
Abstract
Significance:
Infertility is a major global health problem, with nearly half of the cases being associated with male factors. Although reactive oxygen species (ROS) are crucial for sperm cell normal physiological processes, an imbalance between ROS production and antioxidants can lead to oxidative stress that can impair sperm function. Indeed, high semen ROS levels are reported in 30%–80% of infertile men.
Recent Advances:
Male oxidative stress infertility is an uprising classification for idiopathic infertility. Proteomic approaches, including quantitative mass spectrometry (MS)-based proteomics, are being utilized to explore the molecular mechanisms associated with oxidative stress in male infertility.
Critical Issues:
In this review, proteome data were collected from articles available on PubMed centered on MS-based proteomic studies, performed in seminal plasma and sperm cell samples, and enrolling men with impaired semen parameters. The bioinformatic analysis of proteome data with Cytoscape (ClueGO+CluePedia) and STRING tools allowed the identification of the biological processes more prevalent in asthenozoospermia, with focus on the ones related to oxidative stress.
Future Directions:
The identification of the antioxidant proteins in seminal plasma and sperm cells that can protect sperm cells from oxidative stress is crucial not only for a better understanding of the molecular mechanisms associated with male infertility but specially to guide new therapeutic possibilities. Antioxid. Redox Signal. 37, 501–520.
Introduction
Male infertility is a common condition whose incidence has been increasing through the years (52). In the majority of the cases, the causes remain unknown. Oxidative stress, resultant from an imbalance between the production of reactive oxygen species (ROS) and antioxidants levels, has been reported as playing a major role in male infertility (4). Indeed, sperm function impairment due to elevated ROS levels has been ascribed to 30%–80% of male infertility cases (7). Nevertheless, under physiological levels, ROS are crucial to sperm vitality, participating in sperm maturation, hyperactivation, capacitation, acrosome reaction, and fertilization. Yet, when there is a dysregulation of the ratio between the ROS levels and the antioxidant capacity, oxidative stress ensues. Then, elevated ROS levels can induce sperm damage and lead to male infertility.
Oxidative stress seems to be particularly deleterious for sperm motility and well represented in asthenozoospermic men. Asthenozoospermia is a condition characterized by the low percentage of motile sperm in the ejaculate (23). Since ∼20% of semen samples from patients attending fertility treatments have impaired sperm motility (102), the importance of understanding the processes that link asthenozoospermia with oxidative stress is imperious.
The comprehension of the biological processes related to oxidative stress, as well as the knowledge of the proteins related to them, can help us to orientate future studies devoted to finding novel therapies for male infertility. This review aims to highlight the relevance of oxidative stress in men with abnormal semen parameters, in particular in men with asthenozoospermia, by gathering studies that evaluated seminal plasma and sperm cell proteome in this condition using mass spectrometry (MS)-based proteomic approaches. Bioinformatic analysis of the proteins found to be modulated in men with asthenozoospermia allowed to disclose disease-related networks evidencing the most relevant biological processes.
ROS in Semen: The Necessary Evil
Under normal physiological conditions, ROS are essential for sperm maturation, hyperactivation, capacitation, acrosome reaction, and fertilization. Various causes, both biological and environmental, can increase ROS levels to supraphysiological values and result in lipid peroxidation (LPO), sperm DNA fragmentation, and apoptosis, leading to infertility. In sperm cells, a plasmatic membrane sperm oxidase is the main endogenous source of ROS, which is believed to be crucial for the initiation of sperm capacitation, when in right amount (28, 29, 97).
It is still not clear the oxidase that contributes for the mentioned ROS. Recent studies point to the involvement of nicotinamide adenine dinucleotide phosphate (NADPH)-oxidase (NOX)-5 as one probable culprit for increased levels of ROS production (84, 86, 90, 97). However, its association with sperm capacitation is yet to be confirmed (95).
Nitric oxide synthase (NOS) is also a possible source of ROS responsible for sperm capacitation. It was hypothesized that nitric oxide radical (NO•), the superoxide anion, and peroxynitrite, if produced for long enough, are able to supply the sperm cells the necessary ROS to initiate the capacitation process (30, 95). At the mitochondrial level, ROS are generated via electron leakage from the electron transport chain during cellular respiration. However, in normal sperm cells, the amount of ROS generated by mitochondrial function is extremely low (66).
Other studies have shown that mitochondrial ROS have no effect on sperm capacitation (14, 133). Thus, the definitive source of the endogenous ROS necessary to trigger sperm capacitation is yet to be fully understood.
On the contrary, mature sperm cells depend on oxidative phosphorylation for energy production and maintenance of sperm motility. However, immature/abnormal sperm with cytoplasmatic droplets present this mechanism dysregulated, which significantly enhances their ROS production (66, 132). These, together with peroxidase positive leukocytes, are known to be the most significant ROS producers in seminal plasma (6). Also, the vaginal microbiome is responsible for ROS production, which modulates sperm function after ejaculation, in the female reproductive tract (111).
All seminal plasma ROS sources assist the ROS produced by the sperm cell in the induction of sperm capacitation (33). Sperm capacitation is a complex process that is believed to be initiated by cytoplasmic alkalinization, as a consequence of proton extrusion via the Hv1 proton channel, calcium (Ca2+) entry via cation channel sperm-associated protein (Catsper), and bicarbonate (HCO3 −) by sodium (Na+)/HCO3 − cotransporters and solute carriers (61). This is rapidly followed by an increase in cAMP production by adenylyl cyclase, whose activity is also related to the redox balance of the sperm cell, and therefore, ROS production (8, 22, 105).
Then, activation of protein kinase A, protein kinase C, and mitogen-activated protein kinase will lead to the activation of tyrosine kinases that greatly increase the levels of tyrosine phosphorylation and other motifs such as threonine/glutamine/tyrosine and arginine-X-X-(serine/threonine), setting in motion the process to fully capacitate the sperm cell (72, 98, 110, 126). Also, ROS are involved in cholesterol oxidation and removal of cholesterol from the sperm plasmatic membrane, enhancing the fluidity of the plasmatic membrane that promotes the development of the hyperactivation state (19).
Sperm hyperactivation is characterized by a high amplitude and torque force flagellar movement. This process is crucial for the final stage of the sperm journey when extra thrust force is necessary to propel the sperm cell through the zona pellucida (11). Likewise, acrosome reaction also benefits from controlled ROS levels (58) responsible for the increase in membrane fluidity by cholesterol efflux and Ca2+ intake (108); however, its mechanism is not yet fully understood (Fig. 1A). This process occurs when the sperm head enters into contact with the extracellular layer of the oocyte and is characterized by the secretion of proteases such as acrosin that facilitates the sperm cell penetration through the zona pellucida (35).
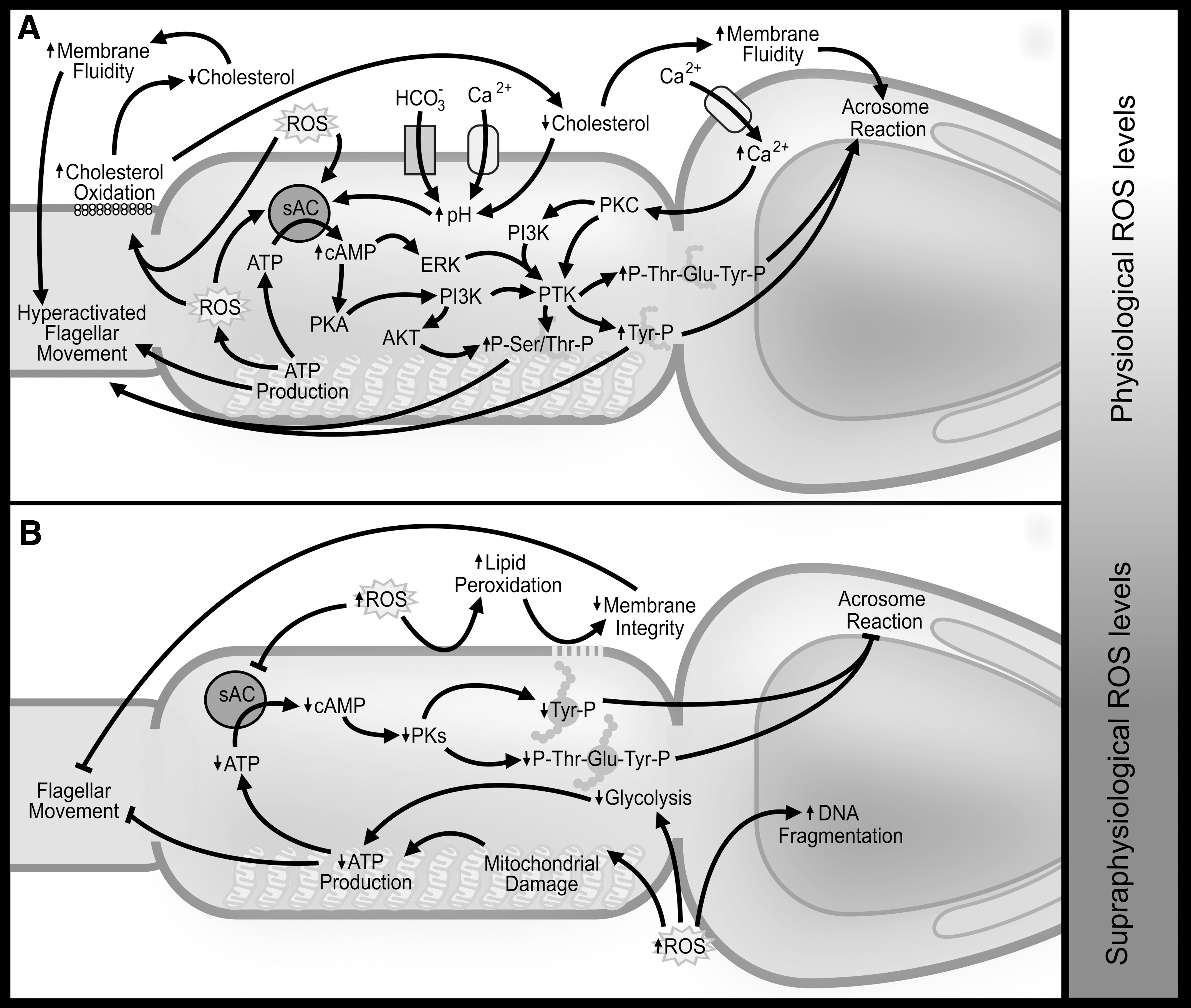
Despite sperm capacitation being ROS dependent, these are only needed in small concentrations, highlighting the importance of tightly regulated oxidative stress. In fact, incubation of sperm cells with supraphysiological ROS has an opposite effect, resulting in significantly less capacitated sperm (58, 110). Sperm cells are particularly vulnerable to oxidative stress due to insufficient antioxidant defenses related to their reduced cytoplasmic content. Also, the high content of polyunsaturated fatty acids (PUFAs) in their plasmatic membrane turns them highly susceptible to LPO, disrupting membrane permeability and damaging flagellar movement (70).
In addition, sperm cells obtain extra condensed chromatin after spermiogenesis, which means that DNA repair is practically inexistent (60). This allows sperm cells to have greater resistance to DNA damage when comparing with other cell types (117); however, when damage occurs, it cannot be repaired. Thus, men with supraphysiological seminal ROS levels have a noticeable increase in DNA fragmentation (40). Also, men with high levels of ROS in semen present sperm cells with a low capacity for ATP production (27). A possible explanation for this is the oxidation of thiol groups in important glycolytic enzymes, which ultimately results in impaired sperm motility (Fig. 1B) (39).
For all these reasons, seminal plasma with low antioxidant capability has been positively correlated with abnormal sperm parameters (64, 78). Seminal plasma antioxidants are able to protect healthy sperm cells from exogenous free radicals and reduce their deleterious effect on their function. However, in the presence of illnesses such as testicular cancer, inflammation, metabolic diseases, and unhealthy lifestyle habits such as smoking, or constant contact with radiation, ROS concentration can increase in semen to unhealthy levels. This will increase the oxidation/reduction potential of the seminal plasma by increasing the number of free radicals in relation to the number of antioxidants capable of scavenging them (5), dysregulating the redox state of the cells.
In the search to demonstrate the importance of ROS management, a study involving 3966 patients with idiopathic infertility described a positive correlation between high oxidation/reduction potential of seminal plasma and abnormal sperm parameters (4). In fact, the literature clearly supports that either the presence of supraphysiological ROS levels or the low antioxidant capacity of the semen has a great impact on sperm parameters and thus in male fertility.
ROS Semen Content in Men with Impaired Semen Parameters
Oxidative stress markers have been evaluated in individuals with impaired semen parameters. Total antioxidant capacity (TAC) and catalase (CAT) levels were found to be significantly lower in patients with asthenozoospermia, asthenoteratozoospermia, and oligoasthenoteratozoospermia in comparison with normozoospermic men. Levels of free 8-isoprostane, a product of arachidonic acid oxidation, were found to be significantly higher in patients than in the control group. Also, CAT levels and TAC were positively correlated with sperm motility and morphology, and free 8-isoprostane levels displayed an inverse correlation with sperm motility and morphology (64).
TAC was found to be significantly lower in asthenoteratozoospermic, oligoasthenoteratozoospermic, and azoospermic men, in comparison with normozoospermic men, and also a statistically significant positive correlation was observed between the TAC and sperm concentration, motility, and morphology (103). These studies support the detrimental role of oxidative stress in semen parameters.
Seminal plasma comprises several types of antioxidants that are constantly involved in free radical scavenging. Enzymatic and nonenzymatic antioxidants that are part of seminal plasma include CAT, superoxide dismutase (SOD), glutathione peroxidase (GPX), glutathione-S-transferase (GST), and peroxiredoxins (PRDXs), as well as water-soluble (uric acid, hypotaurine, tyrosine, polyphenols, vitamin C, ergothioneine, and glutathione) and fat-soluble (all-trans-retinoic acid, all-trans-retinol, alpha-tocopherol (vitamin E), carotenoids, and coenzyme Q10) scavengers (10). Between those, nonenzymatic antioxidants, such as vitamin C and glutathione, and minerals such as zinc, were found to be fundamental to the maintenance of a healthy redox state of seminal plasma (71, 82, 101).
In fact, nonenzymatic antioxidant supplementation was described to increase male fertility potential by reverting abnormal sperm parameters. Supplementation of vitamin C, vitamin E, carnitine, or zinc was also described to increase sperm motility and decrease DNA fragmentation and LPO (9, 43, 53, 81, 104, 134), being that the levels of these nonenzymatic antioxidants are diet-dependent, and not responsive to ROS levels. On the contrary, enzymatic antioxidant (e.g., CAT, SOD, and GPX) presence in the seminal plasma can be modulated by diverse pathways in response to adverse external conditions, such as excessive ROS. Thus, their expression can be modulated with the purpose of compensating for the increase in ROS levels, as that observed in non-normozoospermic men.
Few studies have focused on oxidative stress and its relationship with the alteration of specific proteins in the seminal plasma of men with impaired semen parameters, in comparison with men with normal semen parameters. Wang et al. (138) used a luminol-mediated chemiluminescence assay to determine ROS levels in seminal plasma samples from asthenozoospermic and normozoospermic men, revealing a significantly higher level of ROS in those of asthenozoospermic men.
Also, Herwig et al. (55) determined the levels of oxidative stress in seminal plasma of oligoasthenoteratozoospermic men and fertile volunteers, using an ELISA-based assay to quantify carbonyl protein content. They found higher levels of carbonylated proteins in oligoasthenoteratozoospermic men. These two studies measured ROS levels in men with impaired semen parameters and assessed the differently expressed proteins in seminal plasma samples, using MS-based proteomic techniques (Supplementary Table S1). Other studies categorized participants as ROS positive (ROS+) and ROS negative (ROS−) and analyzed seminal plasma and/or sperm cells using MS-based proteomics.
Hamada et al. (50) measured ROS levels using a luminol-mediated chemiluminescence assay and used liquid chromatography (LC)-MS/MS analysis to evaluate differentially expressed protein in the sperm cells of infertile men. Participants were divided into ROS− (if ROS levels were lower than 20 relative light units [RLU]/s/106 sperm) and ROS+ (if ROS levels were significantly higher). In ROS− sperm, they found significantly higher levels of the antioxidant proteins lactotransferrin isoform 2, lactotransferrin isoform 1 precursor, PRDX1, and manganese-dependent SOD mitochondrial isoform A precursor, in comparison with ROS+ samples, suggesting that these proteins can play a crucial role in the protection of sperm from high ROS levels.
Sharma et al. (121) also grouped infertile men as ROS− (if ROS levels were lower than 20 RLU/s/106 sperm) and ROS+ (if ROS levels were significantly higher). Using LC-MS/MS analysis, they found prolactin-induced protein to be more abundantly present in the seminal plasma of men with increased levels of ROS. A similar analysis was applied to sperm cells and the authors found that energy metabolism and regulation, gluconeogenesis, glycolysis, and oxidative stress regulation were some of the metabolic processes affected in the ROS+ group (121). Also, proteomic analysis of seminal plasma and sperm cell samples from fertile men divided into two groups, control (ROS <102.2 RLU/s/106 sperm) and ROS+ (ROS >102.2 RLU/s/106 sperm), was performed by Dias et al. (32).
NADH:ubiquinone oxidoreductase core subunit S1 was found to be underexpressed in sperm cells of ROS+ group and SOD1 and PRDX4 overexpressed. Fertile men with high ROS levels were suggested as having an effective antioxidant defense system that preserves sperm proteins, and an active proteasomal system for degradation of misfolded proteins (32).
A different classification of participants was used by Agarwal et al. (3): infertile men were subdivided into low ROS (0 to <93 RLU/s/106 sperm), medium ROS (>93 to 500 RLU/s/106 sperm), and high ROS (>500 RLU/s/106 sperm) groups and were compared with fertile men (ROS: 4 to 50 RLU/s/106 sperm) (3). LC-MS/MS analysis of the seminal plasma of these men allowed to find that proteins involved in biomolecule metabolism, protein folding, and protein degradation were differentially modulated in all three infertile patient groups in comparison with fertile controls.
Oxidative Stress-Associated Biological Processes in Seminal Plasma and Sperm Cells of Men with Idiopathic Asthenozoospermia
Envisioning to identify putative biomarkers of sperm oxidative stress, data on the alterations of seminal plasma and sperm cell proteome in individuals with impaired semen parameters were collected. Bioinformatic analysis was accomplished considering information collected from scientific articles identified by searches in PubMed until February 28, 2021, using the terms “seminal plasma proteome semen parameters”, “seminal plasma proteome asthenozoospermia”, “seminal plasma proteome antioxidant” and “seminal plasma proteome oxidative stress”. In the case of search for sperm cell biological sample, “sperm cells” replaced the words “seminal plasma” in the terms described above.
Only articles in the English language and enrolling humans were considered. Reviews and commentaries were excluded. The references listed in the included articles were also hand searched. Since a very small number of studies evaluated seminal plasma and sperm cell proteome in individuals categorized according to ROS levels and taking into account that the criteria used to group the individuals are not consistent about ROS level intervals, we excluded these studies from the bioinformatic analysis and included only studies that did not group participants taking into account ROS levels.
Also, article collection focused on studies that did not include participants with varicocele, testicular cancer, obesity, or other conditions that are known to affect fertility, in a way to further explore idiopathic asthenozoospermia and to not include possible proteomic alterations related to hypoxia and hormonal dysregulation that derive from those conditions. Focus was given to studies that evaluated the levels of proteins in the seminal plasma and sperm cells using MS-based proteomic approaches. These approaches rely on major analytical techniques to identify and quantify proteins in several biological samples and are essential tools for providing insight into biological processes (34).
Three articles were considered in the case of seminal plasma and nine studies for sperm cell samples (Supplementary Tables S1 and S2). Most of the studies included used LC-MS/MS methodology with an Orbitrap mass spectrometer. Also, three of them (one in seminal plasma and two in sperm cell studies) used a proteomic labeling strategy tandem mass tag, allowing more accurate quantification of the protein levels in the samples.
An integrated analysis of the proteins found differentially expressed in seminal plasma and sperm cells from individuals with asthenozoospermia was performed with Cytoscape (v3.5.1) (120), plugins ClueGO (v2.5.7)+CluePedia (v1.5.7) (16, 17). This approach allows the analysis and visualization of the biological processes for clusters of genes in an integrative network. Furthermore, we used STRING v11.0 (128) that allowed the visualization of protein–protein interactions and functional enrichment analysis.
Seminal plasma proteome in asthenozoospermic men: role of oxidative stress in sperm (dys)function
Seminal plasma is the body fluid with the higher antioxidant concentration, which is important for sperm viability (62). Studies report a negative relationship between seminal plasma antioxidant levels and male infertility (41, 57, 127). The seminal plasma antioxidant defense system protects sperm cells during ejaculation and the first steps of the passage through the female reproductive tract (31, 114). The protective role of the antioxidants present in the seminal plasma is particularly important in defending sperm cells from the oxidative stress generated by infiltrating leukocytes, principally neutrophils (115).
In the seminiferous tubules, sperm cells are in an immune-privileged environment, and thus, contact with activated phagocytic leukocytes is reduced. However, after that, sperm cells can be exposed to phagocytes. This is particularly deleterious for sperm in cases of genital tract infection (115, 141).
Although the number of proteomic studies focused on individuals with impaired semen parameters is not very high, bioinformatic analysis highlighted biological processes related to oxidative stress, emphasizing the importance of these processes in seminal plasma to the pathophysiology of asthenozoospermia. According to Figure 2, a biological process associated with oxidative stress that is underlined in the Cytoscape analysis is the “glutathione metabolic process.” This seems to be related to the upregulation of GSTM3 and the downregulation of GSTP1, glutathione reductase (GSR), and Parkinson disease protein 7 (PARK7) proteins.
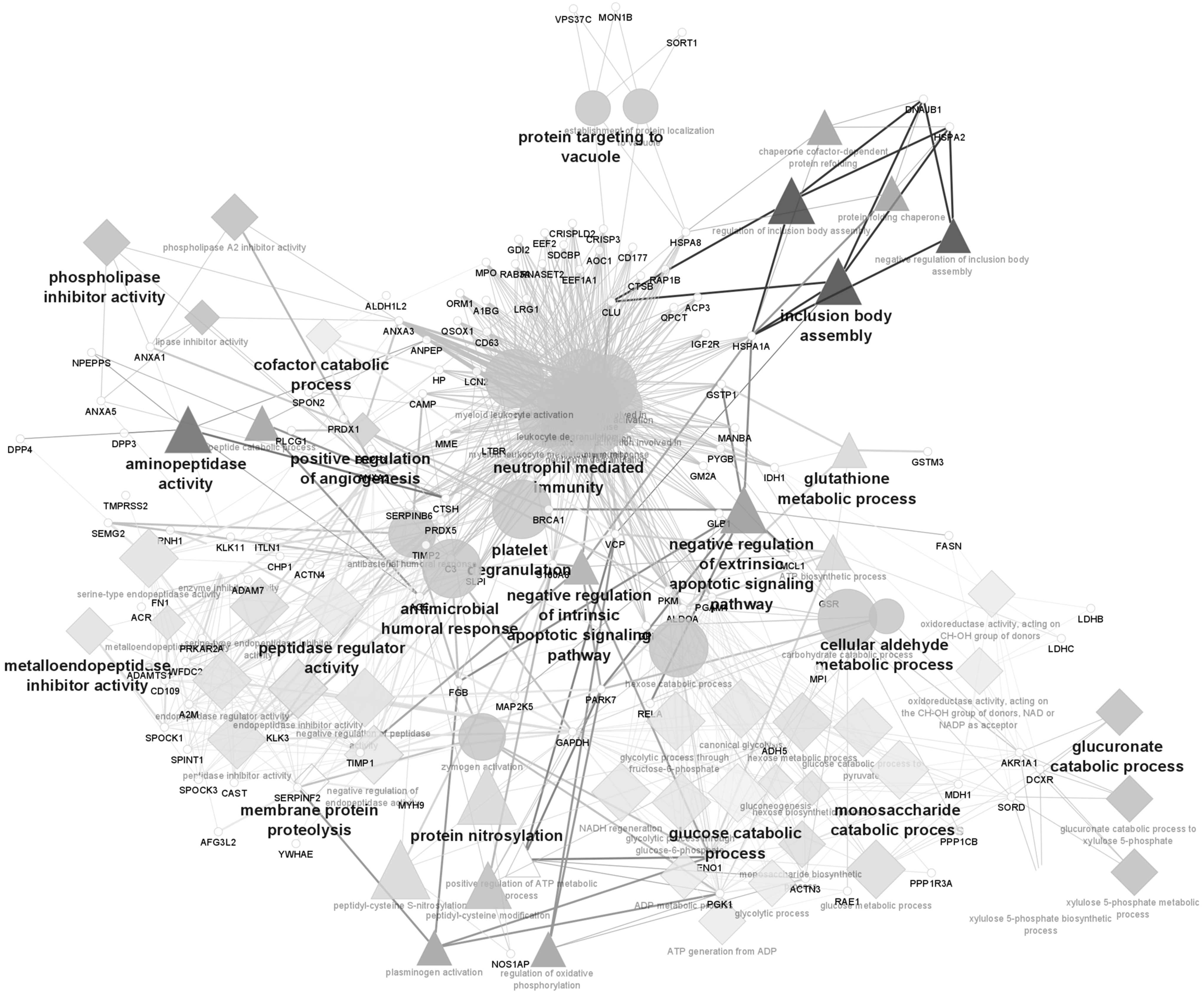
GSTs are important antioxidants that play several roles in sperm (76). They are known to be associated with cell detoxification, preventing lipid membrane peroxidation. They also participate in cellular signaling regulation involved in sperm capacitation and in fertilizing ability. For instance, GSTM3 is involved in sperm-zona pellucida binding events. GSTM3 and GSTP1 are secreted in the epididymis during sperm maturation (76). GSTA1-1 and P1-1 were found to be decreased in subfertile compared with fertile men, but without reaching statistical significance. This can be related to the high interindividual variation of the participants in the study, as well as the multifactorial cause of subfertility. However, the imbalanced presence of these proteins in seminal fluid supports a role in the defense against oxidative stress of sperm cells (109).
Glutathione is a major antioxidant. It can exist in the reduced (GSH) and oxidized (GSSG) forms. GPX, a selenium (Se)-containing antioxidant enzyme, catalyzes the reduction of hydrogen peroxide (H2O2) and lipid peroxide in the presence of GSH, which is converted to GSSG. In turn, GSSG is reduced by GSR in the presence of NAD(P)H (2). Thus, the glutathione system can play a crucial role in the protection of sperm cells, by scavenging H2O2 and lipid peroxides and consequently impairing the chain reaction of LPO of the sperm cell plasmatic membrane (2).
Another biological process highlighted in Cytoscape analysis (Fig. 2), which can be related to oxidative stress, is protein nitrosylation, a post-translational modification where a NO• group is added to a thiol moiety of cysteine (Cys) residues in a reversible reaction. Sperm cells can produce NO• via the enzyme NOS. Two NOS isoforms are present in human sperm cells, endothelial NOS and neuronal NOS (77). A previous study reported various targets for protein S-nitrosylation in sperm from normozoospermic men. Interestingly, an abundance of metabolic proteins and proteins associated with energy generation and cell movement were observed among those targets, suggesting a role for S-nitrosylation in sperm motility (69).
Also, proteins involved in signal transduction were found, accordingly with a role for S-nitrosylation in the modulation of sperm function. GST, heat shock proteins (HSPs), A-kinase anchoring protein types 3 and 4, the voltage-dependent anion-selective channel protein 3, and semenogelin 1 and 2 were among the identified proteins (69). Tyrosine nitration is also a post-transcriptional modification that was associated with decreased sperm motility and with the inability of sperm cells to achieve the capacitated state (87). Thus, protein nitrosylation and nitration seem to be relevant to sperm cell function and its modulation can be related to impaired function.
Another protein found to play a role in sperm protection from high ROS levels is PARK7 protein, also known as DJ-1 protein. This protein is secreted from the testis, epididymis, and prostate, and is involved in protecting sperm cells from oxidative stress (20), through the removal of ROS such as H2O2 (129). This protein was found to be present in the seminal plasma of fertile men (144) and downregulated in asthenozoospermic individuals (138).
STRING analysis further supports the upregulation of oxidation/reduction processes in the seminal plasma of asthenozoospermic individuals (Fig. 3). For example, myeloperoxidase (MPO) was found upregulated in asthenozoospermic individuals in comparison with normozoospermic (138). Neutrophils are the main source of MPO in the body and this protein is used as a marker of neutrophil activation (107).
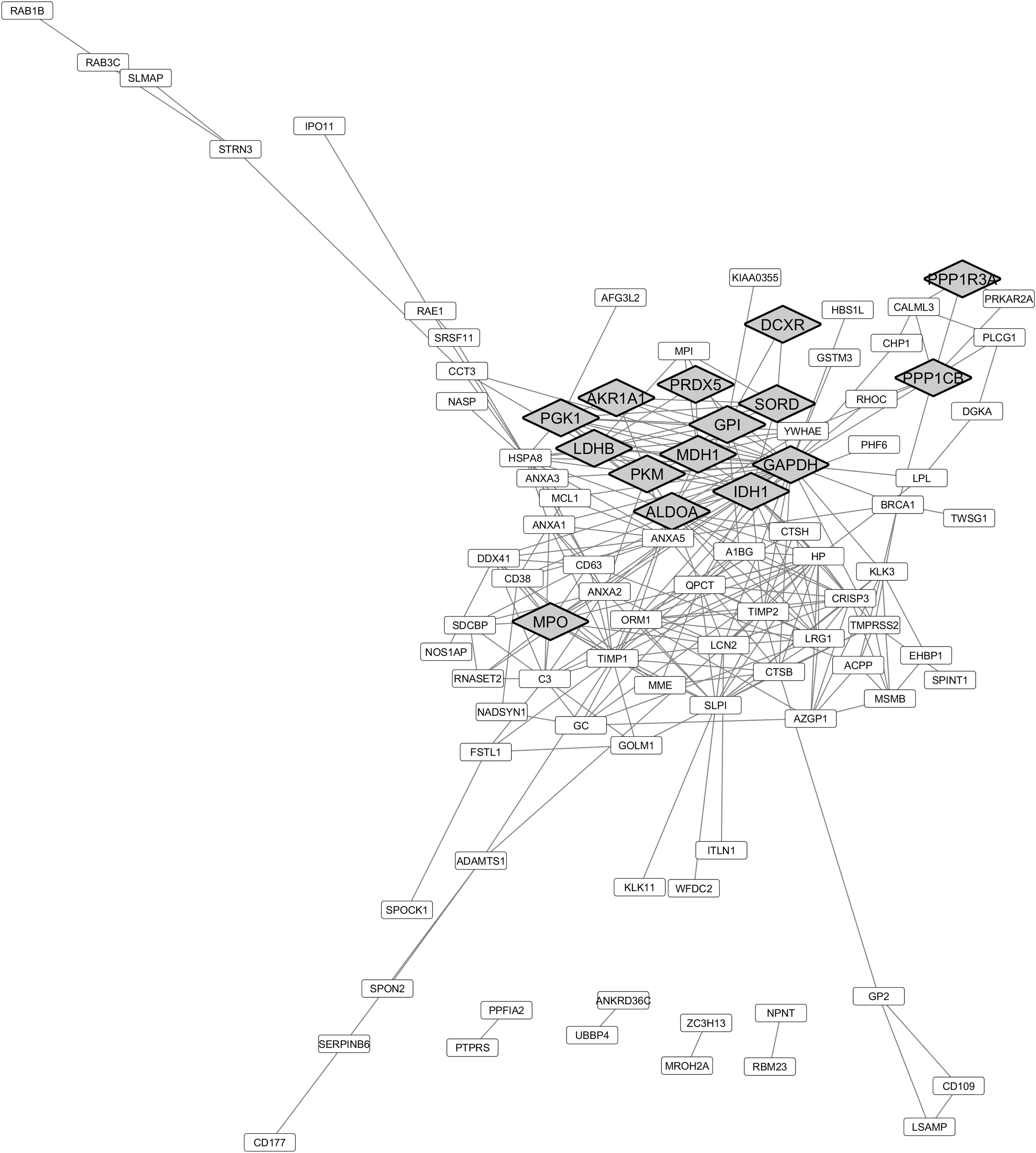
No information was reported in a study by Wang et al. (138) regarding the inclusion or not of samples with high leukocyte levels, and thus, no association with the high MPO levels is possible. Concerning the proteins associated with oxidation/reduction processes identified as downregulated in the seminal plasma of asthenozoospermic individuals (Fig. 4), STRING analysis highlighted the proteins cytoplasmic aconitate hydratase (ACO1), selenium-binding protein 1 (SELENBP1), alpha-enolase (ENO1), alcohol dehydrogenase 5 (ADH5), lactate dehydrogenase C (LDHC), glycogen phosphorylase, brain form (PYGB), sulfhydryl oxidase 1 (QSOX1), amiloride-sensitive amine oxidase [copper-containing] (AOC1), and fatty acid synthase (FASN).

Se is a micronutrient required for normal spermatogenesis (83). SELENBP1 covalently binds Se and is abundantly expressed in the liver, in contrast to the testis (38). However, the downregulation of this protein in the seminal plasma of asthenozoospermic men supports the contribution of SBP1 to impaired semen characteristics.
LDHC belongs to the LDH family, which uses NADH to reduce pyruvate to lactate. LDHC is an isozyme specific to germ and sperm cells. Compared with LDHA and LDHB, LDHC accepts not only pyruvate but also other substrates, such as alpha-ketoglutarate (131). LDHC was reported to be needed for normal male fertility (47). Thus, the reduced levels of this enzyme in the seminal plasma of asthenozoospermic can be associated with impaired fertility.
Furthermore, proteins related to the process “response to oxidative stress” were shown as downregulated in the seminal plasma of asthenozoospermic individuals. These include PRDX1, GSR, GSTP1, PARK7, ATRN, EEF2, HSPA1A, paired amphipathic helix protein SIN3A, and RELA protein (Fig. 4). Among these, GSR, GSTP1, and PARK7 proteins were also highlighted in Cytoscape analysis (Fig. 2), further supporting the relevance of these proteins in protecting sperm from oxidative stress damage.
PRDXs are targets for H2O2 due to the Cys residues at their active site and thus are readily oxidized in cells exposed to low H2O2 levels (94). Indeed, several PRDXs, including PRDX1, were detected in human seminal plasma (99). Decreased levels of total PRDX1 and PRDX6 and higher thiol oxidation of these PRDXs were observed in sperm cells of infertile men, in comparison with control ones (48). These proteins are considered the first line of defense of sperm cells against oxidative stress and their activity was found to be important for proper sperm function and DNA integrity (42). However, after contact with oxidative agents, PRDX must return to its reduced state to regain its antioxidant activity (118). GSTP1 and GSH were linked to the capability to renew PRDX6 activity (42, 145).
In contrast, PRDX1 antioxidant action inactivation by thiol oxidation occurs in two Cys residues, instead of in one Cys residue, as happens in PRDX6, and the thioredoxin/thioredoxin reductase system is responsible for the reactivation of PRDX1 (48). GSTP1, GSR, and the extracellular PRDX1 were found downregulated in men with impaired sperm motility (116, 138), which can point to a dysregulation of PRDX reduction mechanism and thus to the antioxidant capabilities of seminal plasma.
Sperm cell proteome in asthenozoospermic men: role of oxidative stress in sperm (dys)function
Despite having small antioxidant capabilities, sperm cells still present proteins responsible for the regulation of oxidative stress. Also, as seen in the seminal plasma, the expression of these proteins is modulated under unhealthy conditions in comparison with fertile men. The chaperone-mediated autophagy process was found to be upregulated in the sperm cells of asthenozoospermic men (Fig. 5). This biological process is characterized by the unfolding of proteins by chaperones and their consequent transportation into the lysosome for degradation (25).

This autophagy process is particularly sensitive to oxidative stress and has an important role in the removal of oxidized proteins (65), among others. Autophagy has an indispensable interplay in the process of apoptosis (80). The increased expression levels of proteins with roles on chaperone-mediated autophagy could explain the known increase in apoptotic sperm cells present in the ejaculate of asthenozoospermic men (142).
Taking into consideration that ROS levels are also elevated in these individuals, when comparing with normozoospermic men (4), one would assume that this also increases protein oxidation and the need for the clearance of oxidized proteins, explaining the enhanced relevance of the process of chaperone-mediated autophagy in these individuals. The excessive cytoplasmatic ROS levels and oxidative stress noted in the semen of asthenozoospermic men could also be the reason for the upregulated PRDX activity in the sperm cells (Fig. 5).
In fact, the increased expression of proteins such as PRDX2, PRDX5, and PRDX6 (Fig. 6) could be the sperm mechanism to respond to high ROS levels. These proteins protect healthy sperm cells from the ROS produced during the capacitation process (67). The upregulation of PRDX2 and PRDX5 was already described in asthenozoospermic men in comparison with normozoospermic men (116).

As above described, PRDX antioxidant enzymes tend to be upregulated in men with abnormal sperm parameters. Under oxidant conditions, rat epididymal sperm was found to present increased expression of PRDX1 and PRDX6 (75). It is believed that the epididymis epithelium secretes epididymosomes rich in PRDXs to maturing sperm cells in a mechanism to protect them from oxidative stress (96). Another possible explanation for the increased expression of PRDXs would be the antioxidant enzyme synthesis by sperm cells. That, together with replacement of oxidized proteins due to high ROS levels, could explain the fact that processes such as translation and peptide biosynthetic process were upregulated in the network analysis (Fig. 5).
This is a very controversial topic and solid evidence is yet to be presented supporting that sperm cells are capable of translating proteins. On the contrary, it is well documented that sperm cells have a pool of mRNA. However, it is believed that these transcripts are produced and stored before the chromatin compaction during spermiogenesis. Interestingly, the analysis of upregulated proteins in men with asthenozoospermia showed an increase in the regulation of histone ubiquitination process.
In other cell types, histone ubiquitination can either block the transcription or activate the expression of certain genes, depending on the local and the histone where the modification is added (91). Without knowing the class of histones and the residue modified, the information is insufficient to hypothesize if histone ubiquitination serves as a process for chromatin decompression (enhancing the possibility of new transcription) or to further compact the chromatin to protect it from oxidative damage. Another possible explanation for the upregulation of proteins involved in the process of histone ubiquitination is the high prevalence of immature sperm and germ cells in semen of asthenozoospermic men.
During spermatogenesis, especially after meiosis, the regulation of histone ubiquitination is crucial for proper germ cell development (125). Taking into consideration that asthenozoospermic men have significantly more immature sperm cells in the ejaculate when compared with normozoospermic men (26), these cells could be the ones responsible for the upregulated process of histone ubiquitination.
Bioenergetic pathway modulation in asthenozoospermic men
Two processes found to be dysregulated in asthenozoospermic males were glycolysis and tricarboxylic acid cycle. This is in line with the features of asthenozoospermia condition, characterized by a low percentage of motile sperm cells in the ejaculate. These are the two main pathways that sperm cells use to produce energy (36). Both these processes are crucial to ATP production for flagellum movement and consequent sperm motility.
Due to the compartmentalization of the mitochondrion to the middle piece of the sperm cells, it is known that the tricarboxylic acid cycle exclusively takes place in this section, whereas glycolysis takes place in the tail of sperm cells (36, 140). Glycolysis along the fibrous sheath (in the sperm tail) was found to be central for immediate ATP production and delivery, which are important for motility (89).
Enolases are located in the sperm tail and have fundamental roles in glycolysis (46). Alpha-, gamma-, and beta-enolase were found to be downregulated in sperm of asthenozoospermic men (Fig. 7). Interestingly, alpha-enolase was also found to be downregulated in a study comparing the proteome profile of sperm cells from men with high and low levels of seminal plasma ROS (122). This highlights a possible involvement of ROS on glycolysis impairment in sperm of asthenozoospermic men. Also, glyceraldehyde 3-phosphate dehydrogenase is a protein involved in glycolysis and was found to have its efficiency diminished in conditions associated with oxidative stress (39). The oxidation of thiol groups of this protein by free radicals is thought to be the cause of the decreased glycolytic rate in sperm cells exposed to seminal plasma with high oxidation/reduction potential, as asthenozoospermic men are believed to possess (4).

In addition, glycolysis was also found to be linked with bicarbonate production to increase cytoplasmatic pH and develop the capacitation process (79). It was hypothesized that enhanced glycolysis was related to increased pyruvate production and the consequent increased respiration by tricarboxylic acid cycle. The resultant carbon dioxide reacts with water in a reaction catalyzed by carbonic anhydrases to create bicarbonate, which was found to be directly correlated with flagellum beat frequency (79).
This highlights the importance of proper glycolytic rate in sperm motility and capacitation in sperm cells and that oxidative stress can be one of the causes of the dysregulation of this process in asthenozoospermic men.
On the contrary, the noted tricarboxylic acid dysfunction points to a dysregulation of mitochondrial function in sperm cells. In fact, sperm cells of asthenozoospermic men were found to have decreased activity of the respiration chain mitochondrial complexes I, II, and IV when compared with the ones of healthy controls (113). Moreover, significant mitochondrial DNA damage was found in individuals with abnormal sperm parameters (37). Since this particular DNA does not have the same protection as the nuclear DNA provided by the compaction by histones and protamine, mitochondrial DNA is more susceptible to damage.
Moreover, damaged mitochondria can further add to oxidative damage (139) that inflicts further damage to the mitochondrial DNA and decreases the activity of mitochondrial complexes (73), culminating in decreased ATP production (37). This vicious cycle ultimately results in the loss of sperm motility and could lead to apoptosis (74), and thus, infertility.
Midpiece and flagellum structural protein modulation in asthenozoospermic men
As seen above, the decrease in ATP production can have a fundamental role in the loss of sperm motility. However, other factors that can have an impact on sperm motility were found to be dysregulated in asthenozoospermic men. The analysis noted a defect in processes such as microtubule anchoring to the centrosome and cilium movement involved in cell motility (Fig. 5). Thus, sperm from asthenozoospermic men has functional and structural alterations in motility. Oligozoospermic men were reported to have a decreased expression of sperm centrosomal proteins when compared with normozoospermic men (56).
Here, the data suggest that a similar process could happen in asthenozoospermic men. As both centrioles are located in the midpiece of human sperm cells (13), this suggests that the proteins that connect them to the structural proteins of the sperm tail would also be in the midpiece. Interestingly, this is also where ROS are produced in great quantities in sperm, which makes these proteins particularly exposed to oxidative damage. Despite that, this is just a hypothesis and the level of oxidation in the structural proteins of the flagellum should be determined to confirm it. In addition, proteins found to be downregulated such as the pericentriolar material 1 protein (PCM1) were found to be expressed in the midpiece and throughout the sperm tail (143).
It was already demonstrated that impaired axoneme development and centriole defects are present in many cases of asthenozoospermia (13). The latter has implications even after fecundation, during embryo development (119). This points to the process of cilium movement involved in cell motility also being dysregulated in asthenozoospermic men. In this process, proteins involved in the physical support of cilium movement such as proteins with a role in cilium movement were encountered to be downregulated in patients' sperm.
Proteins such as tektins (TEKT) have a fundamental role in the structure of the axoneme. TEKT3- and TEKT4-null mice sperm was found to have decreased progressive motility and deficient tail movement (112, 143). Also, other proteins such as outer dense fiber proteins 1 and 2 were found to be downregulated in asthenozoospermic sperm (Fig. 7); however, the expression of these proteins was not found to be related to the semen oxidative stress (122).
Evidence-based literature suggests that oxidative stress can be the cause of many of the processes dysregulated in sperm from asthenozoospermic men. However, the available information does not associate the morphological deficiencies in the sperm cell axoneme (TEKTs and outer dense fiber protein) with oxidative stress. Functionally, CatSper1 was also found to be downregulated in asthenozoospermic sperm. This protein is primarily responsible for Ca2+ influx in sperm cells. The high cytoplasmatic concentration of Ca2+ in sperm is involved in processes such as hypermotility, capacitation, and acrosome reaction (61). In human sperm, CatSper1 inhibition resulted in decreased sperm motility and acrosome reaction (21, 63).
Previous studies already described that asthenozoospermic men have decreased expression of CatSper1 in comparison with normozoospermic men. Mohammadi et al. demonstrated that vitamin E supplementation increased the expression of CatSper in sperm from aged mice to levels comparable with the ones of young mice (85). This resulted in an increase in sperm viability and motility compared with baseline values. Vitamin E is a powerful antioxidant already studied as a treatment of seminal plasma with low TAC (9, 44). Oxidative stress could be regulating the expression of this Ca2+ channel and impairing sperm functioning. This is not the first time that this hypothesis is described as other authors have also noted this pattern (130); however, no study was found confirming this hypothesis.
Asthenozoospermic men sperm presents downregulated processes of sperm-zona pellucida recognition, binding, and fusion, according to the literature analysis performed (Fig. 5). That could be due to the impairment in the expression of proteins such as sperm acrosome membrane-associated protein 3 (SPACA3) (also known as SLLP1) that have fundamental roles in sperm oocyte binding and membrane fusion (54). A study performed on mice noted that suppression of SPACA3 expression resulted in a complete inhibition of sperm-oocyte binding and fusion (54). Moreover, the expression of this protein in canine sperm cells was inversely correlated with the oxidative stress present in the cryopreservation medium (1), highlighting a possible role of oxidative damage in the downregulation of SPACA3.
The literature herein revised points to a deleterious effect of oxidative damage on glycolysis, mitochondrial function, Ca2+ intake, and sperm-oocyte binding and fusion. However, it stands out that transmembrane proteins are underrepresented, with only a few examples found in the data collection. By being hydrophobic, transmembrane proteins tend to need a specific protocol for proper protein extraction, especially in noncapacitated sperm cells where the plasmatic membrane is rich in cholesterol, further making it difficult for protein extraction. That fact hides an important part of the sperm cell proteome that would help the scientific community to further understand the menace, that is, oxidative damage to sperm cells.
As an example, the expression of transmembrane NOX-5, responsible for the formation of new ROS and oxidative damage in sperm proteins, was found to be upregulated in asthenozoospermic sperm (135). Taking this into consideration, it would be important to further study the transmembrane proteome of sperm cells.
Dysregulated semen antioxidant proteome in relation to oxidative stress
The comparison of the differently expressed proteins in seminal plasma and sperm cells highlighted the up- and downregulation of key antioxidant proteins; some were already aforementioned, but will be further described in this section. This comparison highlights possible key proteins that are over- or underexpressed in both sperm cells and seminal plasma, as well as possible targets for the development of therapies for individuals with asthenozoospermia due to oxidative stress.
Also, possible biomarkers for asthenozoospermia caused by oxidative stress are emphasized that can fasten the diagnostic and treatment of such individuals since the seminal plasma can represent the expression of some proteins in the sperm cells. Among the upregulated proteins in seminal plasma and sperm cells, results showed the proteins arachidonate 15-lipoxygenase B (ALOX15B), PRDX5, and isocitrate dehydrogenase 1 (IDH1) cytosolic. ALOX15B protein is involved in the catalysis of PUFAs; however, during this process, it is also responsible for the formation of ROS. Thus, the overexpression of this protein is pointed to be correlated with increased LPO in sperm cells (136).
Moreover, a study demonstrated that the inhibition of ALOX15B with a selective inhibitor (PD146176) decreased ROS production in sperm and allowed the progression of the capacitation process (137). A protein found upregulated in both sperm cells and seminal plasma that has the capacity to help regulate the increased ROS levels is the PRDX5. This protein was already discussed in the sperm cell section; however, it is interesting to see that it is upregulated in the seminal plasma. PRDX5 is not regarded as an extracellular PRDX in normal conditions, being mostly present in peroxisomes, mitochondria, and cytoplasm (51). The lysis of apoptotic sperm cells and the release of this soluble protein to the extracellular medium could be the origin of the PRDX5 in seminal plasma.
Another study raised a similar hypothesis when noted the presence of PRDX1 in extracellular medium (123). It would be interesting to study if the upregulated PRDX5 resulting from apoptotic or unviable sperm cells could be used to protect healthy sperm cells from oxidative stress by increasing seminal plasma antioxidant capability.
In the case of the cytosolic IDH1, this protein has a role in NADPH renovation that was found to be important for glutathione production, and thus, the redox state of the cell (68). This fact guides us to the protein GSR, which was found to be downregulated in sperm cells and seminal plasma. The correlation of glutathione levels in the seminal plasma with the ones of sperm cells was already described (100). The low representation of the GSR in semen can be linked to the reduced concentration of GSH and the decrease of antioxidant capacity of the medium.
A study by Atig et al. described that sperm from normozoospermic men presented a higher GSH concentration when compared with the one of asthenozoospermic men (12). It is interesting that only GSR is upregulated and not the other enzyme involved in the renovation of the glutathione: the GPX. The decreased activity of this enzyme in the seminal plasma has been described as a biomarker of fertility dysfunction (24, 45). In fact, asthenozoospermic men seminal plasma had decreased GPX activity when compared with normozoospermic males (24). Despite that, contradictory results were reported in GPX protein expression on sperm cells from asthenozoospermic men (116, 124).
It is worth mentioning that protein expression levels and protein activity are not always positively correlated, meaning that despite not decreasing the expression of GPX, the activity was found to be impaired and that is a more valuable information than only the expression per se. By being a selenoprotein, GPX activity was found to be dependent on Se concentration in the seminal plasma (106). Moreover, SELENBP1 and the mineral Se were found to be underrepresented in the seminal plasma of asthenozoospermic men when compared with the values found in fertile men (18, 138). Thus, decreased seminal Se concentration could be the culprit for the impaired GPX activity found in asthenozoospermic patients.
CAT, like GPX, is able to scavenge the free radical of H2O2 and transform it into water, thus decreasing ROS. It is believed that this enzyme has low importance in ROS mitigation in the sperm cytoplasm (93); however, CAT activity in the seminal plasma is regarded as a marker of male fertility. Men with asthenozoospermia had decreased CAT activity in the seminal plasma when compared with the one of normozoospermic men (64). The H2O2 that is neutralized into water by CAT is formed from the ROS superoxide, resultant of the mitochondria, by the SOD. This protein is found to be upregulated in the seminal plasma and in the sperm cell, but in the form of different SODs (SOD3 and SOD1, respectively).
Some studies reported conflicting results regarding the activity of SOD3 in men with abnormal sperm parameters (15, 64); however, the cytosolic homologue (SOD1) of this protein is underrepresented in asthenozoospermic men (49). In mice neuroblasts, the induction of mitochondrial damage resulted in the continuation of superoxide production, but was noticed a decrease in H2O2 production. That was probably due to the downregulation of SOD1 after mitochondrial damage that further produced ROS, which ultimately leads to apoptosis (59). As mitochondrial damage is known to be highly abundant in the sperm cells of asthenozoospermic men (92), a similar process can be happening in the sperm cells of these individuals.
These pieces of evidence point to the dysregulation of asthenozoospermic sperm metabolism that presents enhanced ROS production caused by mitochondrial damage and upregulation of proteins such as ALOX15B. This is accompanied by the downregulation of the primary enzymatic antioxidant defenses, such as SOD and GSR, but the upregulation of other antioxidant proteins (PRDX5 and IDH1) that seem to be trying to regulate oxidative stress, although without much success, when taking into consideration all the evidence available. Thus, antioxidant properties in the semen are found to be crucial for sperm motility and that antioxidant supplementation is able to increase semen TAC and even reestablish sperm motility in men with asthenozoospermia (88).
Conclusion
Recent information indicates that oxidative stress and the consequent oxidative damage are a likely culprit for many of the processes found to be impaired in sperm cells with compromised motility. The involvement of oxidative stress in mitochondrial dysfunction, the loss of membrane integrity, and the DNA fragmentation have irreversible effects on sperm function. Proteomics was found to be a powerful tool for the identification of proteins modulated by conditions such as asthenozoospermia. Despite that, many challenges are still present, and innovation is needed to overcome them.
The great variability within individuals with the same condition and even between the sperm cells of the same individual complicates in achieving consensus. Notwithstanding, the implementation of new methodologies in this topic can further enlighten the effect that oxidative stress has on sperm function. A deeper study of sperm cell membrane subproteome and the development of methodologies applying metabolomic approaches to sperm cells and seminal plasma of asthenozoospermic men would facilitate the assembling of the puzzle pieces that are the effects of oxidative stress in sperm function. In that way, new treatments for infertility could be developed.
Footnotes
Acknowledgments
Inkscape, Cytoscape, and STRING were used to generate the figures in the article.
Authors' Contributions
J.C.R. and R.N.-F. reviewed the literature, wrote the article, and created the figures. F.A., M.G.A., R.F., and P.F.O. conceptualized the concept, and critically reviewed and edited the article.
Author Disclosure Statement
No competing financial interests exist.
Funding Information
This work was supported by “Fundação para a Ciência e a Tecnologia”—FCT to LAQV-REQUIMTE (UIDB/50006/2020), UMIB (UIDB/00215/2020 and UIDP/00215/2020), ITR—Laboratory for Integrative and Translational Research in Population Health (LA/P/0064/2020), and the postgraduation student João C. Ribeiro (Grant No. UI/BD/150749/2020), Pedro F. Oliveira is funded by national funds through FCT—Fundação para a Ciência e a Tecnologia, I.P., under the Scientific Employment Stimulus—Institutional Call—reference CEECINST/00026/2018. The work was cofunded by FEDER through the COMPETE/QREN, FSE/POPH, and POCI-COMPETE 2020 (POCI-01-0145-FEDER-007491) funds.
Supplementary Material
Supplementary Table S1
Supplementary Table S2
Abbreviations Used
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.