
Abstract
Significance:
Hypertension is a multifactorial disease ensuing from the continuous challenge imposed by several risk factors on the cardiovascular system. Classically known pathophysiological alterations associated with hypertension comprise neurogenic mechanisms dysregulating the autonomic nervous system (ANS), vascular dysfunction, and excessive activation of the renin angiotensin system. During the past few years, a considerable number of studies indicated that immune activation and inflammation also have an important role in the onset and maintenance of hypertension.
Critical Issues:
On these premises, it has been necessary to reconsider the pathophysiological mechanisms underlying hypertension development, taking into account the potential interactions established between classically known determinants of high blood pressure and the immune system.
Recent Advances:
Interestingly, central nervous system areas controlling cardiovascular functions are enriched with Angiotensin II receptors. Observations showing that these brain areas are crucial for mediating peripheral ANS and immune responses were suggestive of a critical role of neuroimmune interactions in hypertension. In fact, the ANS, characterized by an intricate network of afferent and efferent fibers, represents an intermediate between the brain and peripheral responses that are essential for blood pressure regulation.
Future Directions:
In this review, we will summarize studies showing how specific brain areas can modulate immune responses that are involved in hypertension. Antioxid. Redox Signal. 35, 1515–1530.
Introduction
Hypertension is one of the most alarming cardiovascular diseases (CVDs) with a continuously increasing number of patients worldwide. Although many drugs are available for therapeutic purposes, the pathogenesis remains unknown in the majority of patients. Hypertension results from one or multiple alterations of the cardiovascular system and, in turn, may disrupt the appropriate functionality of organs typically targeted by high blood pressure—such as kidneys, vasculature, heart, and brain—leading to severe clinical manifestations (66). Dysfunctions of these organs may remain silent for years, allowing the disease to progress in an uncontrolled manner. For this reason, research has been focused in understanding the pathogenetic mechanisms underlying the so called “end-organ damage” and preventing the evolution of organ failure (82).
One of the most common determinants of blood pressure levels is the renin angiotensin system (RAS). As such, an imbalance in RAS activity is typically associated with hypertension in humans and experimental models. A pharmacological increase in the circulating levels of the hormone peptide Angiotensin II (AngII) is a typical model that reproduces hypertension in mice (48). Biological activities of AngII are multiple and are related to the various locations of AngII receptors (ATRs), which are, in fact, expressed by different cell types in peripheral organs as well as by distinct nuclei of the forebrain and brainstem in the central nervous system (CNS) (119).
Further, it has been clearly demonstrated that innate and adaptive immune cells are activated by hypertensive stimuli, promoting their migration toward target organs where they contribute to the progressive tissue damage and consequent organ dysfunctionality. This event is a typical hallmark of hypertension (10, 28, 85). An additional level of complexity is provided by the critical regulatory function exerted by the autonomic nervous system (ANS) on the cardiovascular system's physiology. On this premise, a dysregulation in ANS activity is often associated with the onset and progression of hypertension (33). While typically recognized as a regulator of cardiovascular function, the ANS has been characterized by our studies and others as a critical modulator of immune responses recruited by hypertensive stimuli and is considered relevant for the development of the related target organ damage (9, 12, 90).
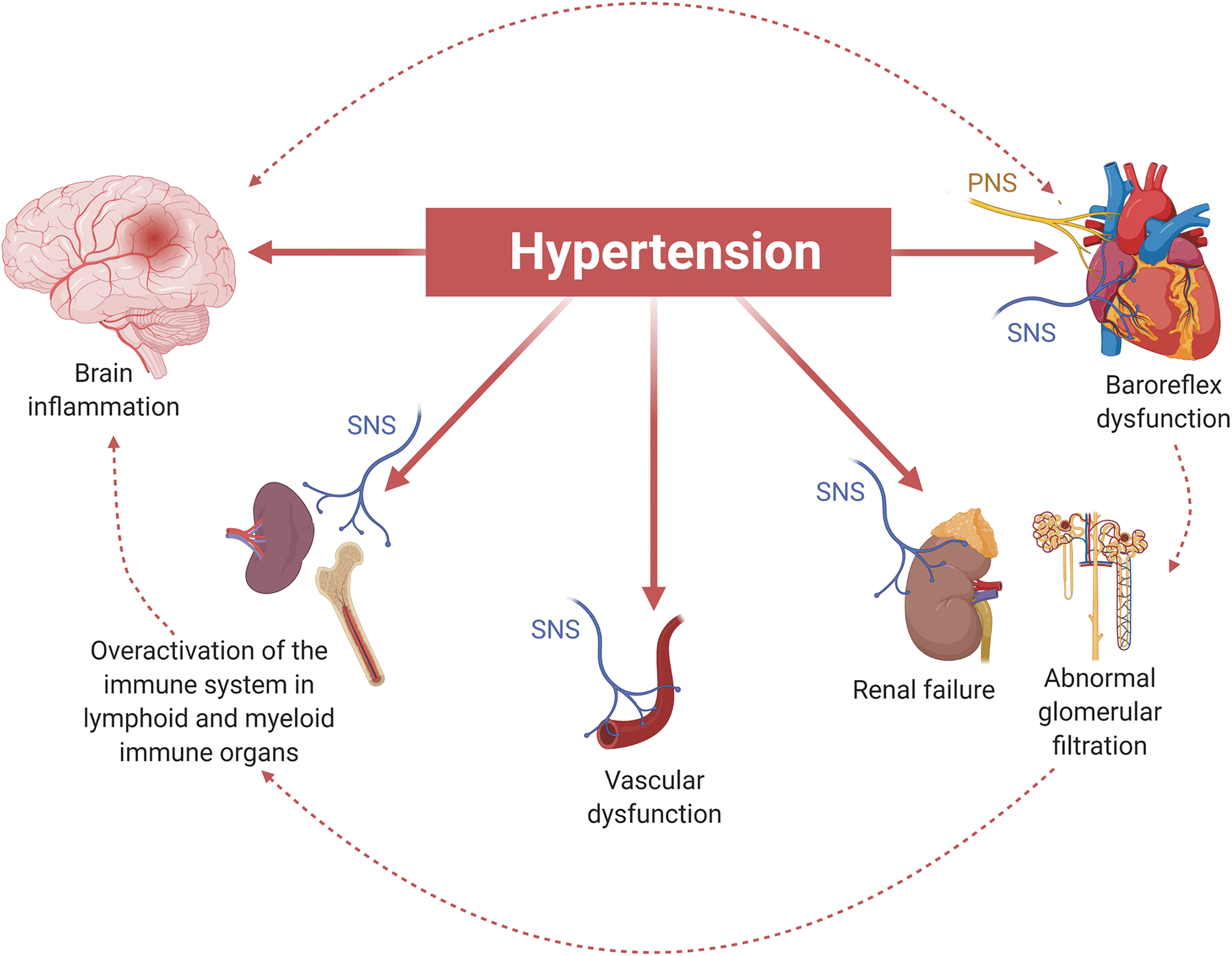
Here, we will provide a revision of the studies that highlight the importance of these critical regulators of blood pressure and their mutual connections (Fig. 1).
Brain Areas Controlling Cardiovascular Function
Circumventricular organs (CVOs) are distinct brain nuclei enriched with ATRs, mediating the effects of AngII and contributing to blood pressure regulation and fluid homeostasis (22, 68). They are characterized by fenestrated capillaries lacking the blood–brain barrier, making neurons in direct communication with substances circulating in the blood (5).
The subfornical organ (SFO) is one of the CVOs anatomically located where the lateral ventricles and the third ventricle converge. The SFO is composed by two distinct areas fulfilling different functions, the core and the peripheral sections (16, 51). The core section is vascularized and communicates with the cerebrospinal fluid through specialized ependymal cells called tanycytes. In turn, these latter ones lack cilia and have processes extending to the perivascular space of SFO capillaries (16, 26). Thus, tanycytes link the cerebral ventricles to the vascular unit, forming an interface between the cerebrospinal fluid and the blood and involving the regulation of fluid exchanges among blood, brain, and cerebrospinal fluid, within the SFO and the other CVOs (67). The peripheral section of the SFO is deprived of vasculature but responds to signals coming from the cerebrospinal fluid (55). The SFO is also a brain area that is fundamental for the organization of neuronal connections with other brain nuclei. In fact, the rostral portion contains efferent projections directed to the median preoptic nucleus (MnPO), the organum vasculosum of the lamina terminalis (OVLT), the supraoptic nucleus (SON), the lateral preoptic area (PA), the paraventricular nucleus of the hypothalamus (PVN), and the suprachiasmatic nucleus (SCN) (102).
Also, together with the PVN, the SFO establishes more specialized connections. The SFO projects to the magnocellular portion of the PVN through efferent neurons of the core, and to the parvocellular neurons of PVN through neurons of the peripheral section (59). In turn, sympathetic efferent projections of the PVN parvocellular neurons are connected to sympathetic neurons of the rostral ventrolateral medulla (RVLM) (18, 62). Defined as the pressor area of the medulla, the RVLM is responsible for controlling the sympathetic activity associated with cardiovascular functions (21, 76). Interestingly, PVN neurons project to the complex of the brainstem composed by the nucleus of the solitary tract (NTS) and the dorsal motor nucleus of the vagus (DMV) (56), implicating regulatory activities of the cardiovascular system established by the modulation of the vagus nerve (58). Thus, the PVN can be considered a crucial hub by which neuronal signals depart to innervate peripheral organs of the cardiovascular system, through the intermediation of the sympathetic nervous system (SNS; by the RVLM) and the parasympathetic nervous system (PNS; by the NTS/DMV complex) (Fig. 2, Table 1).
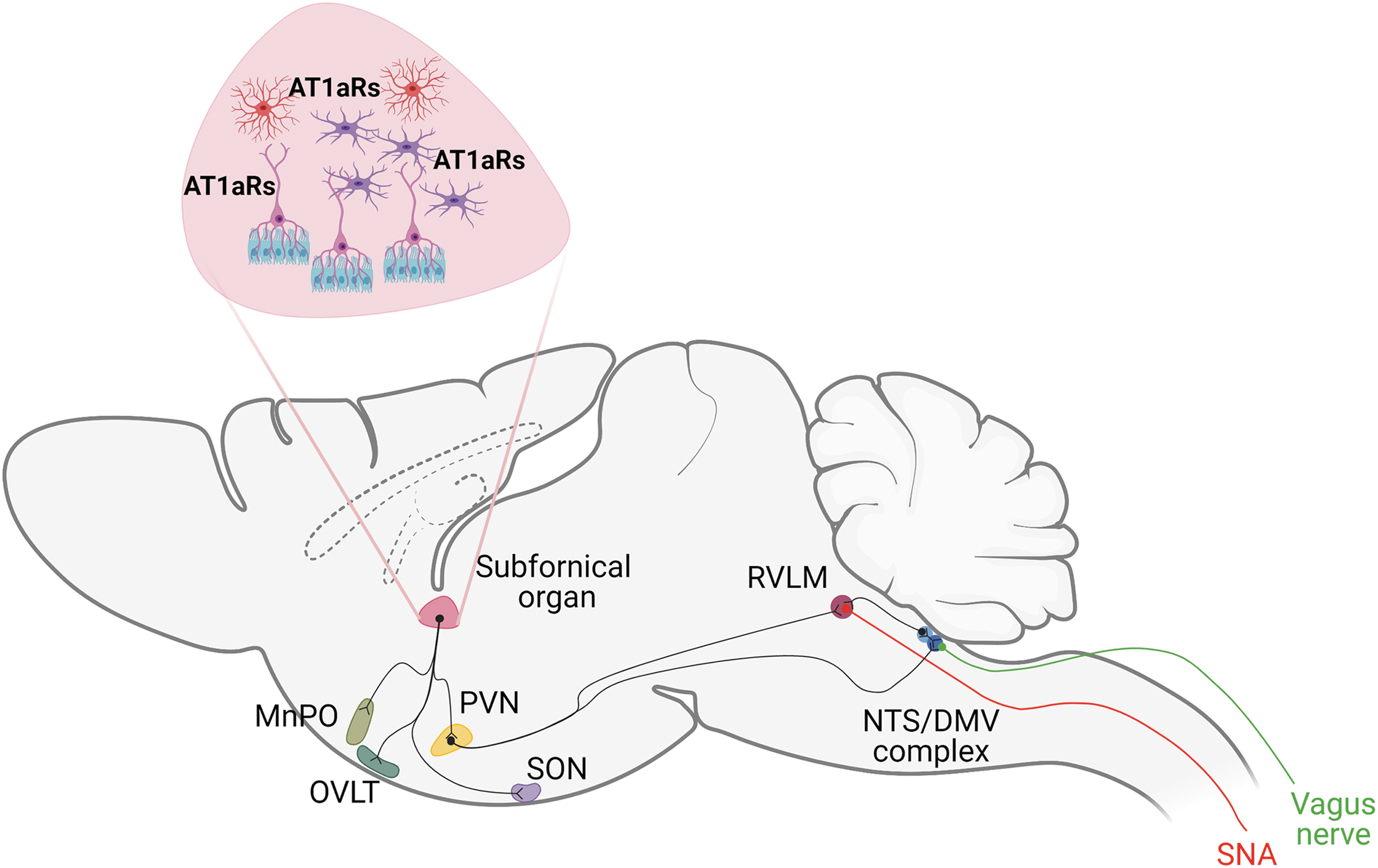
Abbreviations of the Most Important Brain Areas Controlling Cardiovascular Function
Expression of ATRs in Brain Areas Involved in Hypertension
The RAS is one of the principal pathways involved in the development of hypertension (113). Molecular mediators of the RAS are the angiotensinogen, renin, and angiotensin converting enzyme (ACE) that participate in the production of final biologically active peptides, AngI, AngII, AngIII, Ang(1 –7), and AngIV (37). These peptides act through G-protein-coupled receptors (GPCRs) with a wide distribution in many tissues of the cardiovascular system: (i) the AngII type 1 receptor (AT1Rs), (ii) the AngII type 2 receptor (AT2Rs), (iii) the MAS receptor, and (iv) the type II trans-membrane zinc protein receptor with the AngIV binding site (57).
Elevated levels of circulating AngII are critically involved in hypertension, promoting various effects depending on the cell type expressing AT1Rs and/or AT2Rs in different tissues. It is generally considered that AT1Rs and AT2Rs have opposing actions (8, 30, 83, 89), with AT1Rs mediating the “harmful” effects of AngII, as vasoconstriction, cell proliferation, and fibrosis, whereas AT2Rs counterbalance AT1Rs activity by promoting vasodilatation, apoptosis, and antiproliferative effects (19, 38).
The anatomical distribution of AT1Rs and AT2Rs in the brain is particularly important in hypertension, as it contributes toward determining the level of activation of brain areas controlling specific functions of the cardiovascular system (16). Until some decades ago, it was known that, in mice and rats, AT1Rs are enriched in brain areas designated to blood pressure control and fluid balance, whereas AT2Rs are mainly expressed in regions that indirectly communicate with centers controlling the cardiovascular system (69).
AngII has the same affinity for AT1Rs and AT2Rs and, in fact, by cloning the sequence of both receptors, it has been shown that they have the same structure, composed of seven transmembrane domains (69). Hence, many studies provided information that have clarified the physiological and functional roles of these receptors (69). By using transgenic AT2Rs reporter mice combined to an in situ hybridization technique, a detailed description of AT2Rs distribution in the brain was obtained (24, 25). In several works, a high expression of AT2Rs in neurons of intermediate third of the NTS (intNTS) and in the area postrema has been detected (25). Interestingly, they also observe that fibers of AT2Rs-positive neurons terminate in the RVLM and in the PVN but are not directly expressed by the cell bodies of neurons of these brain nuclei (24, 25). For example, some AT2Rs-positive neurons arriving in the PVN were identified as neurons deriving from the MnPO, which is implicated in the regulation of body osmolarity.
The localization of these receptors in different tissues or in various cell types has been long debated. Often, data obtained by immunohistochemistry or in vitro studies indicated a localization of AT1Rs and AT2Rs on non-neuronal cells as microglial cells (70). Conversely, other studies performed in transgenic models demonstrated the presence of these receptors exclusively in neuronal cells but not in microglia or astrocytes (44). On this premise, the availability of commercial antibodies for ATRs that are characterized by low reliability further makes challenging the exact identification of their tissue distribution (50).
Humans have a unique AT1R isoform, whereas mice express two isoforms of ATRs: the type 1a, named AT1aR, and the type 1b, named AT1bR, further adding a level of complexity to the experimental system (7). AT1aR and AT1bR are differentially located in the brain, and both are implicated in the regulation of the cardiovascular system (16). The AT1aR and AT1bR share a 95% amino acid sequence homology. AT1aR is a product of the Agtr1a gene located on mouse chromosome 13; AT1bR is produced by the Agtr1b gene, which is located on mouse chromosome 3 (50). Marked differences have been observed in the non-coding regions of the two genes, thus suggesting that these differences could explain their tissue-specific expression and regulation (69).
Some decades ago, Davisson and colleagues demonstrated that mice with AT1aR deletion, obtained by using homologous recombination in embryonic stem cells (23), were protected from blood pressure increase after central infusion of AngII (23). In contrast, AT1bR-deficient mice, obtained by the same genetic strategy (54), showed normal blood pressure levels but reduced fluid intake through drinking water after central infusion of AngII (23). Taken together, these observations demonstrated an exclusive role for AT1aR in mediating blood pressure regulation and a role in fluid balance modulation for AT1bR (23).
The role of the SFO in hypertension
Although the CVOs are all structurally linked, each organ fulfills a distinct function. Exploiting in vivo models of hypertension and approaches aimed at selectively target CVOs, it is possible to identify specific functions regulated by CVOs.
Very old studies showed that lesions of the SFO hampered the drinking behavior induced by AngII infused in the nucleus of PA, additionally indicating the existence of a connection between the SFO and the PA in mediating this hypertensive phenotype (74, 99). More recently, these concepts were reexamined by utilizing novel techniques. A further specialization of the SFO AT1aR-positive neurons has been shown, suggesting the existence of an additional level of neuronal complexity (74, 75). According to this study, the SFO is composed by excitatory neurons innervating the OVLT and specialized to drive thirst, and salt neurons innervating the bed nucleus of the stria terminalis (99). Activities of these neurons sensing peripheral signaling are modulated by body fluid concentrations (74, 75). Other efficient tools were used to dissect the neural pathways established in this brain area (100). In the brain, the SFO-specific ablation of human angiotensinogen (hAGT) synthesis obtained by microinjection of an adenoviral vector encoding a Cre recombinase (AdCre) in hAGTflox mice resulted in a significant decrease in hAGT, which, in turn, correlated with reduced water intake (94). These results provide genetic evidence that the local de novo synthesis of AngII in the SFO plays an integral role in regulating fluid hemostasis (87, 94). Taking advantage of approaches that mechanically disrupt the SFO, it has also been demonstrated that this brain region regulates blood pressure levels (17).
Moreover, by exploiting a different manipulation of neural signaling obtained by electrical stimulation, an enhanced secretion of two hormones—vasopressin and oxytocin—was shown in circulation (35), as it was the sympathetic outflow (15). This effect was blocked by disrupting the PVN, thus providing evidence that the neuronal communication between this area and the SFO is amplified during hypertensive conditions (36).
The relevance of AT1aR expression in SFO neurons during hypertension was shown in an elegant experiment carried out by injecting in the SFO an adenoviral vector expressing a Cre Recombinase to selectively target AT1aR floxed alleles (51). By inducing an SFO-specific AT1aR deletion, it was, in fact, demonstrated that mice treated with deoxycorticosterone acetate (DOCA) and salt in drinking water (DOCA-salt model of hypertension) were protected from blood pressure increase, polydipsia, polyuria, and sodium intake and from an abnormal overactivation of the sympathetic drive (51). The AT1Rs signaling revealed relevant functions in the DOCA-salt model of hypertension. This model typically reduces RAS peripheral activity but, at the same time, results in an elevated brain RAS activity (53). In fact, a chronic DOCA-salt treatment increased the expression of ATRs in the SFO (45), promoting enhanced RAS activation, which, in turn, significantly contributes to the onset of hypertension.
When it became clear that the immune system participates in the pathogenesis of hypertension, an interest in investigating the SFO as a potential brain area regulating the inflammatory pathways involved in blood pressure regulation emerged. The disruption of the anteroventral third cerebral ventricle (AV3V)—a region that includes the OVLT, the ventral portion MnPO, and the PA, receiving signals from the SFO—was demonstrated to be effective in preventing the effects of hypertensive challenges on blood pressure and fluid intake (78). Further, in the same work, it was shown that in the SFO AngII exerts functions that are necessary for the consequent activation of immune cells and, more specifically, infiltration of T lymphocytes in the vasculature (78). Afterward, another study demonstrated that the intracerebral ventricular infusion of AngII concomitantly enhances peripheral sympathetic outflow and immune activation (51). These findings appear to be of particular importance, as they indicate that the effects of AngII are mainly attributable to central mechanisms, rather than to the direct actions that the hormone exerts on vasculature and immune cells.
Not many years ago and soon after the emerging evidence that psychological stress represents a non-negligible risk factor for hypertension (34), it was shown that psychosocial stressors can activate adaptive immunity and increase blood pressure in mice, similarly to AngII. T cells, activated in response to chronic stress, accumulated into peripheral vasculature and kidneys causing organ damage (71, 79). In addition, there is evidence on the role of the SFO in modulating other peripheral responses during conditions of psychological stress (64). In particular, Krause et al. demonstrated that the delivery of a lentivirus, infused directly in the SFO to knockout AT1R, inhibited adrenocorticotrophin and corticosterone release after acute stress (64). These data pose the question for future studies aimed at dissecting other responses characterizing stress-induced hypertension.
The role of PVN in hypertension
The hypothalamus is a brain region composed by small nuclei with a variety of functions. The PVN is one of the most important centers of the hypothalamus that controls typical activities of the ANS involved in cardiovascular function. As detailed earlier, the PVN is characterized by a complex system of neuronal connections with the SFO. By using neuronal tracers to follow the connections established between the SFO and the PVN in experimental models, it was possible to investigate how neurons are activated during pathological conditions. In response to psychological stressors, it was found that increased levels of blood-borne AngII elevated c-fos expression in the SFO and, moreover, by using an anterograde neuronal tracer, it was shown that neurons activated in the SFO projected to the PVN and were characterized by c-fos activation as well (64).
By combining the Cre-lox system to optogenetics, it was found that the stimulation of PVN neurons promoted the activation of the hypothalamic–pituitary–adrenal (HPA), simulating a condition of stress. In this study, a unique population of PVN neurons mediating the HPA response was identified. These neurons are characterized by the expression of AT1aRs in the parvocellular section and their stimulation proved effective in increasing blood pressure (24). Other studies executed in models of hypertension showed how the effect of SFO stimulation was prevented by PVN ablation, suggesting that this is one of the primary efferent output pathways through which the SFO contributes to blood pressure control (2, 112). Besides the SFO, the OVLT is one of the CVOs communicating with the PVN, implicated as well in the control of cardiovascular homeostasis. A very recent work showed an excitatory pathway in the brain mediated by a specific class of neurons of the OVLT and PVN, which increase blood pressure levels. In fact, the use of an antagonist of vasopressin receptors in the PVN attenuated the blood pressure increase induced by the optogenetic stimulation of afferent fibers in the PVN, derived from the AT1aR-positive neurons of the OVLT (40).
These data pose questions on how the signaling is activated by hypertensive stimuli in the brain and how, in turn, the main neuronal populations expressing AT1Rs are activated by AngII. For example, a work some years ago demonstrated that an acute AngII challenge enhanced the activity of neurons in the SFO, in the PVN, and in the brainstem nuclei that, in turn, provoked an increase in circulating CD4+/IL17+ T cells (118). Also, PVN neurons remained active for a longer time than those observed in other brain nuclei, suggesting a role for PVN in the maintenance of immune activation after the early response to acute AngII (118).
Understanding how the signaling is transmitted to the PVN and then to brainstem nuclei is an intriguing question. On this premise, the PVN works as a hub between CVOs and the brainstem in hypertension. It is known that the expression of brain-derived neurotrophic factor (BDNF) is increased in the PVN in response to hypertensive stimuli (32). This pathway increases sympathetic activity and blood pressure levels (32). In an interesting work, it was shown that hypothalamic BDNF overexpression enhanced blood pressure by reducing the signaling mediated by the catecholaminergic neurons of the NTS. In fact, the NTS established a well-characterized neuronal connection with the PVN, which is important for blood pressure control (105). In this work, BDNF revealed blood pressure elevating effects, by reducing the inhibitory hypotensive outputs from NTS catecholaminergic neurons that, in turn, downregulate the β-receptor signaling (105).
The rostral ventrolateral medulla
The RVLM neurons project to preganglionic neurons in the intermediolateral cell column (IML) of the spinal cord (91) to regulate the sympathetic outflow directed to cardiovascular tissues (46, 48). Hypertensive stimuli, such as AngII and high salt, modulate the excitability of these neurons (103a). The RVLM is also enriched by AT1Rs, and their activation has been associated with increased blood pressure levels in spontaneously hypertensive rats (SHRs). Importantly, salt intake enhances the sympathetic tone by stimulating AT1Rs in the RVLM, supporting the interaction established between the humoral and sympathetic outflow (41, 63). Interestingly, studies have been conducted to dissect the specific classes of neurons involved in hypertensive pathways. In fact, by using drugs against AT1Rs in specific brain areas, it is possible to characterize at the functional level by electrophysiology or at the structural level by immunohistochemistry, classes of neurons activated in other nuclei involved in the regulation of peripheral responses in hypertensive conditions. On this premise, AT1Rs inhibition by an intracerebroventricular injection of candesartan hampered the blood pressure increase induced by chronic stress in rats (117). Specifically, the intracerebroventricular inhibition of AT1Rs by candesartan inhibited the glutamatergic signaling by reducing the expression of N-methyl-D-aspartic acid (NMDA) receptor subunit NR1 and α-amino-3-hydroxy-5-methylisoxazole-4-propionic acid (AMPA) receptors in the RVLM, and it attenuated norepinephrine release in the circulation and hypertensive responses to chronic stress (117).
The RVLM is also an important center of sympathetic vasomotor tone and is characterized by both C1 catecholaminergic and non-catecholaminergic—probably glutamatergic—neurons (47). For example, the inhibition or destruction of C1 neurons corresponded to an attenuation of hypertension (73, 76) and also to a reduction of renal sympathetic nervous activity (76). Another characteristic of this brainstem nucleus is the presence of neurons that are sensitive to hypoxia. In fact, an interesting experiment conducted in SHRs showed that reducing hypoxia in the brainstem parenchyma caused the activation of sympathetic outflow and enhanced blood pressure levels (77).
An innovative emerging biotechnological tool, known as Designer Receptors Exclusively Activated by Designer Drugs (DREADDs), combines genetics and chemical approaches, allowing the identification of specific classes of neurons in vivo in several conditions of health or disease in mice and rats (3). Structurally, DREADDs are mutant muscarinic receptors activated by the inert drug clozapine N-oxide (CNO), administered in drinking water, but not by the native ligand acetylcholine (110). This approach takes advantage of the potentiality of engineered GPCRs that, expressed in distinct tissues or neurons (1), allow to dissect neural circuits underlying the complexity of pathological processes in a spatiotemporal manner (3). Principally, two types of DREADDs are used: the hM3Dq coupled to Gq to induce neuronal activation, and the hM4Di coupled to Gi to mediate inhibition of neuronal signaling (3).
Interestingly, Xiao et al. demonstrated a connection between a distinct class of neurons expressed in the RVLM and the activity of the SNS in the modulation of immune responses in an in vivo model of hypertensive mice, contributing to the development of the disease (114). They performed a series of experiments utilizing this technology to manipulate the activity of SNS and the consequent immune response, responsible for the development and maintenance of high blood pressure levels. A crucial experiment was performed in mice previously treated by an adoptive transfer of T cells isolated from hypertensive mice, which induces a sensitization to low-dose AngII (114). When mice were previously injected with Gi-DREADDs in the RVLM (inhibiting the neuronal signaling in this nucleus), this resulted in protection from blood pressure increase (114). In addition, they showed that the subpopulation of CD8+ effector memory T cells (CD8+ TEM) in the bone marrow, typically remaining in the circulation to patrol tissues and organs such as kidneys after a hypertensive challenge, was reduced by the injection of Gi-DREADDs into RVLM, suggesting the crucial role of SNS in the hypertensive responses and inflammation in target organs (114).
The NTS/DMV complex
The NTS is an integrative site that receives input from sympathetic afferents and arterial baroreceptors mainly mediating baroreflexes in hypertension. The AT1Rs are densely localized within the NTS and their activation stimulates gamma-aminobutyric acid (GABA) B receptor expression dampening the baroreflex, thereby contributing to hypertension (116). The NTS also expresses AT2Rs and, interestingly, their stimulation induces opposite responses, increasing the expression of GABA in the RVLM and mediating a reduction of blood pressure levels. Taken together, these results propose the GABA receptor as a crucial mediator of AT1Rs and AT2Rs activation and thus, a critical pathway in the modulation of hypertensive responses depending on the brain nucleus where GABA receptor is expressed (29).
Although experimental studies confirmed the importance of AT1Rs in mediating hypertensive acute responses to AngII infusion in the NTS, the chronic pathway that this stimulus activates in the NTS is less understood. By using an adeno-associated virus to inhibit AT1aRs expression in the NTS, a reduction of mean arterial pressure and peripheral inflammation was observed (98). Also, the observation that the downregulation of AT1aRs in the NTS led to a reduction in circulating CD4+ and CD8+ inflammatory cells further indicated a link to the activation of immune responses (98).
Closely associated with the NTS, the DMV constitutes a complex comprising vagovagal neurocircuits. In fact, the DMV is the main source of the vagal innervation, reaching almost all visceral organs (84). On the other hand, the NTS receives afferent sensory information from visceral organs and integrates inputs in the CNS, providing an overall coordination of several neuroendocrine functions (6). More recently, it has been demonstrated that the DMV is also directly connected to the peripheral immune system whereby the efferent celiac vagus nerve establishes synaptic connections with noradrenergic neurons of the splenic nerve at the level of the celiac ganglion (65). By an optogenetic approach that activates neurons in the DMV, an increased activity of the splenic nerve outflow was observed, exerting inhibitory functions on tumor necrosis factor (TNF) production. This work provided anatomical evidence of the previously well-described neural control of peripheral immune responses (14). Interestingly, this work identified the anatomical connections of the brain to the spleen circuit previously characterized as a crucial pathway of hypertension (12).
Participation of Innate and Adaptive Immune Responses to Hypertension-Induced Target Organ Damage
The inflammatory process provoking damage to target organs of high blood pressure results from alterations of their homeostatic functions and a huge accumulation of activated immune cells (Fig. 3); thus, the possibility to modulate the mechanisms by which the immune system is activated in hypertension to prevent end-organ damage is an exciting challenge.
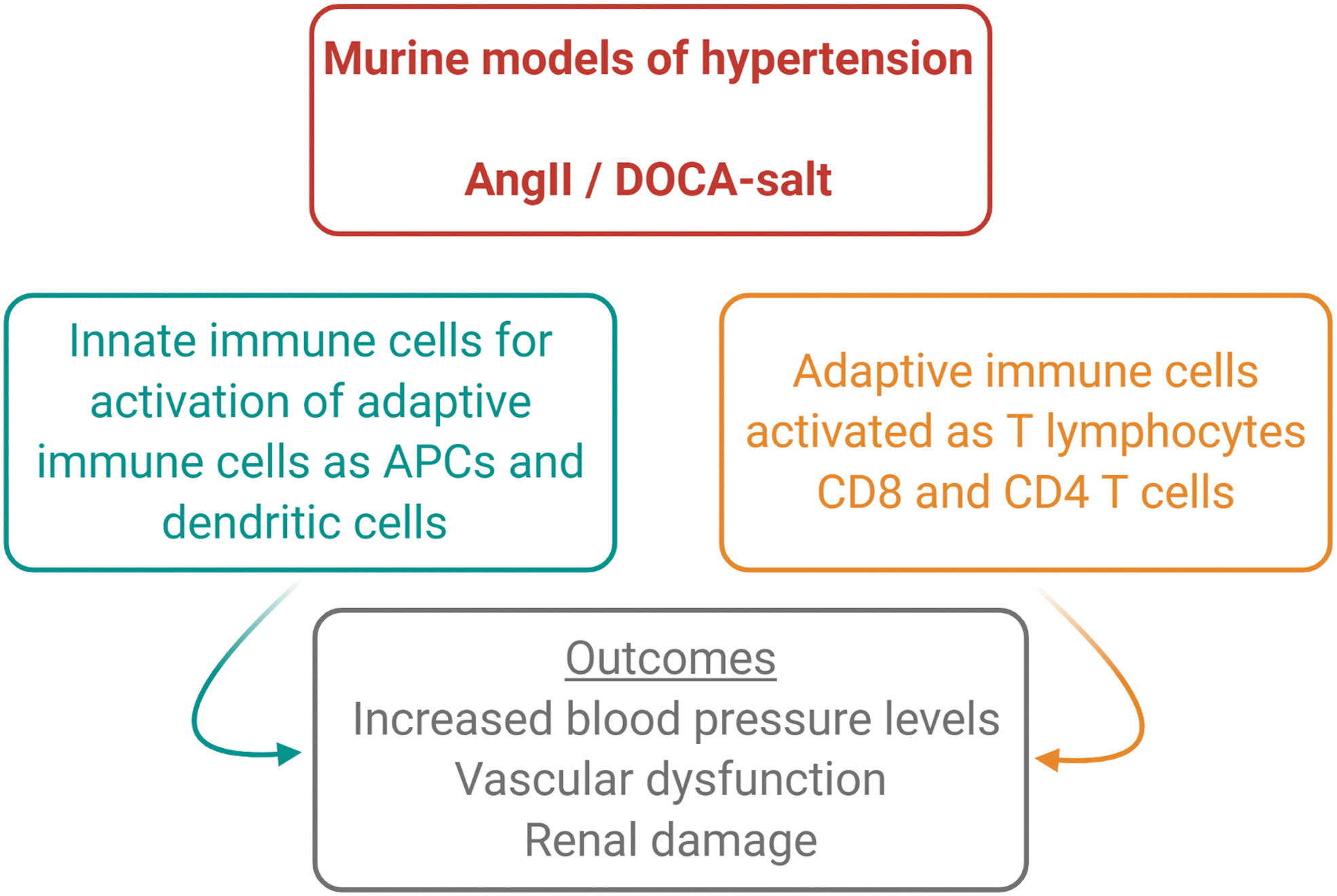
The contribution of innate immunity to hypertension end-organ damage
Innate immune cells play an important role in hypertension as well, contributing to the onset and progression of the disease. On the one hand, it has been shown that hypertensive stimuli prime the activation of myeloid cells by inducing the expression of surface markers of maturation toward antigen presenting cells (APCs) (11, 90). In particular, it was found that stimuli known to induce hypertension, such as AngII and DOCA-salt, upregulate the co-stimulation factor CD86 in splenic myeloid cells, a step of maturation that was indispensable for the consequent activation and egression of T lymphocytes from the spleen in the circulation (11, 90). These results were further suggestive of the existence of an interaction between innate and adaptive immunity, necessary for the onset of hypertension. On this premise, it was previously shown that the co-stimulation axis mediated by the interaction between the coreceptor of T cell, CD28 costimulation factor, and B7 ligands presented by APCs is crucial for hypertension (107).
It is interesting to notice that the dendritic cell (DC) isolated from mice was chronically infused with AngII to determine T cell activation, through the production of IL-17, IFN-γ, and TNF-α (61). The use of different scavenging isoketals hampered the process of T cell activation induced by DCs, protecting from hypertension and renal damage, specifically reducing fibrosis, CD3 infiltration into renal corpuscles and reducing nephrin and albumin concentrations in urine (61).
Further supporting a role of innate immune cells in hypertension, an earlier work showed that the depletion of myeloid cells hampered hypertension progression (109). In particular, by using an elegant mouse model of targeted disruption of myeloid cells, it was demonstrated that in the absence of myeloid cells, mice were protected from vascular dysfunction and showed significantly less infiltrating T cells in the vessel wall on chronic AngII infusion (109).
It has been shown that the response established at the level of peripheral tissues is partly mediated by macrophages, which have an important role in restoring homeostasis after an infection or sterile challenges such as high blood pressure. Tissue macrophages are a heterogeneous class of innate immune cells with different origins, related to the various functions that they carry out in different tissues and diseases. Classically, macrophages have been identified as originating from hematopoietic stem cells. However, in the past years, this concept has been expanded, clarifying that an additional source of myeloid cells might contribute to the innate immune response at the steady state and during pathology. An important work demonstrated that a specific class of resident macrophages derived from the yolk salk is independent from hematopoietic stem cells, further showing the capability of self-renewal of these cells in an independent manner (97).
A similar result was observed in the brain, where it was identified that a distinct population of microglial cells could be equally considered an important therapeutic target for different brain disorders (43). By an innovative multiple fate-mapping approach, a specific class of resident macrophages, populating the arterial wall and originating embryonically from CX3CR1+ precursors, was identified as capable of reestablishing homeostasis by self-renewal, interacting with the ligand CX3CL1, in the context of sepsis. The CX3CR1-CX3CL1 axis emerged as a molecular pathway involved in different functions and in particular in the recruitment of immune cell subpopulations. Whether the activation of this axis could be involved in the target organ damage ensuing in the vasculature during chronic hypertension remains to be investigated (92).
With an innovative approach that combines the analysis of macrophages by fate-mapping and the single-cell RNA sequencing, Weinberger et al. demonstrated that inflammation of adventitial macrophages caused by AngII is enhanced by the recruitment of bone marrow monocytes. Differently, the yolk sac erythro-myeloid progenitors (EMP) showed a different response to AngII related to tissue proliferation and regeneration. These data support the concept that the heterogeneous nature of macrophages is important for the identification of distinct patterns of gene expression in healthy and pathological conditions (108). In fact, the identification of specific populations of resident macrophages activated in the affected vasculature could be a promising target for future therapeutic approaches, as investigated in the contexts of other diseases (31).
The contribution of adaptive immunity in the onset of hypertension and end-organ damage
Pioneer studies highlighted the importance of the immune system in the pathogenesis of hypertension by showing that mice lacking T and B lymphocytes (Rag-1−/−) are resistant to blood pressure increase induced by hypertensive stimuli (49). With an elegant experiment, it was also observed that the adoptive transfer of T cells but not B cells reestablished the hypertensive phenotype (49). More recently, by using CD8 and CD4 T cell specific knockout mice, it was further demonstrated that CD8 knockout mice but not CD4 knockout were protected from blood pressure elevation induced by chronic infusion of AngII, thus clarifying the subtype of T cells with a critical role in the development of hypertension (106). In the following years, Mattson et al. performed an experiment that confirmed those data in a different experimental model. In particular, by deleting Rag-1 gene in Dahl salt–sensitive rats, it was shown that hypertension was attenuated (81). Notably, renal end-organ damage was reversed as well (80, 81). The importance of immune system activation in hypertension was also shown in a further experimental setting. By using scid mice, which lack lymphocytes, it was found that after chronic infusion of AngII, hypertensive responses were significantly reduced, as cardiac and renal injury were (20). During AngII infusion, scid mice manifested upregulation of TNF-α, endothelial nitric oxide synthase (eNOS), and cyclooxygenase-2 (COX-2) in the kidney, associated with an exaggerated natriuresis (20).
The spleen is a main secondary lymphoid organ that holds specific immune functions and reservoir characteristics. The discovery that tissues of the cardiovascular system are infiltrated by immune cells recruited during the course of many CVDs paved the way to intense efforts of investigation on the role of the spleen in CVDs (104). On this premise, a significant amount of works provided evidence that activated lymphocytes infiltrate target organs of high blood pressure, yet without clarifying where T cells originated from. To address this issue, an experimental model of chimeric mice by spleen transplantation was generated whereby the spleen of the C57Bl/6J mouse, which expresses the common leukocyte antigen CD45.2, was replaced by the spleen of a mouse carrying the CD45.1 isoform, making it easy for identifying splenic immune cells from resident or circulating immune cells of a different origin (11). By challenging these mice with AngII and analyzing the CD45.2/CD45.1 ratio in the immune cells infiltrating the peripheral vasculature, it was demonstrated that a significant number of T cells found in high blood pressure target organs was recruited from the spleen (11). Also, splenectomy in mice hampered the typical increase in blood pressure induced by AngII and DOCA-salt (11, 90), thus revealing that the splenic immunity is necessary for the onset of hypertension.
Subsequent studies were aimed at addressing how the splenic immune system gets activated by hypertensive stimuli. The spleen is organized in follicles containing B cells, whereas T cells and DCs are confined in the T cell zone (90). The marginal zone (MZ) separates white and red pulps and represents a connection between the circulation and immune cells (90). Intriguingly, the MZ is also entangled by fibers of the SNS, labeled by tyrosine hydroxylase (TH), suggesting that priming of immune responses might be influenced by neural regulation (72). By exploiting techniques of microneurography, which enable to directly record peripheral nerve activity, it was possible to demonstrate that hypertensive stimuli increase the sympathetic outflow on the spleen, in a way similar to that observed in cardiovascular organs (9, 10). This work led to investigating the neuroimmune mechanisms involved in hypertension and related target organ damage. Noradrenaline release in response to hypertensive stimuli was observed in the splenic zone where innate and adaptive immunity interacted, suggesting the possibility that neural signals might mediate immune priming (11).
Interesting to notice, a growth factor previously known for its angiogenic functions—named placental growth factor (PlGF)—was found to be highly expressed in the region of the spleen where noradrenergic fibers entangle stromal and immune cells (11). More important, PlGF upregulation in response to hypertensive challenges was found to be dependent on sympathetic outflow since a denervation of the splenic innervation completely abrogated its release (11, 90). The observation that the absence of PlGF protected mice from blood pressure elevation after chronic infusion of AngII or administration of DOCA-salt pursued the investigation in neuroimmune functions exerted by this growth factor (90). By transplanting the spleen of PlGF knockout mice into wild-type (WT) control mice and subjecting them to chronic AngII, it was possible to discriminate between splenic and non-splenic functions of PlGF in hypertension (11). The ablation of PlGF in the spleen was sufficient for hampering blood pressure increase, independently from the expression of PlGF in other organs (11). Consistent with this observation, the replacement of a WT spleen in mice with a null background for PlGF was effective in restoring the hypertensive response and the huge infiltration of T cells in vessels and kidneys (11).
Taken together, these data indicated that the spleen is a necessary organ for development of hypertension and that PlGF plays a role in mediating the neural control of splenic immune function. How PlGF senses neural stimuli and converts them into an effective immune response remains to be investigated.
More recently, it was also shown that the activation of splenic sympathetic outflow is an early event in the onset of hypertension and is controlled by the vagus nerve (12). In fact, at odds with expectations, the splenic nerve—a sympathetic post-ganglionic neuron departing from the celiac ganglion—was found to be controlled by the celiac efferent of the vagus nerve, classically considered a parasympathetic nerve. The combination of surgical and electrophysiological approaches showed that a resection of the celiac vagus nerve abrogated nerve firing on the splenic nerve during hypertensive stimuli (12). Further, by implementing microneurographic tools for recording nerve activity at this level, it was possible to measure the increased celiac vagus nerve activity under hypertensive challenges (11). Although these observations made it clear that a vagus-splenic connection controls the splenic immune response recruited by hypertensive stimuli, they also increased the awareness that the well-known ANS control of hypertension should be revisited by taking into account the previously unknown neuroimmune sympathetic outflow (Fig. 4).
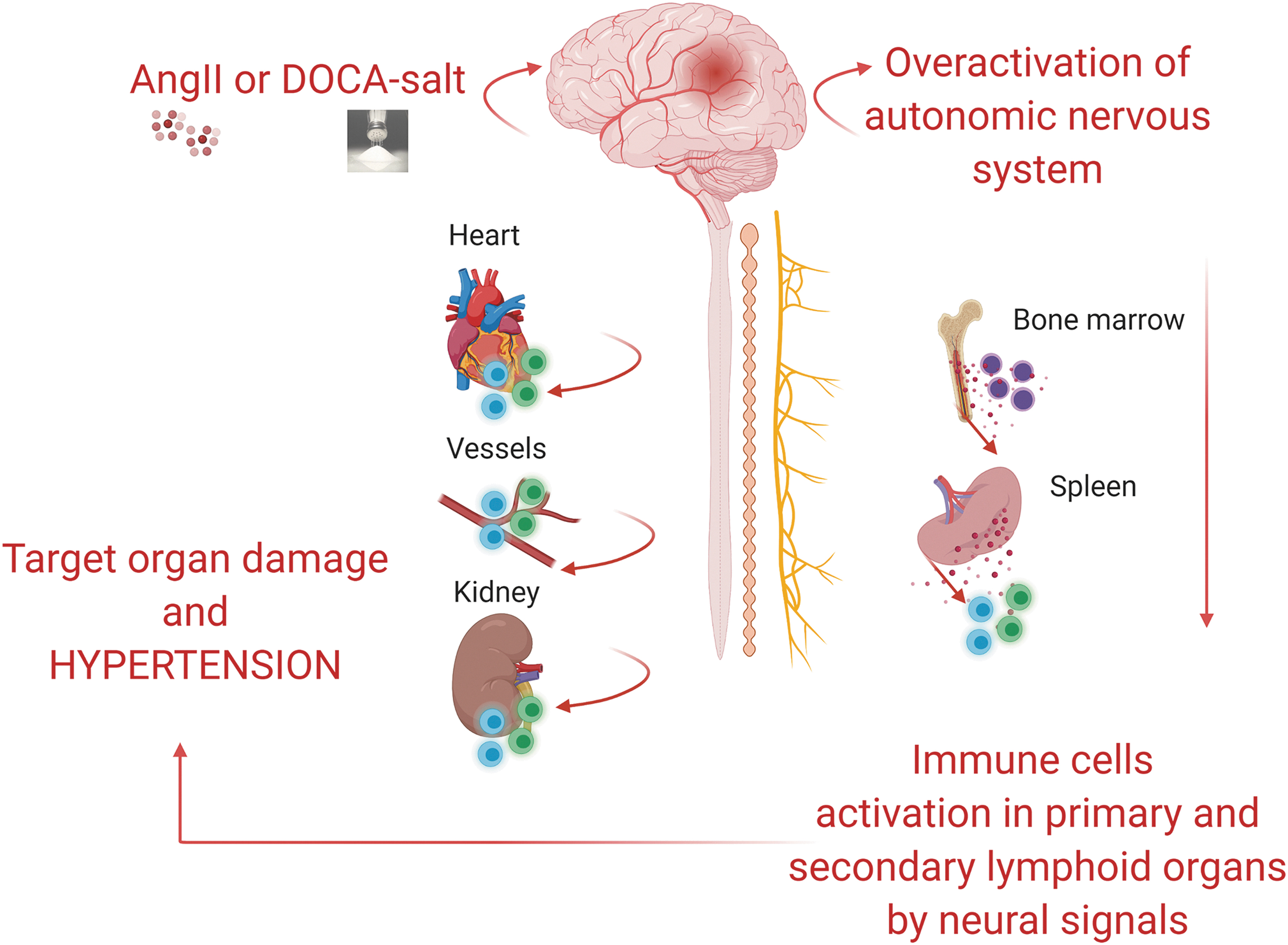
The ANS as an Intermediate Between the Brain and Peripheral Responses to Hypertension
The ANS is composed by two divisions, classically defined as the PNS and the SNS. They appear as two separate branches of the peripheral nervous system parallelly running throughout the body and regulating both common and distinct pathways. Since a long time, the ANS has been known as a critical determinant of blood pressure regulation, controlling homeostatic and altered functions of typical organs of the cardiovascular system. An imbalance in ANS function is often associated with classical outcomes of hypertension, such as glomerular injury in the kidney, cardiac hypertrophy, and vascular damage, which overall represent key determinants of end-organ damage in hypertension (33).
Surgical approaches aimed at reducing the enhanced sympathetic outflow to target organs of high blood pressure were conceived some decades ago, finding promising positive effects in reducing blood pressure levels but remaining at the observational level for many decades (103). In the meantime, a variety of drugs such as diuretics, ACE inhibitors and ATRs blockers, or combinations of these treatments have been commonly used to normalize blood pressure levels and control target organ damage (95).
In more recent times, a renewed interest in techniques allowing surgical denervation of sympathetic fibers innervating kidneys served as prospective new treatments for hypertensive patients, who were resistant to pharmacological approaches (95). The combination of pharmacological and surgical approaches effectively ameliorated blood pressure control in many patients. By applying renal denervation in experimental models, it also became possible to dissect the complexity of activities characterizing afferent and efferent fibers in kidneys (27). On this premise, it was clarified that the efferent sympathetic renal nerves regulate important functions of the kidneys such as glomerular filtration, the exchanges at the level of the renal tubule, and hormones' release. On the other hand, the afferent inputs conveyed to the brain through peripheral nerves allow to establish reflex responses. A classical finding observed in both human and experimental hypertension is the abnormal activation of efferent renal sympathetic nerves that leads to progressive alteration of organ physiology (27).
The observations just cited fueled the interest in investigating how the brain controls the main determinants of blood pressure regulation (96). Although the overall effect of systemically elevated levels of AngII on the ANS and the cardiovascular system are well known, the contribution deriving from brain-mediated actions is still the object of examination. Relevant to this, one of the most innovative concepts in this field was represented by the emerging knowledge of a neuroimmune control of hypertension (9, 10).
One of the first observations that shed light on this notion was obtained in mice that were subjected to a lesion of the CVOs (78). Chronic AngII infusion was unable to mobilize activated T cells toward the vasculature, a typical finding observed in hypertensive mice (78), thus suggesting that a brain-mediated effect could drive immune system activation. Subsequently, it was clarified that the peripheral infusion of AngII caused brain-mediated immune response by elevating sympathetic nerve activity (SNA) in the splenic district, which revealed as an essential step for blood pressure elevation and the onset of hypertension (11, 12).
On a different note, another work showed that neuroimmune interaction might also participate in the process of target organ damage, whereby immune activation in target organs of hypertension—such as the kidneys—is controlled by direct neural regulation. In fact, the bilateral denervation of renal fibers by phenol application on renal arteries protected from AngII-induced hypertension and reduced the activation of DCs, and in turn T cells, thus reducing the overall inflammatory process in the kidneys (115). This was important, because it added further information on the local control of immune processes by neural signals (115). In another model of experimental hypertension, the salt-sensitive DOCA-salt, it was further demonstrated that afferent and efferent renal nerves regulate different functions of the target organ. For example, the denervation of both afferent and efferent nerves in kidneys reduced the process of T cell infiltration associated with DOCA-salt (4). However, with the denervation of only afferents routes this process was not affected, highlighting that a distinct neural pathway is exclusively involved in the regulation of immune responses in hypertension (4).
In contrast, a work on the role of afferent and efferent routes innervating the kidney in a rat model of AngII-induced hypertension described how, by using the same method for nerve ablation, a complete denervation of renal nerves (both afferent and efferent nerves) or a selective denervation of the afferents did not attenuate T cells infiltration in the kidney (39). In addition, in all three experimental groups of denervated rats (with a complete denervation or with a denervation of afferents or efferents renal nerves alone), AngII caused a similar increase of blood pressure levels, but it did not cause a dysfunction of sodium and water balance (39). Given the presence of controversial data in this field, it is clear that many other investigations will be necessary. In fact, additional important studies suggest that this model has certainly a neurogenic origin mediated by the sympathetic outflow in the splanchnic district rather than in the kidney alone (11, 12, 60, 90) (Fig. 5).
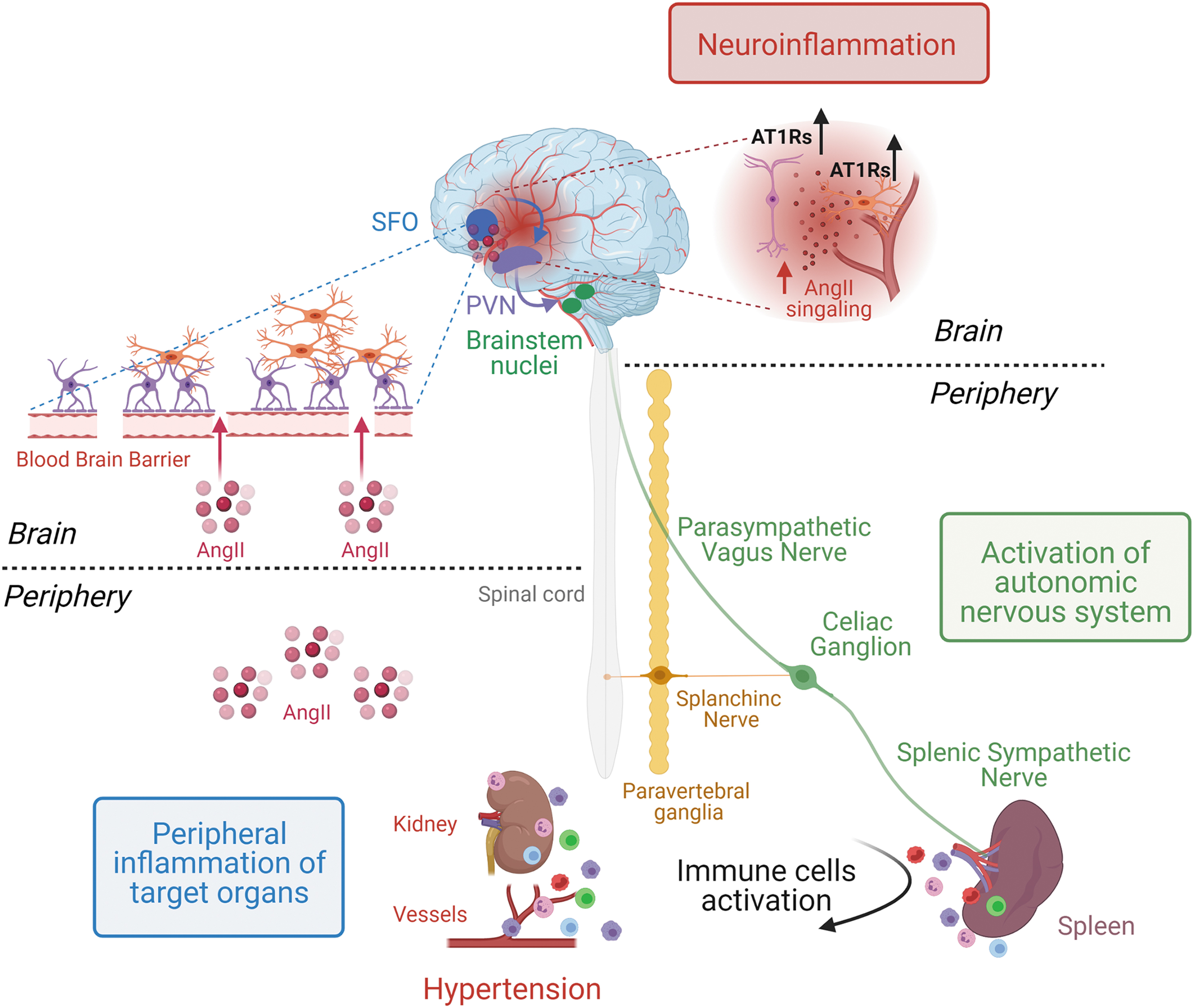
In the past, our group was working on identifying the neural pathway modulating immunity in hypertensive mice (11, 12, 90). On this premise, we established a procedure to selectively denervate the splenic sympathetic innervation by surgical thermoablation (12). In this way, we were able to hamper the neural signal activating the immune response in hypertensive mice. As a consequence, splenic denervated mice were protected from hypertension and organ damage induced by chronic AngII (12). To better characterize the neuroimmune pathway that controls the splenic immune response during hypertension, we further developed an electrophysiological approach to directly measure the sympathetic outflow on the splenic nerve (12). Mice infused with AngII or DOCA-salt showed an increased activity of the splenic sympathetic nerve in a pre-hypertensive phase (12).
More recently, we implemented a bioelectronic medicine approach to directly stimulate the celiac efferent vagus nerve, that is, the pre-ganglionic neuron of the splenic nerve, and evoke the noradrenergic outflow in the spleen (13). The sympathetic fibers innervating the spleen, when activated by bioelectronic approaches, were capable of releasing noradrenaline and promoting the expression of PlGF, which, in turn, primed the egression of CD8 effector T cells, realizing indeed the interface connecting the immune and neuronal systems in the spleen (13).
Therapeutic Relevance of the Central Control of the Neuro-Inflammatory Axis
Discovering the mutual interactions established between the nervous and immune system is becoming increasingly important for various pathologies where inflammatory processes contribute to disease onset and progression. The inflammatory reflex defined for the first time by Tracey and colleagues (88, 93) is a CNS mechanism that is able to modulate the physiological responses of the efferent arm of the vagus nerve mediating inflammation and immune responses (88, 93). This response is a reflex primed by peripheral inflammatory stimuli acting on specific brain areas, which, in turn, control the action potentials of parasympathetic and sympathetic nerves (13, 65) to modulate peripheral immune responses (13). Dissecting components of this pathway could become necessary to study pathologies with a deregulation of the immune system.
An interesting observation that emerges highlights the evidence that components of the neuro-inflammatory axis might be common in various diseases. For example, metabolic disorders such as insulin resistance, type 2 diabetes mellitus, and obesity are characterized by a disruption of the immune system homeostasis, similarly to what was observed in hypertension (52, 88). Similarly, autoimmune diseases display a sustained activation of the immune system, such as the systemic lupus erythematosus or the rheumatoid arthritis (42). Clinical evidence showed that patients with autoimmune diseases are also hypertensive, prompting to hypothesize that common immune-targeting therapies could be used for these patients (101, 111).
Taken together, these data suggest that the common mechanisms are underlying the development or progression of various diseases affecting the immune system and inflammation (86). In fact, researchers studying the role of chronic inflammation in different diseases are interested in translating this concept into therapeutic approaches that would exploit the modulation of the nervous system, to regulate the immune responses. Until a few years ago, the role of the immune system in hypertension was overlooked, as it was the involvement of neuroimmune circuits in the control of cardiovascular reflexes. The use of bioelectronic devices activating the neural reflex modulating immunity in the spleen started to appear as an innovative and pioneering approach for future treatments (Fig. 6). In principle, the idea to find a way to target the neuroimmune pathway by noninvasive therapeutic approaches could be beneficial to modulate the inflammatory responses involved in a variety of different pathologies.

Key Points
Hypertension is characterized by a status of immune activation in the brain and in peripheral organs targeted by blood pressure.
In the brain, the complexity of nervous signals activated by hypertensive stimuli is principally mediated by AT1Rs expressed in brain nuclei controlling cardiovascular functions.
With different roles and functions, the SFO, the PVN, and some brainstem nuclei are important hubs transmitting neuronal signaling from the periphery to the brain and in turn from the brain to lymphoid organs to modulate immune cells activation by the intermediation of ANS.
Chronic immune activation is a crucial pathophysiological alteration involved in hypertension and damage of the affected target organs.
Overactivity of the ANS innervation controlling the cardiovascular system is typically associated with hypertension.
The increased sympathetic activity is also responsible for the modulation of immune response. Once activated, immune cells migrate toward the target organs of hypertension, causing structural and functional damage, as well as contributing to the onset and maintenance of high blood pressure levels.
Conclusions
Taken together, the discoveries reviewed here support the concept that a dysregulation of neuroimmune interactions is critically involved in the onset and progression of hypertension. Although the increasing body of data coming from animal studies and clinical observations substantiated this innovative concept, a gap in translating these aspects in the therapeutic approach to hypertension still exists. On this premise, immunomodulating therapies for CVD are just at the beginning of their clinical investigation and many other studies will be necessary to apply the possibility of neuroimmune modulation in hypertension.
Footnotes
Authors' Contributions
Conceptualization, M.P., and D.C.; writing—original draft preparation, M.P.; writing, review, and editing, D.C.; and funding acquisition, D.C.
Author Disclosure Statement
The authors declare no conflict of interest.
Funding Information
This work has been supported by “Ricerca Corrente” granting program of the Italian Ministry of Health to D.C.