
Abstract
Significance:
The production of antibodies to posttranslationally modified antigens is a hallmark in rheumatoid arthritis (RA). In particular, the presence of citrullination-associated antibodies, targeting both citrullinating enzymes (the peptidylarginine deiminases [PADs]) and citrullinated antigens (anticitrullinated protein antibodies [ACPAs]), has suggested that dysregulated citrullination is relevant for disease pathogenesis. Antibodies to other protein modifications with physicochemical similarities to citrulline, such as carbamylated-lysine and acetylated-lysine, have also gained interest in RA, but their mechanistic relation to ACPAs remains unclear.
Recent Advances:
Recent studies using RA-derived monoclonal antibodies have found that ACPAs are cross-reactive to carbamylated and acetylated peptides, challenging our understanding of the implications of such cross-reactivity.
Critical Issues:
Analogous to the classic antibody response to chemically modified proteins, we examine the possibility that antibodies to modified proteins in RA are more likely to resemble antihapten antibodies rather than autoantibodies. This potential shift in the autoantibody paradigm in RA offers the opportunity to explore new mechanisms involved in the origin and cross-reactivity of pathogenic antibodies in RA. In contrast to citrullination, carbamylation is a chemical modification associated with oxidative stress, it is highly immunogenic, and is considered in the group of posttranslational modification-derived products. We discuss the possibility that carbamylated proteins are antigenic drivers of cross-reacting antihapten antibodies that further create the ACPA response, and that ACPAs may direct the production of antibodies to PAD enzymes.
Future Directions:
Understanding the complexity of autoantibodies in RA is critical to develop tools to clearly define their origin, identify drivers of disease propagation, and develop novel therapeutics. Antioxid. Redox Signal. 36, 389–409.
Introduction
After the description of rheumatoid factor in 1940 (194), multiple autoantibody specificities of uncertain significance have been detected in patients with rheumatoid arthritis (RA) (1, 4, 11, 12, 15, 17, 18, 25, 30, 50, 51, 78, 115, 120, 122, 123, 127, 144, 146, 156, 158, 171, 180, 181, 199, 207). It was the identification of antibodies to citrullinated antigens by Schellekens et al. in 1998 (147), however, which offered for the first time both a useful biomarker for diagnosis and a plausible mechanism responsible for the breach of tolerance to autoantigens in RA (40). The hypothesis that citrullination is a major driver of autoantigen production in RA also provided a feasible explanation to elucidate how previous findings, some of them ambiguous, tailored into a central model of RA pathogenesis. These include the following. (i) Citrullination unified the antigenic specificity of three seemingly unrelated antibody systems strongly associated with RA: the antiperinuclear factor (APF), antikeratin antibodies (AKA), and anti-Sa antibodies (50, 127, 207). (ii) It explained why a broad range of proteins with no obvious relationship, later found to be targets of citrullination, were initially identified as autoantigens in RA (e.g., calpastatin, enolase, collagen, histones, heterogeneous nuclear ribonucleoproteins, vimentin, actin, and others) (4, 11, 12, 15, 30, 51, 120, 144, 158, 171, 181). (iii) Since neutrophils are a major source of citrullination (45), it elucidated the early finding of antibodies to neutrophil antigens in RA (25, 123). (iv) The discovery of anticitrullinated protein antibodies (ACPAs) enlightened the striking association between RA and HLA-DRB1 shared epitope (SE) alleles (87), suggesting that this association is driven through the presentation of citrullinated antigens by molecules encoded by HLA-DRB1-SE alleles (145, 173). (v) Bacterial-induced citrullination provided a mechanistic model to explain the centennial association between periodontitis and RA (73).
The study of autoantigen citrullination also brought new interest about whether other posttranslational modifications (PTMs) are targets of antibodies in RA (39). Antibodies to carbamylated-lysine (i.e., homocitrulline) and acetylated-lysine proteins have gained attention in this regard, mainly because they may offer an additional tool for diagnosis particularly in ACPA-negative RA (91, 153). However, whether and how these PTMs may mechanistically relate to citrullination in RA pathogenesis remains unclear. While initial studies suggested that ACPAs and anticarbamylated protein (Anti-CarP) antibodies were independent antibody systems in RA, there was also evidence that a large proportion of these antibodies were cross-reactive (134, 153). The recent characterization of RA-derived monoclonal ACPAs has confirmed their cross-reactivity with antigens containing homocitrulline or acetylated lysine (KAc) residues (95, 142), suggesting that these antibodies are promiscuous to modified amino acid residues that may share some physicochemical properties with citrulline.
The finding that ACPAs are cross-reactive with other PTMs has important implications for RA and raises many new questions. (i) It may reveal that antibodies to modified proteins in RA relate to a single family of antibodies driven by PTMs with structural similarities (95). (ii) It may challenge existing theories regarding the main role of citrullination as the driver of RA, opening the possibility that other PTMs, such as carbamylation and/or acetylation, may play a primary role in the initial breach of tolerance to self-antigens in this disease. (iii) It may expose that anti-CarP and/or antiacetylated protein antibodies (AAPAs) are indeed cross-reacting ACPA subsets (134), suggesting that carbamylation and/or acetylation are distractors with no relevance for RA pathogenesis.
Interestingly, while PTMs have long been recognized as a potential mechanism to breach tolerance to self-antigens in autoimmune diseases, it is noteworthy that in most cases the antibody response is specific to the antigen containing the modification (54, 55). In the antibody response to modified proteins linked to RA, however, the primary target is the modification but not the antigen, generating cross-reacting antibodies to a broad range of unrelated proteins containing structural epitopes that mimic the primary modified immunogen (142). Understanding the antibody response to PTMs in RA may therefore require a different model to those proposed for other autoimmune diseases (54, 55).
Here, we review some general features about the PTMs targeted by antibodies in RA, including both their strengths and shortcomings to the RA paradigm, and discuss the potential basis and mechanistic significance of their cross-reactivity with ACPAs. In particular, our interest is on understanding why and how a small modification in two polar amino acid residues (i.e., arginine and lysine) is recognized by antibodies in patients with RA. Analogous to the normal antibody response to chemically modified amino acid residues (108, 200), we examine the possibility that antibodies to modified proteins in RA are more likely to resemble antihapten antibodies rather than autoantibodies. Moreover, we discuss the possibility that carbamylated proteins may initiate the immune response to modified proteins in RA. Although malondialdehyde-acetaldehyde (MAA) adducts are also PTMs targeted in RA, anti-MAA antibodies are not included in this review because these antibodies are not specific for RA (5, 138, 187), MAA adducts have no structural similarity with citrulline residues (39), and the shape and charge of MAA adducts are unlikely to fit into the ACPA paratope (67). Thus, while antimodified protein antibodies (AMPAs) in RA may include specificities to any protein modification regardless of their mechanistic association (46, 176), this review only focuses on the analysis of the AMPA subset targeting antigens containing citrulline-like residues.
Overview of PTMs Recognized by Antibodies in RA
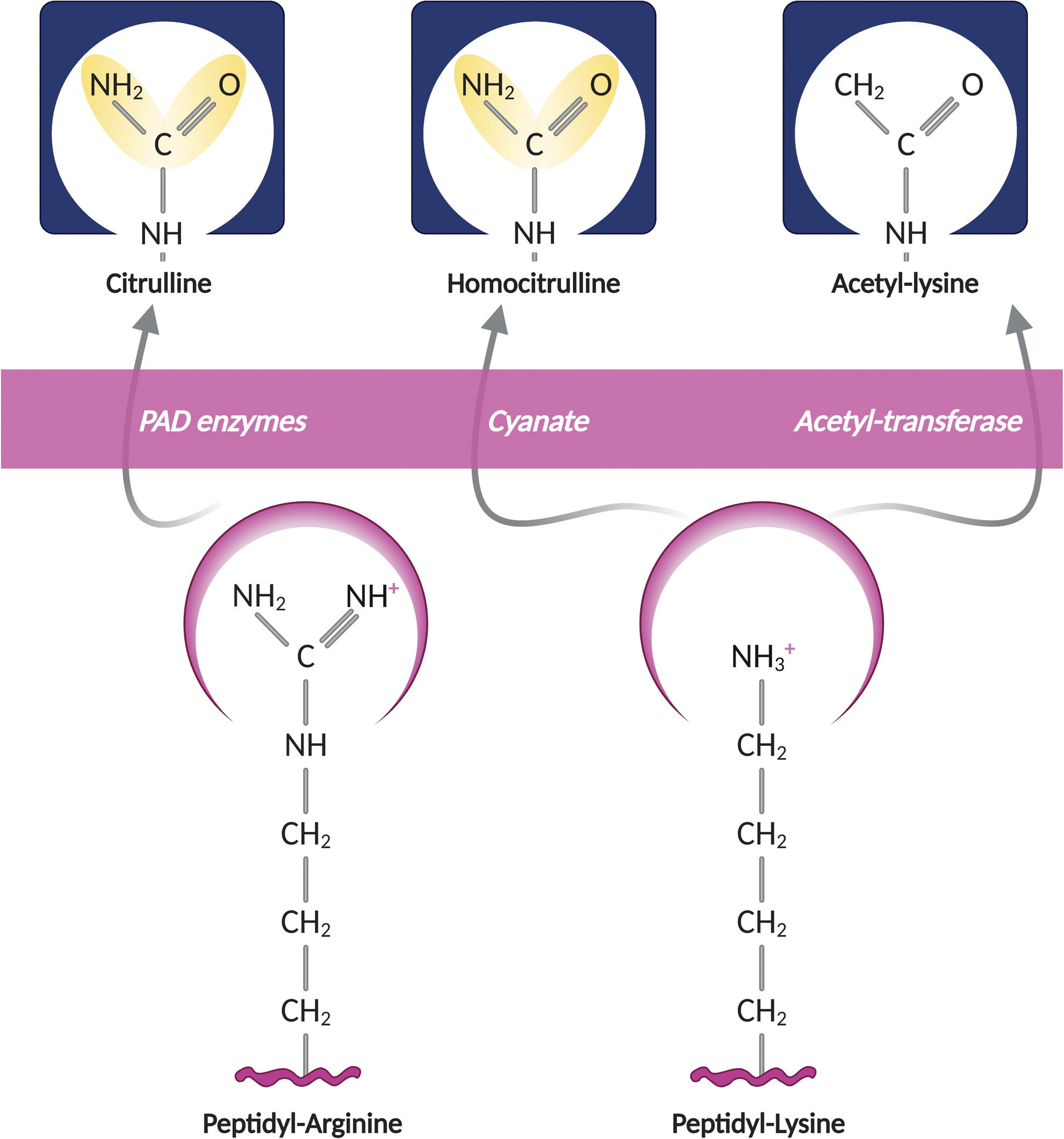
Arginine and lysine are amino acids with a very polar side chain, which makes them highly hydrophilic. They are positively charged at neutral pH and their side chains are the longest in the set of the 20 amino acids used for protein synthesis. Arginine and lysine residues can undergo multiple PTMs, but for the purpose of this review, we only focus on citrullination, carbamylation, and acetylation. These modifications have in common that they neutralize the positive charge of arginine and lysine, creating modified residues with similar shape, charge, and hydrophobicity (Fig. 1), which may render proteins and small peptides the target of a common subset of antibodies in RA. Interestingly, citrullination and carbamylation are generated under opposite redox conditions, suggesting that these modifications are likely induced by distinct mechanisms in RA.
Citrullination and the need of a reducing environment for PAD activation
Citrullination or deimination is the enzymatic conversion of arginine residues to citrulline, which is mediated by the calcium-dependent peptidylarginine deiminase (PAD) enzymes (66). PAD enzymes hydrolyze guanidinium side chains in arginine residues to yield citrulline and ammonia, and this process appears to be irreversible (Fig. 2). The PADs include five members (PAD1–4 and 6) (26, 75, 89, 93, 125, 208), but only PADs 1–4 have demonstrable citrullinating activity (132). PADs are highly conserved and share 50%–55% sequence identity (7), but exhibit distinct substrate preferences and tissue expression (193, 203).
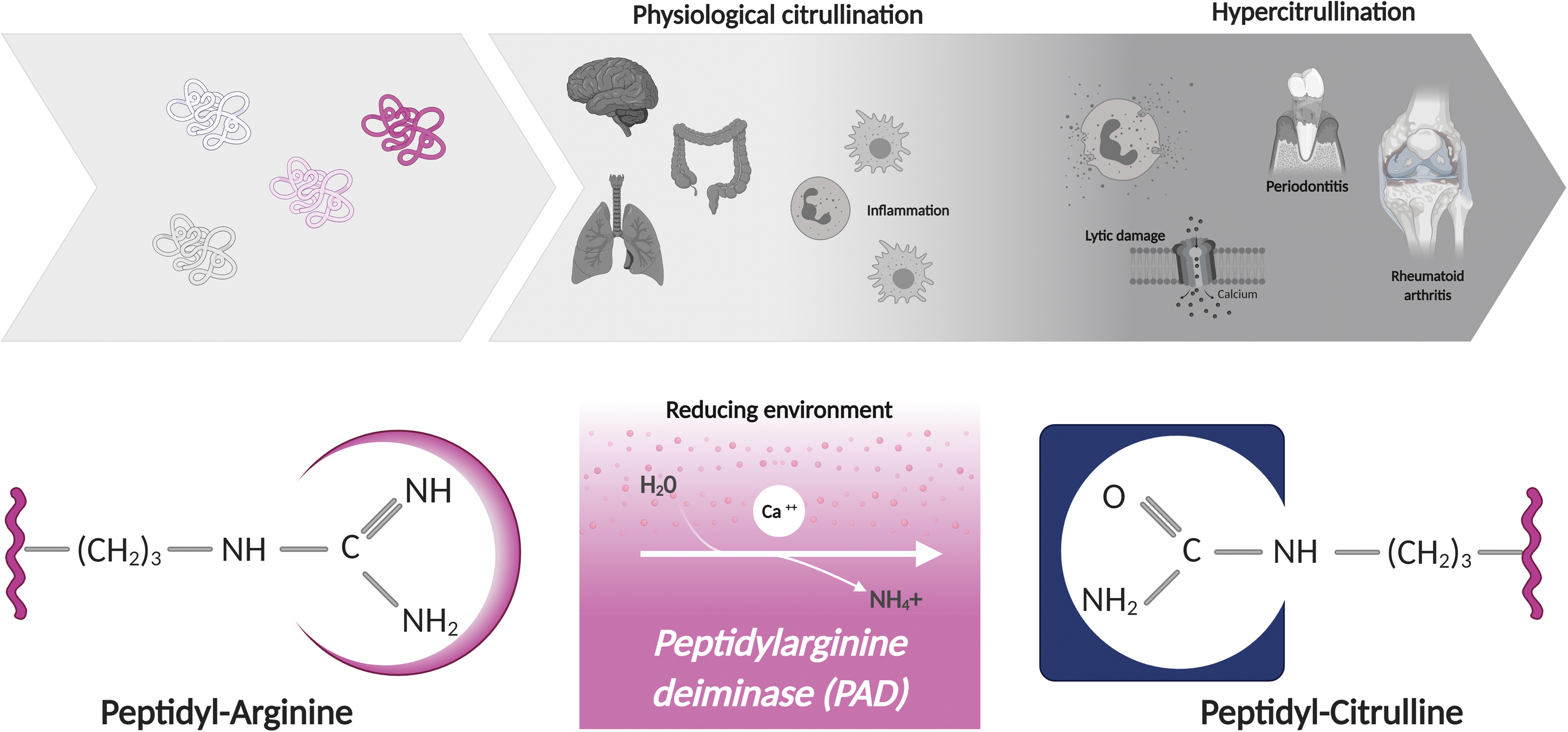
Citrullination is a normal process and more than 200 citrullinated proteins have been identified across multiple tissues in humans (109), which together constitute the human citrullinome. The effect of citrullination on the physiologic function of most of these proteins is unknown. Calcium and redox conditions are important regulators of PAD activity (40). A prominent increase in intracellular calcium as a result of lytic damage is a potent activator of PAD enzymes in cells, which has been associated with the abnormal accumulation of citrullinated proteins (termed hypercitrullination) in target tissues of illnesses such as periodontitis and RA (101, 140). In addition, since PAD enzymes are abundant in myeloid cells, such as neutrophils and monocytes (45, 191), citrullination has been found to increase during inflammation. However, mechanisms that tune PAD activation during physiologic conditions are not well defined.
Interestingly, neutrophils use different redox conditions to counterbalance citrullination and the production of reactive oxygen species (ROS) (Fig. 3), which both have deleterious effects to each other (36, 211). PAD activity is favored by a reducing environment (37, 66), which is required to maintain the active site free thiol cysteine (i.e., C645 in PAD4) necessary for catalysis (7), while ROS inhibit PAD activity likely through oxidation of the catalytic cysteine (36). Conversely, PAD4-mediated citrullination blocks ROS production by targeting components of the phagocyte NADPH oxidase (NOX2) (211). The balance between citrullination and ROS generation by NOX2 in neutrophils is maintained by PAD4, which is physically associated with the cytosolic subunits of NOX2, p47phox and p67phox. This association is essential for the assembly of active NADPH oxidase and ROS production, which is likely amplified by maintaining PAD4 inactive within an oxidizing environment. Upon conditions that activate PAD4, however, citrullination of p47phox and p67phox prevents the assembly of an active NADPH oxidase complex and the production of ROS (211) (Fig. 3).
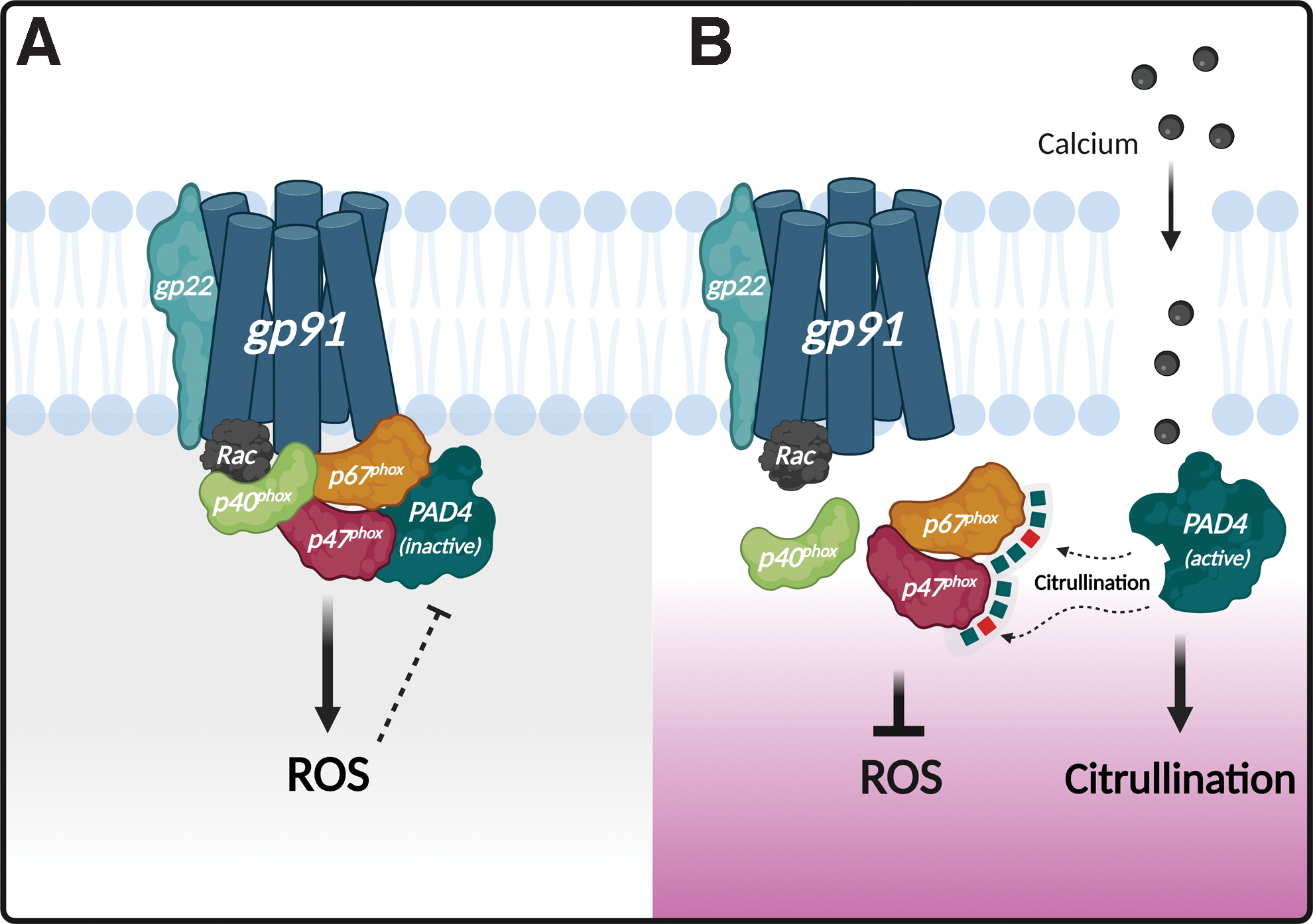
Since the interaction of PAD4 with NOX2 is required for efficient production of ROS (211), this finding explains that PAD4-deficient mice are defective in the production of neutrophil extracellular traps (NETs) (110), which is a process dependent on NOX2 activation (65). Moreover, the finding that PAD4 activation blocks ROS production elucidates why citrullination is deleterious for the killing of phagocytosed bacteria by neutrophils (211), and why citrullination is restricted under conditions of neutrophil activation that requires efficient ROS production, such as the production of NETs (65, 102). Paradoxically, since ROS generation by NOX2 is required for NET formation (65), it was initially thought that ROS production was a trigger of PAD activation (137). This concept was further supported by the finding that PAD inhibitors block the formation of NETs by phorbol 12-myristate 13-acetate (97 –99), which is dependent on ROS generation by NOX2 (65). These findings, however, have not been reproduced by others (94, 102), supporting that NETs induced by activation of NOX2 are independent of citrullination. In contrast to intracellular ROS, which mediate an efficient reduction in PAD activity in cells (36), exogenous hydrogen peroxide can activate citrullination in neutrophils (of histone H3 at least) (126). This finding is likely explained because exogenous hydrogen peroxide induces calcium influx in neutrophils (68), which may generate some PAD activation before its oxidation.
Although oxidative stress is considered an important mechanism of joint damage in RA (72), it is noteworthy that citrullinated proteins accumulate in the RA joints (60, 140, 172, 179, 183, 196). Proteomic analyses of the cellular and soluble components in RA synovial fluid have identified more than 100 citrullinated proteins (60, 140, 172, 179, 183, 196), which include both intra- and extracellular substrates and together comprise the RA citrullinome (60). The finding of hypercitrullinated cells and their citrullinated contents released in RA synovial fluid strongly suggests that intracellular citrullination in RA is driven by NOX-independent mechanisms of cell death, which can efficiently hyperactivate PAD enzymes in cells (102, 140). Regarding citrullination of extracellular substrates, although RA synovial fluid has increased levels of antioxidants (e.g., glutathione reductase and thioredoxin) (143, 161, 206), neither endogenous nor recombinant PADs have catalytic activity in RA synovial fluid unless reducing agents are exogenously added (37, 38). This finding supports that extracellular PADs are inactive by oxidation in RA synovial fluid. However, since PADs released from activated and dying neutrophils are transiently active (37), extracellular citrullination may result from residual PAD activity, which may be enhanced by the presence of anti-PAD4 activating antibodies (43).
Carbamylation is associated with oxidative stress
In contrast to citrullination, carbamylation is a nonenzymatic protein modification generated in conditions associated with oxidative stress (22, 47, 48, 74, 198), and is considered in the group of PTM-derived products (PTMDPs) (69, 70). PTMDPs also include other spontaneous nonenzymatic protein modifications such as oxidation, glycation, glycoxidation, lipoxidation, and carbonylation, which exert deleterious effects in the function of proteins (69, 70). PTMDPs are irreversible, accumulate during life, and are characterized by the spontaneous binding of small metabolites to free reactive groups of proteins, which has a critical influence on protein structure and biological functions, especially in the context of aging, physiological stress, and inflammation (69, 70, 74).
Carbamylation is a chemical modification that results from the addition of a carbamoyl moiety to free functional groups of proteins, peptides, and free amino acids (162 –164). This process mainly results from the reaction of isocyanic acid with a primary amine or a free sulfhydryl group (162 –164), although carbamoyl-phosphate may also act as a carbamylating agent (133) (Fig. 4). Two terms are used to describe this reaction: carbamoylation and carbamylation. Although carbamoylation is the correct term (47), carbamylation, which indeed refers to the addition of CO2 to protein amino groups (111), is the most commonly used. Carbamylation may occur on the α-amino end of proteins or amino acids, or on the ɛ-amino group of lysine residues (generating homocitrulline residues) (47, 162 –164). In addition, carbamylation can also happen on side chains of arginine and cysteine residues (29, 170).
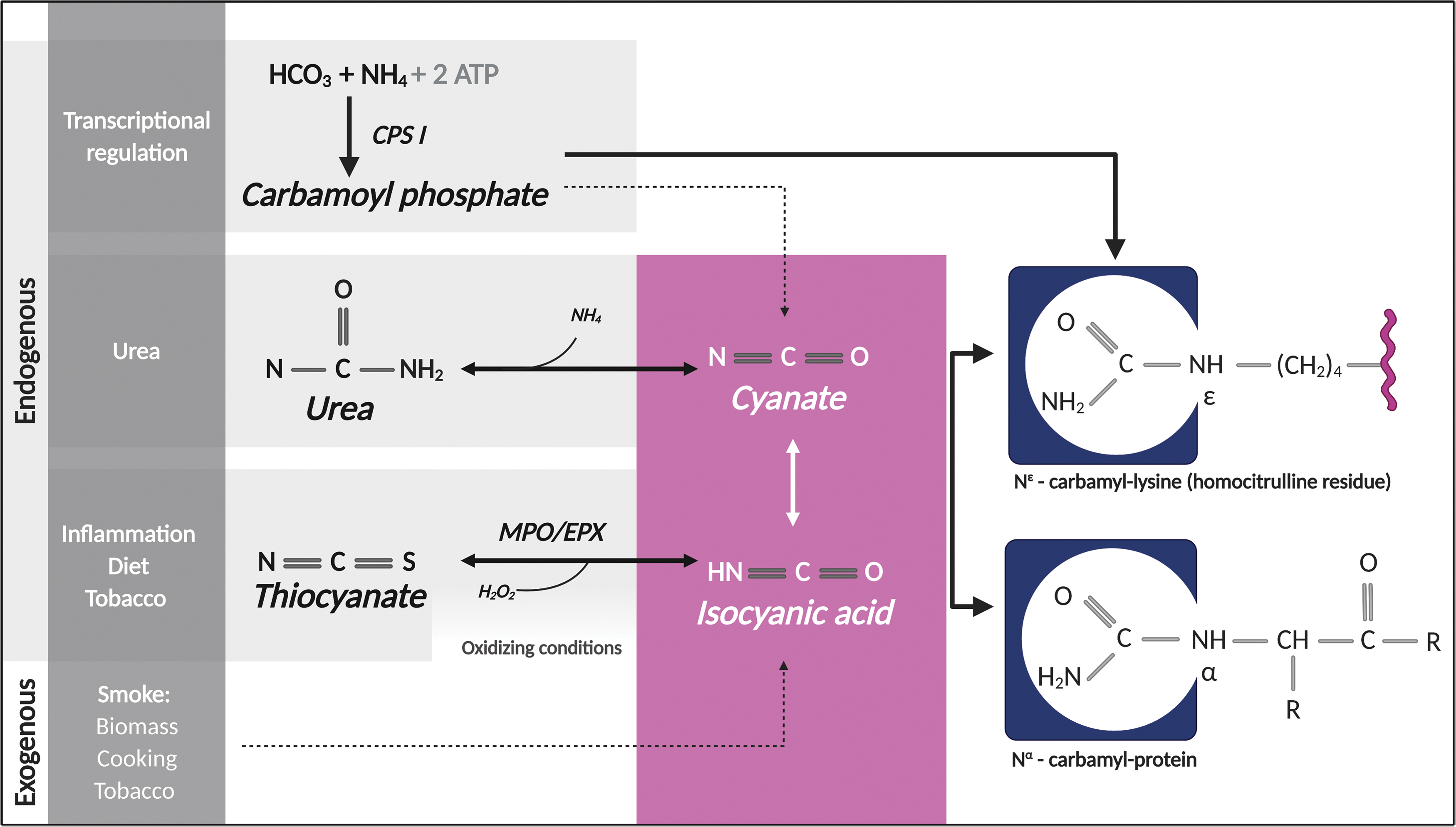
Smoke from biomass burning, biofuel usage, cooking, and tobacco smoking is the source of potential exposure to exogenous isocyanic acid in humans (136). Endogenous isocyanic acid is generated by slow breakdown of urea (this pathway seems to be the major source of isocyanic acid in blood) and through oxidation of thiocyanate (derived from food or smoking) by myeloperoxidase (MPO) or eosinophil peroxidase (EPX) (8, 52, 77, 128, 197, 198). Urea dissociates spontaneously into ammonia and cyanate (cyanate is in equilibrium with its tautomer isocyanic acid) and therefore, carbamylated proteins accumulate in conditions that enhance cyanate levels, such as uremia (52, 53, 59). Leukocyte-derived MPO and EPX can serve as important sources for protein carbamylation at sites of inflammation (8, 197, 198) (Fig. 4). MPO and EPX generate isocyanic acid using hydrogen peroxide and thiocyanate as substrates (8, 197, 198). In addition, MPO-derived chlorinating species decompose thiocyanate and/or urea forming cyanate (48, 86). In the context of inflammation, the requirement of an oxidizing milieu to drive MPO- and EPX-induced carbamylation importantly contrasts with citrullination, which requires a reducing environment and is blocked by oxidation (36).
Carbamylation mediated by carbamyl phosphate has been proposed to involve the direct reaction of the acyl phosphate group in carbamoyl phosphate with a lysine residue (90), although decomposition of carbamyl phosphate resulting in the production of cyanate could be an alternative mechanism (3) (Fig. 4). Carbamyl phosphate is an essential intermediate of pyrimidine and arginine/urea biosynthesis (152). It is generated from ammonia and bicarbonate by the catalytic activity of carbamoyl phosphate synthase (CPS), which includes three enzymes, CPS I and CPS II found in humans and CPS III in invertebrates and fish (152). While CPS I is a mitochondrial enzyme, studies using primary hepatocytes from mice found that CPS I is also a nuclear protein recruited by the aryl hydrocarbon receptor, a transcriptional regulator of xenobiotic toxicity (90). Production of carbamyl phosphate by nuclear CPS I mediates carbamylation of several nuclear proteins in mouse hepatocytes, including histone H1. Interestingly, this process has been suggested to play a role in transcriptional regulation (90), which may have important paradigm-shifting implications to the pathogenic view of carbamylation.
Depending on the altered molecule (e.g., extracellular matrix proteins, collagen, albumin, erythropoietin, hemoglobin, low-density lipoprotein [LDL], high-density lipoprotein, and others), PTMDPs can have different pathophysiological effects. Carbamylation, in particular, has been associated with the acceleration of cardiovascular disease, immune and hemostatic dysfunction, renal fibrosis, and decrease in erythropoietin activity (47). The finding that carbamylation appears to be a regulated PTM in cells is therefore intriguing (90), suggesting that this modification may be reversible (at least intracellularly) and not always detrimental for protein function. Indeed, a recent study suggested that fibroblasts can degrade carbamylated proteins through the ubiquitin/proteasome system (49). Analysis of carbamylation in some cells and tissues has identified carbamylated proteins in neutrophils, skin, and cartilage from healthy individuals (74, 105, 188), and in kidney from healthy rats (29), suggesting that there is some threshold of tolerance to carbamylated proteins under physiological conditions. The complete extent and identity of carbamylated proteins in healthy and disease tissues (i.e., the human “carbamylome”) are yet to be defined. Interestingly, no differences have been found in the content of carbamylated proteins in the joints of patients with RA, osteoarthritis (OA), and healthy controls (188).
Lysine acetylation
Lysine acetylation is a reversible PTM consisting in the transfer of an acetyl group from acetyl-coenzyme A (acetyl-CoA) to the ɛ-amino group of lysine residues (2). Acetylation can occur nonenzymatically by the direct reaction of acetyl phosphate or acetyl-CoA with lysine residues (195). However, in the majority of the cases, acetylation is regulated enzymatically by a balance between lysine acetyltransferases (KATs) and lysine deacetylases (KDACs) (2) (Fig. 5). Since lysine acetylation is a dynamic process, this PTM is widely used in nearly all intracellular compartments to regulate the function and interactions of proteins (195). Nuclear acetylation regulates gene expression, acetylation in the cytoplasm controls protein stability and aggregation, and mitochondrial acetylation regulates cell metabolism (195).
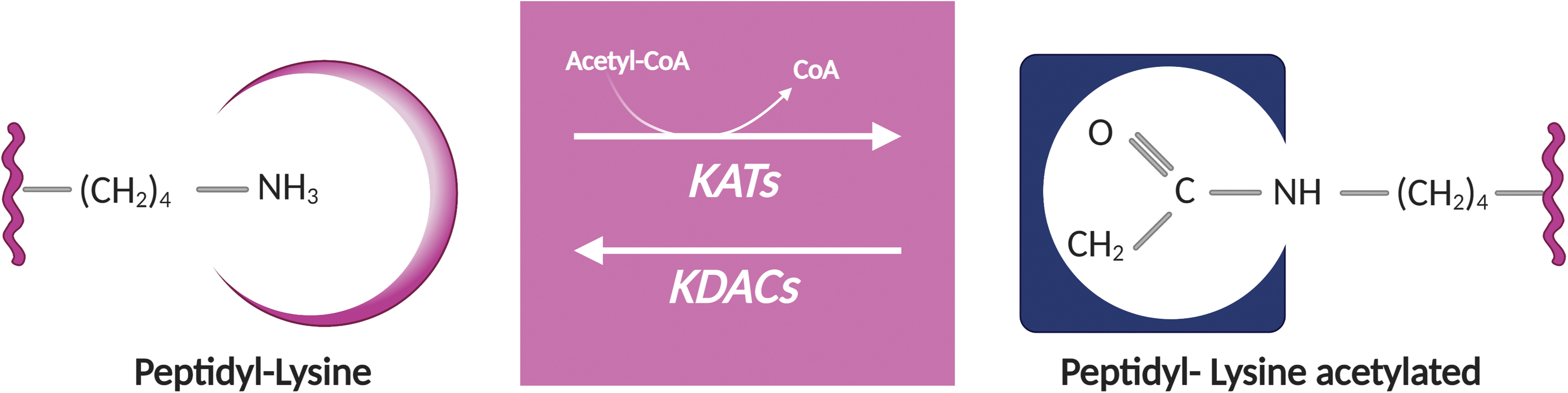
Several thousands of proteins have been identified as targets of acetylation in different cell lines and tissues from human and mouse, which together comprise the acetylome (27, 28). By affecting specific cellular processes (e.g., epigenetic regulation, cellular proliferation, mitochondrial function, and others), aberrant acetylation has been associated with diseases such as cancer and neurodegeneration (2). Whether acetylated proteins are enriched in RA joints is currently unknown.
General Features and Shortcomings of Antibodies to Modified Arginine and Lysine Residues in RA
Antibodies to citrullinated proteins
The discovery of ACPAs resulted from the search for the antigen specificities recognized by APF and AKA (147, 150, 155), two antibody systems identified in patients with RA in 1964 and 1979, respectively (127, 207). Two important premises made by Guy Serre's laboratory guided to the identification of ACPAs. First, filaggrin was identified as the antigen targeted by AKA/APF (150, 155). Second, it was found that AKA/APF only reacted with the acidic/neutral isoform of filaggrin, suggesting that specific modifications in the protein may be required for antibody binding (155). Because filaggrin is highly enriched with citrulline residues, which may explain the migration pattern of the immunogenic isoform (79, 155), this led to the idea that this noncoded amino acid might be present in the epitopes targeted by these antibodies (147).
This hypothesis was confirmed using synthetic peptides derived from the amino acid sequence of human profilaggrin in which arginine residues were substituted with citrulline (147). A 19-mer peptide (amino acids 306–324) was particularly useful in this regard. Nine variants of this peptide in which five arginine residues were changed to citrullines (either individually or in pairs) were reactive to RA sera, while the native sequence did not, providing the first evidence that citrulline residues are a critical target in a subset of antibodies in RA (147). Interestingly, however, there was a remarkable difference in the serum reactivity patterns toward each citrullinated peptide (from 20% to 48% positivity) (147), suggesting that peptidyl-citrulline per se is not the only determinant that confers antibody binding. Instead, the data supported that ACPAs correspond to a pool of antibodies recognizing citrulline residues depending on the context of the neighboring amino acids, which explains their overlapping and nonoverlapping reactivities to multiple citrullinated peptides/proteins (88, 142, 202). The presence of glycine in the +1 flanking position (i.e., citrulline–glycine) is the most common consensus epitope recognized by ACPAs (34, 112, 131, 142, 165, 175), although other consensus binding motifs have been identified for some specific monoclonal ACPAs (e.g., glycine–citrulline or arginine–citrulline–aspartate) (142). In addition to specific amino acid sequences, the presence of varied motifs with structural homology has been proposed to be relevant for antigen recognition among ACPA subsets (149).
Besides the striking association between ACPAs and RA, the importance of citrullination and ACPAs in RA pathogenesis is further strengthened by several findings. (i) ACPAs are produced by plasmacytes within the rheumatoid pannus (117); (ii) their targets (i.e., citrullinated proteins) are abnormally enriched in the RA joints (60, 140, 172, 179, 183, 196); (iii) immune complexes (ICs) containing citrullinated proteins are found in both circulation and synovial fluid from patients with RA (160, 184, 210); (iv) genetic variants in PADI2 and PADI4 (which encode PAD2 and PAD4, respectively) are positively associated with the development of RA (33); and (v) patients with RA develop antibodies to PAD2 and PAD4 (33, 42, 43, 80), including a subset of anti-PAD4 antibodies that favor PAD4 activation and associate with worst disease outcome (43). Together, these findings provide undeniable evidence that citrullination is involved in RA pathogenesis. In particular, the genetic and antigenic features linking the PAD enzymes with RA are difficult to explain mechanistically by antibodies targeting other cross-reactive PTMs (e.g., anti-CarP antibodies and AAPAs) that may question the existence of ACPAs.
Yet, the ACPA paradigm in RA has some limitations. Experimental models of arthritis, considered, by many, mechanistic mimics of RA, have not supported a role of these antibodies in disease pathogenesis. Neither spontaneous nor induced models of arthritis (e.g., tumor necrosis factor alpha-overexpressing mice and collagen-induced arthritis, respectively) develop antibodies to citrullinated proteins (20, 84, 103, 151, 168, 169, 189, 192). Certainly, antibodies to cyclic citrullinated peptides (anticyclic citrullinated peptide antibodies [anti-CCPs]) can be detected in some animal models of arthritis, but it has been demonstrated that these antibodies are not specific to citrulline residues (20, 192). The study of anti-CCPs or any other method of ACPA detection without any control of specificity in experimental models of arthritis is mainly made as a routine test rather than to understand mechanisms.
Moreover, immunization with citrullinated proteins has no advantage to induce arthritis compared with native proteins in mice (159), in some cases the induction of arthritis using citrullinated proteins has been controversial (83, 141), and the antibody response to citrullinated proteins is usually not citrulline-specific (20, 84, 92). Indeed, experimental evidence has demonstrated that B cells reactive to citrullinated proteins are negatively selected both in normal and autoimmune mice (57, 204). In some cases, mouse anticitrulline-specific monoclonal antibodies have been generated by immunization with citrullinated antigens from different species (182). However, different to other PTMs (46, 124, 166, 200), there is no evidence that citrulline residues per se (in autologous proteins) are immunogenic (83).
Antibodies to carbamylated proteins
The study of anti-CarP antibodies resulted from the search for PTMs that may explain the pathogenic role of citrullinated proteins in RA (124, 178). Because attempts of using citrullinated proteins or antibodies raised against citrullinated peptides to induce experimental arthritis yielded modest or negative results (56, 113, 182), it was postulated that other factors might be responsible for triggering arthritis. Since citrulline and homocitrulline are structurally similar, it was hypothesized that carbamylated proteins may be the “missing link” between citrullination and RA (124, 178). In support of this hypothesis, initial studies demonstrated that immunization with carbamylated peptides or autologous carbamylated albumin, but not with citrullinated peptides/albumin, renders mice susceptible to an aggressive form of erosive arthritis following a subsequent intra-articular injection of citrullinated peptides (124). Similarly, adoptive transfer of T and B cells from carbamylated-immunized mice into normal recipients predisposed to aggressive arthritis after intra-articular injection of citrullinated peptides (124). Lastly, the data were strengthened by the finding that homocitrulline-containing proteins are present in the RA joint (124). From these data, it was concluded that the immune-activating effects of carbamylated proteins promote the arthritogenic effects of citrullinated proteins (124). Interestingly, a parallel study showed that antibodies raised in rabbits to carbamylated proteins cross-reacted with citrullinated peptides, opening the possibility that ACPAs may be caused by homocitrulline containing proteins (178).
These interesting observations sparked interest in defining whether patients with RA have antibodies to carbamylated proteins, which was further demonstrated using carbamylated fetal calf serum (FCS) and carbamylated fibrinogen as surrogate antigens (153). In this regard, although carbamylation targets the α-amino end of proteins and side-chain groups of lysine, arginine, and cysteine residues (29, 47, 162 –164, 170), it is assumed that anti-CarP antibodies in RA are exclusively directed against carbamylated lysine (homocitrulline) residues, because of the structural similarity with citrulline (153). However, since the detection of anti-CarP antibodies is made with proteins modified in vitro with potassium cyanate, of which specificity is not exclusive to the ɛ-amino group of lysine residues, whether other modified amino acids besides homocitrulline are also recognized by these antibodies is unclear.
In initial studies, it was found that ∼70% of immunoglobulin G (IgG) ACPA-positive RA tested positive for anti-CarP IgG antibodies, whereas ∼16% of the ACPA-negative RA were anti-CarP antibody positive (153). Using citrullinated FCS and CCP1 [a CCP derived from filaggrin (148)] in competition assays in sera that were reactive to both citrullinated and carbamylated antigens, anti-CarP antibodies showed limited cross-reactivity with citrullinated FCS/CCP1, initially suggesting that these antibodies are not a subset of ACPAs (153). However, further studies using different sets of peptides containing citrulline or homocitrulline residues have shown that some ACPAs and anti-CarP antibodies are cross-reactive (92, 95, 134, 142, 177). These discrepancies may be the result of the distinct targets used for detection and/or competition assays, as the range of epitopes and sites of carbamylation/citrullination is unlikely to be equivalent in synthetic small peptides compared with carbamylated/citrullinated FCS.
In addition to the compelling experimental models that drove initial interest in the study of carbamylation in RA (124, 178), it is noteworthy that in contrast to ACPAs, several experimental models of arthritis develop anti-CarP antibodies (168, 169, 189). These findings suggest that anti-CarP antibodies may be relevant in experimental arthritis, and support the idea that at least in mice, ACPAs and anti-CarP antibodies are not the same antibody. Another important difference between citrullination and carbamylation is that carbamylated residues are highly immunogenic (92, 178) and do not require a xenoreactive response to induce antibodies. Indeed, similar to other chemical modifications in proteins (200), antibodies specific to carbamylated proteins are efficiently generated by immunization with autologous carbamylated proteins (46, 124, 166), making carbamylation an intriguing arthritogenic candidate to initiate preclinical RA. Whereas this notion is supported by the finding that anti-CarP antibodies raised in rabbits by immunization with human carbamylated albumin or carbamylated collagen react with citrullinated peptides (178), it is noteworthy that anti-CarP antibodies generated in mice using carbamylated ovalbumin only reacted with human carbamylated fibrinogen but not with citrullinated fibrinogen (92). These discrepancies may be explained by differences in the host, the type of antigen, or because, as opposed to humans and potentially other species, somehow anti-CarP antibodies from mice have no cross-reactivity with citrullinated sequences. Anti-CarP antibodies generated in mice, however, cross-react with acetylated fibrinogen. This is not surprising, considering that both modifications target the same amino acid residue (lysine) and common epitopes containing either homocitrulline or KAc are likely to be detected by anti-CarP antibodies.
Despite the tempting observation regarding the potential importance of carbamylation in RA, a major caveat in this model is that identifying carbamylated antigens in RA has been challenging. In initial studies, only one peptide containing homocitrulline derived from alpha-1-antitrypsin was found in synovial fluid from an RA patient (190). This importantly contrasts with more than 100 citrullinated proteins found in the RA joint (60, 140, 172, 179, 183, 196). More recently, a proteome analysis of joint tissue (i.e., synovium, cartilage, and synovial fluid) identified multiple proteins containing homocitrulline (188), but their content was not different between OA, RA, and control tissue (only cartilage was analyzed) (188), suggesting that carbamylated proteins in the joints are likely the result of aging rather than a consequence of some pathogenic process.
Antibodies to acetylated proteins
The identification of AAPAs followed the search for antibodies targeting additional PTMs that may mimic citrullination and/or carbamylation in RA. Since KAc resembles homocitrulline (except for the side-chain terminal amine, which is replaced by a methyl group), it was hypothesized that this modification may also be recognized by antibodies in RA (91). However, different to the discovery of ACPAs and anti-CarP antibodies, which was based on the detection of modified antigens with potential biological relevance (147, 153), AAPAs were initially identified using a peptide sequence that does not exist in proteins (91).
A 12 amino acid peptide derived from the sequence of a vimentin variant used to detect ACPAs [termed mutated and citrullinated vimentin, MCV (14)] was changed to include KAc instead of citrulline at position 7, generating a new peptide sequence that does not exist in vimentin or any other protein (91). The rationale of using this peptide to screen for AAPAs may have followed the idea that antigen recognition by these antibodies is only dependent on the presence of KAc, and therefore, the peptide backbone should have no relevance. Nevertheless, it has been clear since the discovery of ACPAs that efficient antigen binding requires accompanying interactions in addition to the presence of citrulline (34, 71, 142, 147, 165, 175). Thus, ACPA reactivity to a peptide containing an ideal ACPA recognition motif in which citrulline was changed for a structurally similar residue (e.g., KAc or homocitrulline) may suggest that some of these antibodies tolerate this change in the antigen, rather than concluding the existence of a new subset of antibodies in RA.
Further studies have followed a similar approach to detect AAPAs, but instead of KAc, acetylated ornithine (which is one methylene group shorter than KAc) has been included at the site of citrulline within the MCV peptide (62). Similar to the MCV peptide containing KAc, ACPA-positive sera reacted with the peptide containing the acetylated ornithine residue (62), suggesting that binding to this peptide is also tolerated by some ACPAs. Importantly, ornithine is not an amino acid present in proteins. Thus, whereas acetylated MCV peptides (either containing lysine or ornithine) may be useful to detect some antibody subsets in RA, their importance as a source of autoantigens in RA is very unlikely.
Analogous to antibodies to carbamylated ovalbumin generated in mice (92), immunization of mice with acetylated ovalbumin generates antibodies that react with human acetylated fibrinogen and to some extent with carbamylated fibrinogen, but not with citrullinated fibrinogen (92). Thus, while the data support cross-reactivity among anti-CarP antibodies and AAPAs generated by immunization in mice, it is clear that these antibodies have no relevance in the recognition of citrullinated epitopes.
Although the initial data suggesting the existence of AAPAs in RA are not convincing (91), further studies have pursued this modification in RA, but using antigens containing real acetylation sites (e.g., acetylated fibrinogen and peptides derived from histones) and monoclonal ACPAs (95, 142). The data from these studies suggest that some ACPAs can react with acetylated epitopes (some even more efficiently than citrullinated epitopes), supporting cross-reactivity of ACPA subsets with some acetylated antigens, but rigorous evidence to confirm that AAPAs are independent antibodies in RA is still lacking.
The Potential Significance of Cross-Reactivity Within and Outside the ACPA Response in RA
Two interesting findings in the study of ACPAs may contain important clues regarding the origin of these antibodies. One is that monoclonal ACPAs are cross-reactive with a large variety of citrullinated peptides, which has been explained by their recognition of specific citrullinated motifs shared among different peptides (142). However, why ACPAs target different motifs and whether these motifs are stochastically or mechanistically selected is unclear. The second is the ACPA cross-reactivity outside citrullinated antigens, particularly with carbamylated proteins, and whether and how citrullination and carbamylation may be mechanistically related to induce the production of pathogenic antibodies in RA.
Cross-reactivity within the ACPA response: citrullination by distinct PAD isotypes as a potential mechanism of ACPA diversity and specificity
A unique feature among ACPAs is that these antibodies recognize linear motifs in which few amino acid residues flanking citrulline are required for efficient binding. Using small peptides (e.g., 9–16-mers) derived from protein sequences in which a citrulline is located around the center of the peptide (which is referred to as position zero), multiple studies have demonstrated that the most common motifs targeted by ACPAs contain glycine in the +1 flanking position (i.e., citrulline–glycine) (34, 112, 131, 142, 165, 175), which is also the most common consensus sequence citrullinated by PAD2 and PAD4 (10, 82, 167). Further studies using monoclonal ACPAs have shown that the citrulline–glycine sequence can have different recognition patterns due to distinct amino acid preferences in the −1 position (142). For instance, the presence of some amino acid residues favors antibody binding (e.g., lysine, asparagine, glycine, arginine, and alanine), while others can interfere with citrulline–glycine recognition (e.g., negatively charged or aromatic amino acids). Similarly, efficient binding of the citrulline–glycine sequence can be influenced by amino acid residues at positions −3, −2, +3, +4, and/or +5, and some ACPA subsets prefer amino acid sequences other than glycine at +1 position (e.g., glycine–citrulline or arginine–citrulline–aspartate) (142).
The recognition of citrullinated antigens based on a consensus sequence (e.g., citrulline–glycine) within common motifs, but not because of protein specificity, explains that ACPAs can bind a broad range of citrullinated peptides (142). However, the preference for unique amino acid residues within ACPA recognition motifs (from −5 to +5 positions) also reveals why not every citrullinated protein is a target for these antibodies (139). Besides the citrulline–glycine sequence, which is a target for both PAD2 and PAD4 (10, 82, 167), it is unclear how other amino acid residues involved in ACPA recognition might be determined. Interestingly, some patterns of ACPA recognition are analogous to consensus sequences that define the distinct substrate specificity between PAD2 and PAD4 (10, 82, 167). Whereas the most common citrullination site for both isotypes is arginine–glycine, the extended consensus sequence (i.e., positions −4 to +4 relative to the arginine) for human PAD2 and PAD4 is different (10, 82) (Fig. 6). Thus, depending on the amino acid context, some arginine residues are only converted to citrulline by one of these PAD isotypes (82). In this setting, it is possible to suggest that ACPA specificity is driven by unique motifs citrullinated by PAD2, PAD4, or by both enzymes. In support of this model, it is noteworthy that the consensus sequence arginine–arginine–aspartate and glycine–arginine (targeted by ACPAs as arginine–citrulline–aspartate and glycine–citrulline, respectively) are citrullination sites preferred by PAD4 (10, 82). Similarly, serine/threonine/lysine/asparagine residues seen in citrulline-flanking regions targeted by some ACPAs are also amino acid residues that favor citrullination by PAD4 (167). The absence of proline at +1 position in ACPA binding motifs may reflect that this amino acid is disfavored by PAD4 (167).

ACPA recognition of specific sequences citrullinated by unique PAD isotypes is also supported by the study of macromolecular substrates. For instance, ACPAs preferentially bind fibrinogen citrullinated by PAD4 (16), and these antibodies also distinguish among proteins citrullinated by either PAD2 or PAD4 in hypercitrullinated cells (139). The lack of ACPA recognition of citrullinated proteins, such as autocitrullinated PAD2 and PAD4, citrullinated bacterial peptidylarginine deiminase from Porphyromonas gingivalis, and citrullinated human serum albumin (35, 41, 104), provides additional evidence that not every citrullinated protein is a target for these antibodies. Thus, despite that ACPAs may not have protein specificity, these antibodies still recognize specific citrullinated proteins within any potential citrullinome (139).
In fact, although ACPAs bind linear epitopes, not every peptide recognized by these antibodies may parallel the primary protein target in vivo. The binding specificities for linear epitopes depend on both their amino acid sequence and their conformational structure (63). Linear peptides can have extensive cross-reactivity because they have a limited number of binding residues. Moreover, peptides displayed on a microarray likely adapt to almost any conformation upon binding to the antibody (“induced fit”), allowing peptides with sequences similar to the epitope to interact with the antibody (63). However, cross-reactive sequences are not recognized when displayed in full-length proteins, likely because the epitope is not exposed in the correct conformation for binding (63). Thus, while the analysis of peptide arrays can be useful to identify consensus sequences related to ACPA binding, the data need to be taken with caution when translated to the real citrullinated protein targets for these antibodies. Indeed, it is possible that the epitopes targeted by ACPAs, and their corresponding proteins, may compose a more restricted repertoire than the expected based on the sole analysis of citrullinated peptides on arrays (142). Similarly, considering that antigen recognition by ACPAs requires additional interactions with amino acid residues flanking citrulline (142), the finding that ACPAs appear to bind small synthetic peptides that end with a C-terminal citrulline is suspicious (21), and highlights the risks of using peptides to identify or confirm citrullinated antigens in RA.
Cross-reactivity outside the ACPA response: potential significance for disease initiation
Since ACPAs recognize a still unknown array of motifs containing citrulline, it is not difficult to reconcile the idea that anti-CarP antibodies and AAPAs belong to a subset of ACPAs that cross-react with carbamylated and acetylated proteins/peptides. Moreover, the reactivity of ACPAs with sequences containing carbamylated or KAc residues may also result from using small cross-reactive peptides (142), which likely have the correct conformation for antibody binding (63). This may be the case for AAPAs, unless some new convincing evidence demonstrates the opposite. However, the case of ACPAs and anti-CarP antibodies is of particular interest in this regard, because there is good evidence to support both that these are different and the same antibodies (92, 95, 134, 142, 176, 177). The proposal that ACPAs and anti-CarP antibodies raise from a common B cell precursor in which each specificity may dominate or parallel depending on the antigenic environment (citrullinated vs. carbamylated) is especially interesting (95), as it may explain why these antibodies can be independent and sometimes overlapping. Yet, there are still some shortcomings to fully support this paradigm.
Neither carbamylation nor citrullination provides an ideal model to individually explain both the initial breach of tolerance and the maintenance of antibody production in RA. Since carbamylated amino acid residues are immunogenic (46, 124, 166), but citrullinated residues are not (20, 83, 84, 92), carbamylation provides a better model to breach tolerance in RA. Paradoxically, citrullinated proteins are highly enriched in target tissues in RA (118, 140, 172, 179, 183), but carbamylated proteins are not abnormally accumulated in RA joints (188). In this scenario, it is difficult to reconcile how the production and pathogenic effect of anti-CarP antibodies could be sustained in the absence of an abnormal source of antigen. Instead, citrullination is a better model to maintain the production of pathogenic antibodies in RA.
An alternative model to explain this paradox is that carbamylation and citrullination are complementary to each other, which is somehow the original idea that drove the interest in studying carbamylation in RA (124, 178). Thus, a proposal is that an initial antibody response to carbamylated proteins may be the bridge to the production of ACPAs in RA. To discuss this model, it is important to first review how minor structural changes in amino acid residues can affect the immune response to self-proteins.
Chemical PTMs are haptens that render self-proteins the target of antibodies
Although self-proteins are not immunogenic, studies by Obermayer and Pick in the early 1900s found that animals immunized with autologous proteins chemically modified by the action of iodine, nitric acid, or nitrous acid (generating proteins containing iodo, nitro, or diazo groups, respectively) generate antibodies that react with the same modified self-proteins, as well as to the same modification in proteins derived from an entirely different species or even from plants (200). The initial thought was that the species specificity of proteins was related to their content of aromatic residues and that substances that combine with the benzene ring will endow the molecule with a new structural specificity. Later studies by Landsteiner and Lampl, however, indicated that the grouping, which determines specificity, may be entirely devoid of aromatic radicals (108). Indeed, other modifications that do not depend on reacting with aromatic radicals, such as esterification, acetylation, and methylation, were also capable of inducing antibodies to proteins containing these modifications (108, 200).
Together, these findings led to the concept that introducing chemical groups into proteins endowed those proteins with antigenic properties characteristic of the chemical groups, while the species specificity (i.e., tolerance) of the protein is lost. Although this concept may be correct, it is important to consider that chemical groups work as haptens. These are small molecules that cannot induce an antibody response unless they are attached to a larger carrier (107). Thus, the immune response to chemically modified self-proteins is not the consequence of a breach of tolerance against self-antigens, but a normal immune response against a hapten. In this scenario, even though these antibodies were generated with and react against modified self-antigens, these antibodies should not be considered autoantibodies but antihapten antibodies.
Relevant to this review, studies in the 1980s by Steinbrecher et al. demonstrated that derivatization of lysine residues by carbamylation renders autologous LDL, albumin, and fibrinogen highly immunogenic in guinea pigs (166). Antibodies generated against these self-antigens reacted with the modified protein, but not with their native counterpart, demonstrating that tolerance to the self-antigens was not altered. Interestingly, however, the specificity of the antibodies against these carbamylated proteins was strikingly influenced by the nature of the antigen. Thus, antibodies produced against carbamylated LDL were able to bind free homocitrulline and reacted equivalently to other carbamylated proteins from the same and other species (i.e., guinea pig carbamylated albumin and human carbamylated LDL), suggesting that these antibodies were specific to the hapten (i.e., homocitrulline). In contrast, reactivity of antibodies generated against carbamylated albumin or carbamylated fibrinogen was not affected by free homocitrulline, and antibodies to carbamylated albumin showed no reactivity to human carbamylated albumin. This subset of anti-CarP antibodies has therefore a unique specificity to carbamylated amino acids within proteins and in the case of antibodies to carbamylated guinea pig albumin, efficient binding required additional amino acid residues specific for guinea pig albumin. Further studies in mice using carbamylated mouse fibrinogen as immunogen also induced the production of anti-CarP antibodies reactive to other carbamylated proteins from the same and other species (i.e., mouse carbamylated albumin and carbamylated ovalbumin) (46), adding evidence that carbamylated amino acid residues are immunogenic.
The heterogeneity in antibody specificities induced by specific chemical modifications is consistent with previous observations by Landsteiner (108, 200), and strongly suggests that the nature of the carrier protein is an important determinant in the specificity of antibodies to modified proteins. Thus, some modified proteins may generate antibodies with high reactivity to the hapten (e.g., homocitrulline), allowing these antibodies to bind any structure similar to the modified amino acid residue independently of the protein carrier or even without a carrier. In contrast, other proteins containing the same modification may generate antibodies, which can only bind the hapten within the protein and in the context of specific residues flanking the hapten. Interestingly, ACPAs/anti-CarP antibodies can behave in both manners; this is, either cross-reactive or with limited specificity (142). Nevertheless, while mechanisms of tolerance exist to peptidyl-citrulline (57, 204), the data support that this is not the case for carbamylated proteins. Thus, despite the notable physicochemical similarities between citrulline and homocitrulline residues, it is clear that they are not equally seen and tolerated by the immune system.
Carbamylation as a potential initiator of pathogenic antihapten antibodies in RA
The temporal detection of ACPAs and anti-CarP antibodies is not different during preclinical RA (19, 154), suggesting that one antibody is unlikely to be the precursor of the other. These studies, however, did not exclude the possibility of cross-reactivity in samples positive for both ACPAs and anti-CarP antibodies. Therefore, it is still feasible that these studies may be detecting a single specificity (i.e., either ACPAs or anti-CarP antibodies) using two distinct assays. Although citrullination or carbamylation may have the same possibility of being the initiator of an AMPA response, the main reason to suggest a model of carbamylated proteins as “antigen zero” is because of their high immunogenicity and the presence of anti-CarP antibodies in experimental arthritis. To establish the basis of this model, the first proposal is that anti-CarP antibodies are more likely to resemble antihapten rather than autoantibodies. This distinction is important because the generation and cross-reactivity of anti-CarP antibodies are more easily explained as an initial immune response against a hapten, instead of evoking a break of tolerance to distinct epitopes in multiple carbamylated proteins.
Interestingly, immunization with a single carbamylated protein can generate diverse subsets of anti-CarP antibodies, some cross-reactive and others with high specificity to homocitrulline (166, 178). Within the diversity of anti-CarP antibodies, it is feasible to propose that the cross-reactive subset could be involved in the production of ACPAs (Fig. 7). Since cellular hypercitrullination generates a broad range of citrullinated proteins (101, 140), it is possible that some of these modified proteins expose citrulline residues on structural epitopes that may be recognized by cross-reactive anti-CarP antibodies. In the presence of a constant source of cross-reacting citrullinated antigens, ACPAs may develop as result of epitope spreading from an initial anti-CarP antibody response (Fig. 7). As citrullination may become the main source antigen, ACPAs may dominate the pathogenic immune response in RA, but with some remnant cross-reactivity to carbamylated proteins. Similar to anti-CarP antibodies, however, ACPAs are likely to remain analogous to antihapten antibodies, which may explain why these antibodies accumulate large numbers of somatic hypermutations without yielding antigen-specific antibodies (174). Anti-CarP antibodies with high specificity to carbamylated proteins, likely unable to cross-react with citrullinated antigens, may represent the antibody subset that remains ACPA negative in RA.
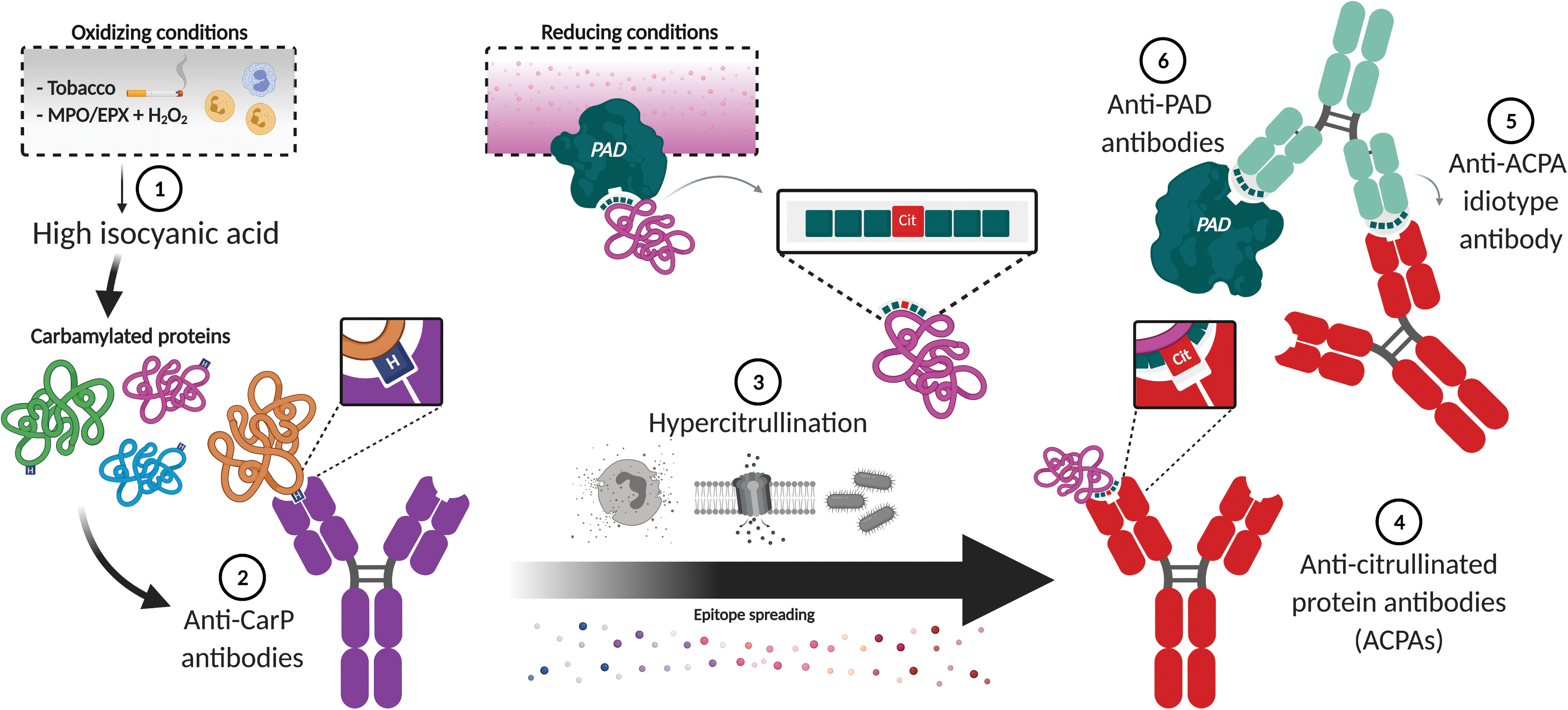
In experimental models of arthritis that develop anti-CarP antibodies, it is possible that mechanisms of hypercitrullination are not required for disease development. Therefore, the absence of a pathogenic source of cross-reactive citrullinated antigens may explain the absence of ACPAs despite the presence of anti-CarP antibodies. Alternatively, some species such as mice may be resistant to produce ACPAs (57, 204).
Potential sources of carbamylation in preclinical RA
Mechanistically, the production of anti-CarP antibodies should be linked to increased exposure to isocyanic acid and the abnormal production of carbamylated proteins. Tobacco smoking is certainly one of the first components to keep in mind as an abnormal source of isocyanic acid in preclinical RA, with the caveat that some epidemiologic studies have not found a direct relationship between tobacco smoking and anti-CarP antibodies in RA (135, 186). Nevertheless, antibodies to carbamylated LDL have been associated with tobacco smoking in healthy controls and smoking mice develop anti-CarP antibodies (106, 129). Moreover, it is intriguing that healthy control neutrophils contain carbamylated proteins (105). Under high cyanate conditions, such as uremia, carbamylated proteins increase in neutrophils and also newly accumulate in monocytes (105). A similar process may occur during smoking, providing a source of inflammatory cells preloaded with carbamylated proteins at sites of inflammation. Thus, despite negative epidemiologic studies, a potential role of tobacco smoking cannot be mechanistically discarded in the induction of anti-CarP antibodies in preclinical RA. Paradoxically, although metals in tobacco, such as cadmium, lead, nickel, manganese, and cobalt (32, 64), are potent inhibitors of calcium-induced PAD activation (66), tobacco smoking is considered a trigger of citrullination in cells (114). Whether tobacco smoking directly activates citrullination in cells, however, has not been experimentally demonstrated.
The abnormal production of carbamylated proteins by leukocyte-derived peroxidases is another potential source of carbamylated antigens in preclinical RA (8, 197, 198). Interestingly, since thiocyanate participates in host defense, extracellular fluids from mucosal sites such as the oral mucosa, lung, and gastrointestinal tract contain the largest amounts of thiocyanate in the body (24). At these sites, peroxidases (e.g., lactoperoxidase, gastric peroxidase, salivary peroxidase, MPO, and EPX) use thiocyanate and hydrogen peroxide to generate hypothiocyanous acid, a relatively weak oxidant with microbicidal activity (24, 96). Isocyanic acid, which is involved in carbamylation, is only a minor product of this reaction (198). However, during chronic mucosal inflammation, such as in asthma, the production of isocyanic acid by leukocyte-derived peroxidases is sufficient to generate carbamylated proteins (197). These findings are particularly important for RA pathogenesis, because chronic mucosal inflammation is likely a major source of carbamylation and it is also considered a potential trigger of RA (73, 85).
Anti-ACPA Idiotype Antibodies As a Model Linked to the Origin of Anti-PAD Antibodies in RA
While this article attempts to propose a model explaining a mechanistic relationship between ACPAs and other antibodies to PTMs in patients with RA, it is intriguing that these patients also develop antibodies to PAD2 and PAD4 (42, 80, 209). The presence of both ACPAs and anti-PAD antibodies in RA is unlikely to be stochastic. However, whether and how the production of these citrullination-associated antibodies is related is still unknown. Although PADs can citrullinate themselves (6, 119, 121, 157), their recognition by anti-PAD antibodies is independent of their citrullination status (41), suggesting that citrullination is not involved in the lack of tolerance to PADs in RA. Several models have tried to explain a mechanistic relationship between ACPAs and anti-PAD antibodies (9, 33). The strongest evidence suggests that ACPAs precede the development of anti-PAD4 antibodies in preclinical RA (33, 61, 100), which highlights the fact that anti-PAD4 antibodies are not required for the development of ACPAs, but ACPAs may be relevant for the production of anti-PAD4 antibodies.
The proposal that ACPAs bind specific consensus sequences recognized by PAD2 or PAD4 may have important implications in the targeting of these enzymes in RA. It is possible to suggest that in some individuals with ACPA-positive RA, the initial antibody response to PAD2 and PAD4 corresponds to anti-idiotypic antibodies directed to ACPA idiotypes recognizing unique sequences citrullinated by either enzyme (Fig. 7). Anti-idiotype-PAD ICs delivered to antigen-presenting cells may further serve as a source of antigen for intramolecular epitope spreading and the induction of anti-PAD antibody subsets, including antibodies that may activate or inhibit the function of PADs (13, 43). Interestingly, despite the high specificity of anti-PAD4 antibodies for RA (>95%) (44, 116), the spontaneous production of antibodies to PAD4 has not been reported in a single mouse model of arthritis, which may be explained because these experimental models cannot develop ACPAs.
Redox Modulation As Therapeutic Target to Control Autoantigen Production in RA
While different mechanisms are likely involved in the production of autoantigens in RA, it is interesting that the redox environment is a common modulator where citrullination and carbamylation concur. Unfortunately, however, these PTMs are controlled by opposite redox conditions. Thus, at least in theory, modifying the redox balance to either block citrullination or carbamylation may paradoxically increase the production of the other modification. This caveat in modulating the redox environment as a potential therapeutic target in RA is not exclusive for citrullination and carbamylation.
Since joint damage in RA has been traditionally linked to excessive oxidative stress (72), several studies have considered the use of antioxidants for the prevention and treatment of RA (23, 31, 58, 76, 81, 130, 185). However, the clinical evidence for therapeutic use of antioxidants in RA is unconvincing. Interestingly, similar to citrullination, other pathogenic processes have been linked to “reductive stress” in RA (201). For instance, inactivation of the ataxia telangiectasia mutated (ATM) kinase due to ROS deficiency in CD4+ T cells in RA induces hyperproliferation and biased differentiation of T cells into interferon-gamma and interleukin-17-producing inflammatory cells (205). Thus, in contrast to the traditional thought of using antioxidants in RA, restoring oxidant signaling using prooxidants was effective in suppressing T cell-induced synovitis in human synovium mouse chimeras (205).
Although a single redox pathway is unlikely to be dominant in RA, it is noteworthy that although the origin of ACPAs and anti-CarP antibodies may be related, these antibodies can distinguish patient subsets within the RA population. Considering that citrullinated and carbamylated antigens are generated under reducing and oxidative conditions, respectively, it is possible that antibodies to these antigens may reflect unique redox environments that may dominate during autoantigen production in individual patients with RA. These antibodies may therefore serve as useful biomarkers to select specific antiredox therapies in RA. A better understanding of the mechanisms involved in the abnormal production of immunogenic modified proteins in RA may aid to support or discard this notion.
Conclusions
A mechanistic association between citrullination and RA is not only based on the presence of ACPAs, but also strengthened by the humoral immune response to multiple components involved in the process of citrullination (i.e., the enzymes and the substrates), by the accumulation of citrullinated proteins in RA joints, and by the presence of PADI2 and PADI4 variants that confer genetic susceptibility to RA. The finding that ACPAs are cross-reactive with other PTMs with structural similarity to citrulline is therefore not hostile to the importance of citrullination in RA, but still demands understanding the significance of such cross-reactivity. The possibility that anti-CarP antibodies and AAPAs are simply subsets of ACPAs requires a major consideration, because as artifacts, these antibodies can unnecessarily spend resources and create false pathogenic hypotheses. There is also the alternative that these antibodies may inform about mechanisms that cannot be revealed by the sole study of citrullination in RA. In this regard, while the independent importance of AAPAs in RA requires a more robust evidence, anti-CarP antibodies have interesting features that may complement the study of ACPAs in RA. Moreover, anti-CarP antibodies and ACPAs may serve as unique biomarkers to distinguish specific metabolic conditions associated with autoantigen production in individual patients with RA.
Independently of any proposal to explain the origin and specificity of the abnormal antibody response in RA, it is certain that these antibodies are distinct from the autoantibody response observed in other autoimmune diseases. These differences may have important implications regarding unique mechanisms of disease initiation, and how modified proteins are processed, presented, and recognized by the immune system in RA.
Footnotes
Authors' Contributions
F.A. conceptualized and designed the article; M.I.T-Z., E.G-B., and F.A. wrote the article draft; and M.I.T-Z. designed the figures. All authors contributed to the preparation of this article and approved the final version for publication.
Acknowledgments
The authors gratefully acknowledge helpful discussions with Dr. Erika Darrah (Johns Hopkins University School of Medicine). Figures were created with
Disclaimer
The content of this article is solely the responsibility of the authors and does not represent the official views of the National Institute of Arthritis and Musculoskeletal and Skin Diseases or the National Institutes of Health.
Author Disclosure Statement
Dr. F.A. is an inventor on a patent for human autoantibodies specific for peptidylarginine deiminase type 3 (PAD3), which are cross-reactive with PAD4, and their use in the diagnosis and treatment of rheumatoid arthritis and related diseases (US patent no. 8,975,033), and on a provisional patent application for anti-PAD2 antibody for treating and evaluating rheumatoid arthritis (US patent no. 62/481,158). Dr. F.A. has received consulting fees, speaking fees, and/or honoraria from Bristol-Myers Squibb, Celgene, and Advise Connect Inspire.
Funding Information
This research was supported by Jerome L. Greene Foundation Scholar and the National Institute of Arthritis and Musculoskeletal and Skin Diseases/National Institutes of Health grant R01 AR069569.