
Abstract
Significance:
Microbial neurotransmitters, as potential targets for cancer therapy, are expected to provide a new perspective on the interaction between the gut microbiome and cancer immunotherapy.
Recent Advances:
Mounting data reveal that most neurotransmitters can be derived from gut microbiota. Furthermore, modulation of neurotransmitter signaling can limit tumor growth and enhance antitumor immunity.
Critical Issues:
Here, we first present the relationships between microbial neurotransmitters and cancer cells mediated by immune cells. Then, we discuss the microbial neurotransmitters recently associated with cancer immunotherapy. Notably, the review emphasizes that neurotransmitter signaling plays a substantial role in cancer immunotherapy as an emerging cancer treatment target by regulating targeted receptors and interfering with the tumor microenvironment.
Future Directions:
Future studies are required to uncover the antitumor mechanisms of neurotransmitter signaling to develop novel treatment strategies to overcome cancer immunotherapy resistance. Antioxid. Redox Signal. 38, 298–315.
Introduction
Cancer immunotherapy, notably CTLA-4 and PD-1/PD-L1 immune checkpoint blockades, has made breakthroughs in various cancers, including melanoma, lung, stomach, colorectal, and liver cancer (Tontonoz and Gee, 2015). Despite these advances, immunotherapy has still failed in many patients; for example, in non-small cell lung cancer (NSCLC) patients, even up to 80% of patients do not respond (Gandhi et al, 2018; Herbst et al, 2020; Reck et al, 2016). A series of independent reports showed notable differences in the composition of gut microbes from responders and nonresponders to anti-PD-1 inhibitors.
By fecal microbiota transplantation (FMT) from patients to germ-free (GF) mice, many beneficial bacteria were found to have a favorable clinical response to anti-PD-1 immunotherapy, including Akkermansia and Alistipes (Routy et al, 2018), Clostridiales and Ruminococcaceae (Gopalakrishnan et al, 2018), and Bifidobacterium longum, Enterococcus faecium, and Collinsella aerofaciens (Matson et al, 2018).
Moreover, our previous study found that a higher relative abundance of Bacteroides vulgatus and Parabacteroides distasonis points to a better response to anti-PD-1 immunotherapy (Huang and Wu, 2021). In addition, two phase I clinical trials proved that modulation gut microbiome could augment the efficacy and overcome resistance to anti-PD-1 blockades in advanced melanoma (Baruch et al, 2021; Davar et al, 2021). Nonetheless, the mechanisms in which microbes, their products, and/or their metabolites influence tumor immunity were still poorly understood.
Multiple studies have suggested that microbial metabolites potentiated the efficacy of immune checkpoint inhibitors (ICIs) (He et al, 2021; Luu et al, 2021; Mager et al, 2020). Gut microbiota can produce and/or metabolize a wide range of mammalian neurotransmitters, including dopamine (DA), norepinephrine (NE), serotonin (5-HT), or γ-aminobutyric acid (GABA) (Roshchina, 2016; Strandwitz, 2018). Manipulation of these neurotransmitters can not only regulate physiological function but also has been shown to affect the initiation or progression of diverse cancers, including colon (Sarkar et al, 2008), pancreatic (Renz et al, 2018), prostate (Zahalka et al, 2017), gastric (Chakroborty et al, 2004), breast (Sarkar et al, 2008), and ovarian cancers (Basu et al, 2001).
Monoamine oxidase A (MAO-A) knockout mice could activate tumor-infiltrating CD8+ T cells by boosting the autocrine 5-HT to enhance the antitumor effect (Wang et al, 2021b). DA, glutamate, neuropeptide Y (NPY), and calcitonin gene-related peptide (CGRP) at low physiological concentrations (∼10 nM) alone or combined could decrease the percentage of PD-1+ T cells and the expression of PD-1 on T cells, and increase the cytotoxicity of hepatocellular carcinoma cells (Levite et al, 2021).
Nevertheless, these studies reveal several gaps in knowledge, including whether manipulation of these microbial neurotransmitters/receptors can potentiate cancer immunotherapy. In this study, we outline the pro/antitumor effect of neurotransmitters/receptors and the influence of microbial neurotransmitters on immune cells and cancer cells, and attempt to put forward a new potential strategy in combination with cancer immunotherapy.
Neurotransmitters Affect Immune Cells to Promote/Control Tumor Progression
Neurotransmitters can be divided into four categories; that is, biogenic amines, amino acids, peptides, and others (Fig. 1). Biogenic amine neurotransmitters were the first to be discovered, including Epi, NE, DA (Kumar, 2003), and 5-HT (Valenstein, 2002). Neurotransmitters are not only released from the brain, peripheral nerve plexus, ganglia, and adrenal medulla by the autonomic nervous system but also can be produced by gut microbes. Specific pathogen-free (SPF) mouse intestines are predominant with free-type NE and DA, while GF mice are mainly glucurono-conjugated type.

When colonized either a mixture of Clostridia species or complete SPF microbiota rich in β-glucuronidase (GUS) activity, the concentration of free NE and DA significantly increased, indicating that gut microbiota can regulate the production of NE and DA via GUS (Asano et al, 2012). 5-HT can also be directly produced by deconjugation via bacterial GUS and sulfatase (Sudo, 2019) (Fig. 2A).

In addition to the direct modulation via microbial enzymes, gut microbiota can indirectly control neurotransmitters production. For example, nucleotide-binding oligomerization domain 1 (Nod1) ligand from gut microbes can be sensed by adrenal chromaffin cells and directly modulates epinephrine secretion (Xiang et al, 2021). Enterochromaffin cells (ECs) can sense nutrients and gut microbiota to secrete 5-HT (De Vadder et al, 2018). It has been proven that gut microbiota can mediate the interplay between metabolites and antitumor immunity. Thus, we summarize the microbial-related neurotransmitter signaling on cancer development and progression, and provide insights and perspectives regarding the rationales and strategies to enhance the efficacy of cancer immunotherapy by targeting neurotransmitter signaling.
Cancer-inhibiting effect of biogenic amine neurotransmitters
In recent years, neurotransmitters have become important microenvironmental components that regulate tissue homeostasis and affect a variety of malignant phenotypes of human cancers (Boilly et al, 2017; Jiang et al, 2020). Growing evidence supports that concentrations of biogenic amines are associated with cancer cell proliferation.
Epinephrine and NE signaling exhibit tumor suppression or cancer progression depending on the immune state
Mounting evidence supports the bidirectional role of Epi and NE in antitumor and protumor in different cancers. Pheochromocytoma is a chromaffin cell tumor induced by excessive production of catecholamines. Catecholamines and their metabolites in urine or plasma could assist in the diagnosis of pheochromocytoma (Lenders et al, 2002). Psychosocial stress, exercise, and receptor agonists and antagonists such as β-adrenergic receptor (β-AR) blockers affect the concentrations of catecholamines. Psychological stress can cause immune dysfunction, thus inducing tumorigenesis and progression (Schoemaker et al, 2016).
Chronic stress-induced immunosuppressive microenvironment promoted the proliferation and activation of myeloid-derived suppressor cells (MDSCs) via adrenergic signaling in a mouse breast tumor model (Mohammadpour et al, 2019). Environmental eustress via an enriched environment activated the antitumor immunity of CD8+ T cells through the sympathetic nervous system (SNS)/β-AR/CCL2 pathway and exerted a better response to PD-L1 immunotherapy (Liu et al, 2021a). Moderate- to high-intensity exercise can upregulate peripheral NE and Epi concentrations to control breast cancer development via the Hippo signaling pathway (Dethlefsen et al, 2017).
In addition, voluntary running was shown to inhibit tumor growth by activating NK cells via Epi and IL-6. In further support of the antitumor effect of Epi, daily intraperitoneal injection of low-dose (0.5 mg/kg) Epi to B16 tumor-bearing mice could mobilize NK cells and β-blockade to blunt tumor growth (Pedersen et al, 2016). In addition, retrospective epidemiological and preclinical studies reported that β-AR blockers could improve the survival of melanoma patients, and enhance the antitumor efficacy of anti-PD-1 inhibitors in mice (Kokolus et al, 2018).
DA inhibits tumor growth and enhances antitumor immunity
DA is a precursor for synthesizing Epi and NE. It is also another catecholamine neurotransmitter, and exerts neuroimmune communication through its receptors in immune cells (Pinoli et al, 2017) (Fig. 2B). DA could improve the chemotherapeutic efficacy for pancreatic cancer in vitro and in vivo by inhibiting the PKA/p38 signal pathway to suppress the polarization of M2 of tumor-associated macrophages (TAMs) (Liu et al, 2021b). In glioma, DA induced caspase-dependent apoptosis and p50/p65 NF-κB signaling pathway to inhibit tumor growth in vivo (Lan et al, 2017).
DA was also reported to inhibit vascular permeability factor (VPF)/vascular endothelial growth factor (VEGF)-induced angiogenesis and delayed tumor growth through D2 receptors in sarcoma tumor-bearing mouse model (Chakroborty et al, 2008). In addition, human T cells express all DA receptors. Dopamine 4 receptor (D4R) agonists exerted the best to activate and rejuvenate T cells. Furthermore, the low concentration of 10 nM DA could decrease PD-1 expression in T cells and induce the proliferation of T cells in vitro (Levite, 2021).
5-HT acts as an immunomodulator in most cancers
5-HT synthesized from the amino acid tryptophan in the brain and the ECs of the intestine is thought initially to increase vascular tone (Walther et al, 2003). 5-HT is absorbed from the bloodstream by platelets and released after activation (Mohammad-Zadeh et al, 2008; Schoenichen et al, 2019).
The protumorigenic effect of 5-HT has been previously shown in lung, bladder, and colon cancer (Cattaneo et al, 1993; Nocito et al, 2008; Siddiqui et al, 2006). For example, 5-HT promotes cell proliferation in multiple tumors through different 5-HT receptor (5-HTR) types (Jiang et al, 2020; Liu et al, 2019) (Fig. 2C). In addition, dysregulated 5-HT signaling has been experimentally observed in epithelial tumors (Sarrouilhe et al, 2015). 5-HT /HTR1E signaling also plays a vital role in preventing ovarian cancer progression promoted by chronic psychological stress (Qin et al, 2021) (Fig. 2C).
Most immune cells express 5-HTR on their cell membranes, as well as other serotonergic components, including tryptophan hydroxylase 1 (TPH1) serotonin transporter (SERT), monoamine oxidase (MAO) (Millan et al, 2008). Most of these 5-HTR affect the activation or inhibition of immune response (Ahern, 2011; Grimaldi et al, 1998). In the intestinal mucosa, 5-HT can act as a proinflammatory or anti-inflammatory tissue protection signal molecule by activating 5-HT7 or 5-HT4 receptors, respectively (Spohn and Mawe, 2017).
It was reported that depleting peripheral 5-HT could downregulate PD-L1 expression on cancer cells, elevate tumor-infiltrating CD8+ T cell accumulation, and delay tumor growth in pancreatic and colorectal cancer mouse models. In addition, the 5-HT inhibitors could augment the effects of anti-PD-L1 inhibitors and induce long-term tumor control (Schneider et al, 2021). Taken together, 5-HT is a key immunomodulation factor in most cancers.
Epi, NE, DA, and/or 5-HT are implicated in tumor growth and progression of multiple cancer types (Fig. 2C). Nevertheless, how the fluctuation of these neurotransmitters in the tumor microenvironment affects cancer progression and how to translate this knowledge into the clinical environment are worthy of further exploration. Further preclinical and clinical data may reveal the molecular mechanisms by which autonomic neurotransmitters regulate cancer at the system and microenvironment level.
Amino acid neurotransmitters in tumor immunity
Role of GABA in tumors
GABA promotes or inhibits tumor growth
GABA mainly acts as an inhibitory neurotransmitter in the mature central nervous system (Li and Xu, 2008). Studies have shown that GABA and its receptors not only exist in nerve tissues (Watanabe et al, 2002) but they also play an essential role in the development of tumors. GABA can promote tumor growth by inducing the polarization of M2 macrophages and limiting the antitumor effect of CD8+ T cells. In contrast, GABA can also exhibit antitumor activity via GABAB receptors (Fig. 3). In humans and animals, GABA receptors were identified on macrophages, dendritic cells (DCs), T cells, and B cells (Alam et al, 2006; Dionisio et al, 2011; Mendu et al, 2012; Tian et al, 2004). Modulation of GABAA receptors has been found to regulate the polarization of macrophages (Januzi et al, 2018), the migration of DCs (Fuks et al, 2012), the secretion of various cytokines (Bhandage et al, 2018), and also the suppression of T cell proliferation (Sparrow et al, 2021; Tian et al, 2019; Tian et al, 1999).

In pancreatic cancer, GABA can promote the proliferation of tumor cells by promoting the upregulation of GABA receptor π subunit (GABRP) expression (Takehara et al, 2007). B-cell–derived GABA limited the antitumor immunity of CD8+ T cells by stimulating the differentiation of IL10+ M2 macrophages through GABAA receptors (Zhang et al, 2021). Moreover, increased GABA in tumor tissues points to a worse prognosis in NSCLC and colon cancer patients.
Further study in the mouse model showed that cancer-cell–derived GABA promoted tumor growth and suppressed the infiltration of CD8+ T cells through β-catenin signaling (Huang et al, 2022a). However, preclinical studies also found that exogenous supplements of GABA exerted an inhibitory effect on tumor growth and metastasis in colorectal, gastric, liver, and NSCLC cells (Al-Wadei et al, 2012; Joseph et al, 2002; Sun et al, 2003). So, GABA modulation may be an effective strategy to improve the response to cancer immunotherapy.
Glutamate stimulates proliferation and migration of tumor cells
Glutamate is the primary excitatory neurotransmitter, which mediates most of the excitatory interaction between different parts of the central nervous system. In addition to its crucial role in the nervous system, glutamate is involved in tumors' occurrence, development, invasion, and migration.
As one of the essential amino acids for protein biosynthesis and the precursor of several other amino acids, glutamate is required for normal and malignant cell growth. Because cancer cells have a higher metabolism than normal tissues due to uncontrolled growth, some genes related to the metabolic chain have been mutated somehow, making cancer cells more dependent on glutamate in various cancers (Yi et al, 2019). Abnormal glutamate signaling can lead to tumorigenesis and may be the mechanism that causes many cancers.
As cancer cells release excess glutamate, nearby cells either die or are transformed to participate in increased glutamatergic signaling by increasing glutamate receptor expression. In this way, cancer cells invade normal tissues and transform normal cell signals (Seidlitz et al, 2009). In glioblastoma, glutamate was increased in the tumor microenvironment. Downregulation of glutamate synergized the response to anti-PD-1 immunotherapy in the GL261 glioma tumor mouse model (Medikonda et al, 2021; Medikonda et al, 2020).
Neuropeptides promote tumor growth by modulating cancer immune microenvironment
Peptide neurotransmitters include but are not limited to substance P (SP), NPY, and CGRP. Apart from traditional roles, the tumor-promoting effects of neuropeptides, especially those of SP and NPY, have been widely reported (Ebrahimi et al, 2020; Garcia-Recio et al, 2013; Moody et al, 2015; Tilan and Kitlinska, 2016). For example, SP mediated by neurokinin-1 (NK-1) receptors is involved in the proliferation, angiogenesis, migration, and invasion of breast cancer (Ebrahimi et al, 2020). NPY is considered to be the growth-promoting and vascularization factor in various cancers, including prostate, breast, and neuroblastoma (Li et al, 2015; Tilan and Kitlinska, 2016). NPY was also reported to modulate immune cell trafficking and alter macrophage function (De la Fuente et al, 1993; Phan and Taylor, 2013; Zhou et al, 2013).
NE-treated Myc-CaP cells promoted the release of NPY, which resulted in the infiltration of myeloid cells and increased IL-6 of TAMs in the prostate tumor microenvironment (Cheng et al, 2019). In addition, the NPY5 receptor (NPY5R) in liver cancer was correlated with tumor growth and survival. NPY promoted the progression of hepatocellular carcinoma by NPY5R activation via TGF-β/NPY/NPY5R axis (Dietrich et al, 2020). Although the oncogenic mechanism of neuropeptides in regulating tumor progression still needs further investigation, the existing evidence strongly supports that neuropeptides inhibition and their receptor blockers may benefit cancer patients.
Gut Microbiota Release Functional Neurotransmitters via Microbial Metabolism
Accumulating evidence supports the notion that gut microbiota potentiates the effectiveness of cancer immunotherapy, especially ICI therapy, mainly through the activation of tumor-infiltrating CD8+ T cells (Gopalakrishnan et al, 2019; Matson et al, 2018; Routy et al, 2018). Microbiota-derived metabolites (induced or produced by the microbiota) play a crucial role in the interactions between gut microbiota and the immune system by modulating antitumor immunity and facilitating the efficacy of cancer immunotherapy. Microbial butyric acid was demonstrated to boost the antitumor effect by enhancing CD8+ T cell immunity in an ID2-dependent manner by IL-12 signaling. Bifidobacterium pseudolongum-derived inosine enhanced the efficacy of immune checkpoint blockade therapy by activating cDC1-dependent Th1 effector function.
In mice monocolonized with B. pseudolongum, treatment with ICIs dramatically elevated inosine concentration in the serum by eight- to nine fold compared with GF mice or mice colonized with bacteria that did not produce inosine (26.16 ± 3.32 μM and 37.5 ± 10.2 μM vs. 3.26 ± 1.01 μM, and 4.8 ± 1.3 μM) (Mager et al, 2020). These findings support the notion that microbial metabolism influences cancer development and response to cancer immunotherapy. Certain gut microbes can produce enzymes to synthesize neurotransmitters and their precursors.
These microbial neurotransmitters can modulate host homeostasis extending beyond the gastrointestinal tract to the entire body, such as the brain, via complex neuronal, immunological, and humoral signaling cascades (Cryan et al, 2019; Fischbach and Segre, 2016). Gut microbiota can also synthesize gaseous neurotransmitters nitric oxide (NO) and hydrogen sulfide (H2S) (Luhachack and Nudler, 2014). Although mechanistic relationships between microbial metabolites and tumor development remain largely unexplored, it may be envisioned that neurotransmitters have the potential to be a new target for cancer immunotherapy (Table 1 and Fig. 4).
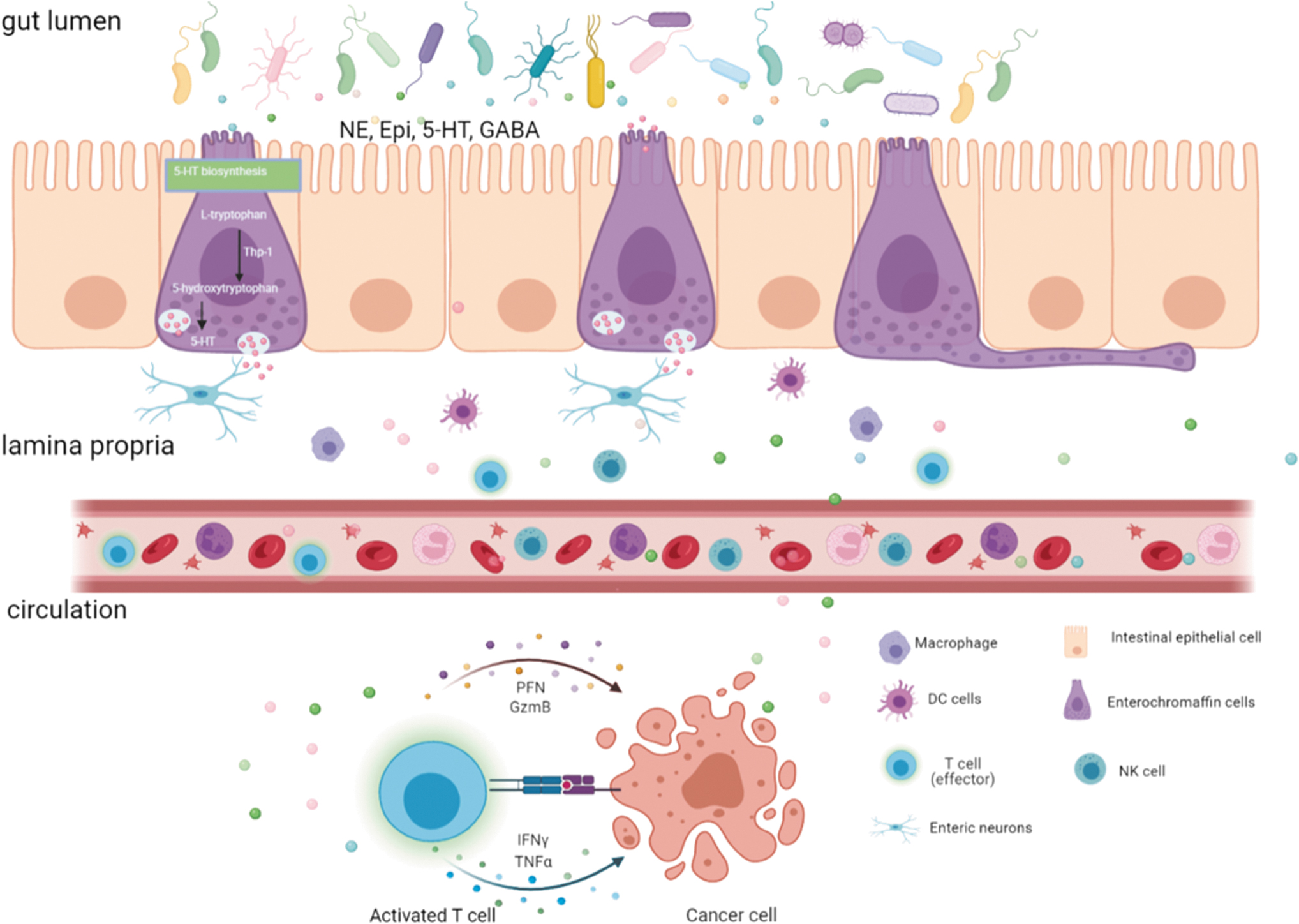
Interactions Between Neurotransmitters and Immune Checkpoint Inhibitors Mediated by Gut Microbiota
DA, dopamine; GABA, γ-aminobutyric acid.
Microbial monoamine neurotransmitters in cancer immunotherapy
Epinephrine and NE
GUS-encoded bacteria can produce biologically active, free forms of NE, Epi, and DA in the gut lumen of mice by comparing the findings among GF mice, SPF mice, and gnotobiotic mice (Asano et al, 2012). In addition, exercise can increase the production of Epi and NE. Exercise could mobilize NK cells via increasing Epi to suppress tumor growth (Pedersen et al, 2016). Other studies reported that >3 h of exercise per week could enrich the abundance of Faecalibacterium prausnitzii, Roseburia hominis, and Akkermansia muciniphila (Bressa et al, 2017).
Interestingly, in patients treated with anti-PD-1 immunotherapy, responders have a relatively high abundance of these bacteria, enhancing the antitumor effect of cytotoxic T cells (Davar et al, 2021; Gopalakrishnan et al, 2018; Routy et al, 2018). Although the direct interactions between these beneficial bacteria and Epi are still unclear, it is interesting to explore whether they can modulate Epi to enhance the antitumor efficacy of anti-PD-1 immunotherapy. Whether the antitumor effect of these bacteria is dependent on Epi/NE remains to be investigated.
Dopamine
DA serves as a precursor of Epi and NE. It was found to remodel tumor blood vessels and improve the delivery of 5-fluorouracil, doubling the amount of the drug in HT29 tumors and increasing its efficacy in chemotherapy (Chakroborty et al, 2011). Peripheral DA exerts antitumor effects via the interactions of DA and immunity, including the activation of CD8+ T cells (Levite, 2016), altering the polarization state of TAMs from M2 to M1 (Qin et al, 2015), inhibiting tumor-infiltrating MDSCs (Hoeppner et al, 2015), and augmenting cytotoxicity of NK cells (Zhao et al, 2013).
Recent evidence suggests that >50% of the peripheral DA has an intestinal source. Tyrosine is the biosynthetic precursor of all three catecholamines. The first step of the reaction pathway is catalyzed by tyrosine hydroxylase (TH) and is rate limiting. This reaction requires oxygen as a cosubstrate and tetrahydrobiopterin as a cofactor to synthesize dihydroxyphenylalanine (DOPA) (Molinoff and Axelrod, 1971). Then, DOPA can be converted into DA by the enzyme dopa decarboxylase (DDC).
Interestingly, gut microbes, such as Enterococcus faecalis (E. faecalis) and Enterococcus faecium (E. faecium), also exhibit TH and DDC activity, indicating the capability to synthesize DA (Wang et al, 2021a). Oral E. faecalis or E. faecium was demonstrated to elevate brain DA levels in mice by converting L-dopa in the gut and blood (Wang et al, 2021a). DA at 10−5 and 10−7 M could downregulate regulatory T (Treg) cell activity, and increase the proliferation of effector T (Teff) cells through DA receptors (D1-R and D5-R) via the ERK signaling pathway (Kipnis et al, 2004).
In a melanoma mouse model, mice monocolonized with E. faecium exhibited the capability to decrease Treg cells and augment Th1 responses to improve tumor control of anti-PD-L1 therapy (Routy et al, 2018). In a pioneering phase I trial of anti-PD-1 immunotherapy by fecal microbiota transplant from responder patients to nonresponders, gut microbiota composition in responders had a higher representation of Enterococcus spp. (Baruch et al, 2021). However, the detailed mechanism is still elusive, and DA may be a potential target for E. faecium to overcome the anti-PD-L1 immunotherapy by activating antitumor immunity.
Elevated 5-HT promotes tumor immune escape
The various effects of 5-HT signaling on immune cells make it a target for studies of tumor immunity. For example, macrophages express 5-HTR2B, and 5-HTR7 signaling stimulates the polarization of M2 macrophages, which promotes tumor development (Nieto et al, 2020; Nocito et al, 2008). 5-HT signaling also promotes activation of Treg cells via suppression of antitumor immunity (Karmakar and Lal, 2021; O'Connell et al, 2006). In the intestinal tract, ∼90% of 5-HT is synthesized by ECs, 50% of which is regulated by gut microbiota. Microbial metabolites, such as SCFAs or secondary bile acids, can act on ECs to induce the transcription of the rate-limiting biosynthetic enzyme, TPH1 of 5-HT.
The effect of gut microbiota on 5-HT production was investigated by measuring peripheral 5-HT levels in conventional mice with normal gut microbiota and GF mice. GF mice have lower 5-HT levels in serum and colon than conventional controls (Yano et al, 2015). 5-HT can act directly or indirectly on the immune system, shaping the microbiota composition and localization. 5-HT can also control gut motility, secretory reflexes, platelet aggregation, bone development, and cardiac function to regulate immune responses.
In mouse models, F. prausnitzii and its supernatant can also modulate intestinal 5-HT levels to protect intestinal integrity (Martin et al, 2015). F. prausnitzii was reported as beneficial bacteria, which was observed to be highly enriched in responders to anti-PD-1 immunotherapy and correlated with higher infiltration of CD8+ T cells in tumors. When performed FMT from responders to the melanoma mouse model, mice with a higher abundance of F. prausnitzii exhibited an improved response to anti-PD-1 immunotherapy (Gopalakrishnan et al, 2018).
Besides, another 5-HT-producing bacterium, Escherichia coli (K-12), was reported to upregulate the expression of PD-L1 in IFN-γ–sensitized HT-29 cells via the NF-κB pathway, which may lead to immune escape (Lee et al, 2020). Collectively, all the evidence indicates that regulating the level of 5-HT by gut microbiota may have an essential role in regulating PD-L1 expression in tumor cells, thus sensitizing response to ICIs.
Microbial GABA signaling augments antitumor efficacy
L-Glutamate and GABA are not only known for their roles as the primary excitatory and inhibitory neurotransmitters in the mammalian central nervous system, respectively. A growing body of evidence suggests that glutamate signaling promotes cancer cell initiation, proliferation, and metastasis through their receptors (Luksch et al, 2011; Stepulak et al, 2014). The GABAergic system may develop an antitumor activity by suppressing cancer cells in the inflammatory response (Jiang et al, 2020). For example, GABA receptor GABRP interacts with KCNN4 inducing an influx of Ca2+ and activating NF-κB signaling. This facilitates macrophage infiltration and tumor progression in pancreatic cancer via the recruitment of CXCL5 and CCL20 in pancreatic cancer (Jiang et al, 2019).
Lower glutamate correlates with a better response to anti-PD-1 immunotherapy in glioblastoma in vivo (Medikonda et al, 2020). As a matter of particular interest, some Bacteroides species can convert glutamate into GABA, which points to a better response to anti-PD-1/CTLA-4 immunotherapy. For instance, Bacteroides thetaiotaomicron and Bacteroides fragilis, the GABA-producing commensals, facilitated the antitumor efficacy of CTLA-4 blockade via the IL-12–dependent Th1 immune response (Vetizou et al, 2015). B. vulgatus and P. distasonis can produce GABA in vitro culture (Strandwitz et al, 2019). Both strains were associated with a better response to anti-PD-1 immunotherapy in NSCLC patients (Huang et al, 2022b).
In addition, Lactobacillus could convert glutamate into GABA (Li and Cao, 2010). Oral administration of live Lactobacillus rhamnosus GG augmented the antitumor activity of anti-PD-1 immunotherapy by increasing tumor-infiltrating DCs and T cells (Si et al, 2022). Lactobacillus reuteri has been reported to alleviate colitis associated with immune checkpoint blockade by inhibiting group 3 innate lymphoid cells (ILC3s) (Wang et al, 2019). A. muciniphila and Pacrabacteroides merdae were found to increase the ratio of GABA/glutamate (Dooling and Costa-Mattioli, 2018).
Notably, previous studies suggested that A. muciniphila could potentiate the response to PD-1 immunotherapy with melanoma, NSCLC, and renal cell carcinoma in an IL-12–dependent manner by recruiting CCR9+CXCR3+CD4+ T cells into tumor sites (Routy et al, 2018). Altogether, these results hint that A. muciniphila may elicit a synergistic antitumor effect via GABA signaling.
However, there is still no study to examine the direct interaction mechanisms between microbial neurotransmitters and cancer immunotherapy. Further research is needed to identify the mode of action of microbial neurotransmitters.
Neurotransmitters Signaling or Receptors as Emerging Targets for Cancer Therapy
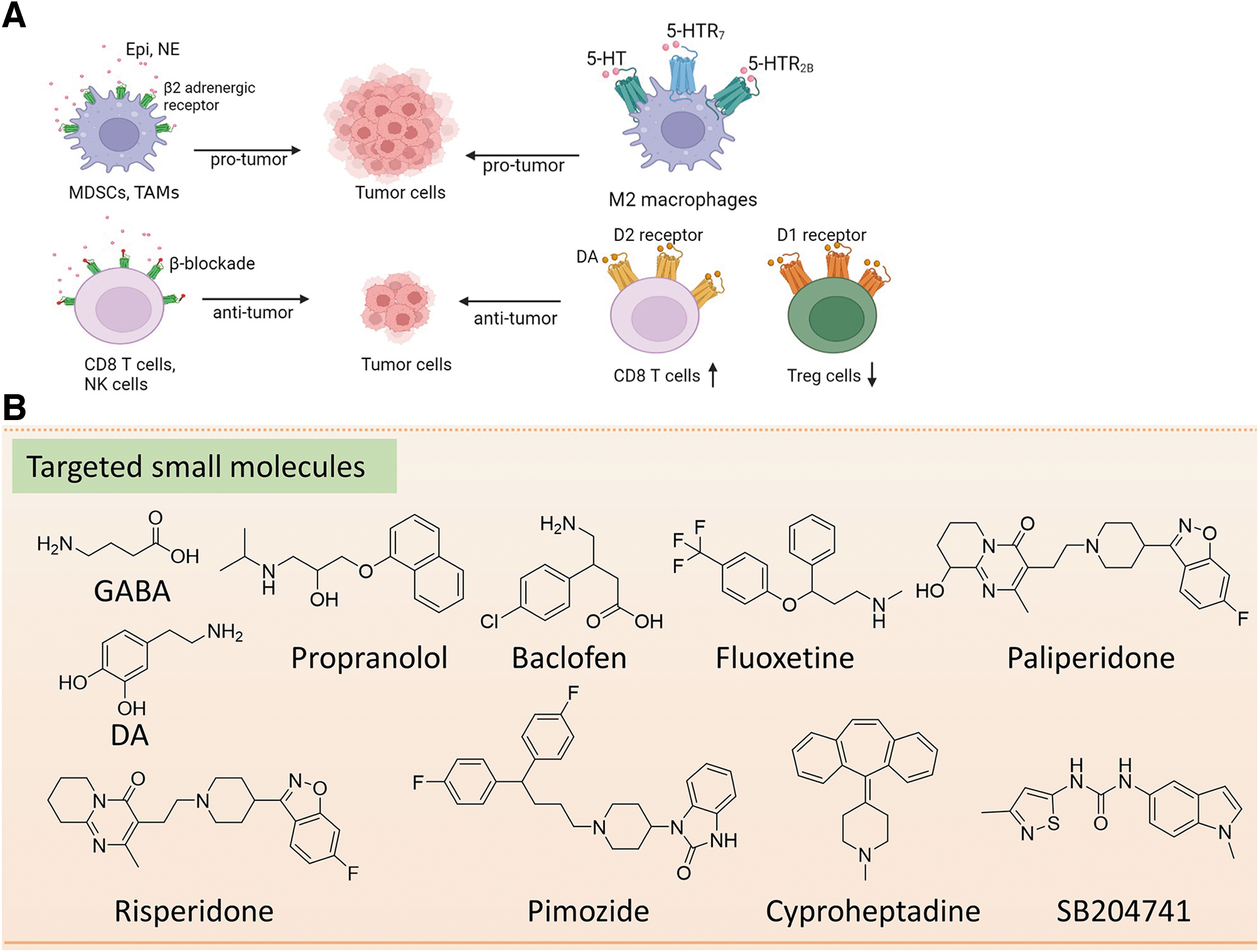
A variety of neurotransmitter receptors, expressed on nerve fibers or tumor-infiltrating immune cells, are critical for tumorigenesis. Thus, antagonists or agonists of their receptors can be scrutinized for their use in cancer therapy (Jiang et al, 2020) (Fig. 5).
Early studies have linked gut microbes with neurobehavioral phenotypes and proposed the concept of the “microbe–gut–brain” axis (Cryan et al, 2019). Mounting evidence supports that certain microorganisms can synthesize and/or regulate various neurochemical substances to influence neurotransmission. They can also synthesize and/or regulate other environmental metabolites to directly or indirectly affect neuronal activity (Jameson et al, 2020; Schledwitz et al, 2021) (Figs. 4 and 5). Thus, the role of microorganisms in regulating neuronal communication through the gut–brain axis is of great significance and has been actively elucidated as a novel therapeutic target for cancer treatment.
Neurotransmitters and their analogs for cancer therapy
The discovery of microbial synthesis of neurotransmitters recapitulates the first discovery of chemical transmitters in the early 20th century (Jameson et al, 2020; Valenstein, 2002). GABA, NE, DA, and 5-HT are also produced by cultured bacteria (Huang and Wu, 2021; Strandwitz, 2018). Recent studies have identified GABA as a potential tumor suppressor for small airway-derived lung adenocarcinoma (Schuller et al, 2008b).
Likewise, DA treatment controls tumor growth by inhibiting angiogenesis (Chakroborty et al, 2004) or regulating diverse immunocompetent cells within the tumor microenvironment (Nasi et al, 2019) (Table 2). The role of small molecule neurotransmitters in tumors is detailed above. Given a potential link between neurotransmitters and antitumor immune response, we next summarized the current development of neurotransmitter receptors in cancer treatment.
Microbial-Derived Neurotransmitters and Small Molecules Targeting Neurotransmitter Receptors for Cancer Therapy
Treg, regulatory T cell.
Targeting neurotransmitter receptors for cancer treatment
Targeting β-AR
Adrenergic receptors α or β (AR-α or β) are widely expressed in most mammalian tissues and cells. β-ARs are expressed in T suppressor lymphocytes, NK cells, B lymphocytes, DCs, and macrophages (Sarkar et al, 2013), which mediate the suppression of antitumor immune response and regulate immune cells within the tumor microenvironment.
At present, epidemiological studies generally support the notion that cardiovascular patients treated with β-AR antagonists, such as propranolol, have a reduced cancer incidence (Grytli et al, 2014; Santala et al, 2019) (Table 2). Preclinical studies revealed that cold-induced stress is sufficient to elevate NE. Propranolol could reduce tumor growth by increasing the secretion of functional cytokines of tumor-infiltrating CD8+ T cells and downregulating the expression of PD-1 on CD8+ T cells in B16-OVA and 4T1 tumor-bearing mouse models.
Furthermore, combination treatment of propranolol and anti-PD-1 inhibitor significantly delayed 4T1 tumor growth by increasing the level of IFN-γ+ CD8+ T cells (Bucsek et al, 2017). In addition, a retrospective analysis revealed that metastatic melanoma patients who received β-AR blockers prolonged overall survival after at least one immunotherapy (IL-2, anti-CTLA-4, and anti-PD-1).
Moreover, the in vivo results further supported the notion that combining β-AR blockers with anti-PD-1 or anti-PD-1/IL-2 could improve tumor control and prolonged survival in the melanoma mouse model (Kokolus et al, 2018). The prospective phase I clinical trial of propranolol and anti-PD-1 ICIs in metastatic melanoma patients provided preliminary evidence of antitumor activity based on safety and tolerability (Gandhi et al, 2021).
Collectively, these data implicate that targeting NE/β-AR signaling is an attractive strategy to improve the efficacy of anti-PD-1 immunotherapy. Still, further data are required experimentally and clinically to classify the potential mechanisms.
Targeting DA receptor
DA exerts its function by binding to D1-like (D1 and D5) DA receptors and D2-like (D2, D3, and D4) receptors. Most immune cells, including monocytes, NK cells, macrophages, DCs, neutrophils, B cells, effector and Treg cells, express DARs (Zhang et al, 2017). DA could directly activate Teff cells at the physiological concentration of ∼10 nM by binding to D2-like receptors or suppress Treg cells via D1-like receptors.
In addition, DA can selectively induce the adhesion, migration, and homing of CD8+ T cells (Levite, 2016). DAR agonists seem to exert inhibitory effects on the growth of various tumors, including gastric cancer, breast cancer, and sarcoma (Roney and Park, 2018). However, whether this anticancer effect is associated with the enhancement of immunity is unclear, and it may provide broad implications for the modulation of DA signaling for cancer immunotherapy.
Targeting 5-HTR
5-HT performs multiple functions by interacting with its receptors. To date, 15 different receptor subtypes have been identified to modulate a wide range of functions on stromal, cancer, and immune cells. The importance of the 5-HT signaling in tumors and the immune system has been well studied by different agonists and antagonists of 5-HTR.
A recent study found that pancreatic cancer cells increase the expression of receptor 5-HT2B to allow glycolysis in tumors under metabolic pressure, thereby promoting tumor growth (Jiang et al, 2017). Likewise, selective inhibition of the 5-HT2B receptor can inhibit tumor angiogenesis by inhibiting endothelial NO synthase and p-ERK1/2 (Asada et al, 2009). Selective inhibition of 5-HT reuptake has been shown to induce anticancer effects by affecting cytokine secretion in mouse models of melanoma (Grygier et al, 2013) (Table 2). The 5-HTR5A agonist, valerenic acid, could attenuate glioblastoma cell growth in vitro and in vivo by increasing innate immune signals, including elevating intracellular ROS and activating AMPK signaling (Lu et al, 2020).
The 5-HTR7 inhibitors paliperidone, risperidone, and pimozide are also being studied to treat glioblastoma (Kast, 2010). Cyproheptadine, a 5-HT antagonist, can inhibit the proliferation of urothelial cancer cells by inhibiting mTOR and β-catenin signaling pathways (Hsieh et al, 2016). In addition, SB204741, a recent specific antagonist of 5-HTR2B, has shown a significant inhibitory effect on tumor growth of pancreatic cancer by inhibiting the Warburg effect (Jiang et al, 2017).
In addition, 5-HTR2B and 5-HTR7 signaling have been reported to promote tumor development through the polarization of M2 macrophages (de las Casas-Engel et al, 2013). 5-HT could boost the expression of PD-L1 in the cancer cell to help the immune escape. Using 5-HT synthesis inhibitors efficiently slowed tumor growth in colon and pancreatic cancers in vivo. And when combined with anti-PD-L1, the tumors were even eliminated (Schneider et al, 2021). Hence, 5-HT signaling is a promising target for cancer immunotherapy.
Targeting GABA receptor
Many tumor tissues have identified ionotropic GABAA, GABAC receptors, and metabotropic GABAB receptors (Kanbara et al, 2018; Sung et al, 2017). These GABA receptors play an essential role in cancer cell proliferation and migration. It is worth noting that GABAA receptors are upregulated in breast, prostate, and pancreatic cancer (Gumireddy et al, 2016; Jiang et al, 2020), while GABAB receptors are downregulated in pancreatic and liver cancers (Schuller, 2018).
Interestingly, in most cases, GABA stimulates the proliferation of cancer cells through the GABAA receptor pathway, whereas it inhibits the growth of cancer cells through the GABAB receptor (Jiang et al, 2020; Zhang et al, 2014). Studies have found that GABAA receptor agonists increase the proliferation of gastric and pancreatic cancer cells by activating the MAPK signaling pathway (Takehara et al, 2007).
In contrast, activation of GABAB receptors inhibits isoproterenol-induced phosphorylation of ERK1/2 to prevent DNA synthesis and cell migration effectively (Schuller et al, 2008b). Moreover, the activation of GABAB receptors by GABA or baclofen can serve as potential targets for cancer treatment (Nieto et al, 2022; Schuller et al, 2008a) (Table 2). Thus, the specific type of cancer or GABA receptor type may lead to different effects of GABA activation on cancer growth/migration.
In addition, since GABA exists in the tumor microenvironment, it may increase the possibility that GABA can regulate the inflammatory response by targeting infiltrating immune cells (Jiang et al, 2020). It may also affect various functional properties of immune cells. The π subunit of the GABAA receptor promotes the progression of pancreatic cancer by regulating KCNN4-mediated Ca2+ in a GABA-independent manner (Jiang et al, 2019). GABRP, a subunit of GABAA, can upregulate the expression of CXCL5 and CCL20 to regulate the recruitment of macrophages in pancreatic cancer (Jiang et al, 2019).
A recent study revealed that B cell-secreted GABA limited the antitumor immune response and controlled the polarization of tumor-infiltration macrophages via IL-10 in the MC38 CRC mouse model. When blocked GABAA receptor or reduced GABA, the tumor growth was controlled with enhanced cytotoxic CD8+ T cells in the tumor microenvironment (Zhang et al, 2021). Altogether, this evidence provides a rationale for manipulating GABA signaling to enhance the efficacy of cancer immunotherapy.
Glutamate receptors play a vital role in the biological effects of glutamate. Extracellular glutamate binds to α-amino-3-hydroxy-5-methyl-4-isoxazole propionic acid (AMPA) receptors (AMPAR) and activates signaling pathways, such as KRAS-MAPK and PI3K, to promote cell proliferation, survival, and migration (Yi et al, 2019). In this way, glutamate can increase the invasion and migration of pancreatic cancer cells (Herner et al, 2011).
Recently, another type of glutamate receptor, N-methyl-D-aspartate (NMDA), has also been shown to activate neural signaling pathways and affect breast cancer brain metastasis (Zeng et al, 2019). The breast-to-brain metastasis (B2BM) activated by NMDA plays a crucial role in spreading metastasis via neuronal signaling pathways (Arvanitis et al, 2020; Zeng et al, 2019). The effects of glutamate on cancer cells are collectively mediated through glutamate metabolism and glutamate receptors.
Current Challenges and Future Directions
Cancer results from a combination of genetic and environmental factors (Hanahan and Weinberg, 2011); it is not surprising that intestinal microbial neurotransmitters play essential roles in cancer occurrence, development, and treatment. While the evidence for interaction between these regulatory elements is overwhelming, the question of how the signals conveyed by neurotransmitters in the gut reach their targets in tumors is still a mystery. Exceptions are colon and other intestine cancers, for which direct interaction between neurotransmitters and cancer cells is possible.
The enteric nervous system (ENS) is a complex network involving ∼500 million neurons (Furness, 2012). It is often referred to as the “second brain.” The ENS can function autonomously (Travagli et al, 2003), but it interacts with the brain and the autonomic nervous system via the vagus nerve (Cryan et al, 2019). Some neurons are well positioned to directly sense neurotransmitters and neuromodulators of microbial origin, such as 5-HT (Li et al, 2012; Watanabe et al, 2002).
Besides, bacterial ion channels have functional roles in information signaling and processing, which needs membrane potential (Vmem) to provide free energy across their membranes. For example, potassium ion channel YugO mediates electrical signaling within a Bacillus subtilis biofilm (Prindle et al, 2015). This ion channel mediates bacterial communication through glutamate metabolism within a biofilm (Liu et al, 2015). These studies suggest that membrane potential could be dynamic and mediate signaling like neurons (Benarroch and Asally, 2020).
Also, it is well established that the brain and the peripheral nervous system strongly influence the immune system and its role in the tumor microenvironment (Bahrambeigi et al, 2019; Chen et al, 2021; He et al, 2021; Karmakar and Lal, 2021; Levite, 2016; Liu et al, 2019; Nasi et al, 2019; Pinoli et al, 2017; Sarkar et al, 2013). It closes the link between signals originating in the gut and those acting on cancers and their microenvironment.
A robust multidisciplinary approach will be required to better understand the communications among gut microbiota, neurotransmitters, and antitumor immunity (Fig. 6). For example, functional metagenomics can help reveal potential mechanisms of host–microbial interactions by identifying effector molecules and functionally essential genes in the human microbiome (Cohen et al, 2015). To further define the role of microbiota-encoded effectors in host–microbial interactions, synthetic biology (such as CRISPR-Cas9 genome editing or in situ microbiome engineering) and bioinformatic analysis of gut microbiome sequencing data were of great significance (Cohen et al, 2017).

Recently, CRISPR-Cas9 genome editing has been applied to Lactobacillus plantarum, Clostridium beijerinckii, and Clostridium difficile (Leenay et al, 2018; Li et al, 2016; Wang et al, 2015). An engineered bacterium was designed to specifically lyse within the tumor microenvironment and activate tumor-infiltrating T cells to reduce tumor growth in a syngeneic tumor-bearing mouse model (Chowdhury et al, 2019), providing proof of concept for engineered bacterial immunotherapy.
In addition, in situ microbiome engineering can be used to manipulate the gut microbiota to generate specific neurotransmitters instead of culturing a specific strain and modifying it in the laboratory (Sheth et al, 2016). Collectively, synthetic biology tools combined with modulation of bacteria's electrical response by neurotransmitters may provide a new approach to modulate bacterial functions such as metabolic reactions, signaling, and gene expression, regarding its capacity to construct new signaling and metabolic pathways or redesign pathways in biological systems (Selberg et al, 2018).
Moreover, the antitumor effect of the modified microbes can be further elucidated by FMT in vivo (Sheth et al, 2016). Besides, culture-independent metagenomic approaches empower to reveal the potential implications for human health. However, <17% of species of human gut microbiota have been cultured (Hugon et al, 2015). Thus, culturomics was developed to culture, identify, and extend our understanding of unknown bacteria.
Although we have taken significant steps in gut microbiota genetic modification, biological and ecological knowledge gaps still need to be filled, which prevent its immediate application. However, it is challenging to evaluate these neurotransmitter-regulatory strategies to modulate native ion-channel circuits and different signaling pathways. The current lack of datasets for physiomic profiling is one of the significant barriers. Nevertheless, such innovative research should be promoted by encouraging crossdisciplinary collaborations between biophysicists, biochemists, and clinicians.
Footnotes
Authors' Contributions
E.L.-H.L. and J.H. designed and conceived the review. E.L.-H.L., J.H., Junmin Zhang, Juanhong Zhang, L.M., and H.Y. wrote the article. M.W., Y.Z., Z.M., E.N., X.Y., and E.L.-H.L. revised, extended, and provided final proof of the article. All authors reviewed the article.
Author Disclosure Statement
No financial or commercial conflict of interest was disclosed.
Funding Information
This work was funded by Dr. Neher's Biophysics Laboratory for Innovative Drug Discovery (Grant No. 001/2020/ALC) and regular grants (File No. 0096/2018/A3 & 0111/2020/A3, 0015/2019/AMJ, and 0114/2020/A3) supported by Macao Science and Technology Development Fund, Macau (SAR) China, 2020 Young Qihuang Scholar funded by National Administration of Traditional Chinese Medicine, the National Natural Science Foundation of China (82003779 and 82001995), the Natural Science Foundation of Gansu Province (20JR10RA010), the Macao Young Scholars Program (AM201926 and AM2020018), FDCT Funding Scheme for Postdoctoral Researchers of Higher Education Institutions (0017/2021/APD), and the Science, Technology and Innovation Commission of Shenzhen Municipality (ZDSYS20190902093601675). This work is also financially supported by the Start-up Research Grant of University of Macau (SRG2022-00020-FHS) and the Faculty of Health Science, University of Macau.