
Abstract
Aims:
Calcium oxalate (CaOx) crystal deposition induces damage to the renal tubular epithelium, increases epithelial adhesion, and contributes to CaOx nephrocalcinosis. The long noncoding RNA (lncRNA) nuclear paraspeckle assembly transcript 1 (NEAT1) is thought to be involved in this process. In this study, we aimed to investigate the mechanism by which NEAT1 regulates renal tubular epithelium in response to inflammatory and oxidative injury triggered by CaOx crystals.
Results:
As CaOx crystals were deposited in mouse kidney tissue, the expression of NEAT1 was significantly elevated and positively correlated with interferon regulatory factor 1 (IRF1), Toll-like receptor 4 (TLR4), and NF-κB. NEAT1 targets and inhibits miR-130a-3p as a competitor to endogenous RNA. miR-130 binds to and exerts inhibitory effects on the 3′-untranslated region of IRF1. After transfected with silence-NEAT1, IRF1, TLR4, and NF-κB were also variously inhibited, and oxidative damage in renal calcinosis was subsequently attenuated. When we simultaneously inhibited NEAT1 and miR-130, renal tubular injury was exacerbated.
Innovation and Conclusion:
We found that the lncRNA NEAT1 can enhance IRF1 signaling through targeted repression of miR-130a-3p and activate TLR4/NF-κB pathways to promote oxidative damage during CaOx crystal deposition. This provides an explanation for the tubular epithelial damage caused by CaOx crystals and offers new ideas and drug targets for the prevention and treatment of CaOx nephrocalcinosis. Antioxid. Redox Signal. 38, 731–746.
Introduction
Kidney stone disease affects ∼5% of women and 12% of men each year and has become more common in the past 15 years (Coe et al, 2005; Scales et al, 2016). Approximately 80% of renal calculi are composed of calcium oxalate (CaOx) crystals, which are gradually deposited in the medullary collecting duct or in the renal interstitium, causing nephrocalcinosis. Although CaOx nephrocalcinosis is usually asymptomatic, it can also induce severe inflammation and renal tubular epithelial cell (TEC) necrosis, which can induce further crystal deposition and eventually lead to end-stage renal disease.
We recently found that the long noncoding RNA (lncRNA) nuclear paraspeckle assembly transcript 1 (NEAT1) is highly expressed in CaOx nephrocalcinosis, and NEAT1 is known to regulate the inflammatory response, reactive oxygen species (ROS) production, and membrane nephropathy progression (Pi et al, 2021; Yang et al, 2021; Zhao et al, 2019). The specific mechanisms by which CaOx crystals induce renal inflammation and oxidative stress injury and further crystal deposition, however, remain unclear.
Another molecule that regulates inflammation, interferon regulatory factor 1 (IRF1), can regulate the expression of IFNs or bind to STAT to regulate the expression of IRFs, which are involved in immune regulation, inflammatory response, and proliferation apoptosis. IRF1 can regulate the responses of several interferon-stimulated genes by regulating chromatin accessibility, which enhances Toll-like receptor (TLR) gene transcription and promotes TLR4 signaling pathways in immune and inflammatory responses (Kang et al, 2019; Liu et al, 2020; Qiao et al, 2013).
In addition, IRF1 can also interact with NF-κB in response to the JAK/STAT pathway and participate in ROS production, which leads to oxidative damage in the renal tubular epithelium (Eckhardt et al, 2014; Lorenzi et al, 2012). IRF1 expression was enhanced in CaOx crystal-induced renal injury in the study by Chen et al (2019). We suspect that IRF1 is involved in the regulatory process of NEAT1 on CaOx crystal-induced renal inflammation and oxidative stress injury.
Innovation
Although clinical techniques are sufficient to treat kidney stone disease, urinary stones still cause pain and financial burdens to many people each year (Scales et al, 2016). Due to the diverse etiology and unclear mechanisms of urinary stones, there are no effective pharmacological means of prevention. Deposited crystal-induced tubular damage is an important process in renal stone formation (Asselman et al, 2003; Thongboonkerd, 2019). We attempted to identify a preventive treatment for renal tubular epithelial injury. In this study, we found that nuclear paraspeckle assembly transcript 1 (NEAT1) participated in regulating calcium oxalate crystal deposition and crystal-induced renal tubular epithelial injury through the IRF1 (interferon regulatory factor 1)-Toll-like receptor 4 (TLR4)/NF-κB pathways, and NEAT1 inhibitors may be potential drugs to alleviate renal tubular injury.
A complex regulatory pathway exists during eukaryotic gene expression. lncRNAs and miRNAs, as noncoding RNAs in organisms, mainly play transcriptional regulatory roles (Anastasiadou et al, 2018). miRNAs are short-stranded small RNAs produced during pre-mRNA shearing that can target and regulate the degradation or translation of mRNAs (Fabian and Sonenberg, 2012). Recent studies have found that lncRNAs, as long-stranded RNAs with abundant miRNA binding sites, can compete for endogenous small RNAs and remove the inhibition of certain gene expressions by small RNAs. This regulatory mechanism is known as the competitor for endogenous RNA (ceRNA) mechanism (Li et al, 2014; Qi et al, 2015; Qin et al, 2018).
In our study, we sought to understand the underlying pathways regulated by NEAT1 and explored the mechanism by which miRNAs regulate CaOx nephrocalcinosis. Our findings highlighted that NEAT1 promotes the IRF1/TLR4 pathway through competition for miR130 to promote renal oxidative injury. These findings establish NEAT1 inhibitors and miR-130 agonists as prospective preventative and therapeutic agents for CaOx nephrocalcinosis.
Results
CaOx deposition in mouse kidneys induces inflammation
To discover key functional gene sets that are potential therapeutic targets for CaOx nephrocalcinosis, we performed transcriptional profiling by RNA sequencing (RNA-seq) on glyoxylate (Gly)- or saline-treated mouse kidneys (GSE192703). Gly-induced CaOx nephrocalcinosis caused the upregulation of 2596 genes and the downregulation of 2161 genes (|Log2FC| ≥ 1.5; p value <0.05) (Fig. 1A–C). Gene Ontology enrichment and differential gene expression confirmed that CaOx crystals led to the upregulation of a spectrum of inflammatory responses and responses to interferons (including IRF1 and TLR4). To further examine the effects of these genes, we built a CaOx nephrocalcinosis mouse model with increasing Gly concentrations. The 100 mg/kg group of mice showed significant crystal deposition in the renal tissue, especially in the corticomedullary junction region.
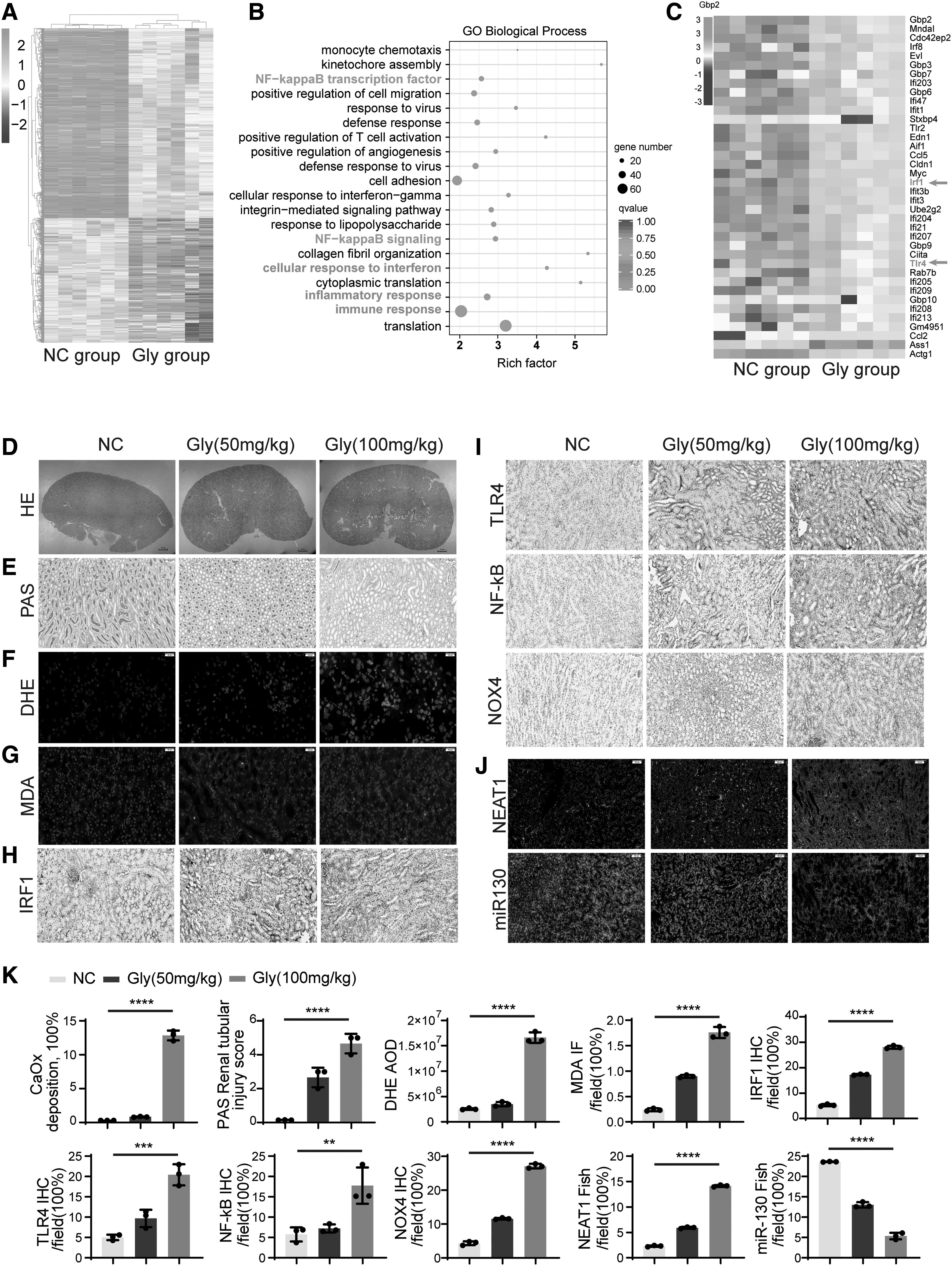
Polarized light optical microphotography, periodic acid-Schiff (PAS), dihydroethidium (DHE), and malondialdehyde (MDA) staining further confirmed that CaOx nephrocalcinosis promoted renal injury and increased CaOx crystal deposition, especially in the corticomedullary junction region, as the Gly concentration increased (Fig. 1D–G). Immunohistochemistry (IHC) staining demonstrated that IRF1, NF-κB, TLR4, and NADPH oxidase (NOX)4 dramatically increased along with CaOx crystal deposition (Fig. 1H, I, K). These data suggest that an imbalance between oxidation and antioxidation may be involved in crystal-induced tubular injury.
NEAT1 expression was significantly increased, while miR-130 expression was decreased in the CaOx nephrocalcinosis mouse model
Through database alignments (starBase and TargetScan), we found that miR-130a-3p has the potential to target and bind both IRF1 and NEAT1, and we conducted further experiments to determine whether there is a regulatory relationship between them. By fluorescence in situ hybridization (FISH) assay, we found that the lncRNA NEAT1 showed a significantly high expression status in mouse kidney tissues with CaOx crystal deposition, while miR-130 expression was reduced (Fig. 1J), and the two were negatively correlated. NEAT1 and IRF1 are key factors involved in the regulation of inflammation and signal transduction in organisms (Eckhardt et al, 2014; Liu et al, 2020; Lorenzi et al, 2012; Qiao et al, 2013; Zhao et al, 2019). The results of combined IHC, in which both IRF1 expression and NEAT1 were increased in kidneys with CaOx crystal deposition, suggest that the two are somewhat synergistic.
Oxidative stress and necrosis in renal tubular cells caused by CaOx crystals
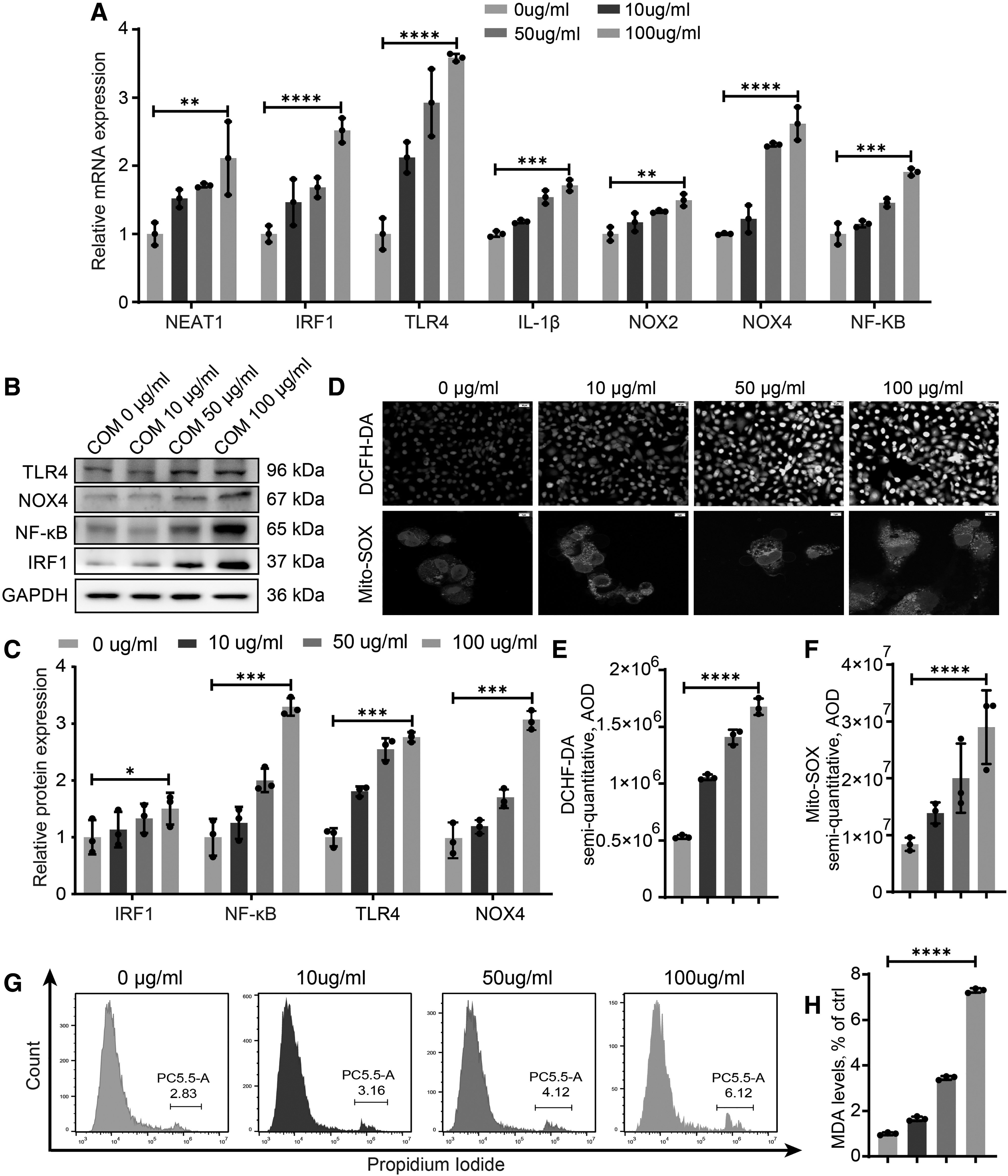
Both clinical and experimental data demonstrate that ROS generation and oxidative injury in renal tubular cells play a pivotal role in nephrocalcinosis progression. We cultured HK-2 cells with different doses of calcium oxalate monohydrate (COM) crystals to demonstrate increasing oxidative cell injury and determine the optimal COM concentration. Figure 2A–C shows that both the inflammatory and oxidative genes IRF1, TLR4, IL-1β, NF-κB, NOX2, and NOX4 increased along with NEAT1 when the COM concentration increased. In addition, the stimulation of COM crystals increased the production of NOX4, promoting a significant increase in ROS production and inducing oxidative damage. Activated IRF1 and NOX resulted in the production of more ROS, which exceeded the capacity of renal TECs to scavenge ROS (Fig. 2D–F, H). Flow cytometry analysis of propidium iodide (PI) staining showed that the rate of necrotic HK-2 cells was also increased when ROS were generated (Fig. 2G).
Inhibiting NEAT1 attenuates CaOx-induced renal inflammation and oxidative-related genes and increases miR-130 expression
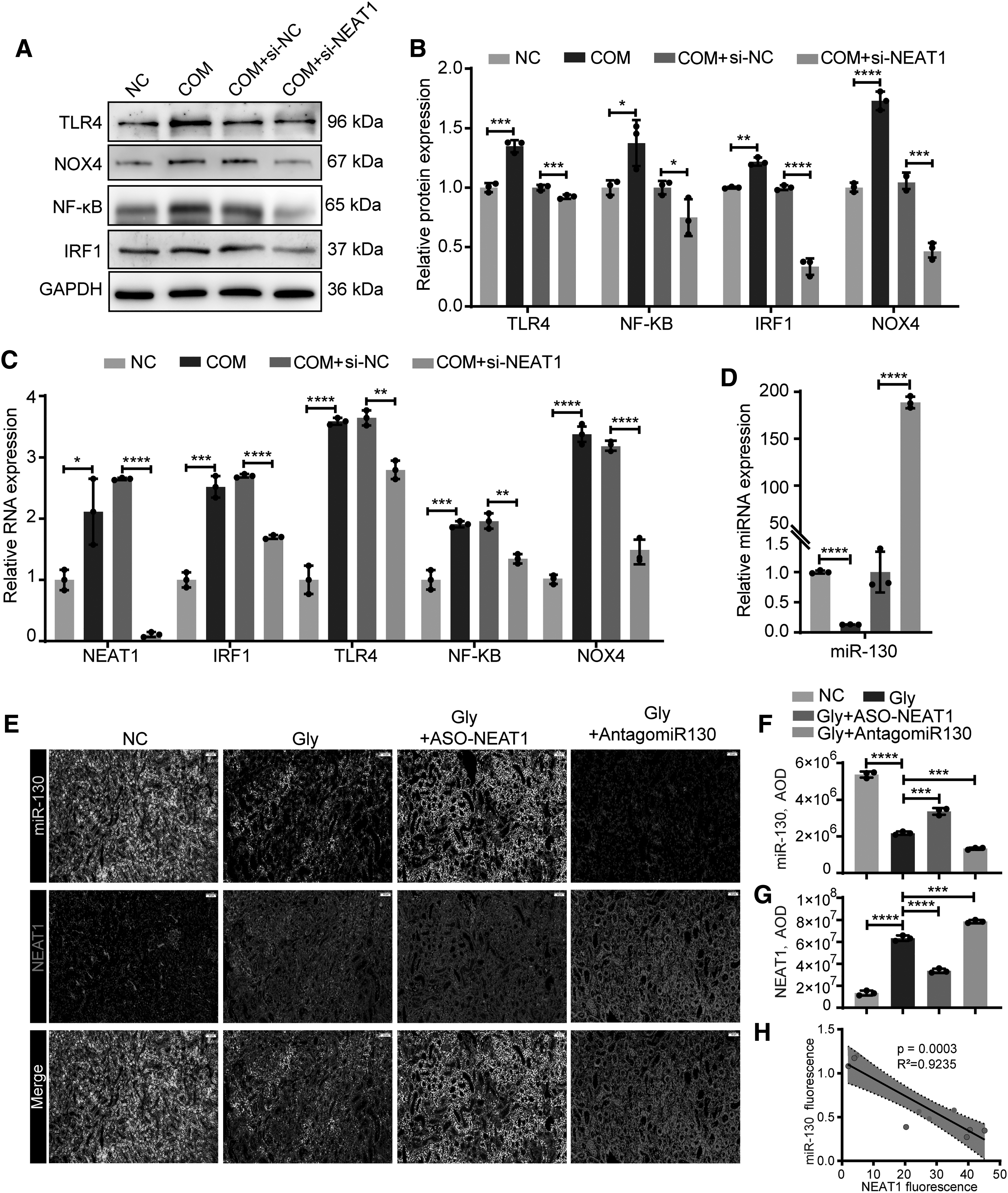
HK-2 cells were stimulated with 100 μg/mL COM for 8 h. To explore the effects of NEAT1, we pretransfected HK-2 cells with NEAT1 small interfering RNA and found that IRF1, TLR4, NF-κB, and NOX4 were significantly decreased at both the RNA and protein levels (Fig. 3A–C). The expression of miR-130 was also decreased with COM stimulation but upregulated with NEAT1 silencing (Fig. 3D). The parallel results were also shown in vivo. We silenced NEAT1 by intraperitoneal injection of antisense oligonucleotides (ASO) of NEAT1 in vivo (Fig. 3E). FISH showed significantly increased NEAT1 and decreased miR-130 staining in CaOx nephrocalcinosis mice. However, these effects were reversed by ASO-NEAT1, and we found that NEAT1 was negatively correlated with miR-130 expression (Fig. 3F–H). In summary, inhibiting NEAT1 attenuates COM-induced inflammation and oxidative damage signaling while increasing miR-130 expression.
miR-130a-3p targets the 3′-untranslated region of IRF1 and NEAT1 to inhibit their expression
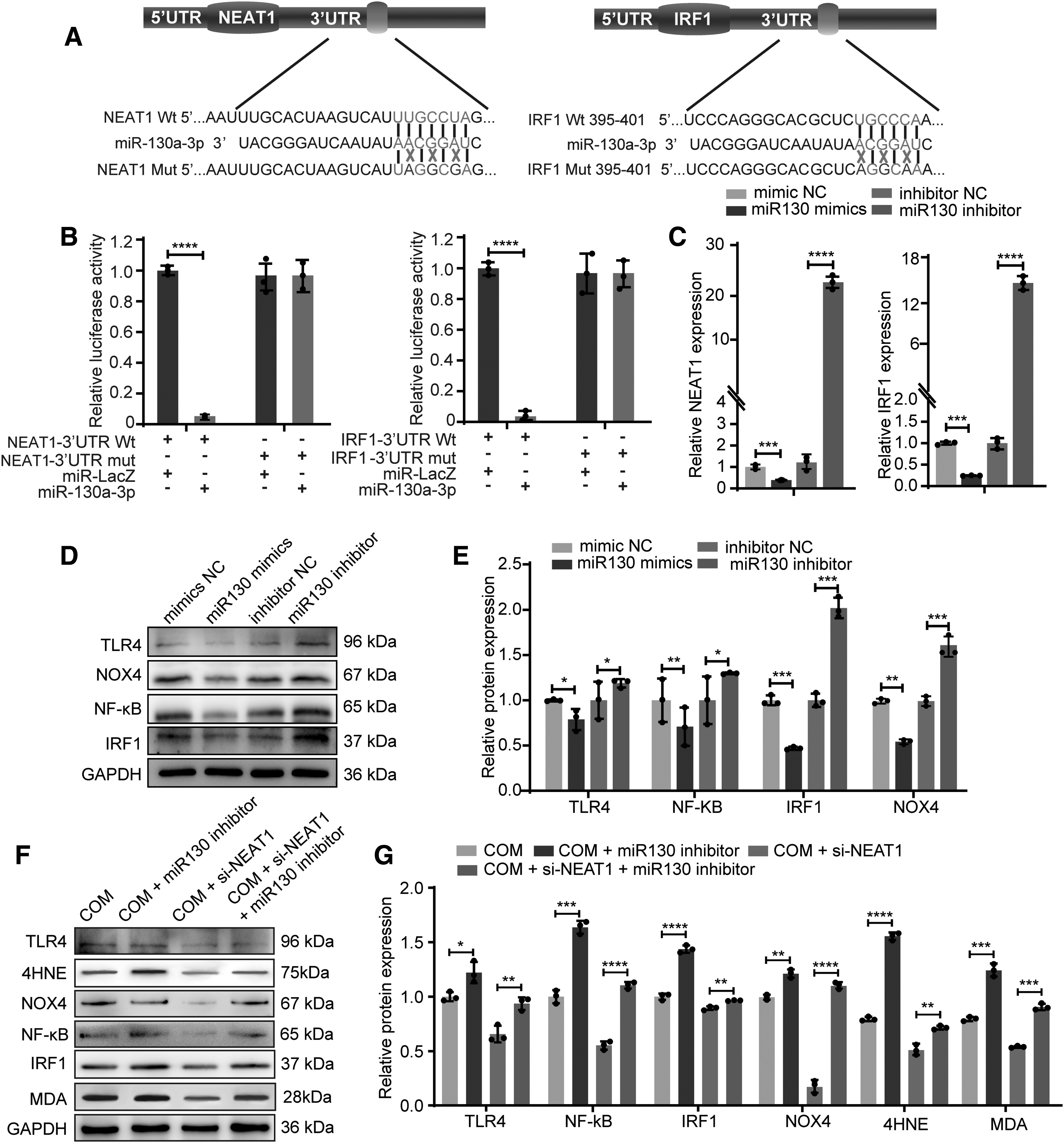
To elucidate the mechanism by which ceRNA regulates CaOx nephrocalcinosis, we focused on miR-130-3p, which was identified as a potential NEAT1 and IRF1 binding target by StarBase 3.0, TargetScan, and miRWalk. Luciferase reporter plasmids containing NEAT1 or IRF1 with a wild-type (WT) or mutant (Mut) miR-130-3p binding 3′-untranslated region (UTR) site were then constructed (Fig. 4A). The dual-luciferase reporter system showed that the luciferase activity of WT-NEAT1 and WT-IRF1 was significantly inhibited by miR-130-3p (Fig. 4B). In line with this, transfection with miR-130 mimics and inhibitors significantly lowered and elevated NEAT1 and IRF1 expression in HK-2 cells, according to polymerase chain reaction (PCR) analyses (Fig. 4C). In addition, Western blot analysis revealed that activating miR-130-3p resulted in decreases in the expression of IRF1, TLR4, NK-κB, and NOX4, while inhibiting miR-130-3p resulted in the opposite effect (Fig. 4D, E). These results indicate that miR-130-3p suppresses IRF1 and NEAT1 by targeting their 3′-UTRs.
NEAT1 regulates CaOx crystal-induced TEC inflammation and oxidative damage via miR-130
To further study the effect of NEAT1 competition for miR-130 on the inflammation and oxidative stress injury caused by COM, si-NEAT1, miR-130 inhibitor, NOX4 inhibitor, or both were cotransfected into COM-stimulated HK-2 cells. Western blot and quantitative PCR (qPCR) analysis revealed that COM-HK-2 cells transfected with si-NEAT1 exhibited lower levels of IRF1, TLR4, NK-κB, and NOX4, while cotransfection with the si-NEAT and miR-130 inhibitor reversed this effect (Fig. 4F, G, and Supplementary Fig. S2). In addition, PI labeling was used to identify COM-induced necrosis in HK-2 cells, indicating that the si-NEAT1 and NOX4 inhibitors dramatically reduced COM-induced necrosis in HK-2 cells, whereas the miR-130 inhibitor abrogated this treatment effect (Fig. 5 A, E).
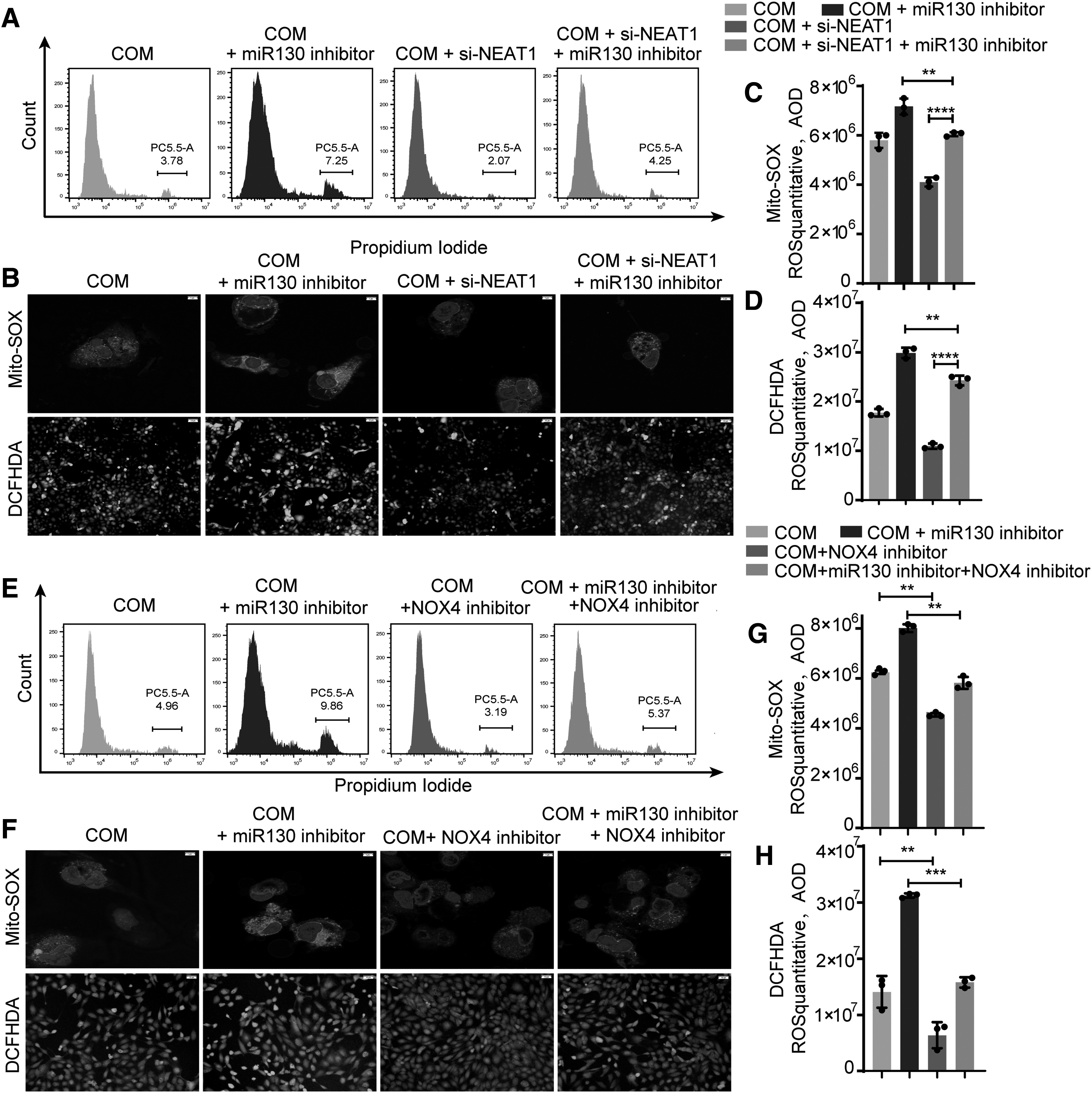
In addition, CaOx crystal-induced intracellular and mitochondrial ROS generation was measured with fluorescence and confocal microscopy. The ROS burst was enhanced by the miR-130 inhibitor and partially attenuated by si-NEAT1/NOX4 inhibitor treatment (Fig. 5B–D, F–H, and Supplementary Fig. S3). Taken together, these findings indicate that miR-130 inhibition augments the inflammatory response and ROS generation, but si-NEAT1 ameliorates these effects and reduces kidney tubular cell injury.
NEAT1 inhibitor attenuates CaOx-induced inflammatory and oxidative kidney damage via the miR-130-IRF1 pathway in CaOx nephrocalcinosis mice
To further explore the mechanism by which NEAT1 inhibits CaOx-induced renal inflammation and oxidative injury and then reduces CaOx crystal deposition, we pretreated mice with ASO-NEAT1 or AntagomiR-130 for 3 days and developed a CaOx nephrocalcinosis mouse model by intraperitoneal injection of glyoxylic acid. ROS bursts and TEC necrosis were associated with the deposition of CaOx crystals in the kidney. To evaluate how NEAT1 inhibition suppresses CaOx crystal deposition and CaOx nephrocalcinosis-mediated renal damage in vivo, we treated CaOx nephrocalcinosis mice with ASO-NEAT1, AntagomiR-130, or both. After 10 days of administration, we sacrificed the mice and collected the kidneys. Transmission electron microscope assays revealed that AntagomiR-130 accelerated mitochondrial swelling and damage in TECs, but ASO-NEAT1 had the opposite effects (Fig. 6A). In addition, similar results were obtained by measuring ROS generation levels with fluorescence microscopy (Fig. 6B, C).
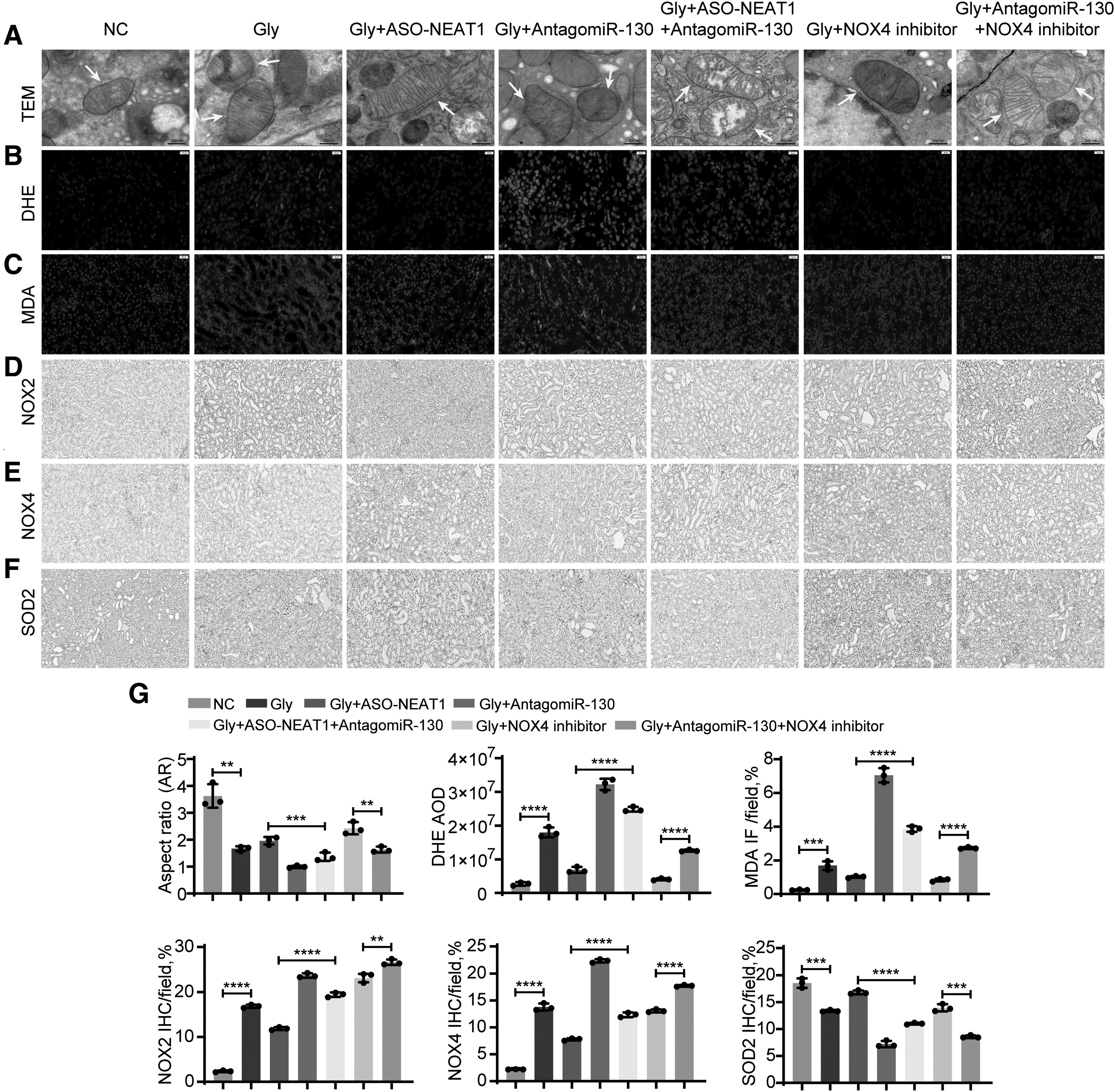
We performed IHC staining of NOX2, NOX4, and SOD2 to evaluate renal redox relate markers in CaOx nephrocalcinosis mice (Fig. 6D–G). Upon treatment with ASO-NEAT1, we observed relief in redox dyshomeostasis and renal tubular injury. However, the therapeutic effects were blocked by AntagomiR-130.
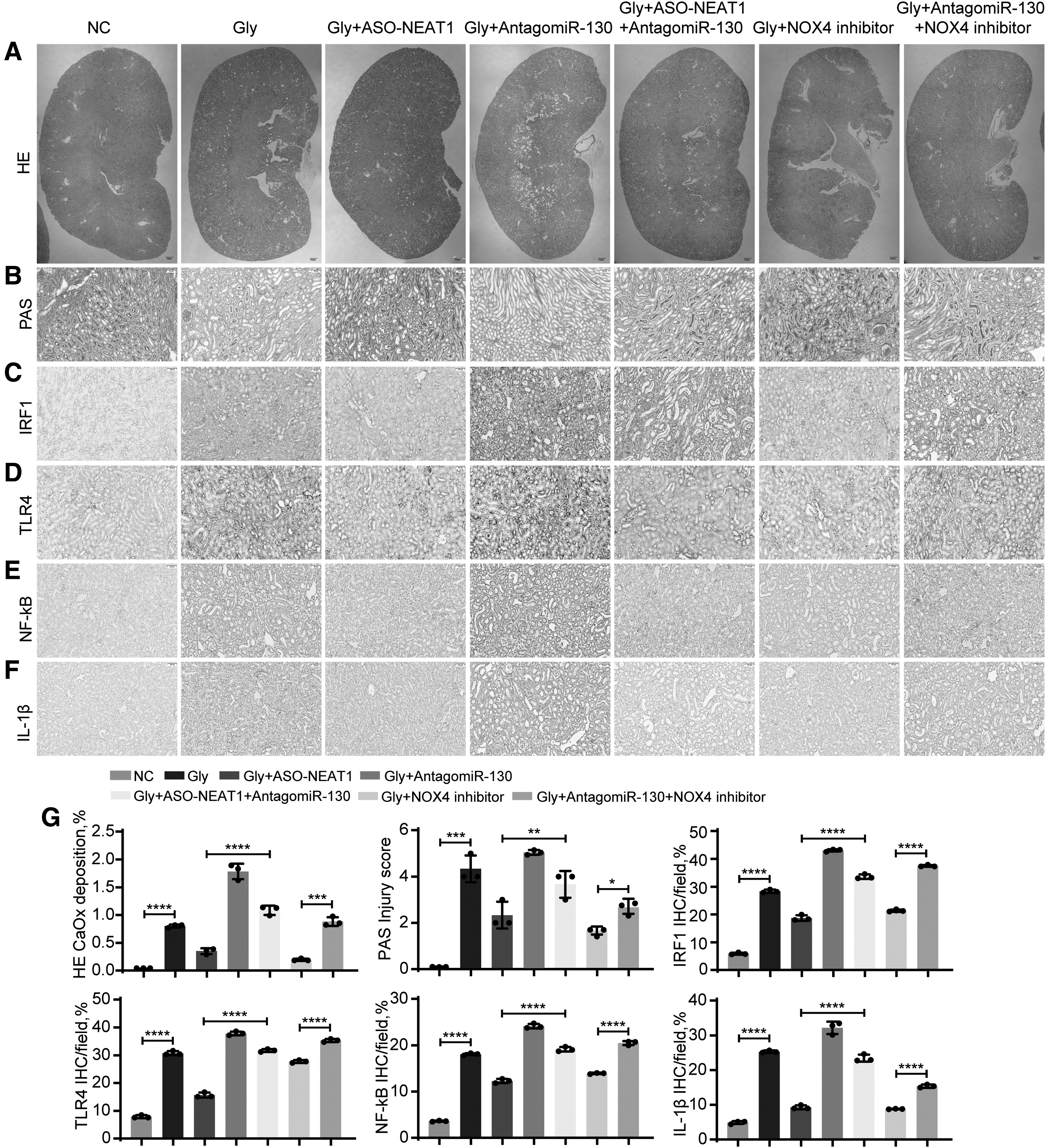
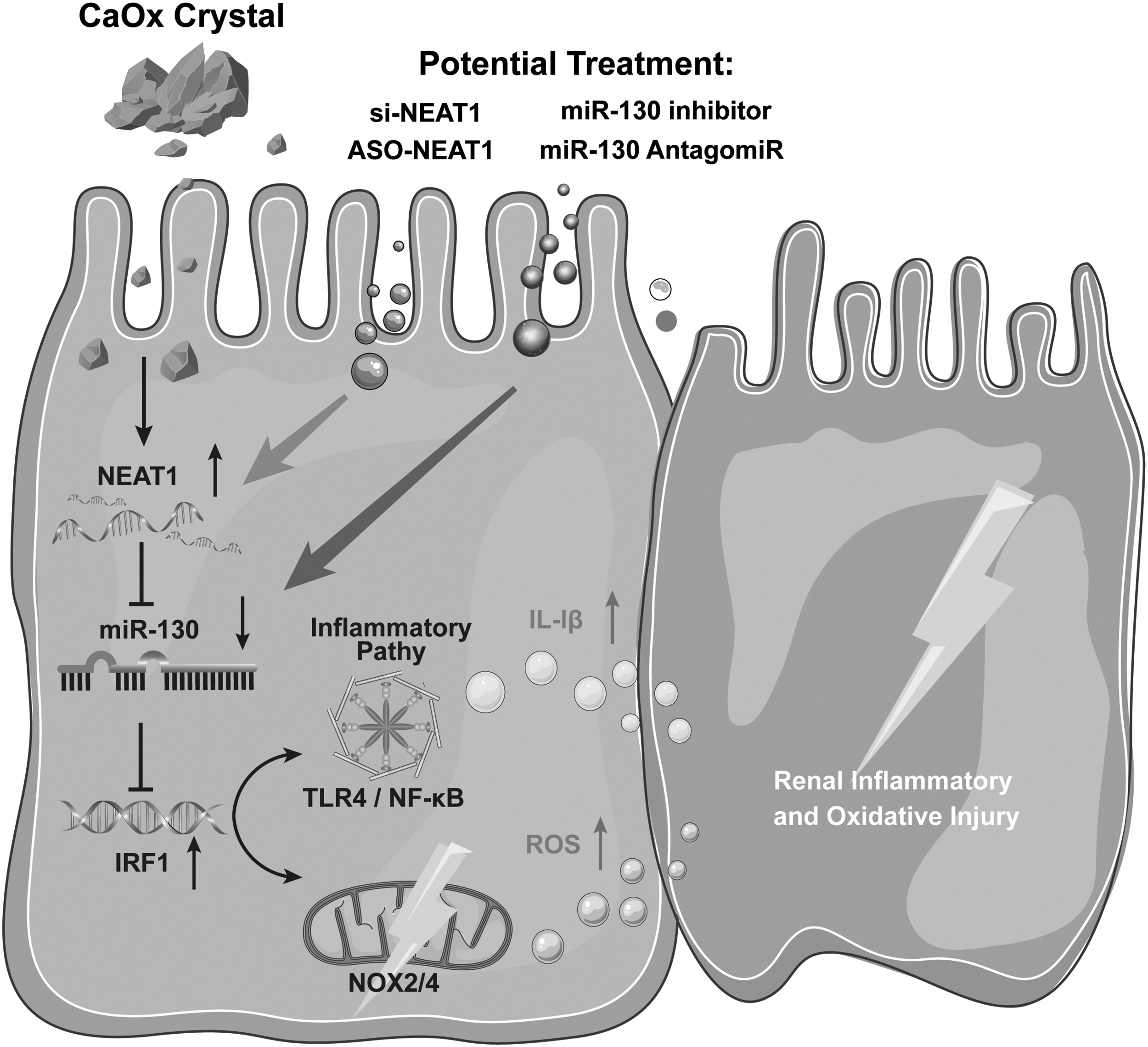
Polarized light optical microscopy and PAS staining further confirmed that AntagomiR-130 reduced the protective effect of ASO-NEAT1 on CaOx crystal-induced renal injury (Fig. 7A, B). Similar results were obtained by measuring IHC staining of inflammatory genes (TLR4, NF-κB, and IL-1β). IRF1 and proinflammatory markers were significantly decreased after ASO-NEAT1 treatment, whereas antagomiR-130 partially reversed this trend and had a significant proinflammatory effect (Fig. 7C–F). Taken together, these results indicate that the NEAT1 inhibitor decreases CaOx crystal-induced inflammatory and oxidative kidney injury by interacting with miR-130-3p and the IRF1/TLR4/IL-1β pathway (Fig. 8).
Discussion
After the removal of stones in the hospital, 50% of patients grow new stones again within 5 years, which is probably related to the presence of old lesions (Goka and Copelovitch, 2020; Vaughan et al, 2019). Previous studies have demonstrated that renal stones are closely related to the immune response and inflammatory processes (Knauf et al, 2019; Mulay et al, 2014). Crystals are known to be cytotoxic and injurious when they are in direct contact with tubular cells. The internalized crystals have been demonstrated to induce tubular cell necrosis and to release numerous damage-associated molecular patterns (Cakir et al, 2017; Dominguez-Gutierrez et al, 2020; Duan et al, 2018; Joshi and Khan, 2019). These particles activate proinflammatory pathways such as the NLRP3-IL-1β, TLR4-NF-κB, and IRF families, which amplify renal inflammatory and oxidative injury and contribute to severe crystal-induced acute kidney injury (Asselman et al, 2003; Khamchun and Thongboonkerd, 2018; Thongboonkerd, 2019).
However, the pharmacotherapeutics for kidney crystal formation remain limited. In principle, the renal inflammatory response is a key regulator of the development of stone disease. Therefore, it is urgent and pivotal to decipher the mechanism to tune down exaggerated immune responses via inflammatory signaling cascades, which can be applied as a therapeutic agent to treat CaOx-induced kidney injury.
The lncRNA NEAT1 has been highly associated with the immune inflammatory response and tissue damage in previous reports and is also involved in TLR4 and NF-κB-related pathways (Bai et al, 2018; Wang et al, 2019; Zhang and Niu, 2019a). Wang et al (2019) reported that the NEAT1-miR-342-3p axis is involved in regulating the metabolism and inflammatory responses of THP1 macrophages. Researchers such as Zhang and Niu (2019a), Gao et al (2020), Yang et al (2020) found that NEAT1 regulation is also present in sepsis-related kidney injury and diabetic nephropathy fibrosis progression, while the role of NEAT1 in CaOx crystal deposition-related damage is still unknown. In our study, NEAT1-mediated promotion of CaOx crystal nephropathy was identified.
In the field of cellular gene expression regulation, the adsorption mechanism of lncRNAs to miRNAs plays an important role in regulating gene expression at the transcriptional and translational levels (Tay et al, 2014). lncRNAs act as ceRNAs to broadly influence the process of disease development. Klec et al (2019) showed that NEAT1 can exert tumor-promoting effects by competing for miRNAs with anticancer effects, while Jiang et al (2019) also reported that NEAT1 can target miR-27a-3p in acute kidney injury progression. Bai reported that NEAT1 can contribute to the secretion of inflammatory mediators by regulating the MAPK/NF-κB pathway through the adsorption of miR-1246 (Bai et al, 2018). A similar effect was exerted by NEAT1 in our study.
We performed bioinformatic searches and analyses to screen and identify miR-130a-3p as a target of NEAT1 and IRF1, with the aim of identifying miRNAs targeted by NEAT1 in crystal deposition nephropathy. Our experimental results confirmed that miR-130 expression was negatively correlated with NEAT1 in the stone model group; NEAT1 acted as a ceRNA to adsorb miR-130, and miR-130 was also previously reported to regulate cell function and modulate the inflammatory response (Su et al, 2015; Zhao et al, 2021). miR-130 was also reported to exert anti-inflammatory effects and reduce tissue damage in diseases such as diabetic kidney injury by upregulating Klotho to inhibit Wnt/NF-κB signaling (Liang et al, 2017; Zhao et al, 2021). Ai also reported that miR-130 targets snoN and is involved in the progression of renal fibrosis through the TGF-β1/Smad pathway (Ai et al, 2020). The pro-oxidative stress effect of NEAT1 may be achieved precisely through the adsorption of miRNAs.
Our mouse model also exhibited mitochondrial stress and excessive ROS production, which were associated with the NOX4 and NF-κB pathways (Qin et al, 2018), and IRF1 could also regulate mitochondrial damage and NLRP3 activation (Zhong et al, 2018). We investigated whether NEAT1 could regulate IRF1 and was thus involved in the development of mitochondrial damage. Based on the results of FISH and dual-luciferase assays, miR-130 could target the 3′-UTR of IRF1. We inferred that NEAT1 regulates CaOx-mediated renal tubular epithelial oxidative damage via miR-130/IRF1. The activated miR-130 could reduce the expression level of IRF1, which then led to downregulation of downstream TLR4 and NF-κB pathways, exerting a therapeutic effect.
Moreover, our results also revealed that silencing NEAT1 released increased levels of miR-130, which also significantly attenuated calcium deposition injury and had a partial therapeutic effect on crystalline nephropathy. Antagonizing miR-130 partially deregulated the inhibitory effects of NEAT1 silencing on IRF1, TLR4, and NF-κB, which promoted oxidative injury. NEAT1 activates NLRP3 via IRF1 and releases inflammatory mediators such as IL-1β and IL-18 (Joshi et al, 2015; Mulay et al, 2013; Zhang et al, 2019b). The expression of NEAT1 was significantly higher in our model of renal calcium deposition than in controls and was positively correlated with TLR4, NOX4, and IL-1β expression. NEAT1 regulated oxidative damage in the renal tubular epithelium through the IRF1/TLR4 and IRF1/NF-κB pathways (Fig. 8).
Our present study revealed that NEAT1 facilitated renal calcium deposition and injury. We also revealed that the sponging effect of NEAT1 on miR-130 promoted the expression of IRF1, which activated a series of ROS responses and induced renal TEC injury (Khan et al, 2021). We used NEAT1 inhibitors and miR-130 activators to alleviate the extent of renal calcium deposition in mice, which could contribute to explaining the pathogenesis of CaOx stones and the search for potential therapeutic strategies. However, thus far, this treatment has only been effective in animals, and more work is needed.
Materials and Methods
Cell culture and transfection
The human proximal renal TEC line HK-2 was purchased from Nanjing Institute of Biological Sciences, Chinese Academy of Sciences, and cultured in Dulbecco's modified eagle medium/nutrient mixture F-12 medium with 10% fetal bovine serum in a 5% CO2, 37°C cell culture incubator.
To control the expression of NEAT1, we synthesized Silence-NEAT1 and ASO-NEAT1 (RiboBio, China) for in vitro and in vivo inhibition of NEAT1 expression (Supplementary Fig. S1), and cells were transfected with the companion ribo FECT™ CP transfection kit (RiboBio). The cells were evenly distributed in six-well plates at a density of ∼60%, and 100 nM silence-NEAT1 was diluted and mixed with 120 μL of 1 × ribo FECT CP buffer in each well. Then, 12 μL of ribo FECT CP reagent was added and incubated for 10 min at room temperature, after which cell culture medium was added and mixed well. After 24 h, subsequent treatments, such as CaOx crystal coculture, ROS analysis, cell necrosis analysis, and total protein and RNA extraction, were performed.
To activate and inhibit miR-130a-3p expression, we used miR-130 mimics/inhibitor (RiboBio) products for cell and animal experiments; cell transfection was also performed with a ribo FECT CP transfection kit by adding 50 nM mimics or inhibitor to six-well plates, and the relevant indices were measured after 24–48 h.
Animal experiments
Specific pathogen free-rated C57BL/6J male mice (6–8 weeks old) were purchased from the Animal Center of Anhui Medical University and used for the experiments. Glyoxylic acid (Sigma) was used to construct a mouse model of CaOx crystal deposition. Mice will be anesthetized with 3%–5% isoflurane (inhalation) at 1–3 L/min and maintained with a mask at 1%–3% at 0.5–1 L/min. After being anesthetized, glyoxylic acid will be injected intraperitoneally at doses of 50 and 100 mg/kg once daily for 7 days. Long-acting miR-130 antagonist (antagomir-130) and ASO-NEAT1 were synthesized by RiboBiotech (Guangzhou, China). Mice received AntagomiR-130 (20 mg/kg; 200 μL) and ASO-NEAT1 (10 nM; 200 μL) through the tail vein on days 1, 4, and 7. Mice at the end of the procedure above will be euthanized by cervical dislocation under isoflurane anesthesia. Then, the kidneys were removed for relevant staining observations according to the operation.
This study was approved by the Animal Ethics Committee of Anhui Medical University (Animal Study Permit No: LLSC20211520).
Western blotting
COM crystals were cocultured with HK-2 cells at a concentration of 100 μg/mL. After 48 h, total cellular proteins were extracted using RIPA buffer (Absin, China), and a bicinchoninic acid kit was used to determine the protein concentration and calculate the loading volume. Loading buffer was added, and the samples were boiled for 10 min to fully denature the proteins. The gel was prepared and loaded, and the samples were separated by electrophoresis and transferred to a membrane, which was incubated with primary antibodies against IRF1 (A00580-1, 1:1000; Boster, China), TLR4 (BA1717, 1:5000; Boster), NF-κB (abs131170, 1:1000; Absin), NOX4 (abs112974, 1:2000; Absin), MDA (ab243066, 1:1000; Abcam, China), 4HNE (ab46545, 1:2000; Abcam), and glyceraldehyde 3-phophate dehydrogenase (GAPDH) (abs132004, 1:5000; Absin) at 4°C for 8 h after being sealed. Then, the membrane was washed and incubated with secondary antibodies at room temperature for 1 h.
An electrochemiluminescence solution was used to develop the blot, and ImageJ software was used for semiquantification. We quantified the proteins by calculating the ratio of the target protein to GAPDH and then normalizing the experimental group data by the mean of the control data (or transfected control groups). All the original blots are shown in the Supplementary Figure S4.
Real-time qPCR
After HK-2 cells were transfected and/or cocultured with CaOx crystals for 24 h, total cellular RNA was extracted using the TRIzol method. The RNA concentration was then measured by a NanoDrop 2000 (Thermo Fisher Scientific, Wilmington, DE). The reverse transcription and amplification process was performed using the HiScript II 1st Strand cDNA Synthesis Kit (Vazyme Biotech Co., China). Real-time qPCR was performed with SYBR Green Master Mix (11201-11203; Yeasen) to assess RNA expression levels on an ABI 7500 qPCR instrument. GAPDH and U6 were used as internal reference genes, and miR-130a-3p was amplified using the stem–loop method. The sequence information of the primers was as follows: TLR4 forward: 5′-AGACCTGTCCCTGAACCCTAT-3′, reverse: 5′-CGATGGACTTCTAAACCAGCCA-3′; IRF1 forward: 5′-CTGTGCGAGTGTACCGGATG-3′, reverse: 5′-ATCCCCACATGACTTCCTCTT-3′; GAPDH forward: 5′-TCGCTCCTGGAAGATGGTGAT-3′, reverse: 5′-CAGTGGCAAAGTGGAGATTGTTG-3′; IL-1β forward: 5′-AGCTACGAATCTCCGACCAC-3′, reverse: 5′-CGTTATCCCATGTGTCGAAGAAGAA-3′;
NOX2 forward: 5′-TGCCAGTCTGTCGAAATCTGC-3′, reverse: 5′-ACTCGGGCATTCACACACC-3′; NOX4 forward: 5′-TGACGTTGCATGTTTCAGGAG-3′, reverse: 5′-AGCTGGTTCGGTTAAGACTGAT-3′; NF-κB forward: 5′-AACAGAGAGGATTTCGTTTCCG-3′, reverse: 5′-TTTGACCTGAGGGTAAGACTTCT-3′; NEAT1 forward: 5′-GCCTTCTTGTGCGTTTCTCG-3′, reverse: 5′-TCCCAGCGTTTAGCACAACA-3′; U6 forward: 5′-CTCGCTTCGGCAGCACA-3′, 5′-AACGCTTCACGAATTTGCGT-3′; hsa-miR-130a-3p stem–loop: 5′-GTCGTATCCAGTGCAGGGTCCGAGGTATTCGCACTGGATACGACATGCCC-3′, forward: 5′-CGCGCAGTGCAATGTTAAAA-3′, reverse: 5′-AGTGCAGGGTCCGAGGTATT-3′.
RNA-seq analysis
Total RNA was extracted from the tissues in RNAlater using an RNeasy Kit (Qiagen). RNA concentration and purity were measured using NanoDrop 2000 (Thermo Fisher Scientific). A total amount of 1 μg RNA per sample was used as input material for the RNA sample preparations. Sequencing libraries were generated using the NEBNext Ultra™ RNA Library Prep Kit for Illumina (NEB). The RNA-seq data used in this study were generated in this article and are available in the Gene Expression Omnibus database (GSE192703). The raw reads were further processed with a bioinformatic pipeline tool, BMKCloud.
Enzyme-linked immunosorbent assay
Oxidative stress markers, including MDA and 8OHdG, were detected in cell homogenates from different treatment groups using enzyme-linked immunosorbent assay kits (E-EL-0028c and E-BC-K028-M; Elabscience, China). The optical density values of 8OHdG and MDA were detected at 450 and 532 nm, respectively, and the concentrations were calculated using the standard curve.
Tissue immobilization and coloration
The kidneys were removed from the envelope, and the tissue was fixed using 4% paraformaldehyde, paraffin embedded, sectioned, dewaxed, and stained. After hematoxylin–eosin staining, the distribution of deposited crystals in the kidney section was observed using polarized light microscopy. The degree of crystal deposition was quantified using Image-Pro Plus. Through PAS staining, glycogen depletion in renal TECs and tubular lumen changes were observed. The tubular lesions were graded on a scale from 0 to 5 as follows: 0 (none), 1 (<10%), 2 (11%–25%), 3 (26%–45%), 4 (46%–75%), and 5 (>76%). Kidney damage was scored according to the following four aspects: tubular necrosis, epithelial cell apoptosis, intraluminal cast formation, and brush border loss.
For IHC, sections were incubated with the primary antibodies: TLR4 (66350-1-Ig, 1:400; Proteintech, China), SOD2 (24127-1-AP, 1:1000; Proteintech), IRF1 (A00580-1, 1:200; Boster), NF-κB (abs131170, 1:200; Absin), IL-1β (abs126104, 1:200; Absin), and NOX4 (abs136444, 1:100; Absin). After being treated with 10 mM sodium citrate to retrieve antigen, the slides were incubated with an endogenous peroxidase blocking solution, and incubated with a primary antibody at 4°C overnight. Then the slides were incubated with a biotin-conjugated secondary antibody and with an enzyme-conjugated horseradish peroxidase (HRP)–streptavidin. 3,3′-Diaminobenzidine was used as substrate to detect HRP. After staining, gradient dehydration and transparent sealing, the images were acquired using a high-throughput Leica SCN400 scanner.
DHE staining
Fresh kidneys that were removed from the renal capsule were frozen and embedded in optimum cutting temperature matrix (SAKURA Tissue-Tek). Frozen renal sections were incubated with 5 μM DHE (Beyotime, S0063, China) at 37°C for 30 min against the light, after which excess dye was removed by three washes with phosphate-buffered saline (PBS) three times. The images were observed under fluorescence microscopy (Olympus IX73, Japan) at 535/610 nm wavelengths. ROS can oxidize DHE to ethidium oxide, which can integrate into DNA and show red fluorescence. The level of ROS was assessed according to the intensity of red fluorescence by ImageJ.
Electron microscopy
Mouse kidneys were removed and dissected, and the tissue at the corticomedullary junction in the middle of the kidney was preserved and fixed with glutaraldehyde electron microscopy fixative. Sections were processed for electron microscopic observation to locate the number of mitochondria in the 5-μm range around the crystal and then 500 nm to observe the mitochondrial ultrastructural changes.
Cell staining and flow cytometry
To detect intracellular and mitochondrial ROS generation and cell necrosis, HK-2 cells were seeded on coverslips in six-well plates at 50%–70% density, cocultured with CaOx crystal or transfection reagents, and then stained with a 2,7-dichlorodihydrofluorescein diacetate (DCFH-DA) assay kit (S0033s; Beyotime, China), Mito-SOX kit (M36008; Thermo Fisher), and PI cell necrosis staining kit (abs9358; Absin). DCFH-DA, Mito-SOX, and PI probes were diluted at 1:1000, 1:2000, and 1:1000, respectively, using a serum-free medium and incubated at 37°C for 10–20 min protected from light. Unbound probes were washed away with PBS three times. Then, the cells were detected by flow cytometry (BD Bioscience) and laser confocal microscopy (Olympus) with excitation and emission at 488/525 nm (DCFH-DA), 510/580 nm (Mito-SOX), and 535/617 nm (PI), respectively. The levels of ROS generation were quantified with ImageJ software.
Dual-luciferase reporter gene assay
To construct reporter gene plasmids, we first used starBase and TargetScan to predict the binding sites of miR-130 targeting NEAT1 and the IRF1 3′-UTR. Then, we artificially mutated the binding site sequence and inserted the sequence into the luciferase vector (pmirGLO). Finally, we obtained vectors containing the WT sequences pmirGLO-NEAT1-WT and pmirGLO-IRF1-WT and the mutant sequence vectors pmirGLO-NEAT1-Mut and pmirGLO-IRF1-Mut. HK-2 cells were cotransfected with miR-130 mimics or negative-control miRNA along with either the WT or mutant 3′-UTR reporter plasmid by using Lipofectamine 3000 (Invitrogen). After 2 days of transfection, luciferase activity was measured using a dual-luciferase reporter assay system (Promega, Madison, WI). Firefly luciferase activity is expressed as relative expression levels and normalized to Renilla luciferase activity.
Fluorescence in situ hybridization
FISH was used to measure the expression of NEAT1 and miR-130 in animal models. Kidney samples were collected, paraffin sectioned, dehydrated by dewaxing, boiled using repair solution, and then cooled naturally before being treated with proteinase K at 37°C for 20 min. Specific probes for NEAT1 (-CY3) and miR-130 (-FAM) (GDP1073 and GDP1071; Servicebio, China) were synthesized, incubated at 37°C for 12 h, and then washed three times. After DAPI was used to stain the nucleus, the cells were observed and scanned by fluorescence microscopy. NEAT1 binding-CY3 was red light, and miR-130 binding-FAM was green light. Semiquantitative analysis was performed using ImageJ. RNA expression is expressed as the average optical density value.
Statistical analysis
GraphPad Prism 8.4 was used to analyze the data. The experimental data are shown as the mean ± standard deviation. We performed t-tests and one-way analysis of variance (ANOVA) to compare the differences among the groups. p Values <0.05 were considered statistically significant.
Use of electronic laboratory notebook
An electronic laboratory notebook was not used.
Footnotes
Acknowledgments
We thank our colleagues in the laboratory of urology at The First Affiliated Hospital of Anhui Medical University.
Authors' Contributions
Q.Y., Q.H., L.H., and H.Z. contributed to the topic design, experimental process, statistical analysis, and discussion of the results. Q.Y. and Q.H. helped summarize and write this article. Q.Y., G.L., S.J., and S.Z. performed the rearing and modeling of the experimental animals for this experiment. Q.H., H.L., and Q.Q. made contributions to the cytology experimental section. All authors have confirmed the final version of the article.
Author Disclosure Statement
No competing financial interests exist.
Funding Information
The National Natural Science Foundation of China (82070724, 82100808), the Natural Science Foundation of Anhui Province (1908085MH246), and Postdoctoral researchers in Anhui Province (2021A482).
Supplementary Material
Supplementary Figure S1
Supplementary Figure S2
Supplementary Figure S3
Supplementary Figure S4
Abbreviations Used
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.