lose its therapeutic action on DN.
mmu_circRNA_0000309 depletion abrogates the inhibitory effect of germacrone on ferroptosis
HG-induced podocyte apoptosis is closely related to mitochondrial injury in DN (14). Therefore, we focused on expounding the effect of germacrone/circRNA on mouse podocyte mitochondrial functions. The podocyte mitochondria were swollen accompanied by a lot of vacuole accumulation, and the mitochondrial membrane was broken and the ridge was also fractured in DN db/db mice (Fig. 5A). Germacrone treatment repaired mitochondrial abnormalities, but this effect disappeared with mmu_circRNA_0000309 knockdown (Fig. 5A). It has been confirmed that mitochondrial damage is mainly caused by the release of reactive oxygen species (ROS), especially lipid ROS, generated followed by the reduction of glutathione peroxidase 4 (GPX4) (8, 52).

GPX4 is a critical enzyme that maintains tissue homeostasis regulated by glutathione (g
Iron was highly expressed, and ferroptosis-related proteins cytochrome c oxidase subunit 2 (COX-2), long-chain-fatty-acid—CoA ligase 4 (ACSL4), and NADPH oxidase 1 (NOX1) were upregulated, and ferritin heavy chain 1 (FTH1) and GPX4 were downregulated in db/db mice. Although germacrone treatment reversed these changes, mmu_circRNA_0000309 knockdown similarly abolished germacrone's inhibitory effect on iron-dependent ferroptosis (Fig. 5F, G). These results illustrate that mmu_circRNA_0000309 knockdown reactivates mitochondrial oxidative damage and iron-dependent ferroptosis, resulting in podocyte apoptosis in germacrone-challenged DN mice.
Correspondingly, the protective effect of germacrone on HG-induced mitochondrial injury and the release of ROS and lipid ROS in MPC5 would be blocked by the decrease of mmu_circRNA_0000309 (Supplementary Fig. S3A–C). Then the JC-1 fluorescent probe was used to analyze the mitochondrial membrane potential (MMP). The results showed that germacrone moderately restored the JC-1 fluorescent content, which was hindered by the silence of mmu_circRNA_0000309, further suggesting that germacrone mediated the improvement of mitochondrial injury implicating with mmu_circRNA_0000309 (Supplementary Fig. S3D, E).
Parallelly, HG also accelerated the contents of iron and inhibited the GPX activity in vitro (Supplementary Fig. S3F, G). The ferroptosis-related proteins COX-2, ACSL4, and NOX1 were upregulated, while GPX4 and FTH1 were downregulated in HG-exposed MPC5 cells (Supplementary Fig. S3H). However, germacrone mediated the restore in these indexes, while mmu_circRNA_0000309 knockdown in turn abolished the role of germacrone (Supplementary Fig. S3F–H). Therefore, ferroptosis possibly implicated the process of mmu_circRNA_0000309 downregulation-mediated proapoptotic effects in germacrone-challenged podocytes.
Ferroptosis inhibition restores the protection afforded by germacrone in mmu_circRNA_0000309-deficient DN mice
To further illustrate the role of ferroptosis in germacrone/mmu_circRNA_0000309-regulated DN progression, liproxstatin-1 (Lip-1), a potent inhibitor of ferroptosis, was applied to interfere with the regulatory role on DN afforded by germacrone or mmu_circRNA_0000309. Consistently, germacrone mediated the decline of BG level, 24-h proteinuria, BUN, Scr, and BUA, and the elevation of Ncr level (Fig. 6A–F), glomerular hypertrophy, and the thickened basement membrane in DN mice (Fig. 6G, H) were all significantly impeded by mmu_circRNA_0000309 knockdown.

However, Lip-1 management almost completely restored the protective effects mediated by germacrone in mmu_circRNA_0000309-deficient DN mice (Fig. 6A–H). In addition to that, mitochondrial injury such as swollen mitochondria, lots of vacuole accumulation, and the broken mitochondrial membrane were relieved after germacrone administration, but received reinjury once treated with mmu_circRNA_0000309 shRNA in germacrone-exposed DN mice. By contrast, Lip-1 administration repaired mitochondrial abnormalities induced by mmu_circRNA_0000309 shRNA (Fig. 7A). Furthermore, mmu_circRNA_0000309 depletion obliterated the inhibitory effects of germacrone on the release of ROS and lipid ROS, and the promotion effects on the contents of GPX4 and GSH activity, which were rescued by the exposure of Lip-1 (Fig. 7B–E).
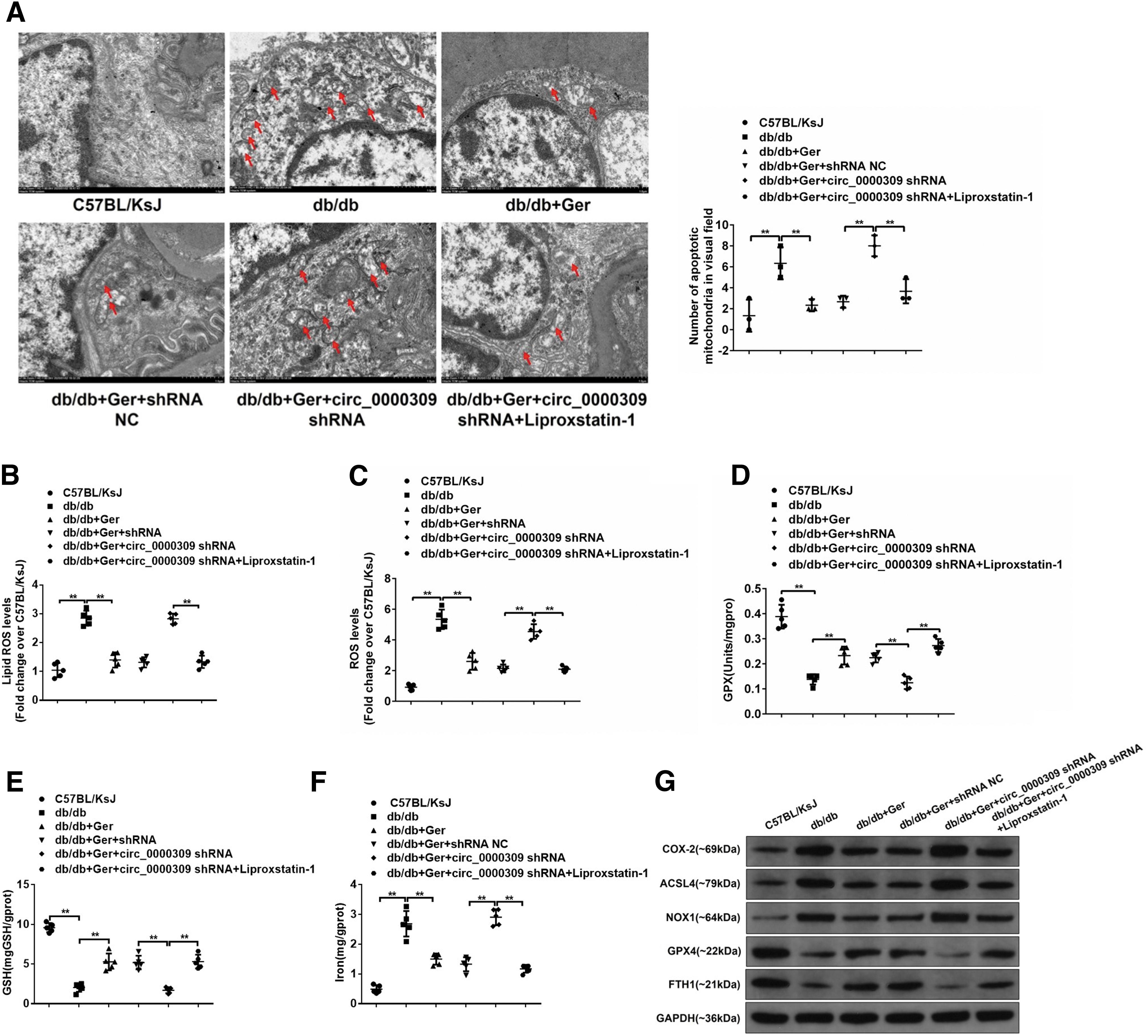
The blocking of mmu_circRNA_0000309 knockdown on the antiferroptotic effect afforded by germacrone was also reversed by the treatment of Lip-1 (Fig. 7F, G). These data indicate that germacrone/mmu_circRNA_0000309 signal pathway-modified DN development has the involvement of ferroptosis progress.
Regulation of mmu_circRNA_0000309/miR-188-3p/GPX4 axis in podocytes
To hunt for the mechanism of mmu_circRNA_0000309-modulated DN progression, we made full use of circBase (
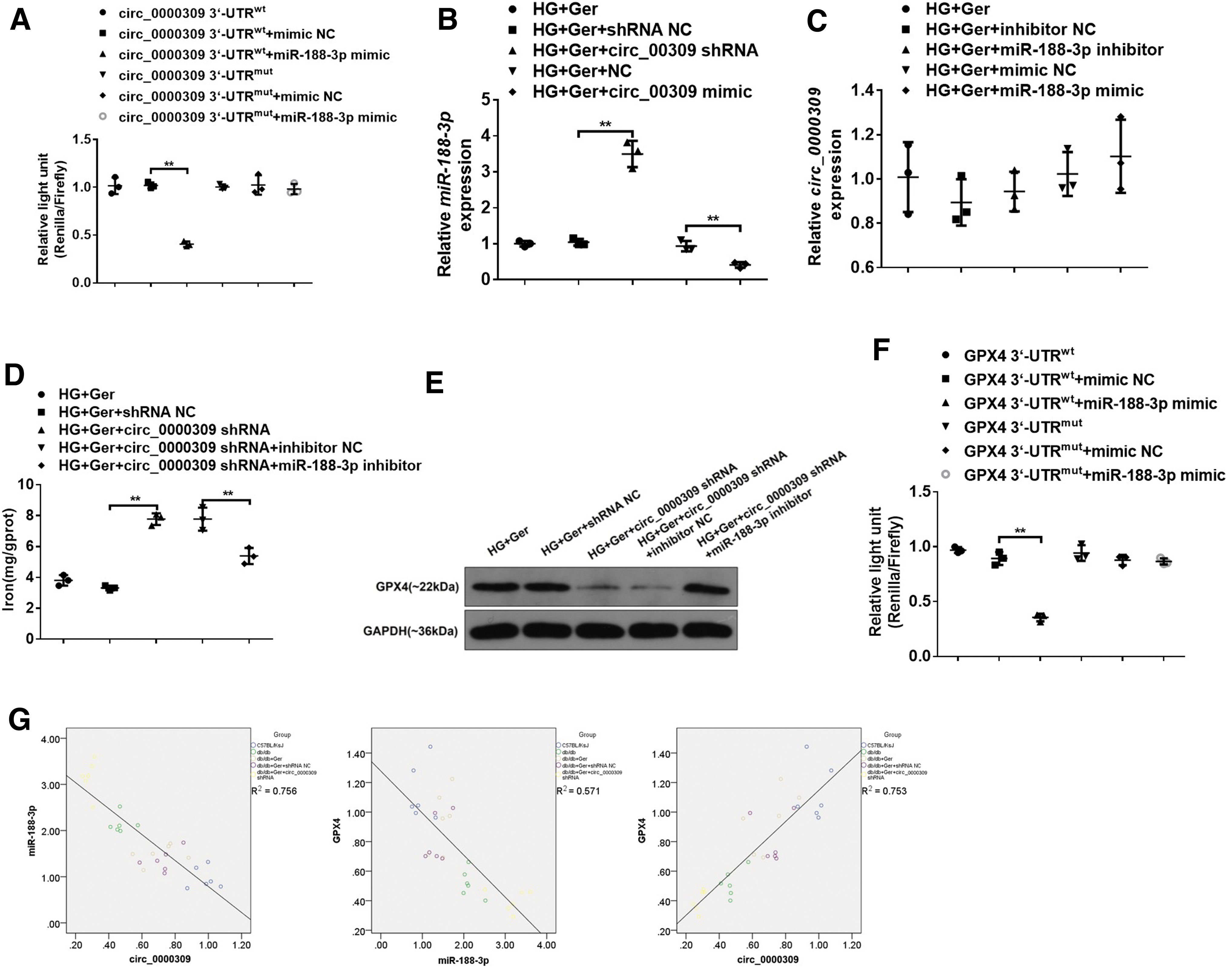
In addition, miR-188-3p expression was visibly enhanced with mmu_circRNA_0000309 knockdown in germacrone-challenged MPC5 cells and declined with exogenous mmu_circRNA_0000309 (Fig. 8B). By contrast, miR-188-3p had a faint effect on mmu_circRNA_0000309 expression (Fig. 8C).
Subsequently, the effect of miR-188-3p on HG-induced podocyte apoptosis was evaluated. As shown in Supplementary Figure S3, apoptotic cells and mitochonrial injury were apparently elevated by transfecting overexpression of miR-188-3p in germacrone-treated MPC5 cells compared with negative control miRNA-transfected cells in the presence of germacrone (Supplementary Fig. S4A–C). Besides, miR-188-3p mimics also prevented the relaxation effect of germacrone on mitochondrial ROS release, and germacrone mediated the elevated GPX activity and the reduction of iron contents (Supplementary Fig. S4D–G).
Germacrone-mediated improvement on the expression of ferroptosis-related proteins was also broken by miR-188-3p mimics (Supplementary Fig. S4H). Of note, mmu_circRNA_0000309 shRNA-mediated increase of iron and the decline of GPX4 expression were abolished by the miR-188-3p inhibitor (Fig. 8D, E), which manifested that miR-188-3p-targeted mmu_circRNA_0000309 and mmu_circRNA_0000309 induced the alteration of podocytes dependent on miR-188-3p. Subsequently, we monitored whether GPX4 was a potential binding protein of miR-188-3p.
Based on the TargetScan, miRD B, and PicTar databases, GPX4 was proven to be the downstream target of the mmu_circRNA_0000309/miR-183-3p axis. The results of the luciferase reporter gene also showed that miR-188-3p directly bound to GPX4 (Fig. 8F). The correlation analysis showed that GPX4 and mmu_circRNA_0000309 were positively correlated, while miR-188-3P was negatively correlated with GPX4 and mmu_circRNA_0000309 (Fig. 8G). These findings mainly signify that mmu_circRNA_0000309/miR-188-3p may join with GPX4 to regulate podocyte apoptosis and the progression of DN.
mmu_circRNA_0000309 regulates podocyte apoptosis in a GPX4-dependent manner
It was verified that GPX4 acted as the target protein of miR-188-3p, and was positively correlated with mmu_circRNA_0000309, and so, we speculated that mmu_circRNA_0000309 monitored ferroptosis-dependent mitochondrial functions through the miR-188-3p/GPX4 axis. Our experiments found that GPX4 overexpressed could give a facelift to the reduction of nephrin and podocin in podocytes caused by the lack of mmu_circRNA_0000309 (Fig. 9A). TUNEL staining results also indicated that exogenous GPX4 notably rescued the proapoptotic effect of mmu_circRNA_0000309 shRNA on podocytes in the presence of germacrone (Fig. 9B, C).

In addition to that, the mitochondrial vacuoles became smaller and fewer in GPX4-overexpressed podocytes compared with the normal podocytes under the cotreated condition of germacrone and mmu_circRNA_0000309 shRNA (Fig. 9D, E). Consistently, circ_0000309 shRNA-mediated release of lipid ROS and the elevation of iron contents, and the decrease of GPX activity and GSH level, were rescued by GPX4 overexpression in the presence of germacrone (Fig. 9F–I). In addition, germacrone inhibited ferroptosis accompanied by the decline of COX-2, ACSL4, and NOX1 and the increase of GPX4 and FTPH1 (Fig. 9J). Although circ_0000309 shRNA exposure abolished the effect of germacrone, proteins that promoted the ferroptosis process such as COX-2, ACSL4, and NOX1 were inhibited, while GPX4 and FTPH1 were impressively elevated in the GPX4-overexpressed cells (Fig. 9J).
These data emphasize that GPX4 overexpression can reverse mmu_circRNA_0000309 silence-mediated mitochondrial injury, leading to podocyte apoptosis by affecting the ferroptosis pathway.
To explore whether germacrone/mmu_circRNA_0000309 signal pathway-modified cell death mainly depended on the ferroptotic effect, cell apoptosis detection was performed in germacrone-alone-treated or germacrone combined with mmu_circRNA_0000309-treated MPC5 cells, which were also exposed with various inhibitors at the same time, including the ferroptosis inhibitor (Lip-1), autophagy inhibitor (3-MA), necroptosis inhibitor (necrostatin-1), and pan-caspase inhibitor (Q-VD-Oph).
As shown in Supplementary Figures S4 and S5, mmu_circRNA_0000309 shRNA impeded the antiapoptotic effects afforded by germacrone in DN mice, while Lip-1 administration almost completely restored the antiapoptotic effects mediated by germacrone (Supplementary Fig. S5A). However, although the other inhibitors, including autophagy inhibitor (3-MA), necroptosis inhibitor (necrostatin-1), and pan-caspase inhibitor (Q-VD-Oph), also had a slight improvement on cell apoptosis, the antiapoptotic effects were moderate and weaker than the ferroptosis inhibitor Lp-1 (Supplementary Figs. S5B and S6A, B). Thus, it is possible that germacrone mainly exerts an antiferroptotic effect on DN progression via upregulating mmu_circRNA_0000309.
Discussion
The rapid oxidative stress stimulated by the HG environment is the main reason for the reduction of podocytes, particularly podocyte apoptosis, which further exacerbates DN progression (11). Triptolide can damage the structure of NADPH oxidase and can weaken ROS reaction to inhibit podocyte apoptosis (27). Spironolactone, as an aldosterone receptor antagonist, brings into play in impeding NADPH oxidase and activating superoxide dismutase to decline podocyte apoptosis (51).
Some recent studies show that antioxidant vitamins are effective in DN treatment by stopping podocyte apoptosis (23, 43). Therefore, reducing podocyte apoptosis is a promising therapeutic strategy for DN. In the present study, we aimed to investigate the role of mmu_circRNA_0000309 on podocyte damage in DN mice under diabetic conditions. Our finding demonstrates that mmu_circRNA_0000309 may display an ancillary function during germacrone-mediated improvement of DN through activating miR-188-3p/GPX4-regulated ferroptosis.
Germacrone participates in anti-inflammatory, antioxidative, and antiapoptotic regulatory mechanisms (64). It is discovered that germacrone treatment evidently suppresses ear inflammation, leading to the decrease in cell apoptosis (35). In the present study, germacrone blatantly improved the abnormal renal functions in DN mice. Besides, germacrone was found to predominantly work in podocytes in the renal corpuscles, which could positively change the expression of podocyte foot process markers, nephrin and podocin, promoting podocyte capability. These results indicated that germacrone also exerted the antiapoptotic effect in podocytes of DN disease.
As mentioned, mmu_circRNA_0080425 upregulation enhances the DN process, and promotes cell proliferation and fibrosis of mesangial cells (31), mmu_circ_0037128 and miR-17-3p also competitively controlled cell proliferation and fibrosis in DN (53). In addition, massive studies have found that circRNAs have myriad miRNA binding sites, which can made them serve as miRNA sponges. The circRNA-miRNA axis can dwindle the obstructive effect of miRNA on target genes and plays a central role in DN (34). The data indicate that circRNAs are a key determinant in controlling cell survival during the DN process.
Based on RNA-sequence analysis, we unanticipatedly outcropped that mmu_circRNA_0000309 was lowly expressed in DN mice, which was notably rescued in germacrone-treated DN mice. We reasonably speculated that germacrone might reduce podocyte apoptosis via an axis centered on mmu_circRNA_0000309.
Podocytes are mature differentiated cells in the glomerulus. The glomerular filtration membrane will be adversely affected in apoptotic or damaged podocytes, thereby aggravating DN progression. Accumulating evidence indicates that abundant signals facilitate podocyte apoptosis, such as p38-MAPK, Smad, Notch/Jag/ICN1 signals can speed up the process of DN (9, 13, 49). Many noncoding RNAs, especially miRNAs are involved in the programmed death of podocytes in DN (54).
As the sponge of miRNAs, circRNAs work with miRNA to coregulate podocyte apoptosis and injury through the antioxidant and anti-inflammatory pathways in DN (18, 62). In the current study, it was found that germacrone would lose its resistance to HG-induced DN symptoms and the elevated podocyte apoptosis in the absence of mmu_circRNA_0000309. The results further proved that mmu_circRNA_0000309 was a switch gene implicated in germacrone-treated DN.
Mitochondrial DNA injury caused by hyperglycemia will accelerate podocyte apoptosis, which is a prevalent mechanism in DN. Podocyte mitochondrial damage is usually coincided with oxidative phosphorylation and COX dysfunction, which will change the expression of inflammatory factors in podocytes (19). It is found that mmu_circRNA_FoxO3 knockout droops glutamate-induced oxidative disability in HT22 cells via the mitochondrial apoptotic pathway (29).
The signaling axis mmu_circRNA-PRKCB/miR-339-5p/p66Shc controls the mitochondrial ROS production by activating NADPH oxidase and the antioxidant enzyme (15). Our research found that the mitochondria in DN mice swelled to produce vacuoles, and the mitochondrial membranes were ruptured. Germacrone treatment could solve the mitochondria dysfunction caused by the aberrant DN status. However, mmu_circRNA_0000309 depletion also abolished the therapeutical effect of germacrone. Potentially, mmu_circRNA_0000309 was involved in mitochondrial function regulation, thereby abrogating the antiapoptotic role of germacrone.
Ferroptosis-like chemistry reaction causes GSH reduction and inactivation of phospholipid peroxidase and GPX4, which can produce iron-dependent ROS to damage cells (61). In renal tubular cells, iron metabolism and lipid peroxidation collection are considered to be the foremost mediators for ferroptosis. Mitochondrial dysfunction brings about iron accumulation in the oxidative respiratory chain and triggers ferroptosis, resulting in cell apoptosis (57).
Our detection discovered that mmu_circRNA_0000309 deletion stopped germacrone from working, leading to a sharp increase of ferroptosis-promoting proteins. These results indicated that germacrone may change the mitochondrial pathological state through mmu_circRNA_0000309, thereby affecting ferroptosis to regulate podocyte apoptosis. Iron-mediated cell injury or ferroptosis is a central player in the pathogenesis of kidney injury. The kidney has an important role in preventing iron loss from the body by means of reabsorption and affecting inflammation. Possibly, germacrone-mediated elevation of mmu_circRNA_0000309 changed the reabsorption function of kidney or inflammatory microenvironment, thereby promoting iron loss from the kidney and inhibiting iron-dependent ferroptosis.
hsa_circ_0109291 sponges miR-188-3p to promote the cisplatin resistance of oral squamous cell carcinoma via adjusting ABCB1 (16); hsa_circRNA_0048179/miR-188-3p markedly upregulates GPX4 and attenuates lipid accumulation in HepG2 cells (60). Combined with the luciferase reporter gene analysis, mmu_circRNA_0000309 sponged miR-188-3p, thereby declining the expression of miR-188-3p. Simultaneously, GPX4 was proven to be positively correlated with mmu_circRNA_0000309. Of note, miR-188-3p could also bind to 3′-untranslated region (UTR) of GPX4 and negatively regulated its expression. Thus, we speculated that mmu_circRNA_0000309 competitively sponged miR-188-3p, and subsequently blocked its inhibitory role on GPX4 expression. miR-188-3p plays a critical role in alleviating cell apoptosis via inhibiting the downstream target genes (17).
Analogously, our data powerfully revealed that the mmu_circRNA_0000309/miR-188-3p/GPX4 axis acted as a core role in podocyte apoptosis by regulating ferroptosis-dependent mitochondrial activity. However, more research is badly needed to investigate whether ferroptosis-mediated mitochondrial injury depends on GPX4. In addition, the mmu_circRNA_0000309/miR-188-3p/GPX4 signaling axis also should be explored using the in vivo DN model.
In the present study, there were still some problems to be solved. First, there were two circRNAs derived from the mouse vascular endothelial zinc finger 1 (Vezf1) gene, including mmu_circRNA_0000309 and mmu_circRNA_0003531. Whether mmu_circRNA_0003531 also plays the similar effects as mmu_circRNA_0000309 remains to be explored in the future. Second, the mmu_circ_0000309 and hsa_circ_0000309 are totally two different genes.
For application to human DN, we found that there were eight hsa_circRNAs that located in the host gene VEZF1, including hsa_circ_0044749, hsa_circ_0044750, hsa_circ_0044751, hsa_circ_0044752, hsa_circ_0044753, hsa_circ_0044754, hsa_circ_0044755, and hsa_circ_0107190. These candidate hsa_circRNAs should be verified using the biopsy samples or kidney tissues of DN patients to screen one or more hsa_circRNAs that play a similar role in human DN as the role of mmu_circRNA_0000309 in a DN mouse model. These should also be verified in the future experiments.
Conclusion
In summary, mmu_circRNA_0000309 silence mediates drug resistance to germacrone in DN mice. Mechanistically, mmu_circRNA_0000309 sponges miR-188-3p, which subsequently upregulates GPX4 expression, thereby inactivating ferroptosis-depended mitochondrial function disorder and podocyte apoptosis.
Materials and Methods (Electronic Laboratory Notebook Was Not Used)
Animal treatments
C57BL/6J mice were purchased from Three Gorges University (Yichang, China), and C57BL/KsJ and male db/db mice were from Changzhou Cavins Laboratory Animal Co. Ltd. (Changzhou, China). All experiments were approved by the Animal Ethics Committee of Zhejiang Provincial People's Hospital, and performed according to specific institutional and national guidelines. The mice were divided into three groups: control C57BL/6J mice, db/db mice, and germacrone-treated db/db mice (db/db+Ger) (n = 10/each group). The db/db+Ger mice received germacrone treatment at a dosage of 10 mg/kg/day, while C57BL/6J mice and db/db mice had been given the same volumes of 0.9% saline simultaneously.
For the shRNA injection, each mouse was injected with 10 nmol cholesterol-modified mmu_circRNA_0000309 shRNA (GenePharma, Shanghai) into the tail vein every 3 days. For the injection of Lip-1 (HY-12726; MedChemExpress), mice were challenged by intraperitoneal injection of 10 mg/kg Lip-1. In db/db mice appeared the symptoms of mesangial hyperplasia, glomerulus mesangial matrix expansion, podocyte loss, basement membrane thickening and other nephropathies at 12 weeks. Thirteen weeks after modeling, mice were used for subsequent experiments.
Measurement of biochemical parameters
Fasting blood glucose was monitored with a fully automatic biochemical analyzer 7020 (Hitachi, Japan) at 2-week intervals by using one drop of tail blood. At the end of the experiments, mice were placed in metabolic cages, and 24-h urine was collected for determination of proteinuria. The urine samples were centrifugated at 300 g for 10 min. After removing the precipitates, samples were stored at 80°C until analyzed. The BUN, Scr, Ucr, and BUA were measured with the corresponding ELISA kits.
Glutathione peroxidase activity
NADPH has a characteristic absorption peak at 340 nm, and the activity of GSH was calculated by NADPH rate (38). NADPH was determined by the Nano-100 Micro Spectrophotometer (Aosheng Instrument, China) at 340 nm for fixed time intervals (15 s), and the result was expressed as nanomole of NADPH oxidized to NADP per minute per milligram protein.
Glutathione peroxidase activity and iron contents
Cells or 10% tissues were homogenized and centrifuged to take the supernatants, and the GPX activity and iron level were measured with the standard protocols of the Glutathione Peroxidase Cellular Activity Assay Kit (CGP-1; Sigma) and iron content test kit (E-BC-K139; Elabscience, Wuhan, China), respectively.
Cell culture, treatments, and transfection
Primary mouse podocyte MPC5 purchased from the Institute of Basic Medicine, Chinese Academy of Medical Sciences, was cultured in 10% fetal bovine serum (FBS) RPMI-1640 (SH30809.01B; Hyclone) medium containing 20 U/mL recombinant human IFN-γ (TL-105; T&L Biotechnology, Beijing), supplemented with type I collagen in flasks, 100 U/mL penicillin (DY14011; Hyclone) for proliferation (1:3) at 37°C. To promote the differentiation of podocytes, cells were cultured in 5% FBS RPMI-1640 medium without IFN-γ at 37°C for 10–14 days (change the medium every 2 days). MPC5 cells were treated with 25.5 mM mannitol to stimulate cell differentiation. Glucose treatment, 25.5 mM, for 24 h (started at 0 h) served as the HG treatment group.
For the stable Gpx4 overexpression podocyte cell line, GPX4 CDS fragment was subcloned into the pEGFP-C1 plasmid. For cell transfection, pEGFP-C1 and recombinant pEGFP-C1-Gpx4 plasmids were transfected into cells using the TurboFect transfection reagent (R0531; Thermo Fisher Scientific). Then, cells were screened by G418 for 10 days, and the stable Gpx4 overexpression podocyte cell line was confirmed by Western blot.
For shRNA transfection, mmu_circRNA_0000309 shRNAs (shRNA-1: CAGCCGAACTTCGTTGGTTTCAACT; shRNA-2: CCGAACTTCGTTGGTTTCAACTATT; shRNA-3: TCAAGAAACCCAGTAAGCCTGTCAA; NC shRNA: UUCUCCGAACGUGUCACGUTT) were transfected into MPC5 cells for 48 h. Subsequently, cells were harvested for RNA isolation, and real-time PCR was used to determine the expression of mmu_circRNA_0000309 in MPC5 cells.
Hematoxylin/eosin staining
The kidney tissue samples were fixed in 4% PFA (30525-89-4; Sinopharm, Shanghai, China), dehydrated, embedded in paraffin, and cut into 5-μm-thickness sections. After dewaxing with xylene and gradient alcohol, the sections were dyed using hematoxylin for 5 min and washed with distilled water. Then the slices were counterstained using 1% hydrochloric acid (GB622-89, Xinyang Chemical Reagent, Henan, China) ethanol for 10–60 s and washed with distilled water. Subsequently, the sections were dyed with 0.5% eosin (71014544; Sinopharm) for 5 min and washed with distilled water for 10 s. After mounting, the results were observed under a microscope.
PAS staining
Five-micrometer-thickness sections were placed in 10 g/L periodic acid (10450-60-9; Nanjing Reagent, Nanjing, China) solution for 20 min, and then washed with ddH2O for 5 min. Then the slices were incubated with Schiff (DG0005; Leagene, Beijing, China) dye solution for 1 h, rinsed with tap water for 10 min, stained with hematoxylin (CTS-1099; MXB, Fuzhou, China) for 5 min, and rinsed again with tap water for 10 min. The results were observed under a microscope.
Immunofluorescence staining
The glass slides were placed in 24-well dishes and the same density cells were added. After 48 h, the glass slides were washed with phosphate-buffered saline (PBS) thrice for 5 min. Then the slices were fixed with 4% paraformaldehyde for 15 min, washed with PBS, and permeabilized with 0.2% Triton X-100 (ST795; Beyotime Biotechnology, Shanghai, China) at room temperature for 10 min. After washing with PBS for 5 min, the slices were blocked in 10% bovine serum albumin at room temperature for 1 h.
The primary antibodies against podocin (ab50339; Abcam) and nephrin (ABIN2177857; Antibodies-Online, Germany) were incubated with the slices at 4°C overnight. Followed by staining with the CyTM3-conjugated secondary antibody goat anti-rabbit IgG (BA1032; Boster Biological Technology, Wuhan, China), the nuclei were visualized by DAPI (C1002; Beyotime Biotechnology) staining. Images were taken using a fluorescence microscope (BX53; Olympus, Japan).
Tissue sections were deparaffinized and stained according to the standard protocol of the TUNEL Apoptosis Detection Kit (12156792910; Roche Applied Science, Switzerland). Subsequently, the sections were conducted with immunofluorescence staining to detect apoptotic cells.
RNA-seq
circRNA microarray analysis was contracted by NUMBUI Corporation (Shenzhen, China). In brief, the TRIzol reagent (15596-026; Ambion) was used to extract total RNA from mice kidney tissue. All total RNA samples were amplified, and then microarray hybridization was performed on the basis of Arraystar Mouse circRNA Array (6x7K; Arraystar, Inc., Rockville, MD) and scanned by the Agilent G2505C Scanner. circRNA chip data combined with miRNA target gene prediction software were made full use of to find cotargeted miRNAs of mRNA and circRNA, and constructed a ceRNA network.
miRNA screen
miRNA screening made full use of circBase (
RNA isolation and real-time PCR
Total RNA was extracted from tissues and cells based on the standard manufacturer's protocol with the TRIzol reagent (15596-026; Ambion). RNA was reversed transcribed into cDNA. The primers are shown below: U6-Forward, 5′-CGCTTCGGCAGCACATATAC-3; U6-Reverse, 5′-AAATATGGAACGCTTCA-CGA-3′; mmu-miR-188-3p-Forward, 5′-TGCGCCTCC-CACATGCAGGGT-3′, Loop: 5′-GTCGTAGGATAGTGCAGGCAAACGAGGTATTACT-3′, Reverse, CCAGTGCAGGGTCCGAGGTATT; mmu_circ_0000309-Forward, 5′-ACGAATGGGATGAACAGG-3′, Reverse, 5′-ACAAGCACGATGAAATGG-3′; Mus GPX4-Forward, 5′-CAAAGTCCTAGGAAACGCCC-3′, Reverse, 5′-CCTTGGCTGAGAA TTCGTGC-3′; and Mus β-actin Forward, 5′-CACGATGGAGGGGCCGGACTCATC-3′, Reverse, 5′-TAAAGACCTCTATGCCAACACAGT-3′.
Other primers of circRNAs are shown in Table 1. The primers were designed by using Primer Premier 5 software (Premier Biosoft International, Palo Alto).
Primer Sequences of Quantitative Reverse Transcription-Polymerase Chain Reaction
The reaction condition was 2 min of 50°C, followed by 40 cycles of 95°C/10 min, 95°C/30 s, and 60°C/30 s. Then the expressions of these genes were determined by using SYBR Green Master Mix (Q111-02; VAZYM E) and standard real-time fluorescent quantitative PCR system protocol for reaction.
Flow cytometry
Podocytes were digested using trypsin (T4799; Sigma), resuspended in 1 × binding buffer, and collected by centrifugation at 1000 g for 3 min. The cell suspension was treated with the Annexin V-APC/7-AAD Double Stain Apoptosis Detection Kit (KGA1026; KeyGen Bio, Nanjing, China) in darkness for 15 min at 37°C. The apoptotic cells were then detected by flow cytometer (CytoFLEX; Beckman).
For the inhibition assays, MPC5 cells were pretreated with antiferroptotic reagents Lip-1 (0.5 μM, HY-12726; MedChemExpress), autophagy inhibitor (3-MA, 5 mM, M9281; Sigma), necroptosis inhibitor (necrostatin-1, 30 mM, HY-15760; MedChemExpress), and pan-caspase inhibitor (Q-VD-Oph, 0.05 μM, HY-12305; MedChemExpress) for 24 h. Subsequently, cell apoptosis was measured by flow cytometer. ROS activity within the cells was detected using the DCFH-DA ROS Assay kit (S0033; Beyotime Biotechnology).
The results were also measured by flow cytometry. The lipid peroxidation accumulation (lipid ROS) was assessed by flow cytometry using the C11-BODIPY probe (D3861; Invitrogen). In brief, the cells were rinsed twice with PBS following incubation. Adhered cells were stained with C11-BODIPY (2.5 μM) solution in PBS in darkness for 30 min at 37°C. Cells were then collected with 0.05% trypsin solution, suspended in fresh complete medium, and immediately analyzed with a flow cytometer. MMP detection was also evaluated by flow cytometry.
Briefly, MPC5 cells were digested with 0.25% trypsin without EDTA, and the cell pellets were collected and centrifugated at 1000 rpm for 5 min. After removing the supernatant, the cells were washed with PBS for three times. One microliter JC-1 diluted in 1000 μL incubation buffer was added in the cell solution, allowing incubation for 15 min. Subsequently, the cells were collected and washed twice with 1 × incubation buffer. Then the MMP content was measured by flow cytometry.
Transmission electron microscopy
Fresh tissues were fixed in an electron microscope fixative (G1102; Servicebio, Wuhan, China) for 2–4 h, washed with 0.1 M phosphate buffer (PB, pH 7.4) for 15 min × 3 times, and fixed in 0.1 M PB containing 1% osmic acid (PBGP18456; Zhongjingkeyi, Beijing, China) for 20 min at room temperature. After washing with 0.1 M PB for 15 min, the slices were dehydrated using conventional alcohol gradient. According to the method of uranyl acetate (YS014569; Amresco)-lead citrate (L885990; Macklin, Shanghai, China), the slices were stained and dried, the images were observed under a transmission electron microscope (HT7700; Hitachi, Japan), and the collected images were analyzed.
Luciferase assay
MPC5 cells were seeded in 48-well plates at 80% confluence. The wild type and the mutant 3′-UTR of miR-188-3p and GPX4 sequences were cloned into the pRL-TK plasmid vector. Then, MPC5 cells were cotransfected with mmu_circRNA_0000309 mimics and control mimics by Lipofectamine 2000 (11668027; Thermo). After 24 h, the transfected MPC5 cells were examined using a Dual-Luciferase Reporter Assay System.
Western blot
Cells or tissue proteins were extracted with the RIPA lysis solution (P0013B; Beyotime Biotechnology) and quantified by BCA (P0010; Beyotime). Antibodies against GAPDH (1:1000, Ab181602; Abcam), GPX4 (1:3000, Ab125066; Abcam), podocin (1:1000, Ab50399; Abcam), nephrin (1:1000, Ab58968; Abcam), FTH1 (1:1000, DF6278; Affinity), COX-2 (1:1000, Ab15191; Abcam), NOX1 (1:2000, DF8684; Affinity), ACSL4 (1:2000, Ab155282; Abcam), PARP (1:2000, 9542; CST), and caspase 3 (1:1000, ab184787; Abcam) were used to incubate with the membrane. After washing with TBST for 10 min × 3 times, the blots were incubated with horseradish peroxidase-conjugated goat anti-rabbit/anti-mouse IgG (1:5000, BA1054, BA1053; Boster Biological Technology) for 1 h and visualized with an ECL Chemiluminescent Detection Kit (P1050; Applygen, Beijing, China).
Statistical analyses
All experiments were expressed as mean ± SEM and performed at least three independent times. Differences between the groups were analyzed with Student's t-test and two-way analysis of variance (ANOVA) using GraphPad Prism (version 8.0). *p < 0.05 indicates a significant difference between the different groups. The analyses were performed using SPSS (version 25.0) software.
Footnotes
Authors' Contributions
Q.H. and. Y.W. conceived the idea and designed the experiments. J.J. and Y.W. conducted the experiments. J.J., D.Z., and M.L. analyzed the results. Q.H. and Y.W. wrote the article. Q.H. and Y.W. supervised and supported the study. All authors reviewed the article.
Ethical Approval and Consent to Participate
All experiments were approved by the Animal Ethics Committee of Zhejiang Provincial People's Hospital, and performed according to specific institutional and national guidelines.
Availability of Data and Materials
All data generated or analyzed during this study are included in this published article.
Author Disclosure Statement
No competing financial interests exist.
Funding Information
This work was supported by The Construction of Key Projects by Zhejiang Provincial Ministry (WKJ-ZJ-1915 and WKJ-ZJ-2017), The Zhejiang Province Chinese Medicine Modernization Program (2020ZX001), The General Project of Zhejiang Education Department (Y201942823), The Construction Fund of Medical Key Disciplines of Hangzhou (2020–2024), Zhejiang Provincial Traditional Chinese Medicine Science and Technology Project (2022ZB273), and Clinical and Experimental Research of YSHS Granule.
Supplementary Material
Supplementary Figure S1
Supplementary Figure S2
Supplementary Figure S3
Supplementary Figure S4
Supplementary Figure S5
Supplementary Figure S6
Abbreviations Used
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.