
Abstract
Significance:
Diabetes is a major source of neuropathy and neuropathic pain that is set to continue growing in prevalence. Diabetic peripheral neuropathy (DPN) and pain associated with diabetes are not adequately managed by current treatment regimens. Perhaps the greatest difficulty in treating DPN is the complex pathophysiology, which involves aspects of metabolic disruption and neurotrophic deficits, along with neuroimmune interactions. There is, therefore, an urgent need to pursue novel therapeutic options targeting the key cellular and molecular players.
Recent Advances:
To that end, cellular targeting becomes an increasingly compelling drug delivery option as our knowledge of neuroimmune interactions continues to mount. These nanomedicine-based approaches afford a potentially unparalleled specificity and longevity of drug targeting, using novel or established compounds, all while minimizing off-target effects.
Critical Issues:
The DPN therapeutics directly targeted at the nervous system make up the bulk of currently available treatment options. However, there are significant opportunities based on the targeting of non-neuronal cells and neuroimmune interactions in DPN.
Future Directions:
Nanomedicine-based agents represent an exciting opportunity for the treatment of DPN with the goals of improving the efficacy and safety profile of analgesia, as well as restoring peripheral neuroregenerative capacity. Antioxid. Redox Signal. 36, 122–143.
Background
Diabetic neuropathic pain: the scale of the therapeutic challenge
Diabetic peripheral neuropathy (DPN) is a major and irreversible complication of diabetes and the most common form of neuropathy worldwide, affecting 30%–50% of patients with diabetes (71, 166, 175). In a recent report, the National Center for Chronic Disease Prevention and Health Promotion estimated that 30.3 million people in the United States live with diabetes (9.4% of the population) and as many as 84.1 million adults (33.9% of U.S. adults) are prediabetic (17). This means that approximately two of every five U.S. adults have diabetes or prediabetes, with the potential for this prevalence to continue increasing.
The pain, impaired nerve regeneration, and sensorimotor deficits associated with DPN represent a significant source of disability, ulceration, and infection, which can necessitate amputation (57, 76, 80, 133). Further, currently available treatments achieve significant pain relief in only half of the patients (160). The high incidence of diabetes poses an additional threat to overall health through increased complications post-peripheral nerve injury, a common occurrence due to trauma and/or surgery. Further, the axonal regeneration is significantly impaired by hyperglycemia in diabetes. Several animal models have demonstrated that nerve regeneration is impacted through multiple intersecting mechanisms (146). An additional challenge in the treatment of DPN is that current therapies do not necessarily support nerve regeneration, which is associated with the restoration of function and the alleviation of pain symptoms. The complex neuro-immune interactions driving impaired neuroregeneration in diabetes represents a source of new cellular targets that could both help reverse this trend and treat DPN pain.
A major cause of DPN is hyperglycemia, which increases oxidative stress (7). The distal nerve endings in the hands and feet are especially vulnerable to such damage, resulting in the typical “stocking and glove” distribution of sensory loss and pain (Fig. 1). Loss of intraepidermal nerve fibers in patients and rodent models of pre-diabetes, metabolic syndrome, and overt diabetes has been widely reported (16, 122, 163), and it reflects the impaired nerved regeneration capacity (80). Along with impaired nerve regeneration and wound healing in general, persistent pain and other sensory disturbances are common complications of diabetes (157, 176, 179, 182). Approximately 15%–20% of patients with DPN report the most common and early symptom of DPN: pain in the feet, often described as a stabbing or burning sensation, a symptom of small myelinated fiber involvement. Although the symptoms can spontaneously resolve within a year of onset in about half of the patients with proper glycemic control, others suffer from persistent symptoms, resulting in a significant detrimental impact on their quality of life (
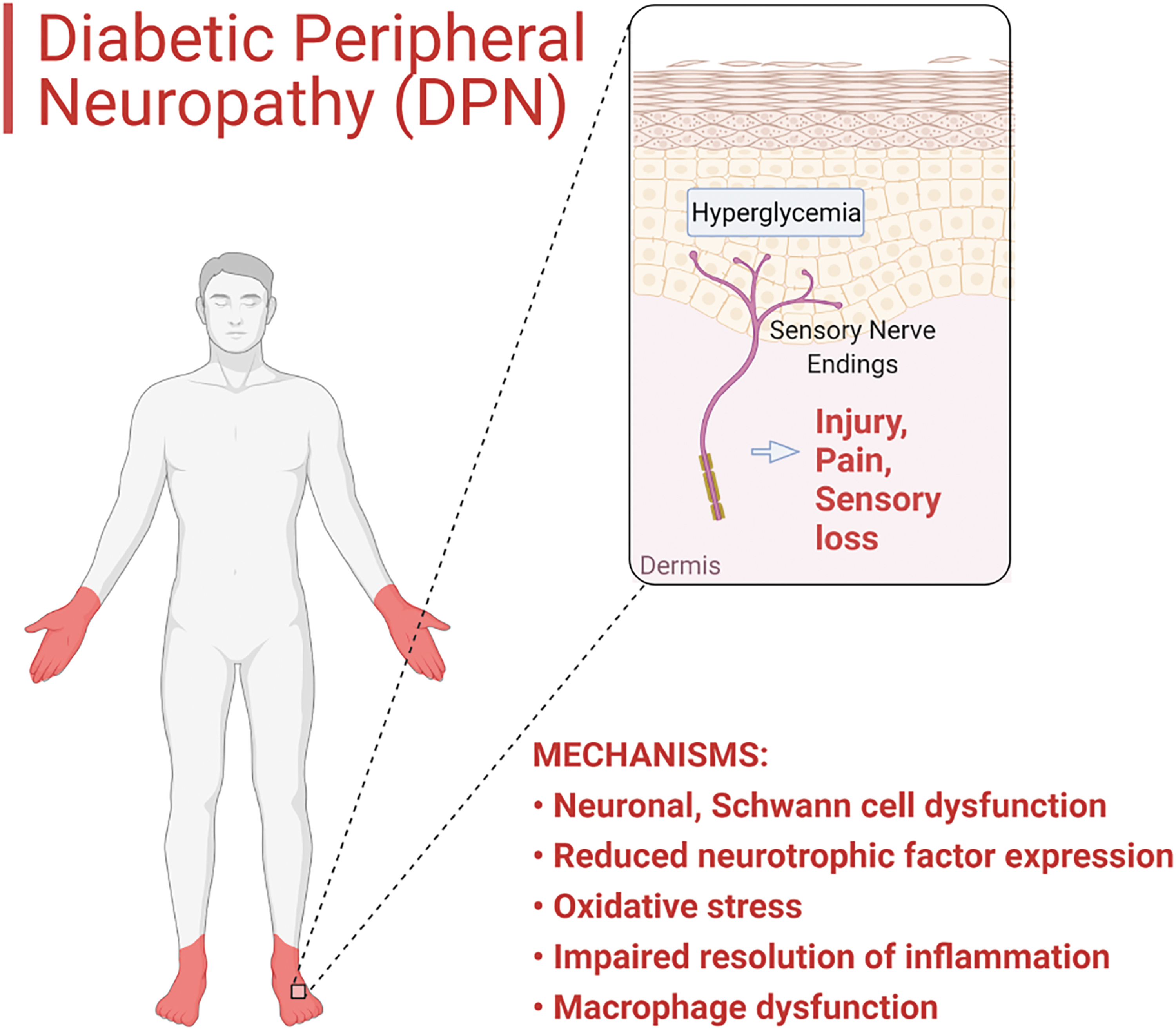
First-line pharmacotherapies for DPN overlap significantly with those used to treat other types of neuropathic conditions, since neuropathy-specific medications are currently lacking (119). The first-line therapy of DPN includes antidepressants (e.g., duloxetine, venlafaxine, amitriptyline, and other tricyclic drugs) and gabapentinoid antiepileptic drugs (e.g., pregabalin, gabapentin) (58). In other words, the most utilized pharmacological targets in DPN are monoamine transporters (inhibition of norepinephrine and serotonin reuptake) and voltage-dependent ion channels (typically Ca2+ channels), respectively. Currently, the clinical practice tends to target the central nervous system (CNS) and lacks medications that directly treat peripheral nerve damage, leading to an emphasis on prevention as a key component in diabetes care (58, 119, 135).
The need for new DPN diagnostics and therapeutics
It is evident that no single treatment for DPN can be regarded as comprehensively effective and completely safe, and no currently used therapeutics treat or influence the main underlying cause of DPN: neuronal damage, which is at least partly driven by complex interplay between the nervous and immune system [in addition to metabolic disruption, vascular damage, and other non-neuronal cells or processes, which are discussed in detail in other reviews such as (150, 159)]. Therefore, the introduction of novel agents or therapeutic delivery approaches are needed to advance into the clinical practice with the goal of improving analgesic efficacy and/or enhance the regenerative processes of nerves in DPN. There is tremendous potential for cellular and targeted therapies, including tissue/cell targeted, nanotechnology-based treatments that could not only treat but also reverse underlying pathologies driving DPN. All currently used treatments for DPN, both systemic and local, do not seem to be disease-modifying; they do not cause any long-term changes that could potentially reverse cellular dysfunction, reduce pain, and improve regeneration. In addition, these chronic treatments often lead to the development of tolerance and reduced responses, which then require increases in dosing and elevate the risk of side effects. Finally, current treatments, both single-agent and combinations, are very difficult to personalize, leaving the clinician to make indirect judgments based on potential improvement in symptoms as the gauge of therapeutic success. This is why in this article we propose two specific roles for targeted nanotechnologies in DPN, namely therapeutic and diagnostic. We will survey accessible targets for these developments and propose adapting or tailoring some of the existing nanotechnologies applied to other diseases such as cancer, autoimmune and inflammatory diseases (e.g., rheumatoid arthritis, or inflammatory bowel disease). In this article, we lay out the specific cellular targets that we believe are the most well understood (with particular regard to neuro-immune interactions) and exhibit the greatest potential for novel DPN therapeutics and diagnostic nanotechnology development.
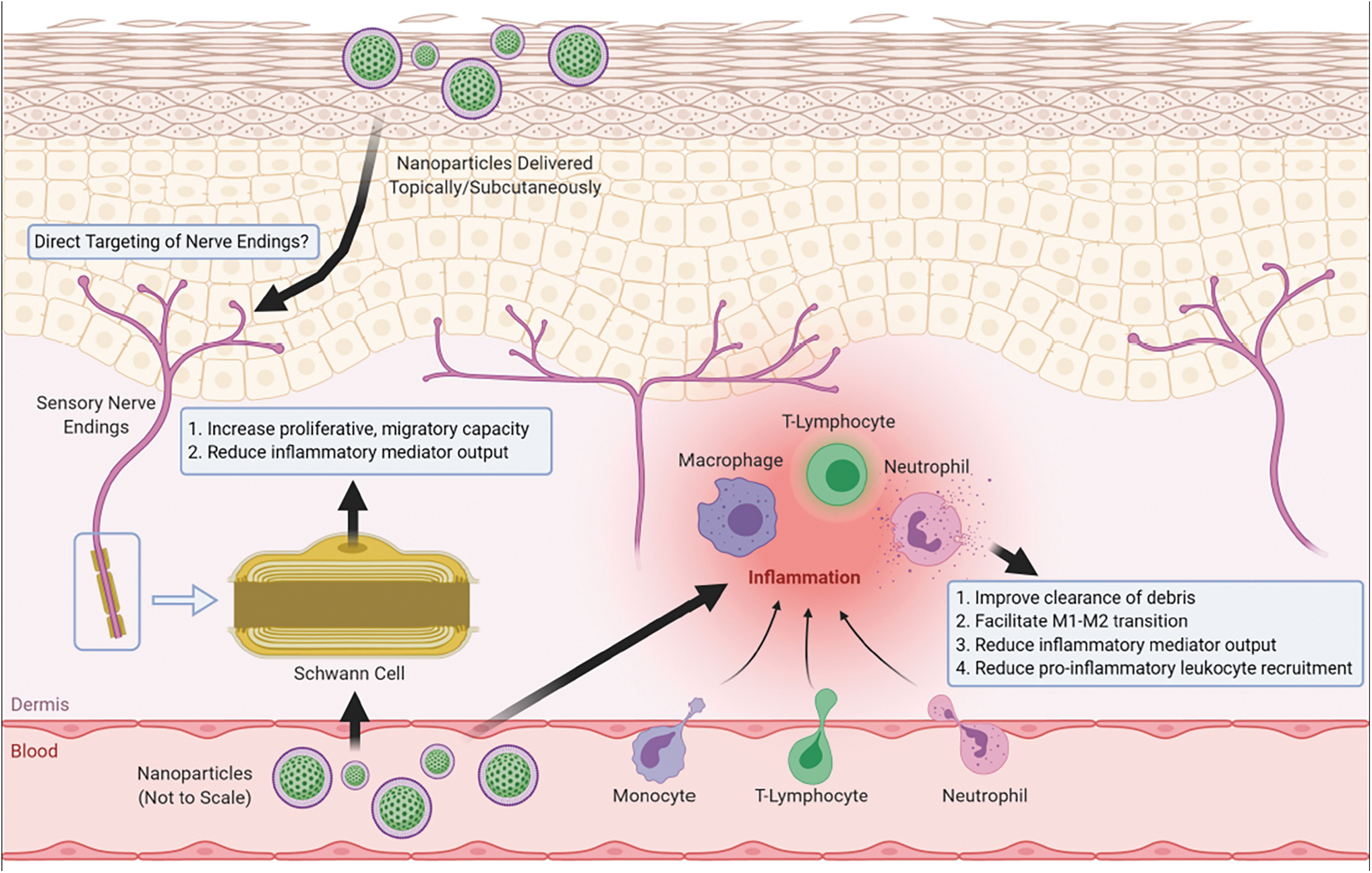
“Nanomedicine”, or the use of nanotechnology in medicine, has been extensively researched for decades, from therapeutic drug delivery technologies to diagnostics and sensors of biological changes during the course of disease. Our purpose here is to highlight the potential of nanomedicine as applied to the treatment and diagnosis in DPN and propose how nanomedicine can further advance our understanding of this complex problem. In upcoming sections, we lay out molecular and cellular targets, starting from current first-line therapeutic targets in DPN to novel targets recently uncovered in preclinical research. The focus will be the key cellular and molecular players in the peripheral nervous system (PNS) and PNS-immune system interactions. The CNS-targeted nanotechnologies currently developed to treat neurodegenerative diseases are extensively reviewed elsewhere (2, 129, 158), and only those applicable for peripheral targeting will be included here. We present examples of applicable nanotechnologies that could engage these targets to both treat DPN pain and potentially support healing persistent neuronal damage in patients with diabetes. The current proposed novel mechanistic strategies are summarized in Figure 2.
Nervous System Targets as Current First-Line Therapy of Pain in DPN
Monoamine transporter inhibitors
The only monoamine transporter inhibitor that is approved by the U.S. Food and Drug Administration (FDA) and the European Medicines Agency (EMA) for the management of DPN is duloxetine (135). Duloxetine is a serotonin and noradrenaline reuptake inhibitor (SNRI). Serotonergic and noradrenergic neurons in the superficial dorsal/posterior horns of the spinal cord are a key point in pain transmission (177). These descending inhibitory pain pathways inhibit ascending input from the periphery, a process that is augmented by inhibiting the reuptake of serotonin and noradrenaline (21, 177). Tricyclic antidepressants (TCAs, e.g., amitriptyline, desipramine, nortriptyline) can be also utilized for the management of DPN. The TCAs work on the same principle as SNRIs, but they exert a more complex mechanism of action: Besides inhibiting 5-HT and NA uptake, they also antagonize (among others) the muscarinic acetylcholine-, histamine- (H1), α1 adreno-, and 5-HT2A receptors (54). These medications bring pain relief for a number of patients, but unfortunately, they do not show neuroprotective, or neuroregenerative properties.
Ca2+ channel subunit modulators
Beside duloxetine, the other FDA and EMA approved drug for the treatment of DPN is the anticonvulsant pregabalin, an inhibitor of the voltage-gated calcium channel α2–δ subunit (135). Several clinical studies proved its effectiveness in DPN (50, 51, 141), though there are discrepancies reported in terms of its effectiveness (137). Nevertheless, pregabalin is prescribed on a routine basis for the treatment of DPN. The important role of α2–δ1 in pain transmission is also supported by knockout animal studies in several different models (100, 125). In contrast, elevated expression of α2–δ1 leads to prolonged Ca2+ responses, that is, α2–δ1 shows a strong effect on depolarization-induced intracellular Ca2+ signaling in dorsal root ganglia (DRG) neurons (31, 42). Of note, the α2–δ1 subunit consists of different splice variants with varying affinity for gabapentinoid drugs, and based on a recent study, a low-affinity variant is upregulated in damaged DRG neurons in rats (95). This could be a significant feature behind the inconsistent efficacy of the α2δ ligand drugs in DPN, but further future studies are needed to address this issue.
Combinatorial/opioid-based approaches
In case of inadequate response to first-line therapies, a few other approaches to treat DPN are available. The American Diabetes Association (ADA) and other internationally applied guidelines suggest the use of drug combinations selected from the first-line treatments (
First-line therapy targets and nanotechnology
Monoamine transporter inhibitors, TCAs, and opioids have demonstrated pain relief effectiveness when administered to peripheral targets in preclinical studies. This is due to the presence of their respective targets in the periphery. A limited number of examples where first-line treatments have been formulated into nanoparticles exist in the literature at varied, typically early stages of development. Pham and Kim (132) recently summarized selected examples of poly(lactic-co-glycolic acid) nanoparticles packaged with small-molecule analgesics (e.g., morphine), natural products (curcumin), siRNA, and proteins, designed for both systemic and parenteral administration for treatment of NP (Table 1). Most of these formulations provided extended pain relief that lasted from 2 days to several weeks. However, some of the examples utilized rather invasive routes of administration (intrathecal and intra-hippocampal injection), which may present a significant barrier to further clinical development.
Nanomedicine Examples for Neuropathic Pain Treatment [Adapted from Pham and Kim (132)]
CCI, chronic constriction injury; IL, interleukin; MIA, mono-iodoacetate; NP, neuropathic pain; PLGA, poly(lactic-co-glycolic acid); SNL, spinal nerve ligation; STZ, streptozotocin; TNF, tumor necrosis factor.
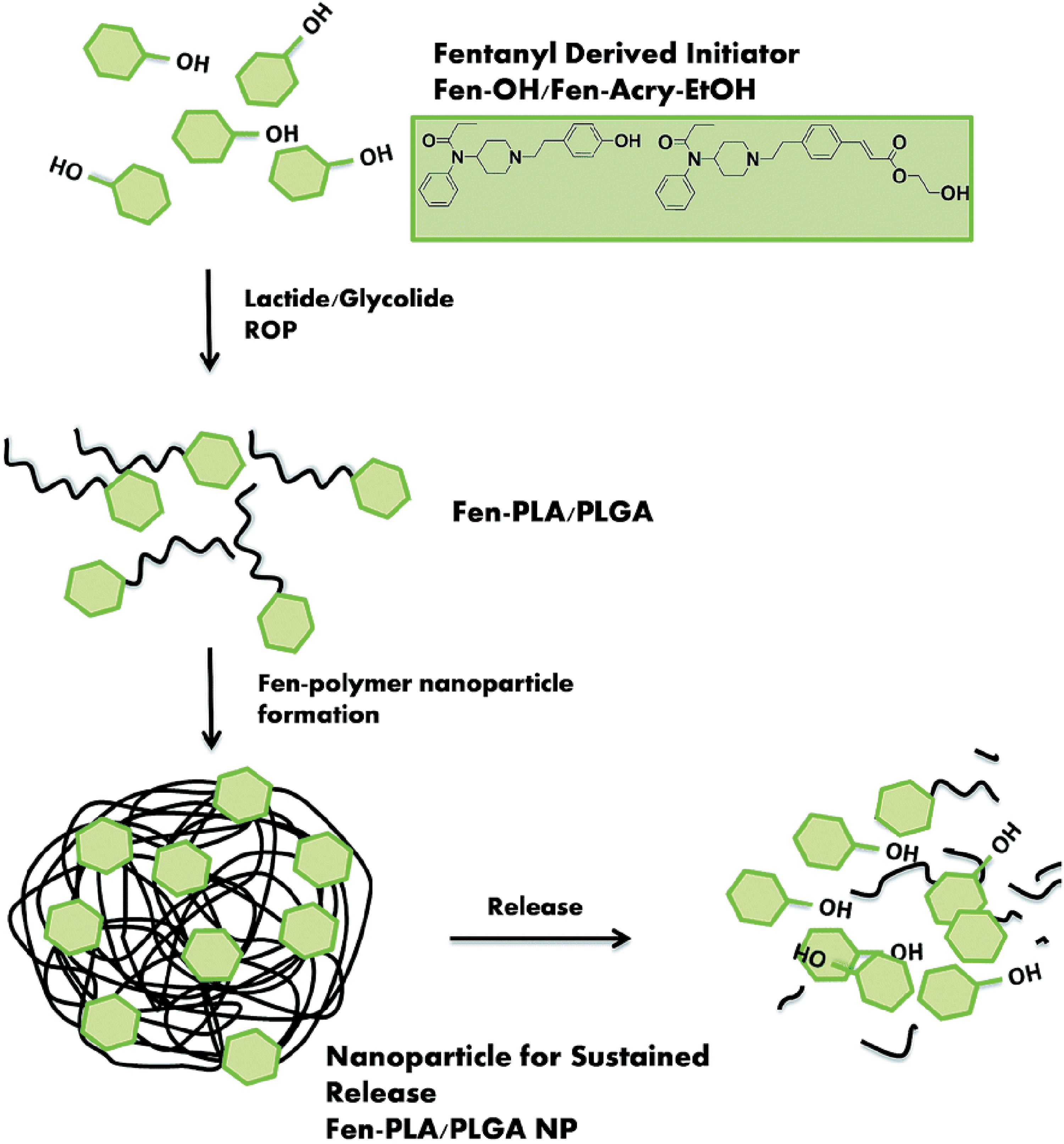
As mentioned, opioids present unique challenges for DPN pain treatment due to dangers of overdose, dependence, and addiction. To overcome these challenges, an elegant synthetic strategy was recently employed where fentanyl-bearing poly(l-lactide) and poly(lactic-co-glycolic acid) (Fen-PLA/PLGA) nanoparticles were synthesized (89). These biocompatible Fen-PLA/PLGA nanoparticles were synthesized as polymeric products by using fentanyl-based initiators to provide controlled opioid release. Further, the researchers used biodegradable microneedles to provide precise and noninvasive administration, avoid potentially toxic drug burst release, setting the stage for much improved patient compliance once technology advances to clinical use. Figure 3 shows the key design features of this innovative nanotechnology.
Based on studies showing peripheral opioid analgesia (10, 93), inhibiting the passage of opioids across the blood–brain barrier and making them exert analgesic effects in the periphery was the motivation for another example of opioid-based nanoparticle design. In this study (55), morphine was conjugated to a hyper-branched polyglycerol via a cleavable linker, which released the drug only in inflamed tissues due to elevated leukocyte esterase activity. The large molecular size and hydrophilicity of this construct prevents the drug passing through the blood–brain barrier, and hence it avoids central side effects. This is one of several examples where chemical modification of existing clinically used opioids was used to suppress side effects and achieve more targeted delivery to the periphery (111).
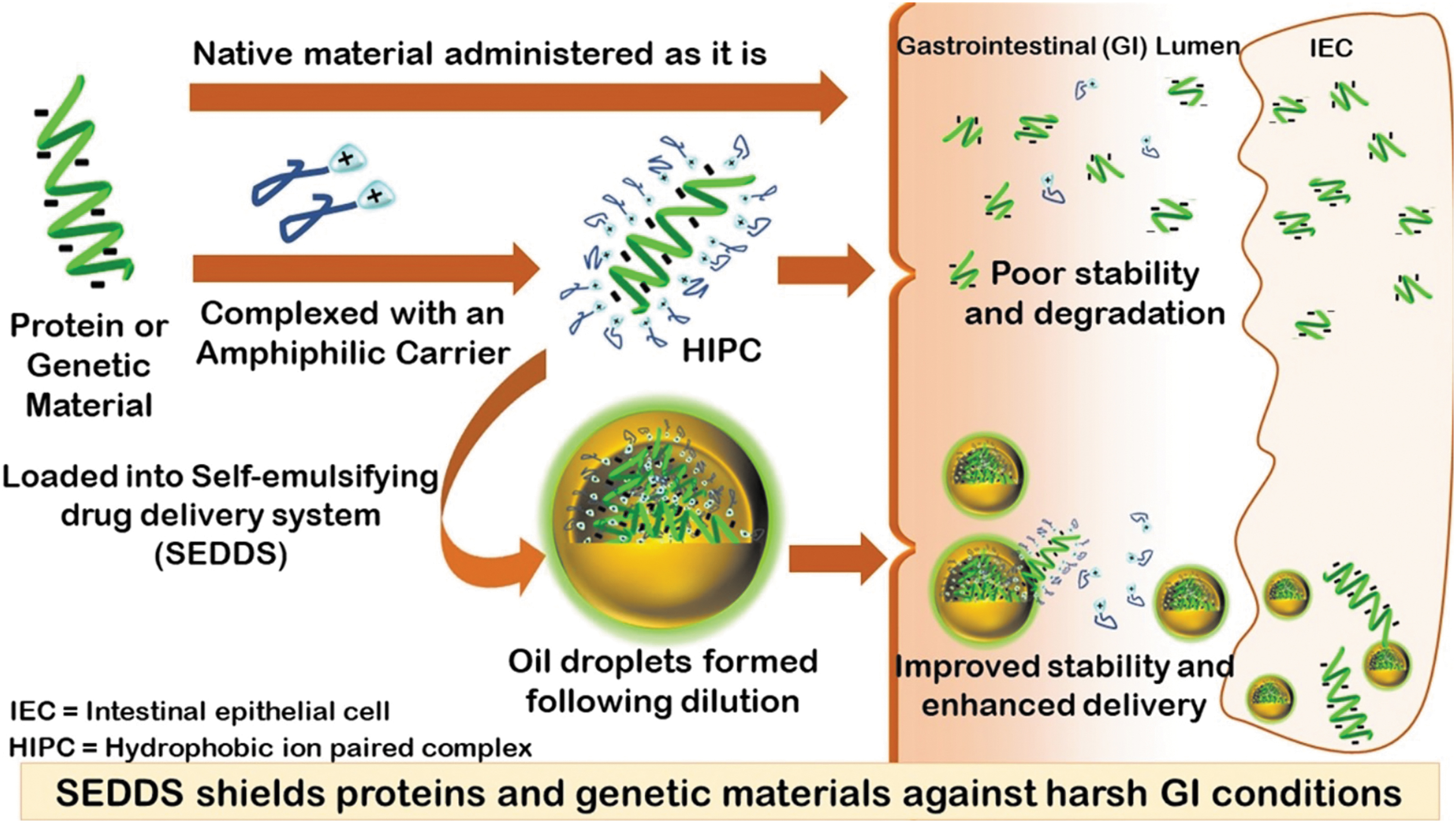
Duloxetine was also formulated into nanoparticles for improving its pharmacokinetic profile, stability, and safety. In one example, duloxetine chloride was loaded into solid lipid nanoparticles (SLNs) prepared by sonication with size less than 100 nm (124). Particles were tested in an animal model of depression, where treatment reduced depressive symptoms in the forced swim test. In a more recent study in rats, duloxetine-loaded solid nanoparticles improved depression-induced changes in forced swim and tail suspension tests after an intraperitoneal injection (139). Solid self-nanoemulsifying drug delivery system (S-SNEDDS) has also been used to improve the pharmacokinetic profile of duloxetine in a rat NP model. In this instance, duloxetine was loaded in combination with the anti-inflammatory and neuroprotective natural product curcumin (91). Self-emulsifying systems, including S-SNEDDS, have unique structural features that allow for self-assembly on oral delivery. Typically, a drug is incorporated into an oil/surfactant/co-solvent mixture and the nanosystem forms in situ in the stomach after disintegration of the capsule carrying the mixed materials (22). Figure 4 shows the typical process for drug delivery in this class of nanomedicines. The advantage of such an approach is that it can provide protection from the harsh stomach environment (low pH, proteases), even for macromolecules such as protein and DNA (1). Currently there are no examples where this technology was applied to NP and DPN, but we posit that this approach could open doors for effective oral delivery of biologics for DPN treatment, using a delivery route that is highly preferred by the majority of patients.
These promising developments offer hope that re-packaging known first-line drugs into nanomedicines could become viable treatment options for DPN pain. Although these examples demonstrate improved drug stability and pharmacokinetic profiles, leading to improved efficacy, they do not utilize nanomedicine to its full potential, since there is no cell-specific targeting. We propose that cell-specific targeting approaches may offer further improvements in the overall efficacy and safety of first-line drug treatments, administered either systemically or locally.
Topical treatments
As an alternative to systemic treatments, other non-first line therapeutic options include topical therapeutics and electrical nerve stimulation (not a subject of the present review). Local analgesia is highly preferred over systemic therapies as it can avoid serious side effects, including the development of dependence and addiction. Capsaicin is a commonly used local agent in different NP states and several studies show significant pain relief after its application, alone or in combination with other therapeutics. Finnerup et al. found that the use of high concentration capsaicin patches is supported by high quality of evidence, although the balance between desirable and undesirable effect is low; consequently, they recommended its use as second-line therapy (48). Some studies also suggest the use of lower concentration capsaicin products (creams or patches), but the literature is not yet conclusive, and further studies would be needed for strong recommendation (48, 61, 170). Capsaicin is a potent agonist of the transient receptor potential vanilloid 1 (TRPV1). TRPV1 is predominantly expressed on unmyelinated C nerve fibers, and it is highly involved in the activation and sensitization of peripheral nociceptors (21). Prolonged use of capsaicin results in desensitization, and therefore the inhibition of pain transmission. TRPV1 (and other TRP channel) ligands might be a novel future group in the treatment of NP (19, 149, 180). Others have reviewed in detail the role of TRP channels in NP (15). Lidocaine (5% strength) patches are also available for the treatment of NP, as a second-line topical strategy. There are studies suggesting significant improvement in DPN (13, 40), yet Finnerup et al. showed in their meta-analysis that the evidence supporting its use in NP is low quality, but with high tolerability and a preferable safety profile (48). Further studies are needed to determine the appropriate place of topically applied local anesthetics in the therapy of peripheral NP diseases (39). Lidocaine, similar to other local anesthetics, is an inhibitor of voltage-gated sodium channels. Therefore, it is an inhibitor of pain transmission via blockade of depolarization and action potential propagation. Subtype-selective sodium channel inhibitors are an area of active research, and they might be a future tool to provide more specificity in the management of NP. The roles of sodium and other ion channels in different NP conditions have been reviewed elsewhere (35, 136).
Nanotechnology for PNS targeting
Though locally targeted therapies are considered safer than systemic therapies, they still lack specificity and targeting, both of which would be critical for effective and personalized DPN treatment. Locally acting nanotechnology-based analgesics have been demonstrated as viable options for the treatment of pain related to PNS injury.
de Araujo et al. recently summarized the current state-of-the-art for lipid-based nanocarriers, such as lipid nanoparticles, nanocrystals, and liposomes developed for local analgesia (34). We present here a few examples that either locally deliver well-known analgesics or engage targets involved in controlling DPN pain and have been evaluated in nerve injury or DPN animal models.
Local anesthetic delivery with lipid-based nanosystems (liposomes, microemulsions, lipid nanoparticles) has been pursued with the goal of prolonged analgesia and improved local efficacy (34). Bupivacaine lipid-polymer hybrid nanoparticles were synthesized by self-assembly and nanoprecipitation. These nanoparticles showed extended analgesic effects in mice, up to 20 h, compared with 1 h with free drug delivered subcutaneously. This extended efficacy was attributed to both improved drug stability and delayed drug release (108). A similar formulation of ropivacaine demonstrated the same improvements in analgesia in both mice and rats (24). A recent study compared tetracaine-loaded poly(
In addition to local anesthetics, natural products have been investigated as local analgesics packaged as nanomedicines. Capsaicin magnetic nanoparticles designed for local pain relief demonstrated much improved efficacy at TRPV1 in vitro and longer analgesic effects in vivo in inflammatory pain rodent models (14). Kopach et al. found persistent analgesia improvements with combining the Na+-channel blocker QX-314 and capsaicin delivered locally by using biodegradable microcapsules (87). The drug combination showed long-lasting analgesic effects and was more effective than capsaicin alone when tested in the complete Freund's adjuvant (CFA) inflammatory pain mouse model.
Although these microcapsules are not nanosystems per se, nor is the CFA pain model directly relevant to DPN, this study is important as it highlights huge untapped potential for local analgesia where a combinatorial approach engaging multiple targets provides long-lasting analgesia. Taken together, these examples indicate that currently well-established molecular targets for local analgesia hold potential for treatment developments using locally delivered nanotechnology. Nanotechnology can improve both efficacy and safety and ultimately provide the long-lasting analgesia needed in patients with DPN.
Non-Neuronal Cell Targets and Their Role in DPN
Targeting Schwann cell dysfunction in DPN
Schwann cells are essential in the maintenance of normal physiological functions of peripheral neuronal cells. The complex functions of Schwann cells include being an insulating barrier to ion movement, hence allowing fast conduction, and physiological response to nerve injury (e.g., phagocytosis of debris) (44). In the event of nerve damage, Schwann cells are able to transition from a myelinating to a regenerating phenotype, while also undergoing proliferation. It was shown in rats that after neuronal injury (sciatic nerve lesion), there is a marked increase in neurotrophic factor expression (such as NGF, BDNF, NT-4, p57NTR, IGF) in Schwann cells distal to the damaged nerve (44, 45). Several studies suggest that Schwann cells support axonal outgrowth by not just physical support (production of basal lamina proteins), but also by producing a wild variety of growth factors. It has been also postulated that neurotrophin binding on the cell surface of Schwann cells (possibly mainly via p57NTR, the low affinity receptor of NGF) might provide neurons with trophic support and chemotactic guidance for regenerating neurons (172).
Hypoxia, hyperglycemia, and increased oxidative stress—the typical attributes of DPN—can contribute both directly and indirectly to Schwann cell dysfunction (44). It is not yet clear whether the necessary transition of Schwann cells from a myelinating to a regenerating phenotype is compromised in diabetes, but it could potentially explain the deficits observed in regeneration. Ekstrom and Tomlinson showed that neuronal regeneration is improved by treatment with ganglioside (glycosphingolipids, which are particularly abundant in the neuronal membrane), presumably via an effect on Schwann cells (46, 178). The study of Jolivalt also suggests the possibility that influencing the function of Schwann cells might improve NP symptoms and nerve regeneration. They investigated the effects of prosaposin, a secreted neurotrophic and myelinotrophic factor (75). Among the several described functions of prosaposin (e.g., inducing neuritogenesis, promoting maturation of muscle), it was previously shown that it promotes the myelinating phenotype of Schwann cells (68). Jolivalt et al. found that prosaposin secretion is reduced after nerve injury, which may contribute to impaired neuroregeneration in diabetic animals (75). The supplementation of a peptide fragment of prosaposin improved regeneration in diabetic rats (improved NP symptoms and nerve regeneration distance and mean axonal diameter of regenerated axons). The mechanism of action of prosaposin is not yet fully understood, but its effect on Schwann cells might be a significant contributor in the improvement of neuronal functions, though further studies are needed to clear this question.
Schwann cells targeting with nanotechnology
The extent to which influencing the function of Schwann cells in DPN will be a future clinical tool is yet unclear, but several recent literature reports demonstrate potential for nanotechnology in modulating Schwann cell behavior to promote nerve regeneration. In a recent study, Razavi et al. (140) applied NGF-loaded chitosan nanoparticles and tested them in vitro for their effects on the differentiation of adipose-derived stem cells into Schwann-like cells. Though limited in scope, this study demonstrates promise for further development where NGF could be delivered specifically to Schwann cells, leading to their modulation in DPN. In another study, liposomes were used to transfect Schwann cells to express neurotrophin-3 (194). The resultant modified Schwann cells were found to promote repair of sciatic nerve defects. In a recent study, a natural product, syringic acid, delivered with mPEG-PLGA-PLLSA-PEAL nanoparticles was found to enhance the proliferative and migratory ability of Schwann cells and contribute to the functional recovery of injured sciatic nerves (102).
It is yet to be confirmed whether these specific approaches would be beneficial for DPN; however, these studies show that Schwann cells are amenable to cellular modulation using nanotechnology. Further, Schwann cells belong to a complex dynamic multicellular vascular structure, the blood–nerve barrier with endoneurial endothelial cells, perineurial cells, pericytes, basement membrane, and axons found to play a significant role in neuropathies (165). This structure is critical defense against chemical and biological insults at the peripheral nerves, which can be utilized for nanotechnology modulation with a single or multi-target approach. As discussed in the following chapters, Schwann cells play a critical role in neuro-immune interactions in DPN, which further strengthens the argument that these cells should be considered as nanotherapeutic targets.
Neuroimmune Interactions in DPN
A rapidly developing area of pain research is the investigation of interactions between neuronal processes and immunological pathways. It has become clear that these interactions are crucial in the development and maintenance of a majority of pain states (12, 148). A plethora of inflammatory mediators are known to sensitize nociceptors and are, therefore, associated with pain and the pathophysiology of DPN (18, 27, 163). In the following subsections, we describe the inflammatory processes and immune cells involved in the development of DPN, and how they could potentially be targeted by novel therapies and diagnostics.
Macrophage infiltration is a well-known phenomenon in different NP conditions (traumatic, metabolic, toxic, degenerative, etc.), as macrophages are the most prominent leukocytes involved in nerve injury and regeneration (29, 59). Tissue-resident macrophages initiate an inflammatory cascade in response to the site of injury and mediate the recruitment of neutrophils and monocytes to the injured area. Monocytes recruited to sites of nerve injury differentiate into additional macrophages that secrete pro-inflammatory mediators, including prostaglandin E2 and cytokines such as tumor necrosis factor-α (TNF-α), interleukin (IL)-1, and IL-6 (99, 186). These inflammatory mediators further amplify the inflammatory response and contribute to the maintenance of neuropathy, pain, and sensory loss (4). Consequently, DPN is associated with macrophage accumulation at site(s) of nerve terminal degeneration (153), at nerve terminals (i.e., in the skin), and in the more proximal nerve (29, 143). In the case of neuropathy, inflammatory-polarized macrophages accumulate in the vicinity of injured nerves to play a role in the regulation of neuroinflammation and function as a common peripheral regulator of NP.
As there is a strong interplay between neurotrophic factors and processes and the immune system, directly aiming at immunological targets might be an interesting novel therapeutic approach in the management of DPN. Kennedy and Zochodne investigated the abnormal macrophage participation in nerve repair in diabetic animals (79). They concluded that impaired macrophage recruitment and impaired macrophage function in diabetes may underlie impaired clearance of debris and therefore sub-optimal regeneration. The question that inevitably rises is whether normalizing macrophage functions could lead to improvement of DPN, and which targets are most likely to achieve this outcome. Here, we will focus on those targets that are currently most accessible for nanomedicine development and targeted drug delivery approaches as potential new DPN treatments.
Macrophage targeting in DPN
Macrophages make appealing nanomedicine targets, due to their early and ongoing roles in the pro-inflammatory process. Based on the tissue environment at the site of injury, macrophages assume one of two heavily studied phenotypes: pro-inflammatory (M1) or anti-inflammatory/pro-healing (M2), though it is likely that these are two extremes on a continuum of potential phenotypes. Classically activated M1 macrophages accumulate in the area of inflammation in the early stage and participate in pro-inflammatory phase where debris clearance is facilitated. These macrophages later polarize into alternatively activated M2 macrophages, relieving inflammation and improving repair by playing a neuroprotective role.
It is critical that M1 macrophages, as well as monocytes and T helper cells (Th), undergo activation at the appropriate times after injury to orchestrate the clearing of damaged tissue or apoptotic cells, stimulation of angiogenesis, and promotion of cell proliferation (37). Targeting macrophages after experimentally induced NP with a macrophage-targeting toxin or blocking macrophage-derived inflammatory cytokines and chemokines with selective inhibitors offers promise by preventing continued NP in rodent models (73, 82, 83, 86). Direct delivery of macrophages as cellular therapy has been shown in experimental studies to enhance cellular repair, but there is a lack of clinical translation associated with the difficulty and potential hazards of administering live cells. As a result, there is a benefit in recruiting endogenous macrophages through target drug delivery over delivering exogenous macrophages (52). Experimentally, the benefit of recruiting macrophages in tissue healing has been demonstrated with multiple methods. One such approach implanted biodegradable vascular grafts seeded with human bone marrow cells, expediting the recruitment of host monocytes and macrophages with the secretion of monocyte chemoattractant protein-1 (142). In addition, encapsulated gelatin/lactic acid-based micelles containing the macrophage recruitment agent SEW2871 in gelatin hydrogels promoted tissue healing in a rat model (84).
The key macrophage receptor regulating M1–M2 conversion is CD163. CD163 scavenges hemoglobin:haptoglobin complexes, which leads to the nuclear translocation of the transcription factor Nrf2 (NF-E2-related factor 2), upregulation of heme oxygenase (HO)-1 cytoprotective protein, and release of the anti-inflammatory cytokine IL-10 (96). Data suggest that a range of natural and synthetic anti-oxidants may drive M1 to M2 switching through Nrf2/HO-1, leading to improved kidney function in diabetes (74, 96, 121). Therefore, Nrf2, which is the transcriptional regulator upstream of HO-1, might be a novel target in diabetes management and DPN. Although Nrf2 activation has been mainly investigated in kidney and heart diseases during diabetes, Nrf2 agonists showed mixed results in clinical trials (with adverse events including heart failure and fluid overload (26, 36, 130). Further study is needed to establish whether M1–M2 transition induced by the Nrf2 pathway could be beneficial in DPN management.
Toll-like receptor (TLR) signaling is also crucial for appropriate activation of macrophages. Through the stimulation of TLRs, macrophages produce pro-inflammatory cytokines and chemokines. Co-stimulatory molecules that are critical for adaptive immune responses are also induced, and they regulate their own transcription (169). Mohammad et al. reported that TLR expression/signaling is disrupted in diabetes (e.g., expression of TLR3 and TLR5 was significantly higher in diabetic animals), which may contribute to the impaired macrophage function (118). The exact role of these changes in the future therapy of diabetes and DPN needs to be further investigated. Given that the hyperglycemic diabetic milieu is known to drive pro-inflammatory polarization of macrophages (60, 174), it is plausible to suggest that measures designed to reduce this pro-inflammatory polarization will have a positive impact on nerve regeneration in diabetes. Macrophages are an important source of cytokines (IL-1, IL-6, IL-12, TNF-α), which are necessary for driving regeneration (80). Kamijo et al. investigated the process of Wallerian degeneration in spontaneously diabetic biobreeding Worcester rats (78). They showed a marked delay in Wallerian degeneration and decreased endoneurial macrophage activity in diabetic nerves, indicating their interrelation. This study gives evidence that macrophage influx and clearance activity is impaired in diabetes (decreased phagocytic activity by macrophages). Interestingly, these changes were not demonstrated in aldose reductase inhibitor (ARI) treated diabetic rats. How exactly ARI treatment improves macrophage function is not clear, but it seems that activation of the polyol pathway may influence both intrinsic and extrinsic mechanisms governing nerve fiber regeneration. Other articles provide further details about ARI treatment and how it might be utilized for the treatment of DPN (56, 77, 178).
Targeting macrophages with nanotechnology for the management of NP
Considering the earlier findings together, it is becoming clearer and clearer that macrophages play a highly significant role in the development of NP. Therefore, agents aiming at influencing their activity represent promising therapeutic candidates in the management of NP diseases, including DPN. The rapidly developing nanotechnologies available to target macrophages also offer a promising novel approach to specifically influence macrophages, hence alleviating DPN. The ability of macrophage-targeted nanomedicine to selectively and specifically influence macrophage behavior in vivo lies in their ability to uptake nanoparticles by endocytosis and/or phagocytosis (52). Nanoparticle uptake can be facilitated by a variety of targeting and surface modification strategies. Two basic strategies exist for targeting macrophages in the treatment of inflammation, with nanoparticles revolving around the existence of macrophages in two distinctive phenotypes. One such method involves macrophage depletion by nanoparticle-loaded drugs, whereas the other employs re-education by nanoparticles carrying specific cytokines to the relevant microenvironment to skew M1-polarized macrophages to M2 (70). Although achievable by using nanoparticle-based approaches, macrophage depletion is not an ideal strategy due to potential immunosuppressive effects and creating defects in infection defense. For that reason, macrophage-directed drug delivery aimed at suppression of intracellular pro-inflammatory pathways using small molecules or siRNA is a safer and more effective alternative. Modulation of macrophage phenotype with nanoparticles is an attractive therapeutic strategy, and a recent review highlights the new developments in macrophage-targeted polymeric nanoparticles (5). These theranostic nanoemulsions (NEs) allow visualization of macrophage infiltration as a surrogate measure of inflammation by multimodal imaging strategies, combining both fluorescence and magnetic resonance imaging.
Recently, we have developed and evaluated theranostic NEs for targeting macrophages via an intravenous injection in a rodent NP model with the goal of suppressing injury-induced neuroinflammation (144, 181). The NEs are widely used colloidal dispersions in pharmaceutical applications as a biocompatible strategy to increase bioavailability of poorly soluble or lipophilic drugs (88, 115, 138, 147, 151). The NEs can be designed to have multimodal imaging capability, as well as exceptional colloidal and photo stability allowing incorporation into gels and can be produced on an industrial scale (103). We have established a robust multimodal theranostic perfluorocarbon nanoemulsion (PFC-NE) platform that simultaneously delivers a therapeutic agent and monitors responsiveness to therapy, integrating therapeutic and diagnostic functionalities (66, 67, 72, 103, 123, 126 –128). The novel colloidal nanosystems, PFC-NEs are triphasic because they consist of: perfluoropolyether (fluorous phase), hydrocarbon oil (synthetic or natural/organic phase), and water (aqueous) phase. These small-sized (100–140 nm) PFC-NEs are ideally suited for theranostics, due to their surface-area-to-volume ratio that allows for effective loading of drugs and multiple imaging agents, and can provide sustained release, and the ability to functionalize the surface with cell targeting ligands (macrophage targeting ligands) (67, 72, 126).
Macrophages at sites of injury, whether hematogenous or resident, produce numerous pro-inflammatory mediators (e.g., chemokines, cytokines, cyclooxygenase-2 [COX-2] induced prostaglandins) (134). COX-2 levels are upregulated after neuronal injury (43, 168), and persistent overexpression of COX-2 in infiltrating macrophages has been strongly linked to chronic (late phase) neuroinflammation and NP in both rats and humans (43, 109, 110, 168). Our prior and ongoing work confirms that COX-2 inhibition in monocyte-derived (hematogenous) macrophages by targeted delivery of COX-2 inhibitors (e.g., celecoxib) can reduce tactile allodynia to baseline levels in a rat model of NP (72). To effectively suppress COX-2 in nerve injury infiltrating macrophages, we used nanoemulsions designed for selective targeting of macrophages with an extended and prolonged drug release profile. COX-2 inhibitor (celecoxib [CXB]) loaded nanoemulsions (CXB-NEs) are designed to delay intracellular drug release by 3–5 days. This delay in COX-2 inhibition is critical to allow M1 macrophages to support initial injury debris clearance and prepare the area for neuronal regeneration (20). Muzamil Saleem et al. have shown that targeted macrophage COX-2 inhibition with celecoxib loaded nanomedicine (CXB-NE) leads to a phenotype shift from M1 to M2 at the site of neuronal injury in an NP model in rats, whereas there was no response to drug-free control (drug free NE), Figure 5 (120). In a related study in an inflammatory pain mouse model, we have also demonstrated that a single dose of macrophage-specific COX-2 inhibiting nanomedicine provides extended reversal of mechanical hypersensitivity—over 30 days in males and 10 days in females (104).
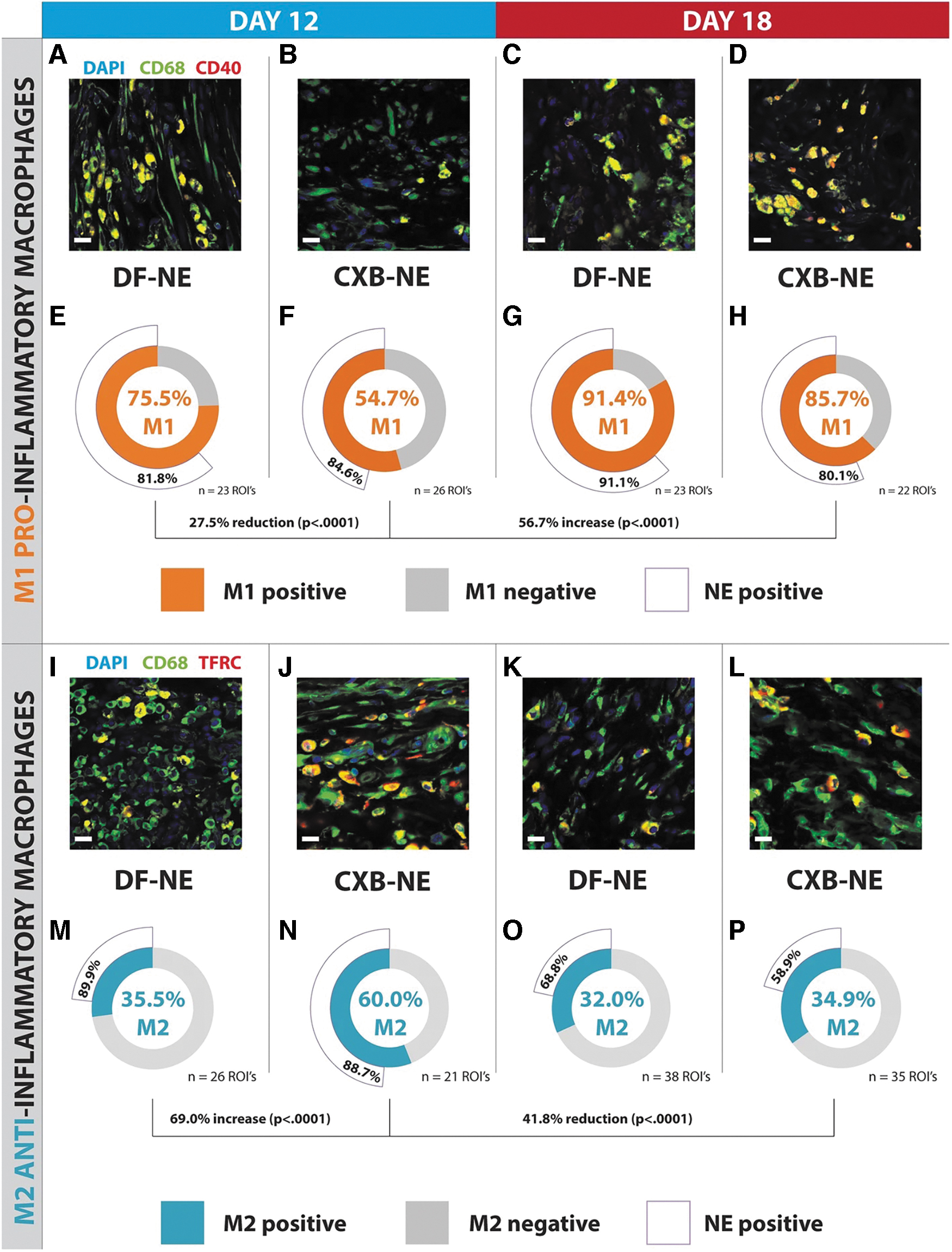
In an in vitro study (3), human macrophages were transfected to overexpress CD163 by using polyamine nanoparticles. The CD163-overexpressing cells exhibited reduced levels of pro-inflammatory cytokines and increased levels of anti-inflammatory cytokines after stimulation with lipopolysaccharide (LPS), further suggesting the critical role of CD163 in macrophage polarization. A more recent study by the same group showed differences between macrophages obtained from patients with DPN and healthy donors. Briefly, they transfected macrophages with CD163 by using polyamine nanoparticles and then studied their response in vitro to LPS challenge. They found that DPN patient-derived macrophages showed a pro-inflammatory phenotype in an unstimulated state, which was reversed by CD163 transfection (4). Whether any of these macrophage-targeted nanotechnology approaches may be applicable to DPN is currently unknown and subject to further investigation.
Angiotensin II signaling in neuroimmune interactions
We have recently reviewed the complex role of the renin–angiotensin system in NP development (8). Consistent with a pro-inflammatory role for angiotensin II (Ang II) signaling, it has been suggested that angiotensin converting enzyme (ACE) inhibitors or angiotensin receptor blockers can be anti-inflammatory (145). It has also been demonstrated that nerve injury is associated with increased Ang II production. This “local” (i.e., non-circulatory) production of Ang II represents a novel component of inflammation, neuropathy, and pain (153, 155, 156). Importantly, clinical trials of Ang II receptor blockers for pain relief have shown promising results (62). Importantly, the AT2R signaling within the immune system is involved in NP, and Ang II receptor positive (AT2R+) macrophages at the site of nerve injury are required in the process. (152, 153, 155, 156). These effects are consistent with the hypothesis that Ang II stimulates the production of growth factors, cytokines, and chemokines (184), an observation consistent with the induction of inflammation-related genes seen in transcript analysis in other disease models (162).
From a nanomedicine design perspective, it is important to emphasize the finding that Ang II exerts a chemotactic effect on macrophages when injected into hindpaw skin (23, 85). Further, data suggest that Ang II production at sites of nerve injury influences macrophage infiltration. We recently demonstrated the expression of Ang II receptor genes in tissue biopsies and functional expression in primary mouse macrophages, and mouse and human cell lines in vitro (153, 155). At the same time, it was shown that exogenous Ang II (at sub-nanomole quantities) is sufficient to induce pain hypersensitivity.
These data support the hypothesis that macrophages respond to pain-promoting Ang II released by injured nerves in DPN. We have shown that Ang II can drive macrophage infiltration, and macrophage density is elevated in the vicinity of injured nerves in mice and humans (Fig. 6) (153). Accordingly, the inhibition of Ang II receptor signaling reduces mechanical and cold hypersensitivity by targeting infiltrating macrophages. We have also shown that Ang II acts via the type-2 Ang II receptor (AT2R) on macrophages to produce reactive oxygen species (ROS), which sensitize adjacent sensory nerves. Key experimental findings were recapitulated in human neuron cultures and patient samples (153, 155). Antioxidant-based nanomedicines are therefore an attractive target, as detailed in previous sections for activators of the antioxidant regulator gene Nrf2. In addition, cerium oxide nanoparticles exhibit antioxidant properties and have demonstrated a reduction in ROS-induced neuronal loss in models of Alzheimer's and multiple sclerosis. However, their efficacy in peripheral neuropathies has not been tested (92). Nevertheless, they may offer ancillary benefits for DPN in the form of accelerated wound healing (25).
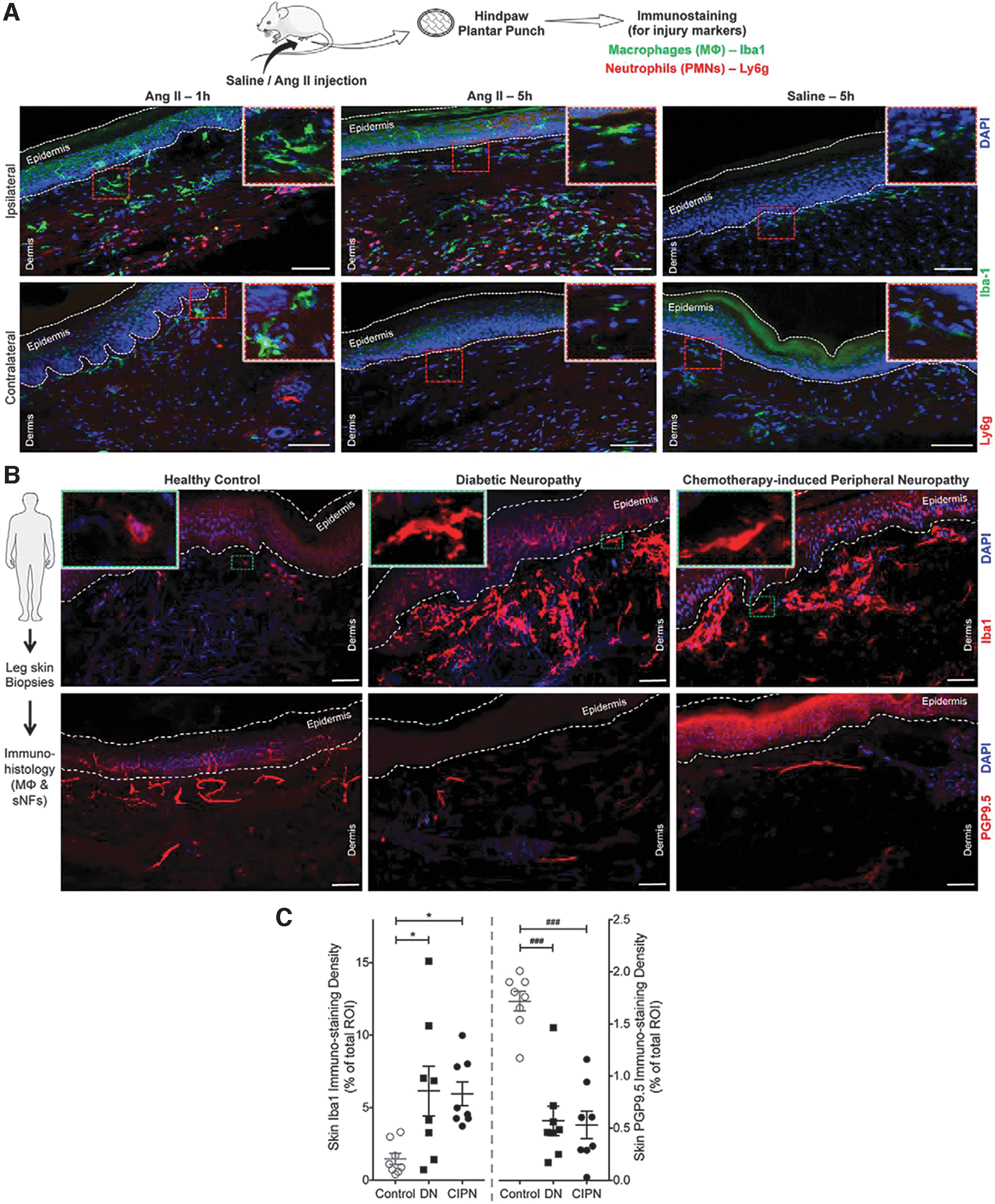
Systemic or local injection of an AT2R antagonist strongly attenuated mechanical and cold hypersensitivity in a mouse model of traumatic neuropathy (153, 155). These results suggest that long-term inhibition of local Ang II signaling could reverse sensorimotor disruption in DPN. High-fat diet feeding to wild-type mice is used as a model of the early stages of type 2 diabetes (28). The resultant diet-induced obese mice show an increase in tactile hypersensitivity similar to that seen in type 2 diabetes, a finding that has been independently verified (143, 164). Prior reports have suggested a beneficial role for ACE inhibitors in diabetic neuropathy, but the beneficial effect of such an approach was ascribed to improvements in endoneurial blood flow rather than neuroimmune crosstalk (113). These data indicate that Ang II generated in the vicinity of neuronal injury potentially contributes to pain sensitivity in diabetic neuropathy and similar disease states.
Other (Non-Macrophage) Immune Cell Types Involvement in DPN
Our understanding of the complex machinery of neuroimmune interactions in the development of NP continues to grow and is still far from being fully understood. Besides the tremendous changes caused by prominent macrophage infiltration, there are now clear signs of the contribution of other immune cells to the development of pain. Although macrophages seem to be the most promising targets for the future treatment of DPN (112), other immune cell types are also involved.
Yao et al. showed that in patients with trigeminal neuralgia, neutrophils, lymphocytes, monocytes, platelets, albumin, as well as the neutrophil/lymphocyte ratio and monocyte/lymphocyte ratio were all elevated (188). In the study of Sutherland, CD4+ and CD8+ T cells and CD20+ B cells were localized in human ganglia of patients suffering from post-herpetic neuralgia, years after the viral infection (167). In this section, we describe findings suggesting a role of different immune cells in the development and maybe the future management of NP.
During the complex process of Wallerian degeneration induced by nerve damage, Schwann cells release numerous proinflammatory cytokines and chemokines (e.g., IL-1α, IL-1β, IL-6, TNF-α, LIF, MCP-1) (191). The release of these inflammatory mediators results in phospholipase A2 upregulation by Schwann cells, which, in turn, generates lysophosphatidylcholine (LPC) (53). The LPC acts as an “eat me” signal for macrophages and triggers the classical complement pathway, hence the influx of neutrophils. Neutrophils are the earliest inflammatory cells that infiltrate damaged nerves and tissues, with a peak usually after 3 days of injury. Neutrophils release different inflammatory mediators as well (such as TNF-α, IL-1β, IL-2, IL-6, bradykinin, etc.), leading to hyperalgesia and the recruitment of other immune cells, such as macrophages (101). About a decade ago in the study of Perkins and Tracey, it was shown that neutrophils play an essential role in the early development of hyperalgesia after peripheral nerve injury (131). The administration of neutralizing antibodies against neutrophils alleviated the pain symptoms within the first 48 h after injury but proved to be ineffective after this period. There are recent studies showing the contribution of neutrophils in the development of mechanical allodynia in an experimental model of multiple sclerosis, but their exact role in the development of different neuropathies and the emergence of pain symptoms needs further investigation (63, 189).
Current findings shed light on the role of different T cell subtypes in the development of autoimmunological neuropathies, rather than DPN (191). As an example, CD8+ T cells seem to be the main contributors to the development of Guillain–Barre syndrome (GBS) and chronic inflammatory demyelinating polyneuropathy (CIDP) and Schwann cells seem to be involved in these complex mechanisms as well (187, 191). This indicates that besides macrophages, Schwann cells are capable of interacting with these cell types. Indeed, Tang et al. investigated the interactions under diabetic conditions. They found that Schwann cells under high glucose level upregulate the expression of different C–X–C motif chemokine ligands, which leads to the attraction and activation of cytotoxic CD8+ T cells, resulting in the release of different inflammatory modulators (171). There are additional studies showing that Schwann cells interact with natural killer T cells: In patients suffering from CIDP and GBS, markers of antigen-presenting cells, as well as co-stimulatory molecules were detected on Schwann cells, indicating the capability of antigen presentation by these cells (114, 116). On the other hand, Krukowski et al. found that CD8+ T cells increase DRG IL-10 receptor expression, and consequently they are required for the resolution of chemotherapy-induced NP in mice (90). These somewhat contradictory results might be explained based on the variation in the damaging insult and subsequent nerve injury response. This evidence further strengthens the strong interplay between immune and neuronal cells and should be further investigated in DPN models. Hartlehnert et al. showed how Th cells promote the development post-traumatic nerve inflammation and NP (64). They proved the capability of Schwann cells to express class II major histocompatibility complexes (MHC-II); hence, they might act as antigen presenting cells (APCs), that is, they described them as potentially conditional APCs. They found that the deletion of MHC-II in myelinating Schwann cells reduces both mechanical and thermal hyperalgesia. These beneficial changes were accompanied by a reduction of intraneural CD4+ T cells. These results indicate that Schwann cells modulate local immune responses in the peripheral nerves, as well as the involvement of T cells in neuroimmune interactions accompanying the development of pain.
Davoli-Ferreira et al. investigated the role of regulatory T cells (Treg) in NP by using a partial sciatic nerve ligation model in mice (33). They showed that these cells infiltrate and proliferate in the site of peripheral nerve injury. Interestingly, they found that Treg counteracts NP development, possibly via IL-10 signaling and the inhibition of the CD4 response. There are several other studies showing the beneficial role of Treg in the attenuation of NP as well (6, 97). Most recently, Ding et al. showed that the depletion of Treg counteracts the protection of NP mediated by gut microbiota changes, and it results in an increase of IFN-γ-producing Th1 cell infiltration in the spinal cord (41). Based on these results, boosting Treg functions and the inhibition of CD4 response might be beneficially utilized in the treatment of NP diseases.
Neutrophil and T cell targeted nanomedicine
Biomaterials and nanosystems designed specifically to engage immune cells beyond macrophages have been extensively investigated in recent years, and they are thoroughly reviewed elsewhere (98). Here, we will highlight a few examples of nanomedicines targeted to T cells and neutrophils. As expected, most neutrophil targeting examples have been evaluated in tumor immunology contexts and for the purpose of improving cancer therapy. Polypoly(sialic acid) decorated liposomes were shown to be specifically taken up by neutrophils in vivo due to the presence of sialic acid receptors on the neutrophil cell membrane (107). Wang et al. successfully inactivated the pro-inflammatory functions of neutrophils with albumin-based nanoparticles loaded with the tyrosine kinase inhibitor piceatannol, resulting in reduced vascular inflammation (183). Another study employed a different approach, wherein instead of targeting neutrophils, the nanoparticles were formulated with neutrophil membrane surrounding the polymeric core. These unique neutrophil-nanoparticles showed remarkable anti-inflammatory effects in an arthritis model (190).
T cells are, by far, the most challenging cells to effectively target in vivo with specific nanoparticles for both therapeutic intervention and diagnostics. However, successful examples of modulating T cell behavior in living subjects by T cell targeted nanosystems have been reported (161). Briefly, to achieve gene delivery into T cells, T cell-targeting anti-CD3e f(ab′)2 fragments were coupled to the surface of biodegradable poly(β-amino ester)-based nanoparticles. This is a typical approach for targeting T cells where antibodies or antibody fragments are used to achieve target specificity, as opposed to macrophages where simpler surface modifications (e.g., size, charge, PEG-ylation, etc.) and small-molecule or peptide incorporation is sufficient to secure targeting and intracellular uptake. Maleimide-thiol chemistry was recently used to conjugate anti-CD4 f(ab′) antibody fragments to the surface of PEG-ylated PLGA nanoparticles to target CD4+ T cells and deliver eggmanone, a specific Hh inhibitor, as a T cell directed immunomodulation approach (65). The use of these technologies in DPN may currently seem like a distant prospect. However, as our knowledge of the role of T cells in DPN increases and new intracellular targets are discovered, having access to already established T cell targeted nanoparticles is useful for two reasons: (i) We can utilize nanosystems to achieve very specific T cell modulation with pathway inhibitors or other types of probes that can lead to new mechanistic insights; and (ii) targeting T cells may open new therapeutic opportunities for DPN.
Conclusions
Though the pathophysiological complexity of DPN represents a formidable therapeutic challenge, the recent increase in our knowledge and appreciation of neuro-immune interactions offers a suite of new potential targets to modulate pain and neuropathy. These advances have been accompanied by the development of multiple novel nanomedicine-based agents, offering new capabilities and specificity of drug delivery. Uniting these two rapidly progressing fields of research raises the possibility that new treatment options to relieve pain and perhaps promote nerve regeneration will be generated, in DPN and other NP states. This is particularly true for cell-specific targeting approaches, which appear to offer the greatest potential.
Footnotes
Authors' Contributions
M.B.: Writing—Original draft; Conceptualization. J.M.J.: Writing—Original draft; Visualization; Conceptualization. A.J.S.: Writing—Review and editing; Visualization; Conceptualization. All authors read and approved the final article.
Author Disclosure Statement
No competing financial interests exist.
Funding Information
A.J.S. is supported by funds from The University of Texas MD Anderson Cancer Center, the Rita Allen Foundation and NIH grant UG3NS116929. A.J.S. and J.M.J. are supported by the Department of Defense partnering awards W81XWH-20-1-0276 and W81XWH-20-1-0277.