
Abstract
Significance:
The ultimate manifestations of life, birth, survival under various environmental pressures and death are based on bioenergetics. Hibernation is a unique survival strategy for many small mammals that is characterised by severe metabolic depression and transition from euthermia to hypothermia (torpor) at body temperatures close to 0°C. These manifestations of life were made possible by the remarkable “social” behavior of biomolecules during billions of years of evolution: the evolution of life with oxygen. Oxygen was necessary for energy production and the evolutionary explosion of aerobic organisms.
Recent Advances:
Nevertheless, reactive oxygen species, formed through oxidative metabolism, are dangerous—they can kill a cell and, on the other hand, play a plethora of fundamentally valuable roles. Therefore, the evolution of life depended on energy metabolism and redox-metabolic adaptations. The more extreme the conditions for survival are, the more sophisticated the adaptive responses of organisms become. Hibernation is a beautiful illustration of this principle. Hibernating animals use evolutionarily conserved molecular mechanisms to survive adverse environmental conditions, including reducing body temperature to ambient levels (often to ∼0°C) and severe metabolic depression. This long-built secret of life lies at the intersection of oxygen, metabolism, and bioenergetics, and hibernating organisms have learned to exploit all the underlying capacities of molecular pathways to survive.
Critical Issues:
Despite such drastic changes in phenotype, tissues and organs of hibernators sustain no metabolic or histological damage during hibernation or upon awakening from hibernation. This was made possible by the fascinating integration of redox-metabolic regulatory networks whose molecular mechanisms remain undisclosed to this day.
Future Directions:
Discovering these molecular mechanisms is not warranted only to understand hibernation in itself but to help explain complex medical conditions (hypoxia/reoxygenation, organ transplantation, diabetes, and cancer) and to even help overcome limitations associated with space travel. This is a review of integrated redox-metabolic orchestration in hibernation. Antioxid. Redox Signal. 40, 345–368.

Introduction
During billions of years of Earth's evolutionary history, life has adapted and changed in response to numerous environmental challenges. As a result of these adaptive processes, evolution gave rise to various life forms, including diverse anaerobic and aerobic, unicellular and multicellular organisms. During evolutionary history, the development and survival of new life forms primarily depended on their intrinsic capacity to adapt to ever-changing environmental conditions. This was also true for aerobic multicellular organisms from the moment that oxygen appeared at substantial levels in the atmosphere, or more precisely, the appearance of photosynthetic cyanobacteria that produced it.
The evolution of life with oxygen led to the vast expansion of aerobic organisms, guided by efficient bioproduction of chemical energy in the form of adenosine triphosphate (ATP), with oxygen as the terminal electron acceptor in the process of mitochondrial oxidative phosphorylation. The high rate of energy production provided by oxygen is the basis for diversity at the level of biomolecules and more complex biological entities such as organelles, cells, organs, and organisms.
Bioenergetic pathways ensured the maintenance of complex structures at all levels of the biological organization by virtue of highly adaptive metabolic capacities necessary for growth, reproduction, and the evolutionary explosion of aerobic organisms (Carter, 2003; Halliwell and Gutteridge, 2015).
In addition to the paradox that a by-product of cyanobacterial metabolism gave rise to oxygen-based life, aerobic life paradoxically depends on potentially harmful by-products of oxidative metabolism, reactive oxygen species (ROS); primarily, superoxide anion radical (O2 •−), hydrogen peroxide (H2O2), and hydroxyl radical (OH•). However, certain ROS (primarily H2O2 and possibly O2 •−) are essential signaling molecules that regulate vital physiological processes. Precisely for this reason, we look at antioxidant defense (AD) as an adaptive evolutionary response that eliminates ROS, halts oxidative damage to biomolecules, and prevents oxidative stress development as an integrative part of the homeostatic signaling system in redox biology.
In accordance with Lehninger's (Fisher, 2001): nothing in evolution exists to play a single role, redox-dependent transcription factors emerged and diverged from the evolution of life with oxygen as an integrative link between redox and metabolic regulation of fundamental physiological processes. The evolution of nuclear factor erythroid 2-related factor 2 (Nrf2) role as a master regulator of redox-metabolic integration is one striking example of this principle. Thus, the adaptive potential of aerobic organisms was increased by uniting seemingly paradoxical pressures into an essential evolutionary driving force. For details, see excellent review papers (Gutteridge and Halliwell, 2018; Taverne et al., 2018).
The paramount molecular principles of redox-metabolic integration that evolved globally on Earth are maintained and persist in all aerobic (and anaerobic) cells. On the molecular level, the differences are evident in the specific protective mechanisms against free radicals during adaptation to extreme environmental conditions, such as hibernation.
Fascination and the Beauty of Sustaining Life Through the Lens of Hibernation
The beauty and fascination by fundamental physiological principles drive research in the field of hibernation through increasing understanding of underlying molecular mechanisms of bioenergetics and energy metabolism. To achieve homeothermy, that is, maintain constant body temperature at ∼37°C, mammals and birds evolved the unique ability to generate body heat at the expense of cellular biochemical reactions; that is, endothermy (Prosser, 1991). In terms of bioenergetics, endothermy is quite expensive, which is why the metabolic rate in mammals is four to seven times higher than those in respectively sized reptiles.
Maintaining constant body temperature requires allocating more than two-thirds of the total metabolic energy toward heat production (Carey et al., 2003). Extreme environmental conditions such as subzero temperatures and food scarcity make it virtually impossible for homeotherms to survive without a well-designed coping strategy. Under such constraints, the only path toward survival for many mammals is through hibernation, which requires abandoning homeothermy and allowing body temperature to fall near-ambient levels (Carey et al., 2003; Mohr et al., 2020; Storey, 2010). In this way, hibernation saves enough energy to sustain life until spring (Fig. 1).

The first reported knowledge of hibernating mammals dates back to Aristotle and Schneider (384-322 BC) (1878). Hibernation is a broad term and generally refers to a wide class of heterothermic organisms that, especially in the winter months, reduce their metabolic rate. Several different groups of mammals can hibernate, ranging from rodents and certain primates to bears (Geiser, 2004). These animals are less active in winter, when a lethargic (dormancy) state, hypometabolism, hypothermia, and a decrease in heart rate are the most important manifestations of such a condition (Giroud et al., 2021; Staples, 2014).
The spectrum of these changes is relative and species specific. Some of the best studied species among hibernators are the hedgehog, marmot, bat, and ground squirrel (Geiser, 2004; Lyman and Chatfield, 1955; Wu and Storey, 2012). For example, the brown bear hibernates in a dormant state in winter, but its body temperature does not radically decrease (∼30°C), nor does the degree of metabolic depression (Lyman and Chatfield, 1955).
On the contrary, in many hibernating small mammals and one type of bird, this state is defined as deep hibernation, that is, torpor, with a radical decrease in body temperature and metabolic rate lasting from several days to several weeks (Carey et al., 2003; Storey and Storey, 2004; Storey et al., 2010). Hence, the term hibernation in this review is used as a synonym for deep hibernation, that is, torpor.
The most studied hibernating species is the ground squirrel. In times of hibernation, these “warm-blooded” animals behave as “cold-blooded” organisms, sleeping through the unfavorable conditions of winter and waking up in the spring ready for reproduction (Lyman and Chatfield, 1955). This multimonth hibernating period during the winter is characterized by periodic phases of awakening from torpor (torpor–arousal cycles) when the metabolic rate increases several times and the organism returns from hypothermia to normothermia.
Both the state of torpor and awakening from it are characterized by a unique metabolic transition at the level of the organism, organs, tissues, and cells, supported by redox-metabolic reprogramming (Carey et al., 2003; Sheriff et al., 2013; Storey et al., 2010; Wei et al., 2018). Metabolic depression and hypothermia are characteristics of the hibernation phase (winter), whereas normal (or increased) metabolic activity and euthermia are inherent to other circannual phases: spring, when reproduction occurs shortly after awakening from hibernation; summer, when hibernators intensively feed, accumulate fuel reserves, and prepare to enter hibernation in late autumn; and winter, when hibernators cycle through long periods of torpor interrupted by brief periods back in euthermia (Fig. 2) (Storey, 2010; Storey and Storey, 2005).
Hibernation is an evolutionarily conserved survival strategy in extreme environmental conditions (Bouma et al., 2012). Accordingly, hibernation is a seasonal phenomenon dictated by a circannual rhythm and entrained by photoperiod (Mohr et al., 2020; Storey et al., 2010). Specific stimuli and molecular mechanisms involved in the preparation, entry, and awakening from hibernation are still poorly understood. In general, these stimuli include external, exogenous and internal, endogenous stimuli. Exogenous stimuli mainly include photoperiod, food scarcity, and low temperatures (Mohr et al., 2020).
Endogenous stimuli are in sync with photoperiod, and include the central nervous system (especially the suprachiasmatic nucleus), hypothalamus, endocrine activity, and gonadal development (Armitage and Shulenberger, 1972), which are all under the control of the endogenous circannual clock (Davis, 1976; Kenagy, 1980). Life cycle and circannual rhythm (propagated by endogenous and environmental signals) direct the recurring pattern of metabolic reorganization and redox-metabolic integration in hibernators.
Among the most intriguing regulatory mechanisms are the ones that enable hibernators to quickly and without consequences occasionally wake up from torpor (deep phase of hibernation), and rewarm their bodies to enter a brief phase of euthermia, achieving complete metabolic and body temperature restitution (Mokrasch et al., 1960). These processes occur without negative consequences for tissues and organs with no detectable damage on the histological and molecular level, even though the organism repeatedly oscillates between the radically different physiological states of hypothermia in hibernation and euthermia upon awakening.
Remarkably, hibernators are resistant to ischemia/reperfusion injury, oxidative damage, cold (hypothermia)-induced tissue damage, inflammation, thermal stress, metabolic stress, and their cytoprotection is enabled through antiapoptotic signaling (Rouble et al., 2013). It is important to note that the organs of hibernators are as susceptible to metabolic damage in the spring as they are in nonhibernating species, which indicates that the redox-metabolic mechanisms that support hibernation are not present in the organs of hibernators all year round (Storey, 2010). Therefore, metabolic adaptations must be induced just before the onset of hibernation or as a direct response to entering torpor (Storey, 2010).
Suspended animation is enabled by a plethora of metabolic adaptations, most of which require changes at multiple levels of regulation (Biggar and Storey, 2014; Carey et al., 2003; Morin and Storey, 2009). By hibernating, small mammals can save >90% of the energy that would otherwise be allocated toward maintaining a constant body temperature by allowing a drop to near-ambient temperature, sometimes as low as 0°C–5°C (Barnes, 1989). Metabolic rate can be reduced to as little as 1%–5% relative to basal metabolic rate in euthermia, and heart rate can drop from 350–400 beats per minute to only 5–10 beats per minute.
A similar depression of breathing occurs that may decrease from >40 cycles per minute to less than one cycle per minute in hibernation, with more extended periods of apnea (McArthur and Milsom, 1991; Zatzman, 1984). In addition, organ perfusion can drop to <10% of the average during euthermia; for example, blood flow to the brain can be reduced by 90% (Storey, 2010). The function of several organ systems is also severely suppressed, such as renal and digestive functions and immune and inflammatory responses (Carey et al., 2003; Storey et al., 2010).
On the molecular level, transcription- and translation-related processes are suppressed. Downregulation of the processes mentioned above results in a distinct systematic suppression of the euthermic physiological phenotype (Storey and Storey, 2004; Zatzman, 1984). These tissue responses represent a general principle operating at the level of the whole organism, whereas tissue-specific variations serve particular purposes. This sophisticated response is evolutionarily conserved and written in genes and behavior.
Coordinated metabolic suppression that, in fact, represents a newly established homeostasis in hibernation is made possible by establishing complex yet robust mechanisms of biochemical control. Initiation of specific adaptations on genetic, epigenetic, transcriptional, translational, and post-translational levels creates the necessary framework for cells and tissues to maintain viability, and function under systemic hypothermia and hypometabolism (Staples, 2014). The result is the stabilization of vital cellular functions and metabolic rearrangements that are established during torpor and re-established immediately upon awakening, all of which is supported by specific adaptations at the level of redox regulation (Buzadzic et al., 1990; Petrovic et al., 1983).
Endocrine activity in hibernators shows some general patterns, with species-specific differences in the way of providing energy substrates before entering hibernation (fat storing or food storing), body weight gain (spring and summer when the reproductive period ends), body weight loss (immediately before entering and during hibernation), sex, as well as torpor duration (Dark, 2005; Giroud et al., 2021). Lyman and Chatfield (1955) showed at the histological level that thyroid and adrenal gland activities are suppressed during hibernation, that ovaries and testes atrophy before entering hibernation and start to activate only up to several days before coming out of hibernation (woodchuck and ground squirrel).
Boswell et al. (1994) monitored seasonal variations in body weight and insulin, cortisol, and corticosterone levels in golden-mantled ground squirrels (Spermophilus saturatus) during the active season from April to August. The authors found that, regardless of sex, the insulin concentration is maximal before entering hibernation (at the end of August, when body weight is the highest), which is followed by higher concentration of adrenal glucocorticoids to support the anabolic effect of insulin. This insulin peak (hyperinsulinemia) is followed by increased insulin resistance in peripheral tissues, especially in white adipose tissue (WAT).
Buck et al. (2002) also found that the highest level of circulating insulin in 13-lined ground squirrels (Ictidomys tridecemlineatus) is in the autumn before entering hibernation (four times higher compared with spring euthermic animals), that it continues to increase slightly in the first months of hibernation, but during late hibernation, it returns to the basal level, which is maintained in the phases of awakening from torpor. Cortisol concentration is the highest after coming out of hibernation (April), decreases during mating season (May, June), and peaks in both sexes in August, before entering hibernation, when the body mass is at its highest.
Shivatcheva et al. (1988) monitored cortisol levels in European ground squirrels (Spermophilus citellus) throughout the year, including the period of hibernation (from October to March), as well as during periods of awakening from torpor. The authors also found that the cortisol level is highest during spring, summer, and early autumn before entering hibernation. Cortisol levels decrease in torpor; it is the smallest in the first phase of torpor (from October to January) and increases in the second phase (from January to March).
In the phases of complete euthermia during intermittent awakening from torpor, cortisol levels are high and reach the values of euthermic active animals. The plasma levels of leptin in the little brown bat (Myotis lucifugus) increase before the onset of body weight gain and in adiposity, and decrease before entering hibernation despite the maintenance of body weight and adiposity (Kronfeld-Schor et al., 2000). When it comes to thyroid hormones, the results are controversial, which is probably the result of different methodologies.
Berrada et al. (2000) showed that serum T3 and T4 levels are reduced in both naturally occurring and induced hibernation, as well as in hypothyroid jerboas (Jaculus orientalis). The authors believe that all these conditions are accompanied by a depression of thyroid gland activity and the level of thyroid hormones in the circulation as preparation for hibernation. It is generally believed that before entering hibernation and during hibernation, there is an involution of the endocrine glands, which implies dormant thyroid and adrenal glands, the atrophy of ovaries and testicles before entering hibernation, which remain in that state during torpor, and begin to activate at the end of torpor (Lyman and Chatfield, 1955).
Hibernator's Life Requires Metabolic Reprogramming
Oscillations in metabolic profiles that generally follow circannual patterns are regulated at transcriptional, post-transcriptional, translational, and post-translational levels in a tissue-specific manner (Biggar and Storey, 2014; Morin and Storey, 2009).
The degree of metabolic suppression is organ specific; it is profound in the gastrointestinal system, liver, and muscles of hibernators, and less pronounced in the brain, heart, and brown adipose tissue (BAT); the latter tissues being the most responsible for coordination, support, and thermogenesis even in the systemically depressed metabolic phenotype (Carey et al., 2003; Heller, 1979; Hittel and Storey, 2002a; Sonntag and Arendt, 2019; Stancic et al., 2018; Storey et al., 2010). However, hibernators maintain neuronal activity, as evidenced by sustained sensitivity to auditory, tactile, and thermal stimulation (Sonntag and Arendt, 2019), as well as cardiac contractility and ADs (Buzadzic et al., 1998; Buzadzic et al., 1997; Buzadzic et al., 1990; Lee et al., 2007; Stancic et al., 2018; Vucetic et al., 2013). Still, an over-reaching principle of metabolic reorganization emerges, which is synonymous with the hibernator's life cycle.
In anticipation of hibernation, physiological adjustments occur long before the onset of hibernation. For example, in late summer intensive feeding behavior occurs and is accompanied by the synthesis of triacylglycerols that are stored in large WAT depots (Carey et al., 2003), resulting in an increase in body mass by >50% (Sheriff et al., 2013). In support of this behavioral change, the expression of metabolic enzymes involved in lipid biosynthesis is strongly upregulated at this time, especially in the liver (Storey, 2003), whereas glycogen reserves and gluconeogenesis are maintained at a constant level (Hindle et al., 2011).
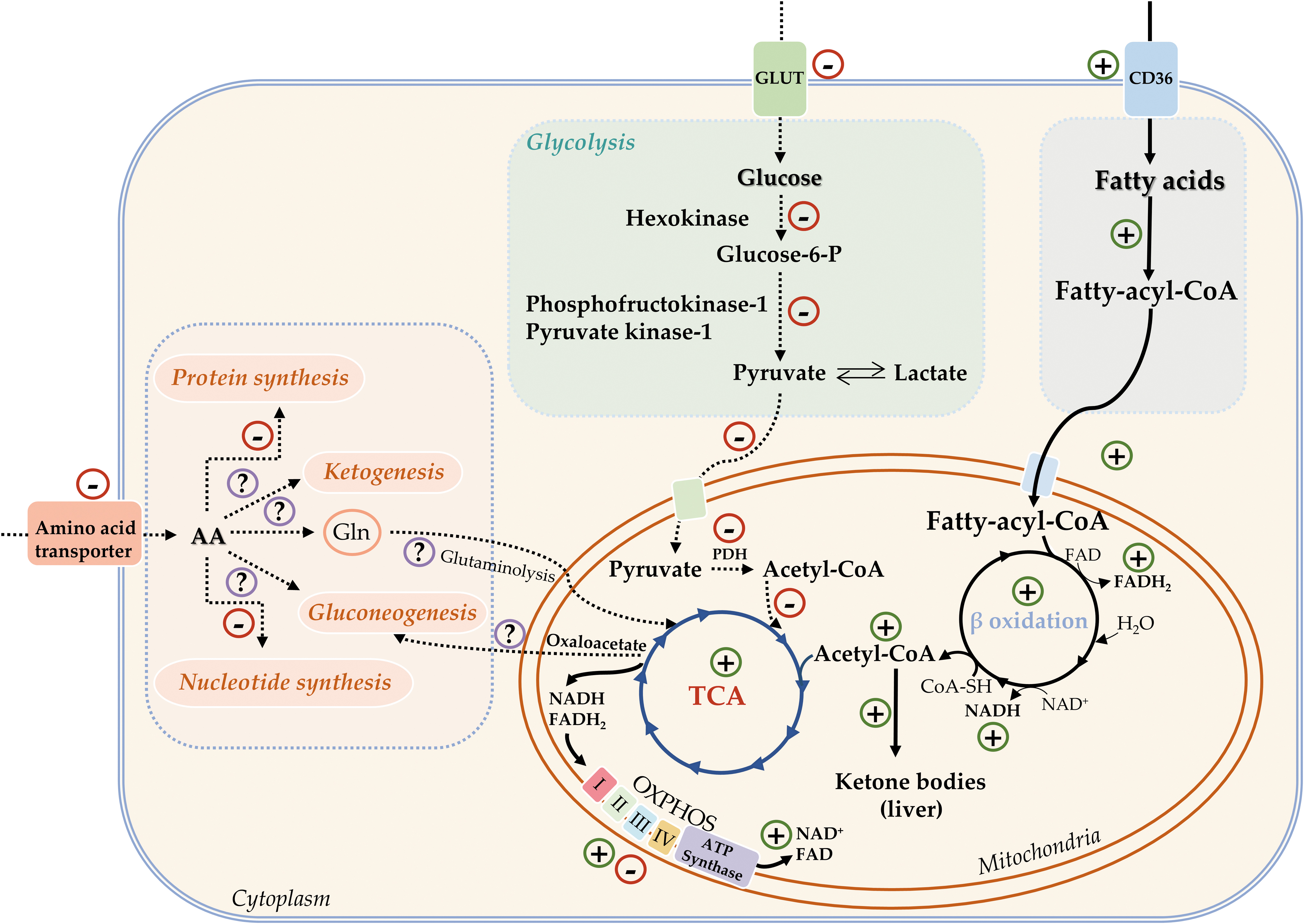
In general, hibernation is characterized by the strong suppression of biosynthetic processes, including glucose, fatty acid, protein, and nucleic acid synthesis (Cai et al., 2004; Eddy and Storey, 2003; Frerichs et al., 1998; Storey, 2004; Storey, 2003). This helps save energy for survival and favors catabolic processes. A complex metabolic switch redirects energy metabolism from carbohydrates (glucose) to lipids (fatty acids) during torpor (Eddy and Storey, 2004; Tashima et al., 1970).
Accordingly, the expression of glycolytic enzymes is downregulated by controls at key regulatory points of the pathway (hexokinase, phosphofructokinase 1, pyruvate kinase 1) (Khu et al., 2014; Smolinski et al., 2020; Stancic et al., 2018), as well as at the level of glyceraldehyde-3-phosphate dehydrogenase (Soukri et al., 1996a; Soukri et al., 1996b). Circulatory glucose concentrations are also reduced (Burlington and Klain, 1967). In addition, activation of pyruvate dehydrogenase (PDH) kinase 4 mRNA and protein expression (Andrews et al., 1998; Buck et al., 2002) inhibits PDH activity via phosphorylation (Wijenayake et al., 2018), disabling the entry of carbohydrate (as pyruvate) derived from glycolysis into the citric acid cycle.
Instead, the circulatory concentrations of fatty acids are increased, and fatty acids become the primary fuel for ATP synthesis during hibernation (Buzadzic et al., 1990; Wilson et al., 1992). Lipid catabolism is activated by the induction of key enzymes involved in fatty acid mobilisation from WAT (Bauer et al., 2001; Cochet et al., 1999; Wilson et al., 1992) and β-oxidation (Elaine Epperson et al., 2010; Hindle et al., 2011; Stancic et al., 2018), whereas fatty acid biosynthesis is halted (Hampton et al., 2011; Storey, 2003). Moreover, fatty acids are the only source of “metabolic” water during hibernation (Galster and Morrison, 1976).
At the same time, the lipolytic product glycerol serves as a substrate for gluconeogenesis and restitution of glucose and glycogen reserves during short-term arousals back to euthermia (Galster and Morrison, 1975; Lyman and Chatfield, 1955). Of these new gluconeogenic substrates, about three-fourths are derived from fat, and one-fourth is supplied from proteins (Galster and Morrison, 1975). However, whether gluconeogenesis occurs during hibernation is still in question (Green et al., 1984), with the prevailing consensus that it is, in fact, attenuated (Gehnrich and Aprille, 1988; Mohr et al., 2020).
Fatty acids are the dominant source of energy during hibernation for the heart (Burlington et al., 1972; Storey, 2003) and indirectly for the brain, when in the absence of glucose the brain uses ketone bodies synthesized from fatty acids by the liver as its only source of energy. Supported by fatty acid oxidation (Andrews et al., 2009), ketone body synthesis is enhanced in hibernation and interbout arousal as indicated by higher circulatory levels of β-hydroxybutyrate (Andrews et al., 2009; Feng et al., 2019; Regan et al., 2019). However, amino acids are probably not the main carbon source for ketogenesis, given that muscle proteins are “protected” from degradation during hibernation (Gao et al., 2012; Storey, 2004).
Preserving skeletal muscle mass and function through the winter is a priority. One aspect of this is to limit amino acid catabolism (Hindle et al., 2011), so that muscle function remains intact upon waking up, despite long periods of inactivity, which, in humans and most other mammals, would result in muscle atrophy. Hence, in general, amino acids do not contribute to the energy potential in hibernation, even though total muscle mass has been shown to decrease over the winter (Galster and Morrison, 1976; Wang, 1989). There are still conflicting opinions about the degree of muscle proteolysis, and whether and to what extent amino acids can contribute to gluconeogenesis and ketogenesis during hibernation (Storey, 2004; Storey, 2000; Tashima et al., 1970).
Mitochondrial oxidative phosphorylation supports the catabolism of fatty acids as the primary fuel source for ATP synthesis, while the oxidative phosphorylation rate is very low during torpor and very high during arousal due to shivering and nonshivering thermogenesis and remains at an intermediate-to-high level over the arousal period (Staples, 2014). However, this response is accompanied by tissue-specific induction of individual components of the respiratory chain complexes, ATP synthase, and cytochrome c oxidase expression (Eddy et al., 2006; Hittel and Storey, 2002b; Storey, 2003).
For example, mitochondrial oxidative phosphorylation in the brain is increased during torpor and interbout arousal (Ballinger et al., 2017; Epperson et al., 2010), even though mitochondrial copy number does not change during annual cycles (Ballinger et al., 2017). However, in the prehibernation period, induction of mRNA and protein expression of oxidative phosphorylation components has been observed, and this is maintained during torpor as a precondition for rapid entry into interbout arousal and re-establishment of the euthermic state (Stancic et al., 2018).
It is important to keep in mind that even though metabolism occurs at low perfusion rates and oxygen concentrations, the organs of hibernating mammals are not ischemic or hypoxic. Instead, oxidative metabolism is fine-tuned to the new metabolic landscape of hibernation (Green, 2000; Lindell et al., 2005; Storey, 2003). In euthermia, a high metabolic rate and normoxia are re-established, especially during the interbout arousal process when the metabolism and oxygen consumption are at even higher rates.
Ion Homeostasis in Hibernation: Illustration of Muscle Protection
The skeletal muscles of ground squirrels have a high tolerance to atrophy and dysfunction after long periods of inactivity despite a 14%–65% reduction in body mass (Carey et al., 2003). The maintenance of the ion gradient across cell membranes, especially the regulation of Ca2+ concentration in cells, is more effective at low temperatures in hibernators (Carey et al., 2003). This response is tissue specific, as with NaK-ATPase, which is under dominant post-translational regulation through reversible phosphorylation, and whose activity is decreased during torpor in skeletal muscle, kidney, and liver, and unchanged in the heart and intestinal epithelial cells (Carey et al., 2003).
On the contrary, Storey reported that ∼200 genes are upregulated in skeletal muscle during hibernation, six of which are membrane transporters responsible for the transport of organic cations, anionic and neutral amino acids, monocarboxylates, bicarbonate, Na+, K+, and Cl− (Storey, 2004). The authors emphasize the importance of this change in cold acclimation and in hibernation, in addition to coordinated and regulated suppression of metabolism, reversible phosphorylation, preconditioning, increased levels of AD, suppression of gene expression, proteolytic suppression, etc.
Xu et al. (2013) provide a very intriguing strategy for muscle protection in 13-lined ground squirrels (I. tridecemlineatus) through activation of the protective “endurance exercise pathway.” Namely, muscles of ground squirrels in torpor exhibit a shift to slow-twitch Type I muscle fibers, with activation of the peroxisome proliferator-activated receptor-γ coactivator-1α (PGC-1α)–mediated pathway, increase in antioxidant capacity without evidence of oxidative stress, decline in apoptotic susceptibility, and enhancement of mitochondrial metabolism.
AD in Hibernators
The specifics of AD organization during the life cycle of hibernators raise several important questions regarding redox regulation (Fig. 3). Different phases of the circannual rhythm: spring—waking up from hibernation and reproduction; summer—intense nutrition; autumn—preparation for hibernation; and winter—deep hibernation accompanied by occasional awakening are each characterized by specific redox profiles, parallel to metabolic reorganization. To enable large-scale physiological adaptations, reprogramming at the level of AD and redox regulation is essential.
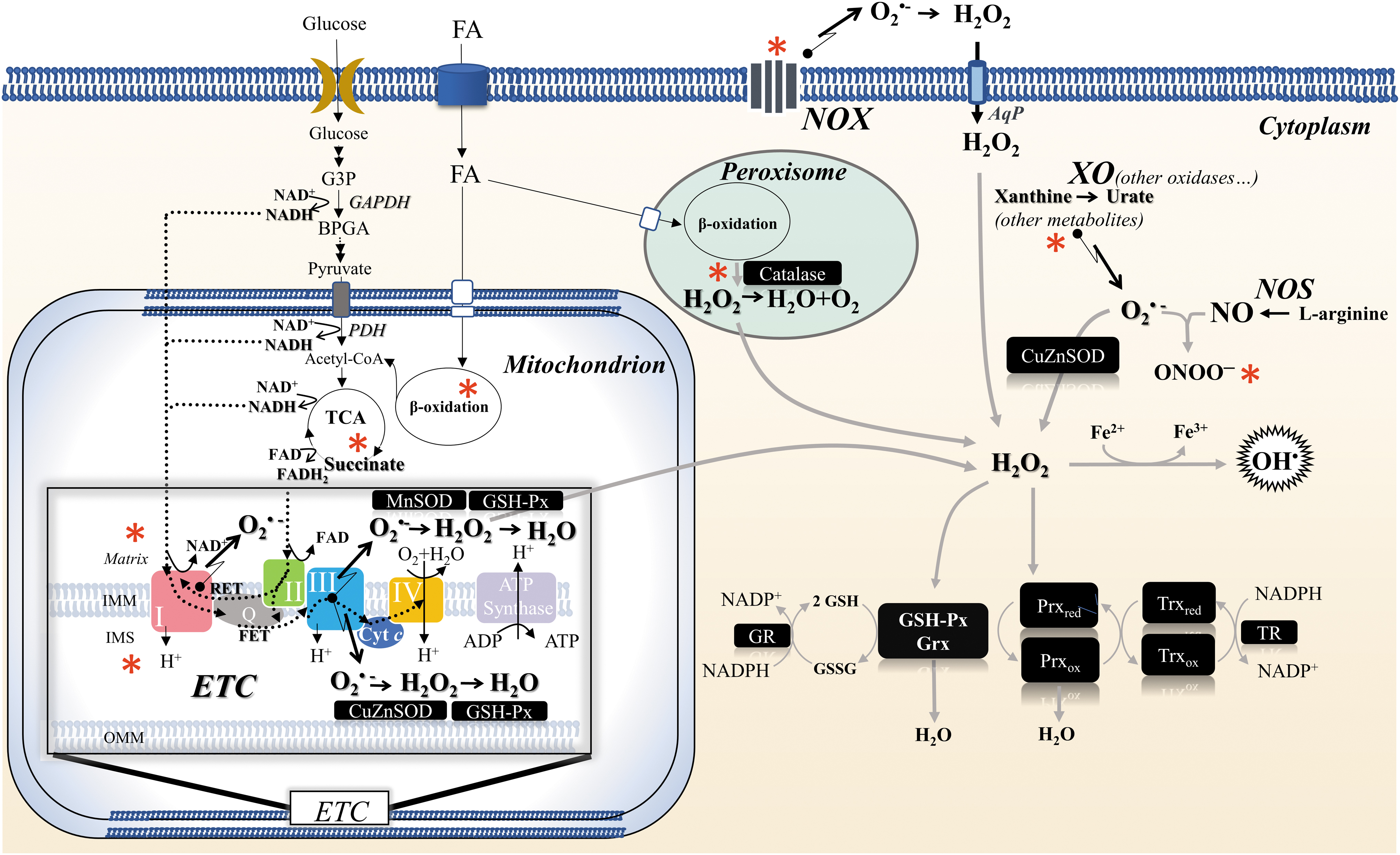
In fact, the importance of strong AD for animal survival under diverse environmental stress conditions is well established, with both constitutive and inducible responses involved. Moreover, metabolic adaptations to adjust redox biochemistry are tissue specific and sometimes need to be established well before entering hibernation.
Accordingly, a “preparation for oxidative stress” strategy has been identified during which animals show upregulation of antioxidant capacity (enzymes or small molecules) while under stress or hypometabolic conditions to maximize a rapid antioxidant response when oxygen levels are suddenly elevated again during arousal from torpor. Indeed, preparation for oxidative stress has now been described in multiple situations, including response to hypoxia/anoxia, dehydration, thermal and heavy metal stresses, as well as during hypometabolic states of torpor, dormancy, estivation, and hibernation (Giraud-Billoud et al., 2019).
In hibernators, preconditioning plays an essential role both in preparation for hibernation and during hibernation, so that animals are ready to awaken quickly and transition from hypothermia to euthermia. For example, in Arctic ground squirrels (Spermophilus parryii), plasma ascorbic acid concentrations increase three- to fivefold during torpor but are rapidly utilized during arousal to combat a surge in ROS generation associated with the oxygen-dependent thermogenesis needed to rewarm the body (Drew et al., 2002).
BAT plays a special role in hibernation due to its thermogenic role, metabolic specificities, and unique redox regulation. Buzadzic et al. (1990) examined changes in ADs in BAT in European ground squirrels (S. citellus) that are awake and active during spring and autumn, and hibernate during winter.
A marked increase in glutathione peroxidase (GSH-Px) activity and ascorbic acid levels occurred in autumn with constant levels of other components of AD, compared with spring levels, indicating preconditioning before hibernation. Hence, despite overall metabolic rate depression during hibernation, a generally high level of AD was found in BAT with increases in copper–zinc superoxide dismutase (CuZnSOD), manganese superoxide dismutase (MnSOD), GSH-Px activity, and ascorbic acid levels during hibernation as compared with spring, regardless of levels recorded in autumn.
Other components of AD were maintained at the same level. Obviously, in addition to preparing for hibernation, preconditioning for interbout arousal takes place during hibernation itself. This was indicated by the results of Petrovic et al. (1983), which showed increased total SOD activity, not just in BAT but also in the liver, kidney, heart, spleen, and lungs of European ground squirrels (S. citellus) upon arousal, as compared with hibernation levels.
It is truly fascinating that redox preconditioning takes place in preparations for deep torpor (hypometabolic state) and continues during hibernation as preparation for interbout arousal from torpor (hypermetabolic state). The systemic role of BAT in both states is vital to support and maintain the hibernator's life. Similar changes have been found in the liver of hibernating animals, where AD was either maintained at spring levels or increased, despite a considerable decrease in metabolic activity (Buzadzic et al., 1990).
When considering whether changes in AD could be related more to low temperatures than hibernation per se, Buzadzic et al. (1990) pointed out that changes in ADs in BAT and liver represent an adaptation to hibernation, and that they are not a consequence of low temperature alone. This was confirmed by results obtained from European ground squirrels (S. citellus) that remained active in winter and did not enter hibernation while kept at an ambient temperature of 7°C with a rectal temperature of 34°C. Biochemical parameters such as the circulating concentrations of glucose and free fatty acids were the same as in active euthermic animals in spring, confirming this, even if BAT tissue weight was increased four times, as in torpor.
Other results (Vucetic et al., 2013) from euthermic European ground squirrels (S. citellus) acclimated to low temperature for 21 days and ground squirrels in torpor in the prehibernation period (autumn, September) shed new light on the effect of low temperature and preconditioning itself.
In general, the protein expression of antioxidant enzymes was increased during hibernation in BAT, muscle, retroperitoneal and subcutaneous WAT with certain tissue-specific differences. For example, while CuZnSOD, MnSOD, and catalase (CAT) were increased, GSH-Px was decreased in BAT; CuZnSOD, CAT, and GSH-Px were increased, and MnSOD was decreased in the muscle; CuZnSOD was increased, whereas MnSOD and GSH-Px were decreased in retroperitoneal WAT; GSH-Px was increased in gonadal WAT; CAT was increased, whereas GSH-Px and MnSOD were decreased in subcutaneous WAT.
An increase in the protein expression of AD components in hibernation in these tissues was also found in ground squirrels kept at low temperatures. These results clearly indicate that preconditioning as a preparation for torpor occurs in prehibernation periods (autumn), and that it is further stimulated by low temperatures.
In a similar experimental setup, a detailed analysis of AD and metabolic profiles in the heart of hibernators (European ground squirrels, S. citellus) confirmed previous claims, and firmly linked redox and metabolic integration (Stancic et al., 2018). In parallel with the increase in the protein expression of CuZnSOD, MnSOD, GSH-Px, and thioredoxin (even with no change in enzyme activity) during torpor (hypometabolic state), the protein expression of respiratory complexes I, II, III, and IV, as well as ATP synthase and cytochrome c, was markedly increased. It is important to note that the absence of changes in the expression of the hypoxia-inducible transcription factor-1α confirms that the heart is not hypoxic.
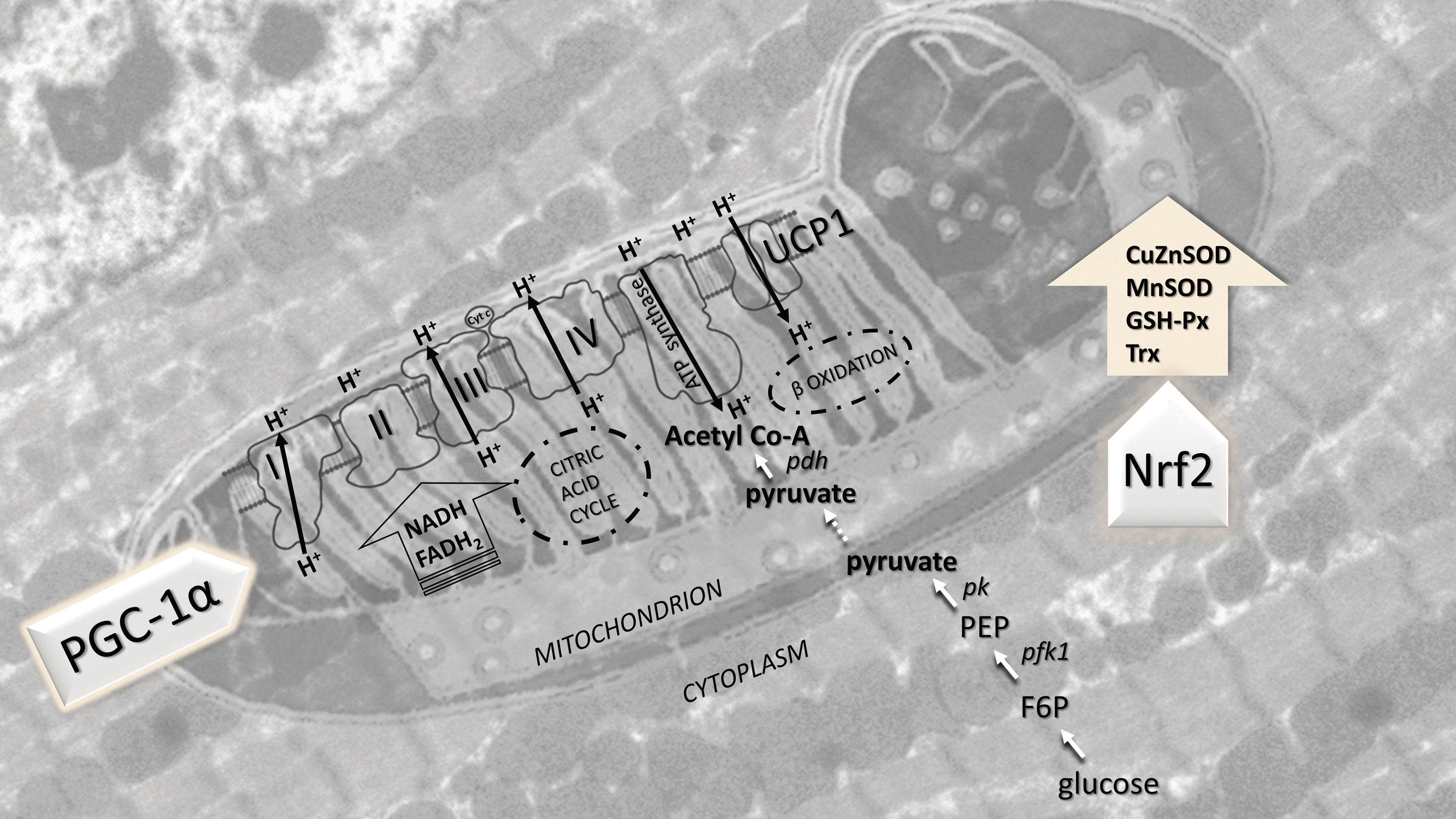
A newly established redox-metabolic homeostasis is essential for hibernation, especially in the preparation and preconditioning for awakening from hibernation. This synchronized redox-metabolic response is regulated by transcription factors. For example, PGC-1α protein expression was increased twofold (Stancic et al., 2018), and Nrf2 mRNA and protein levels were increased 1.5-fold in the heart of 13-lined ground squirrel (I. tridecemlineatus) in hibernation (Pier et al., 2008).
Moreover, Nrf2 protein levels were elevated during entry into torpor, deep torpor, and early stages of arousal but returned to euthermic values in fully aroused ground squirrels during interbout (Pier et al., 2008). Induction of mitochondrial metabolism and oxidative phosphorylation in the heart during hibernation was accompanied by inhibition of glycolysis (protein expression of phosphofructokinase 1 was reduced by 90%), inhibition of the glycolysis product, pyruvate, entry into mitochondria (PDH protein levels were reduced by 50%), and an increase (twofold) in acyl-CoA dehydrogenase protein expression (Stancic et al., 2018).
This confirms that in hibernation, the heart intensively uses fatty acids stored in the prehibernation period to provide ATP, primarily by oxidative phosphorylation. This fascinating redox-metabolic synchronization in the heart of hibernators is schematically illustrated in Figure 4.
Transcription Factor Control of AD During Hibernation
Three prominent transcription factors have been identified as having significant roles in cellular AD during hibernation. These are Nrf2, NF-κB (nuclear factor kappa-light-chain-enhancer of activated B cells), and FoxO3a (forkhead box class O member 3) (Ma, 2013; Morgan and Liu, 2011; Zhang et al., 2019). All can be classed as redox-sensitive transcription factors and have been linked to the upregulation of genes/proteins involved in AD (and other actions) during hibernation. All three have also been shown to be upregulated in tissues of hibernating ground squirrels, as compared with euthermic controls.
Nuclear factor erythroid 2-related factor 2
When inactive, Nrf2, a basic leucine zipper transcription factor, is bound to a repressor protein Kelch-like ECH-associated protein 1 (Keap1) in the cytoplasm. However, under electrophilic or oxidative stress conditions, Nrf2 is released from Keap1 (i.e., H2O2-mediated Nrf2 phosphorylation or Keap1 oxidation/phosphorylation) and translocates into the nucleus, where it dimerizes with small Maf proteins (or other partners) and binds to the antioxidant response element in the promoter region of selected genes (Ma, 2013).
Prominent antioxidant genes regulated by Nrf2 include heme oxygenase-1 (HO-1, the inducible isoform), glutathione S-transferases, CuZnSOD, thioredoxin reductase 1. Other genes regulated by Nrf2 include those associated with thioredoxin and peroxiredoxin metabolism, including multiple NADP-dependent dehydrogenases that supply the NADPH reducing power to the thioredoxin, peroxiredoxin, and glutathione systems (Fig. 5).

Studies of Nrf2 involvement in hibernation of 13-lined ground squirrels, Ictidomys (formerly Spermophilus) tridecemlineatus, showed that Nrf2 transcript levels were significantly elevated in multiple tissues during hibernation at a body temperature (Tb) of ∼5°C, as compared with euthermic controls (held at 21°C, Tb 36°C–38°C) (Ni and Storey, 2010). Nrf2 protein levels (both 57 and 100 kDa subunits) also increased in several tissues during hibernation (2–5 days at Tb ∼5°C); for example, by 1.6, 2.2, and 3.2-fold for liver, heart, and BAT, respectively. These changes in Nrf2 were matched by 1.5- to 1.8-fold increases in the Nrf2 partner protein, MafG.
Further study assessed the cytoplasmic versus nuclear distributions of Nrf2 57 kDa and MafG, and found significant increases in both proteins in the nuclear fraction of liver (∼1.4-fold) during hibernation with opposite reductions in the cytoplasmic content. This supports an active role for Nrf2-mediated gene transcription during hibernation against a background of strong global suppression of gene expression.
As a result, various tissues showed significantly increased protein levels of Nrf2 target genes (especially during entry into torpor), including HO-1, CuZnSOD, and aflatoxin aldehyde reductase (AFAR1) (Ni and Storey, 2010; Pier et al., 2008). Overall, these data support a strong role for Nrf2 action in readjusting antioxidant potential during hibernation.
A subsequent study revealed more about Nrf2 regulation and action during hibernation with an analysis of the Nrf2 system in I. tridecemlineatus skeletal muscle, a tissue that contributes shivering thermogenesis to the arousal process (Tessier et al., 2021). Responses by Nrf2 (both 62 and 100 kDa forms), Keap1 (the negative repressor of Nrf2), and small Maf proteins (MafF, MafG, MafK) that form heterodimers with Nrf2 to activate transcription of downstream genes were assessed over a 5-point time course of torpor-arousal.
Prominent responses included (a) a strong suppression of MafF and MafG proteins during entrance into torpor; (b) a pronounced increase in MafK protein in prolonged torpor compared with euthermic controls; and (c) strong increases in Nrf2 (100 kDa) and the downstream antioxidant enzyme, CAT, when ground squirrels aroused from torpor, contributing to antioxidant needs in the shivering muscle. Post-translational modifications of Nrf2 were also identified and showed very strong increases in serine phosphorylation of Nrf2 p62 during prolonged torpor, which were sustained in early arousal but declined during interbout.
Acetylation of Nrf2 was also high during early arousal and interbout time periods. These parallel increases in both serine phosphorylation and lysine acetylation that span late torpor and arousal suggest that Nrf2-mediated transcriptional controls over gene expression are very important to the arousal process before declining again after interbout euthermia is re-established. This is not unexpected given the high rates of oxidative stress associated with rewarming the body during arousal, as illustrated most clearly by the rapid depletion of ascorbic acid as ground squirrels rewarm (Drew et al., 2002).
Previous studies have suggested that serine phosphorylation promotes Nrf2 nuclear localization, whereas acetylation promotes Nrf2-DNA binding. Hence, coordinated changes in these two post-translational mechanisms are not unexpected and contribute to a smooth activation of Nrf2-mediated gene transcription as ground squirrels exit torpor.
Nuclear factor kappa-light-chain-enhancer of activated B cells
The redox-sensitive transcription factor NF-κB is also involved in AD during hibernation in I. tridecemlineatus. Under normal conditions, NF-κB is bound to its inhibitor protein, IκBα, in the cytoplasm, but under stress two upstream regulators of NF-κB, tumor necrosis factor receptor 1 (TNFR1) and Fas-associated protein with death domain (FADD), become active, and then trigger the IκB kinase (IKK) that phosphorylates IκBα to halt it from binding with NF-κB (Napetschnig and Wu, 2013). This leads to NF-κB phosphorylation and translocation to the nucleus, where it binds to the promoter region of selected genes (Fig. 6).

An initial study showed that protein levels of both p50 and p65 subunits of NF-κB were elevated in skeletal muscle during torpor and arousal phases (but not during entry into torpor), as compared with euthermic controls (Allan and Storey, 2012). Furthermore, both phospho-IκBα and IKK content increased strongly during arousal, suggesting that NF-κB is the most active during this transition to mediate the expression of genes needed for a return to euthermic function. Indeed, protein levels of an NF-κB target, the MnSOD, were consistently elevated in skeletal muscle (by ∼2-fold) during both hibernation and arousal phases, and HO-1 also increased during the early hours of arousal (Allan and Storey, 2012).
These data provide additional evidence of a need for significant ADs, particularly as animals arouse from torpor when oxygen uptake and oxygen-based thermogenesis by BAT and skeletal muscle soar to very high levels. It is well known that shivering thermogenesis by skeletal muscle is an important contributor to heat generation during arousal from torpor (in addition to brown fat), and that superoxide can be produced from 11 different sites within mitochondria (Brand, 2016). Although shivering does not begin until the squirrel body is partially rewarmed, the high rates of ATP turnover needed to support shivering would be facilitated by the action of intramitochondrial MnSOD in rapidly dissipating superoxide.
A study that surveyed eight ground squirrel tissues also showed that NF-κB–mediated processes were activated during torpor (5 days of continuous torpor with Tb of 5°C–8°C) in selected tissues, particularly in brain (Hadj-Moussa et al., 2020). For example, in the cerebral cortex of hibernating animals, both TNFR1 protein and phospho-FADD levels were elevated by 2- to 4-fold, phosphorylated active NF-κB (S536) increased twofold, and NF-κB binding to DNA increased significantly. This led to a marked increase in the levels of a well-known downstream protein target (c-Myc).
Brainstem showed a similar pattern with particularly strong increases in phospho-FADD and phospho-NF-κB (S536), whereas other tissues showed more muted responses. Transcription factor DNA-binding ELISAs were also used to measure the relative degree of NF-κB binding to DNA during hibernation and showed a significant increase in NF-κB binding to DNA in cerebral cortex during prolonged torpor (Hadj-Moussa et al., 2020). These data suggest that ADs are particularly important for neuroprotection of brain during hibernation; brain being crucial to regulating whole-body oscillations of torpor/arousal. Indeed, brain is very sensitive to knockout of NF-κB subunits (p50 or p65) or enhanced inhibition by IκB; such manipulations lead to major defects in learning and memory in mice (Kaltschmidt and Kaltschmidt, 2009).
Forkhead box class O member 3
Forkhead box transcription factors of the O subclass (FoxO) also contribute to AD in hibernation. This subclass is involved in regulating diverse processes, including apoptosis, cell cycle arrest, and AD, all of these being important considerations for hibernation. FoxO3a is known as a tumor suppressor and promoter of longevity. It is regulated by the action of a variety of protein kinases, in both cytoplasmic and nuclear compartments, with either activating or inhibitory effects (Wang et al., 2017). For example, c-Jun N-terminal kinase (Jnk)-mediated FoxO phosphorylation is conserved across the animal kingdom, and is known to be triggered by oxidative stress or UV radiation (Iordanov and Magun, 1999) (Fig. 7).

Analysis of ground squirrel skeletal muscle showed that FoxO3a protein levels increased by 3.2- to 4.5-fold when animals entered torpor, as compared with euthermic controls (Wu and Storey, 2014). Phosphorylation of FoxO3a at Ser-253 paralleled the rise in FoxO3a protein, and p-Thr-32 levels also rose when ground squirrels entered torpor. Both FoxO3a and its coactivator protein β-catenin showed higher nuclear localization during torpor, and DNA binding by FoxO3a increased ∼2-fold.
As a result, mRNA levels of three downstream targets of FoxO3a were elevated when ground squirrels entered torpor: catalase, cyclin G2, and p27. Their protein products can each contribute important cell preservation functions to the hibernator. CAT is a major antioxidant enzyme that rapidly breaks down hydrogen peroxide to water and oxygen, significantly minimizing H2O2 levels during torpor and/or arousal.
By contrast, cyclin G2 and P27 are involved in inhibitory regulation of the cell cycle that is profoundly suppressed during torpor (Wu and Storey, 2012). Cyclin G2 is upregulated by growth-inhibitory stimuli and thought to promote quiescence of the cell cycle, whereas P27 belongs to the well-known Cip/Kip family of cell cycle inhibitors.
Gasotransmitters in Hibernation: The Role of H2S, NO, and CO
The role of ROS in the regulation of various physiological and pathophysiological conditions is well studied in nonhibernators. However, there are little data on ROS signaling pathways in hibernation, especially during specific annual cycles such as prehibernation, torpor, interbout arousal, and awakening from hibernation. It was suggested that ROS play an important role in awakening from hibernation (Wei et al., 2018). H2O2 as a signaling molecule was shown to play an important role in the Nrf2-dependent induction of endogenous antioxidant protection during interbout arousal, late torpor, and awakening from hibernation (Wei et al., 2018).
In the recent decades, intensive research has been carried out on the role of gasotransmitters, hydrogen sulfide (H2S), nitric oxide (NO), and carbon monoxide (CO) in metabolism regulation, especially in terms of mitochondrial oxidative phosphorylation in numerous nonhibernating species. However, despite such progress, there are still relatively little data on the role of gasotransmitters in hibernation.
Blackstone et al. (2005) showed that H2S reduces metabolic rate and body temperature in mammals through specific reversible inhibition of complex IV of the electron transport chain. The authors concluded that in doses up to 80 ppm, H2S induces a suspended animation-like state in mice, similar to the induction of hibernation, torpor, or aestivation in hibernating animals. However, Baumgart et al. (2010) and Hemelrijk et al. (2018) showed that the hypothermia-inducing effects of H2S are lost at an oxygen concentration of 21%.
Recent data also showed that H2S plays an important role in metabolic suppression during hibernation. Jensen et al. (2021) showed that the effect of H2S in the liver of hibernating 13-lined ground squirrels (I. tridecemlineatus) depends on its concentration in torpor and upon awakening, which is primarily determined by the activity of two key enzymes involved in its metabolism, cystathionine β-synthase (CBS), responsible for H2S synthesis, and membrane-bound sulfide:quinone oxidoreductase (SQR), responsible for H2S degradation (oxidation) in the mitochondria.
In vitro, on isolated liver mitochondria from ground squirrels, the authors postulated that when SQR is inhibited, H2S accumulation induces a hypometabolic state during torpor via inhibition of oxidative phosphorylation. In addition, in vivo blood plasma and liver levels of H2S were higher in torpor than upon awakening from torpor and during summer in normoxic and normal metabolic state. Increased CBS activity in torpor, as well as increased availability of substrates cysteine and cystathionine in the plasma of 13-lined ground squirrels (I. tridecemlineatus) (D'Alessandro et al., 2017) and the lungs of Syrian golden hamsters (Mesocricetus auratus) (Talaei et al., 2012), also support this claim.
Although there is no definitive evidence for the involvement of H2S in the induction of hibernation, H2S plays an important role in regulating the hibernator's metabolism both in hypothermia (torpor) and in euthermia (awakening from torpor and active state in spring and summer). In addition to its role in regulating mitochondrial energy metabolism, H2S can achieve other beneficial effects in hibernators. Among other things, Talaei et al. (2012) showed that in the lungs of Syrian golden hamsters (M. auratus), H2S produced in torpor (as indicated by an increased expression and level of CBS) contributes to tissue remodeling by suppressing the inflammatory response and inhibiting gelatinase activity.
To what extent do H2S plasma levels correspond to its levels in the tissues, cells, and cell compartments (mitochondria), is there a tissue-specific expression and activity of enzymes involved in its synthesis and degradation, and what is their post-translational regulation (e.g., acetylation), concentration-dependent response, or connection with redox-sensitive transcription factors (primarily Nrf2), are only some of the open questions about the role of H2S in redox-metabolic regulation in different life phases of hibernators.
For more detailed mechanisms and other possible pathways of H2S signaling in hibernation, including protective ones, see excellent review papers (Giroud et al., 2021; Jensen and Fago, 2021). It should be taken into account that the induction of a suspended animation-like state with H2S is less efficient in rats, unlike in mice, and even inefficient in other species such as piglets, pigs, sheep, and humans (Cerri, 2017).
The vascularization is vital for maintaining overall tissue and organ homeostasis in hibernation concerning their metabolic activity. In hibernation, peripheral resistance enables the redirection of blood to vital organs such as the heart, brain, lungs, and BAT. In general, NO has an important role in regulating vascular tone. Saitongdee et al. (1999) showed that in hibernating hamsters, the percentage of endothelial nitric oxide synthase (eNOS)- and endothelin (vasoconstrictor)-positive endothelial cells was decreased in renal and mesenteric arteries in hibernation, and increased in the early phase of awakening from hibernation and in euthermic animals.
The authors conclude that the endothelium has an active role in regulating vascularization relative to the reduced metabolic activity in these tissues (10% of the euthermic level in kidneys and almost entirely suspended in the intestines) and reduced blood flow (Zatzman, 1984).
Moreover, in the European ground squirrel (S. citellus), Sandovici et al. (2004) showed lower immunohistochemical reaction for eNOS (but not neuronal and inducible NOS), as well as lower eNOS mRNA level and activity in glomeruli during torpor and awakening from torpor compared with nonhibernating euthermic animals. As the authors did not find changes in eNOS protein levels in the interstitium, they concluded that NO has a specific role in regulating ultrafiltration in hibernation without endangering the tubular structure.
Moreover, Kudej and Vatner (2003) showed that NO is involved in maintaining myocardial blood flow in the woodchuck (Marmota monax) during hibernation almost at the control level, even if the body temperature is lowered to ∼13°C and the heart rate is lowered from 86 to ∼27 beats per minute. In parallel, the authors showed that splanchnic blood flow is reduced.
On the contrary, Popovic (1964) showed that the cardiac output of hibernating ground squirrel (Citellus tridecemlineatus) in torpor (body temperature of 7°C) is ∼65 times lower than that in the active state, just as much as the oxygen consumption of the hibernating animals was reduced. However, the arteriovenous difference in oxygen blood content was unchanged.
Another gasotransmitter, CO, is also of interest in hibernation, not only in the regulation of vascularization but also in the metabolic processes. Ni and Storey (2010) examined changes in the expression of HO-1, the inducible isoform in CO synthesis, and showed that it is strongly increased in the liver, kidney, heart, and brain during torpor in 13-lined ground squirrels (Spermophilus tridecemlineatus) when compared with euthermic controls (mRNA level 1.4–3.8 times and protein level 1.5–2.0 times).
Since HO-1 is one of the primary targets of Nrf2 and since Nrf2 and its heterodimeric partner protein, MafG, strongly increase in torpor (including nuclear translocation in the liver), the authors conclude that the induction of HO-1 in torpor is an integral part of the overall increase in AD in hibernation as a result of the production of the antioxidant biliverdin (and consequently bilirubin), also a product of the reaction catalyzed by HO-1.
Accordingly, the results of Mountassif et al. (2007) and Baker and van Breukelen (2009) showed that plasma bilirubin levels in torpor of jerboas (J. orientalis) and bile levels in hibernating ground squirrels (Spermophilus lateralis) increased five times. Moreover, biliverdin levels in the liver of 13-lined ground squirrels (S. tridecemlineatus) are 15–20 times higher during awakening and interbout periods compared with animals before entering hibernation or full torpor (Nelson et al., 2009).
Moreover, it should be acknowledged that H2S, NO, and CO regulate a wide variety of physiological processes, including oxidative phosphorylation and energy metabolism. The final effects of gasotransmitters depend on their concentration and their synergistic/antagonistic effects, which are also dependent on and specific to the oxygen concentration in mitochondria (hypoxic or normoxic conditions) (Pun et al., 2010). The molecular mechanisms of their complex interactive responses have not yet been elucidated even in nonhibernating species, but they certainly play an important role in both nonhibernators and hibernators.
Between Metabolic Reprogramming and AD: Oxinflammation in Hibernation
The pathophysiology of ischemia/reperfusion injury involves several important mechanisms, including induction of inflammatory responses, ROS signaling, and altered ionic homeostasis (Kalogeris et al., 2012; Wu et al., 2018). ROS and oxidatively damaged biomolecules are involved in the etiology of several pathological conditions, and are intimately tied to inflammatory pathways, inducing oxinflammation (Valacchi et al., 2018; Valacchi et al., 2017). Hibernating mammals represent an ideal, naturally occurring model for the study of mechanisms underlying the induction and protective strategies against oxinflammation.
In a well-controlled study, Bogren et al. (2014) showed that arctic ground squirrels (Urocitellus parryii) are highly resistant to multiorgan damage and systemic inflammation after ischemia/reperfusion events. Moreover, this protective response, not observed in nonhibernators (rats), seems to be evolutionarily conserved in hibernators since it is independent of the season and decrease in body temperature, and it is most likely regulated by the modulation of metabolic responses during ischemia/reperfusion.
In general, Logan and Storey (2018) pointed out that inflammation is suppressed in hibernation. The authors investigated advanced-glycation end-product (AGE) and AGE receptor (AGE-RAGE) pathways in BAT and WAT of hibernators (I. tridecemlineatus), and showed that these tissues are not in a state of inflammation, despite the upregulation of proinflammatory responses, especially during arousal from torpor. A recent study showed that despite a considerable increase in body weight during late summer, BAT and WAT of I. tridecemlineatus did not show increased expression of pro- or anti-inflammatory cytokines, while BAT-specific increase of interleukin-1α and soluble epidermal growth factor receptor expression may serve specific purposes other than inflammation and tissue remodeling such as suppression of adipogenesis, cell differentiation, lipid oxidation, or other energy-expensive processes (Logan and Storey, 2021b).
In line with this, activation of innate immunity, Nucleotide-binding oligomerization domain, Leucine-rich Repeat, and Pyrin domain, and Absent in melanoma 2 inflammasome signaling was shown in I. tridecemlineatus BAT during arousal, and was associated with the protection of metabolically active BAT and the support of nonshivering thermogenesis during arousal (Logan and Storey, 2021a). This response was also shown to be tissue specific since no inflammasome activation was found in WAT.
In addition, the systemic anti-inflammatory response of hibernators includes upregulation of serpins (especially A1, α1-antitrypsin and A3, α1-antichymotrypsin), which protects tissues from injury by suppressing circulating proteases involved in inflammation over weeks of continuous torpor (Storey, 2010; Storey, 2004). The explanation for such responses could lie in the balance between the induction of ADs and proinflammatory pathways, as well as the modulation of downstream inflammatory pathways.
Similarly, Carey et al. (2000) stated that even though the GSH/GSSG ratio is lower and intraepithelial lymphocyte content is increased in the intestine of I. tridecemlineatus during torpor, this represents a new equilibrium that provides better protection for enterocytes under stress.
In addition to the fact that no infiltration of polymorphonuclear leukocytes was found, the authors state that high NF-κB activation is directed toward protecting intestinal epithelial cells that favor a pro-oxidized state during hibernation and not toward inducing inflammatory responses. On the contrary, in Pelodiscus sinensis, the total GSH content in the brain decreased during hibernation and was restituted after awakening, whereas in the liver GSH content increased after arousal from torpor (Zhang et al., 2017). Moreover, GSH/GSSG ratio remains stable in the brain, while it shifts to a reduced state in the liver. These results also corroborate the tissue-specific overall adaptive homeostatic response.
It is generally accepted that inflammation is absent in the brain during hibernation. Although the brain, like other organs in hibernation, is not hypoxic, it goes through distinct phases of ischemia/reperfusion (reoxygenation) during different phases of the hibernation/arousal cycle, especially during interbout arousal. Even in euthermia, arctic ground squirrels (S. parryii) are remarkably tolerant of global cerebral ischemia (Dave et al., 2006). Dave et al. (2009) emphasized the important role of protein kinase C ɛ in the maintenance of ion homeostasis in the brain, which is essential for protection against ischemia in arctic ground squirrels.
Owing to specific mechanisms that are not yet fully understood, the brain is preconditioned to tolerate ischemia/reperfusion events. Ischemic preconditioning triggers a number of molecular mechanisms aimed at protecting internal organs. For example, remote ischemia preconditioning specifically induces eNOS in the human mammary artery (Jankovic et al., 2021). The maintenance of the ischemic brain tolerance in euthermia suggests that this response is conserved in hibernators. In the case of the hibernator brain, a number of cellular and molecular mechanisms are involved in neuroprotection; for details, see an excellent review paper (Dave et al., 2012).
However, as previously emphasized, preconditioning at multiple regulatory levels (e.g., AD, metabolism, inflammatory responses, etc.) probably represents different parts of the same, evolutionarily refined response that protects hibernators in a tissue-specific manner over different life cycle phases: euthermia, torpor, and interbout arousal. Further understanding of this phenomenon and the mechanisms underlying tissue resistance to inflammation in hibernation is of immense importance because of its potential to illuminate mechanisms underlying numerous medical conditions in which ischemia/reperfusion accompanies tissue and organ injury.
Between AD and Thermogenesis: BAT as a Hibernating Gland
BAT is a thermogenic organ that plays an important role in regulating heat production and whole-body metabolism. BAT proliferation peaks before hibernation and provides animals with heat to warm their bodies during each interbout arousal from torpor (Ballinger et al., 2016; Rouble et al., 2014). Mitochondria are the focal point not only for ATP synthesis and thermogenesis but also for glucose, fatty acid, and amino acid metabolism (Fisher, 2001).
In addition, mitochondria are essential in triggering cell death—apoptosis (Ravagnan et al., 2002), and they are also the primary sites of ROS production (Handy and Loscalzo, 2012). As highly dynamic organelles, mitochondria change their number, structure, and morphology depending on the metabolic demands of the cell (Cogliati et al., 2016; Hendriks et al., 2017). In response to numerous internal and external stimuli, all aspects of mitochondrial dynamics are under the tight control of several transcription factors (Scarpulla, 2012; Scarpulla, 2008), which includes the coordination of the nuclear and mitochondrial genome (Jornayvaz and Shulman, 2010).
Reprogramming of the mitochondrial function is most pronounced in highly metabolically active tissues such as BAT and in response to extreme environmental conditions, such as hibernation (Hadj-Moussa et al., 2018; Stancic et al., 2018; Staples and Brown, 2008). BAT mitochondria are equipped with a specific molecular adaptation—the presence of uncoupling protein 1 (UCP1), a unique protein expressed in the inner mitochondrial membrane. UCP1 activity increases the conductance for protons and provides them with an alternative pathway from the mitochondrial intermembrane space to the mitochondrial matrix.
In this way, uncoupling of the cellular respiration from oxidative phosphorylation occurs, thereby releasing the energy of the proton-motive force as heat. The physiological role of UCP1 in nonshivering thermogenesis is the production of heat without shivering, and is firmly linked to the animal's overall metabolic state and metabolic needs, as is well described in the literature (Jankovic et al., 2017; Jankovic et al., 2015; Otasevic et al., 2011; Vucetic et al., 2011). However, uncoupling cellular respiration from oxidative phosphorylation also has other important physiological roles. For example, uncoupling is involved in the regulation of electrochemical potential and consequently ATP synthesis (Ricquier and Bouillaud, 2000; Skulachev, 1998).
In addition, Skulachev (1998) pointed out that high electrochemical potential can threaten cell physiology through increased O2 •− production in mitochondria. In this regard, uncoupling mediated by UCP1 can play an antioxidant role, perhaps as the first line of defense in the regulation of O2 •− production, before O2 •− dismutation by SOD. The same author emphasized the role of UCP1 not only in BAT but also in other tissues, including the role of other UCP isoforms (e.g., UCP2 and UCP3 in animals, UCP5 in plants). Moreover, other studies pointed out that the UCP function is redox regulated, where UCP1, 2, and 3 can be activated by O2 •− and other ROS by-products (Echtay et al., 2002; Mailloux and Harper, 2011).
Skulachev (1998) further introduced the term “mild uncoupling” to describe a potentially universal tissue mechanism of UCP-mediated AD in preventing a “parasitic” one-electron reduction of O2 to O2 •−. Oelkrug et al. (2014) postulated that this role of UCP1 in buffering mitochondrial ROS production is evolutionarily conserved in mammals, emphasizing that “the association between UCP1 and ROS prevention seems stronger than the association between UCP1 and thermogenesis under mild cold conditions.” Moreover, UCP1 expression has been shown to play a role in controlling mitochondrial ROS production in skeletal muscle (Adjeitey et al., 2013).
Not only thermogenic but the antioxidant role of UCP1 is also equally vital in hibernator tissues, both for maintaining the temperature of individual tissues during torpor and nonshivering thermogenesis in BAT during arousal as well as the control of ROS production during the hyperoxia that accompanies re-establishment of euthermia (Fig. 8). Ballinger et al. (2016) showed that mitochondrial respiratory capacity, mitochondrial DNA (mtDNA) copy number, and UCP1 mRNA were significantly increased in BAT during hibernation and interbout arousals relative to the spring season, whereas UCP1 protein levels were not significantly increased in the 13-lined ground squirrel (I. tridecemlineatus).
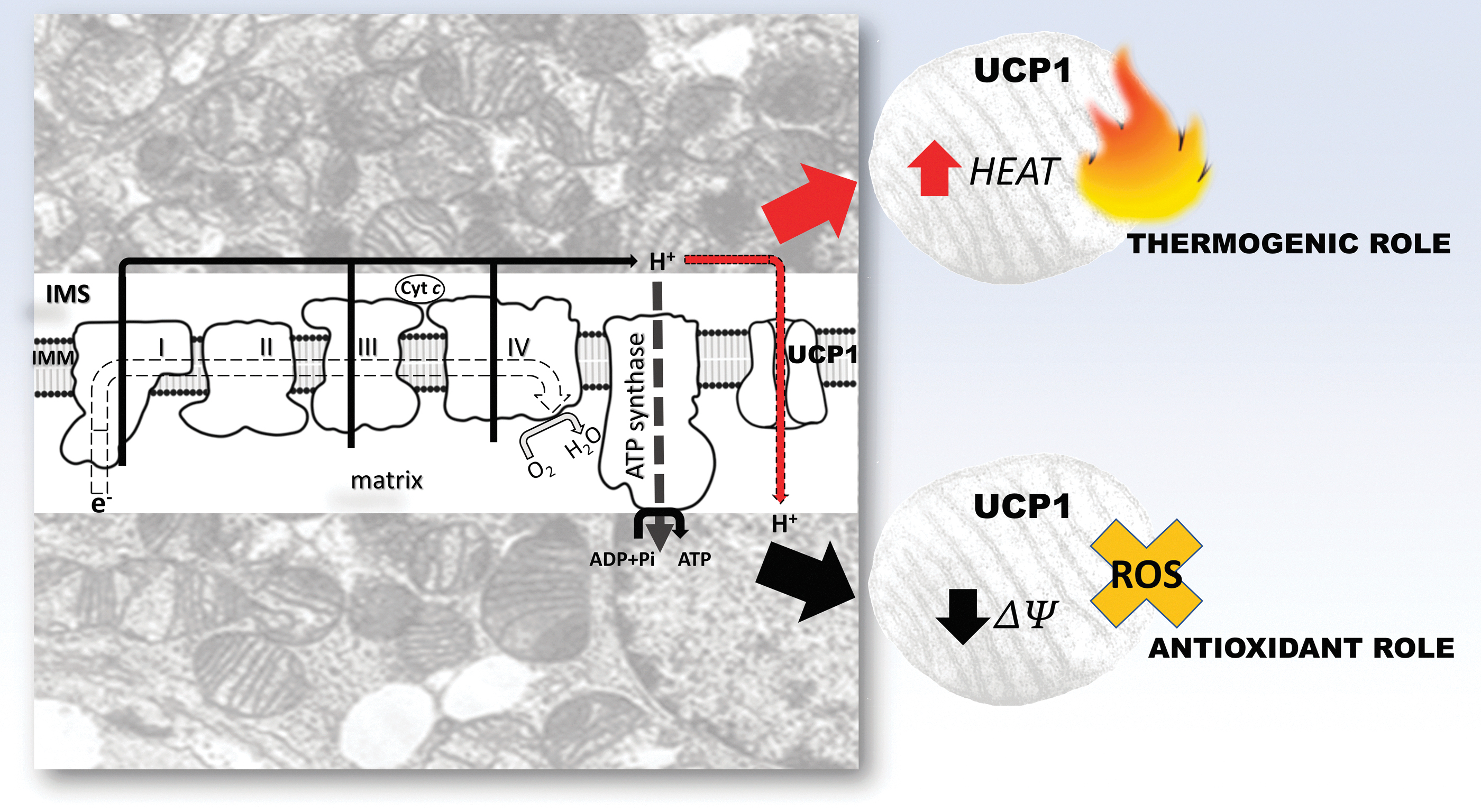
The authors concluded that with an increased number of mitochondria, the oxidative capacity and thermogenic function of BAT were increased in hibernation. This is of great importance, especially during periods of arousal from hibernation when it is necessary to rapidly increase the body temperature to the euthermic level of 37°C. Brustovetsky et al. (1990) showed that uncoupling and increased respiratory chain activity in the liver mitochondria of ground squirrels (Citellus undulatus) simultaneously take place during arousal.
That UCP1 potentially has both thermogenic and antioxidant roles in the brain of 13-lined ground squirrel (I. tridecemlineatus) was shown by Laursen et al. (2015). The authors found that neurons from the central and peripheral nervous system show UCP1 protein expression, that the expression patterns are seasonally dependent, and that they are the highest during hibernation. Functionally active UCP1 in the brain and the fact that the brain is warmer than BAT and WAT in ground squirrels highlight the importance of local thermogenesis for maintaining minimum brain function in deep torpor and reducing ROS production during arousal.
In addition, the regulation of mitochondrial function in hibernators may be dependent on other variables, such as temperature and membrane lipid composition. The temperature changes affect the efficiency of mitochondrial oxidative phosphorylation both in artificially induced conditions (therapeutic hypothermia) and naturally occurring conditions, such as torpor in hibernators.
Compared with 37°C, the efficiency of the electron transport system respiratory flux at lower temperatures (28°C) is ∼40%–75% lower in the mouse brain (Pamenter et al., 2018). However, the authors showed that mitochondrial respiration is much more efficient at low temperatures since the mitochondrial H+ gradient was more tightly coupled. In addition to temperature, mitochondrial function in hibernators is affected by membrane lipid composition, that is, mitochondrial membrane phospholipid composition, as well as by the shift in energy substrate utilization in different phases of the annual cycle.
Heim et al. (2017) found small changes in membrane lipid composition in different tissues of golden-mantled ground squirrels (Callospermophilus lateralis), especially in BAT. However, the authors concluded that these changes are inconsistent with functional adaptation, and rather represent seasonal variations in relation to uptake and utilization of fatty acids. In addition, the authors found that in accordance with the shift toward lipid metabolism during hibernation, fatty acids have the greatest influence on mitochondrial respiration, especially in prehibernation and torpor phases.
Conclusions and Biomedical Perspectives
“She may be sleeping with eyes beyond all evil realm; Beyond all things, illusions, beyond reality; And with her there sleeps, unseen, her beauty; She may be living and will come after this dream; She may be sleeping with eyes beyond all evil realm”—Serbian poet Vladislav Petkovic Dis (She may be sleeping).
Perhaps these verses most realistically describe the fascinating creation nature has designed over millions of years—hibernation. Indeed, when we observe a hibernator in torpor, their dream hides incredible secrets, the secrets of living with reduced oxygen and the secrets of the molecular mechanisms underlying the regulation of redox-metabolic adaptations.
Indeed, studies of mammalian torpor/hibernation are discovering the “outer limits” of biochemical adaptations among living creatures on Earth. This is not only fascinating in itself but it holds the tantalizing goal of ushering in the necessary advancements in life science, biomedicine, and the future of suspended animation.
Understanding the molecular mechanisms of redox and metabolic regulation involved in hibernation, which are disturbed in numerous pathophysiological conditions, such as metabolic syndrome and type 2 diabetes, is a challenge to modern medicine. Hibernators are obese, insulin resistant, and hyperinsulinemic, but they do not exhibit any pathophysiological features associated with these conditions; obese hibernators lose their body weight during torpor, and they are ready for life and reproduction upon awakening, controlling crucial biochemical parameters without having vascular problems or developing hypertension (Martin, 2008).
Hibernators keep the myocardium functional during torpor, which may help us understand and prevent the state of “myocardial hibernation” in patients with chronic ischemic heart disease (Kudej and Vatner, 2003). Moreover, hibernators are more resistant than nonhibernators to ischemic conditions even in the euthermic phase, which, bearing in mind that a large percentage of patients after cardiac arrest die from brain injuries, indicates the necessity of establishing new therapeutic protocols to alleviate cerebral ischemia (Dave et al., 2006).
In addition, in nonhibernators, hypothermia <20°C–25°C induces ventricular fibrillation, which also occurs in cardiac surgery when hypothermia slows down the metabolism. Simultaneously, the time for intervention, which is proportional to the lowering of the temperature, is reduced. Hibernators have developed resistance mechanisms to ventricular fibrillation even when the heart temperature is much lower and lasts for weeks (Johansson, 1991).
Moreover, cells and tissues of hibernators are protected from death in torpor and upon awakening from it. One of the protective mechanisms is based on the cell cycle regulation and tissue-specific expression of antiapoptotic proteins, especially in the heart and brain (Rouble et al., 2013). This mechanism can help overcome various stressful conditions in humans, including oxidative stress, through the induction of AD, which is regulated by redox-sensitive transcription factors, primarily Nrf2 and FoxO (Wei et al., 2018). Increased expression of AD components before entering hibernation is an additional mechanism that creates a basis for better protection through preconditioning (Stancic et al., 2018; Vucetic et al., 2013), which opens up the possibility of prevention from oxidative damage in numerous conditions where the redox homeostasis is disturbed.
Therapeutic hypothermia induced by artificial torpor based on naturally occurring mechanisms in torpor would be useful in treating stroke and septic shock (Cerri, 2017). Cold organ ischemia during storage and transport for transplantation leads to metabolic collapse, ischemia, energy crisis, reperfusion injury, oxidative stress, edema, and autolysis. All of these processes are suppressed in hibernation, while organs and tissues are preserved at very low body temperatures without injuries at the histological and molecular level, while oxidative injuries during the transition from torpor to euthermic state (ischemia/reperfusion) upon reoxygenation are prevented (Storey, 2004).
Improving therapeutic approaches based on sophisticated protective mechanisms that evolved in hibernators against ischemic conditions would help attenuate cerebral ischemic injury of neurons in numerous neurodegenerative conditions (Dave et al., 2012), as well as in other organs from systemic inflammation (Bogren et al., 2014).
The fundamental mechanisms of hibernators are an excellent natural model for the study of aging and longevity, and may provide an answer to whether the aging process is slowed during hibernation (Lyman et al., 1981). This includes understanding the complex signaling pathways that hibernators develop in response to environmental conditions such as insulin resistance, metabolic adaptations, mitochondrial uncoupling, and stress resistance (Ulgherait et al., 2020; Wu and Storey, 2016), especially oxidative stress, with a developed strategy that is in line with Harman's free radical theory of aging (Harman, 1992).
Also, hibernators do not show any signs of bone and muscle atrophy upon awakening from torpor, and understanding the principles of this adaptive response may be useful for patients who are bedridden for long periods or cosmonauts (Andrews, 2007). Ultimately, these studies could pave the way toward new research in biomedicine, which could take hibernating astronauts to Mars and beyond (Bouma et al., 2012; Dirkes et al., 2015; Hadj-Moussa and Storey, 2019).
The beauty of life is exemplified in hibernation. The search for this knowledge is nothing but an attempt to understand the integration of spirit and matter and their existence in various states of life. That is why the hibernator, even when awake in euthermia, slowed down in hypothermia, or in hibernation, when the body temperature is 37°C, 20°C, or 0°C, may still be asleep.
Footnotes
Acknowledgments
We thank Tamara Zakic and Marta Budnar Soskic for their kind help in schematic design and figures production.
Authors' Contribution
A.J., A.K. (Aleksandra Korac), B.B., K.B.S., and B.K. wrote the article; A.J., A.K. (Andjelika Kalezic), A.K. (Aleksandra Korac), B.B., and B.K.S. performed reviewing of the article; A.J., A.K. (Andjelika Kalezic), A.K. (Aleksandra Korac), B.B., B.K.S., and B.K. performed editing of the article; and B.K. contributed to conceptualization. All authors consented to the final article.
Author Disclosure Statement
The authors declare no conflict of interest. The funders had no role in the design of the study, in the collection, analyses, or interpretation of data, in the writing of the article, or in the decision to publish the results.
Funding information
This research was funded by the Ministry of Education, Science and Technological Development of the Republic of Serbia, grant numbers 451-03-47/2023-01/200007 and 451-03-47/2023-01/200178.