
Abstract
Significance:
A growing body of evidence has demonstrated that the commensal microbiome is deeply involved in the host immune response, accounting for significantly divergent clinical outcomes among cancer patients receiving immunotherapy. Therefore, precise screening and evaluating of functional bacterial strains as novel targets for cancer immunotherapy have attracted great enthusiasm from both academia and industry, which calls for the construction and application of advanced animal models to support translational research in this field.
Recent Advances:
Significant progress has been made to elucidate the intervention effect of commensal microbiome on immunotherapy based on animal experiments. Especially, correlation between gut microbiota and host response to immunotherapy has been continuously discovered in a variety of cancer types, laying the foundation for causality establishment and mechanism research.
Critical Issues:
In oncology research, it is particularly not uncommon to see that a promising preclinical result fails to translate into clinical success. The use of conventional murine models in immunotherapy-associated microbiome research is very likely to bring discredit on the preclinical findings. We emphasize the value of germ-free (GF) mice and humanized mice as advanced models in this field.
Future Directions:
Integrating rederivation and humanization to generate humanized GF mice as preclinical models would make it possible to clarify the role of specific bacterial strains in immunotherapy as well as obtain preclinical findings that are more predictive for humans, leading to novel microbiome-based strategies for cancer immunotherapy. Antioxid. Redox Signal. 37, 1291–1302.
Introduction
Immunotherapy has led a promising direction of cancer treatment, realizing more lasting efficacy and fewer side effects compared with radiation or chemotherapy (Sanmamed and Chen, 2018; Wei et al, 2018). Still, a large proportion of cancer patients do not respond to immunotherapy and some patients could even experience hyperprogressive disease (HPD) during treatment (Borcoman et al, 2019; Haslam and Prasad, 2019; Kim et al, 2019; Ribas et al, 2016). Notably, commensal microbiome has been frequently indicated to be closely related to cancer immunotherapy efficacy, and the exploitation of gut microbiota resources to modulate host immune response has attracted enormous attention from academia and industry (He et al, 2021; Roviello et al, 2022; Zhou et al, 2021).
Understanding how specific bacterial strains are involved in the host immune response is of great importance to the development of microbiome-based strategies to optimize cancer immunotherapy. However, for immunotherapy, preclinical results based on conventional animal models often fail to translate corresponding molecular mechanisms in humans, let alone predict clinical outcomes (Dholakia et al, 2021; Groisberg et al, 2019; Seyhan, 2019; Waldman et al, 2020). In this context, advanced animal models for screening and evaluation of immunotherapy-associated functional bacteria are undergoing rapid evolution.
In this study, we summarize current studies in depicting the potential role of commensal microbiome in cancer immunotherapy. We discuss the construction of germ-free (GF) mouse strains and humanized mouse models along with their value in cancer immunotherapy research. With the increasing need for researchers to elucidate the intricate relationship between microbiome and immunotherapy efficacy as well as improve predictability of preclinical studies in this field, more advanced mouse models should be developed.
Commensal Microbiome in Cancer Immunotherapy
Microbiome and immune checkpoint blockade
In the past 10 years, research and clinical application of immunotherapies, including immune checkpoint blockade (ICB), targeted antibodies, adoptive T cell transfer, cancer vaccines, and oncolytic viruses in a growing range of cancer types, have reflected the importance of modulating the immune system in cancer treatment (Dobosz and Dzieciatkowski, 2019; Mellman et al, 2011; Oiseth and Aziz, 2017).
Remarkably, immunotherapy using immune checkpoint inhibitors (ICIs) targeting programmed cell death protein 1/programmed cell death ligand 1 (PD-1/PD-L1) and cytotoxic T lymphocyte-associated antigen 4 (CTLA-4) has led vast advances in cancer treatment over the past decade (Sanmamed and Chen, 2018; Wei et al, 2018). ICIs bring fewer side effects than traditional radiotherapy and chemotherapy through activating patients' own T cell immune response and the approved indications of ICIs are increasing (Vaddepally et al, 2020).
However, the clinical response to ICB is heterogeneous between different individuals and it is still difficult to achieve a durable response even in selected patients (Borcoman et al, 2019; Haslam and Prasad, 2019; Kim et al, 2019; Ribas et al, 2016). There is mounting evidence supporting that patients' commensal microbiome is closely related to the efficacy of ICIs.
Considering that the gastrointestinal tract is the largest immune organ with a huge microbial load (McDermott and Huffnagle, 2014), modulating the composition of gut microbiota has been regarded as a promising approach to improve the host response to ICB. In a study investigating the anticancer efficacy of CTLA-4 inhibitor ipilimumab, microbiota composition was found to be a key factor. Certain Bacteroides species in both tumor-bearing mice and patients facilitate ipilimumab treatment probably through modulating interleukin-12 (IL-12)-dependent TH1 immune responses (Vetizou et al, 2015).
Routy et al found that the progression-free survival and overall survival of patients with advanced nonsmall-cell lung cancer and renal cell carcinoma who had taken antibiotics before and after the first dose of anti-PD-1 immunotherapy are significantly lower than those of patients who did not take antibiotics. The subsequent animal experiments showed that supplementing tumor-bearing mice with Akkermansia muciniphila can induce dendritic cells (DCs) to secrete IL-12, which in turn recruits CCR9+CXCR3+CD4+ T cells into tumor beds and restores immunotherapy sensitivity (Routy et al, 2018).
In another study with regard to the effects of microbiome on the responses of melanoma patients receiving anti-PD-1 therapy, high diversity and abundance of Ruminococcaceae and Faecalibacterium were proposed to be favorable for enhancing the antitumor efficacy, while low diversity and high relative abundance of Bacteroidales were related to reduced tumor infiltrating immunocytes and dampened antigen presentation capacity (Gopalakrishnan et al, 2018).
Recently, two independent phase I clinical trials of anti-PD-1 immunotherapy for patients with melanoma in Israel and the United States reported that transplanting the gut microbiota from responders to nonresponders can activate CD8+ T cells and the corresponding immune cascade in the recipients' tumor microenvironment (TME) (Baruch et al, 2021; Davar et al, 2021).
On the contrary, immune-related inflammatory side effects have caused widespread concern with the expansion of ICIs' application. The occurrence of such adverse events differs in patients, in which the potential role of the composition of host microbiota needs to be fully elucidated (Postow et al, 2018). Clinically, ICI-induced colitis is a notable autoimmune toxicity caused by enhanced T cell responses after administration of ICIs, and the handling of this problem is critical to continuous treatment and patient outcome (Abu-Sbeih et al, 2018; Chang et al, 2021; Dougan et al, 2021; Michot et al, 2016).
The first prospective study to characterize the composition of gut microbiota before the development of CTLA-4 blockade-induced new-onset colitis demonstrated that members of the Bacteroidetes phylum were correlated with resistance to colitis in a subset of patients (Dubin et al, 2016). Data from an induced mouse model showed that decreased abundance of Lactobacillus was associated with severe ICI colitis after ICB treatment. This toxicity could be suppressed by supplementing probiotic Lactobacillus reuteri, which dramatically diminished the mucosal numbers of group 3 innate lymphoid cells (ILC3s) triggering the protective effect (Wang et al, 2019).
Wang et al (2018) reported the first case series that refractory ICI-induced colitis was abrogated with fecal microbiota transplantation (FMT) probably through affecting the populations of host CD8+ cytotoxic T lymphocytes (CTLs) and CD4+ FoxP3+ T cells, although the cohort in this study only includes two patients and the utility of this modality needs further validation. Despite the fact that the crucial role of microbiota in ICB therapy has been increasingly recognized and new discoveries are being reported continuously, some key issues including the applicability of certain “beneficial” bacteria in assisting ICB across cancer types and the criteria for selecting FMT donors and recipients remain ambiguous.
These problems underscore the need for more rigorous in vivo preclinical studies so as to defuse latent risks of FMT in the context of ICB and guide clinical practices in this field (Elkrief and Routy, 2021; Fillon, 2021; Woelk and Snyder, 2021; Zipkin, 2021).
Microbiome and chimeric antigen receptor-T cell therapy
Likewise, much more knowledge is required to unearth the likely potential of commensal microbiome in other cancer immunotherapies. Because they follow some shared immunological principles such as the recognition of tumor-specific antigens, the activation of cytotoxic T cells, and the release of cytokines, to stimulate a patient's immune response. For example, chimeric antigen receptor (CAR)-T cell therapy, by genetically modifying T cells harvested from a patient's or a healthy donor's peripheral blood and infusing them into the patient, is capable of inflicting targeted cytotoxicity on tumor cells with specific antigens on their surface (Sadelain, 2017). Similar to ICB, the outcomes of CAR-T cells are heterogeneous among patients and cancer types and the overall response rate still needs to be improved.
As one of the latest strategies especially in the treatment of hematological neoplasms, CAR-T cell therapy has also aroused research interest in speculating its engagement with the commensal microbiome ever since the FDA approved the first two CAR-T cell therapies (Maude et al, 2018; Neelapu et al, 2017) for the treatment of B cell acute lymphoblastic leukemia and large B cell lymphoma, respectively. As early as 2017, researchers addressed that broad-spectrum antibiotics impaired the sustained curative effect of adoptive CD4+ T cells in tumor-bearing mice. In contrast, depletion of the gut microbiota of mice with B cell lymphoma through antibiotic administration did not show any impact on the efficacy of CD19+ CAR-T cell therapy although it was connected with enhanced CAR expression and declining CD19+ B cells in the spleens (Kuczma et al, 2017).
Interestingly, however, in a retrospective study, intestinal microbes were identified as biomarkers for the efficacy and toxicity of the therapy. Bacterial taxa, including Oscilliospiraceae, Ruminococcaceae, and Lachnospiraceae, were found enriched in the cohort of “complete response (CR),” while Peptostreptococcaceae in “no CR.” The analysis of fecal samples from patients who later developed CAR-T cell-induced cytokine release syndrome (CRS) or neurotoxicity showed a higher abundance of Lachnospiraceae, while the richness of Peptostreptococcaceae was found in patients free from toxicity (Smith et al, 2018). Another in vitro cellular assay and syngeneic murine model-based study highlighted the role of two microbial metabolites pentanoate and butyrate in augmenting the therapeutic outcome of CTLs and CAR-T cells (Luu et al, 2021).
At present, the number of published researches depicting the possible interplay between commensal microbiome and the efficacy and adverse events of CAR-T cell therapy is still far fewer than that of ICB, and various speculations concerning this issue are yet to be confirmed. It is mainly because CAR-T cells are now only applicable to limited cancer types and limited number of patients and the intestinal environment in recipients of CAR-T cells has often been affected by chemotherapies previously received, leading to faltering attempts to draw any conclusions about that (Abid et al, 2019; Innao et al, 2020). Hence, reproducible in vivo preclinical studies preferably based on animal models with controlled microbial background are recommended to demonstrate if the relationship between host microbiome and CAR-T cell therapy exists and clarify the underlying mechanisms.
In addition, a growing number of studies are trying to incorporate the ICB function into CAR-T cells either by PD-1 gene knockout or in situ secretion of anti-PD-1 proteins (Chen et al, 2021; McGowan et al, 2020; Suarez et al, 2016), in the hope of enhancing the response rates and expanding the currently limited application scope of CAR-T cell therapy. Given the aforementioned findings on ICB, commensal microbiome could be an important influencer in determining the outcomes of these novel immunotherapies as well.
The role of extraintestinal microbiota in immunotherapy
Although huge preclinical and clinical attention has been drawn to the multiple effects of the gut microbiota on cancer immunotherapy, the role of extraintestinal microbiota, especially in nongastrointestinal cancers, is also highlighted by researchers. Nejman et al analyzed the intratumor bacteria from patients with metastatic melanomas to see if there exist associations between intratumor microbiota and a differential host response to ICB. Indeed, they found that patients who responded to ICB possessed more abundant Clostridium in tumors, whereas enrichment of Gardnerella vaginalis was significant in tumors from nonresponders to ICB (Nejman et al, 2020). In another study, evidence showed that airway dysbiosis with Veillonella parvula was associated with a poor prognosis. This microbiota signature could affect the response to immunotherapies through modulating the IL-17-driven inflammatory pathway (Tsay et al, 2021).
It is also noteworthy that the accumulation of specific commensals within the TME, which is capable of intervening immunotherapy, could be attributed to gut microbiota translocation. For example, Bifidobacterium from the gastrointestinal was reported to migrate to the TME in tumor-bearing mice and enhance the efficacy of anti-CD47 immunotherapy through activating the stimulator of interferon genes (STING) signaling pathway inside DCs (Shi et al, 2020). A recent study also validated that intratumoral monocytes produce type I interferon in response to microbiota-derived STING agonists, leading to improved ICB efficacy in melanoma patients (Lam et al, 2021).
GF Mice and Their Value in Cancer Immunotherapy Research
The necessity of the application of GF mice
An animal model that excludes the possible influence of native microbial background is an ideal choice for FMT and antibiotic-treated mouse models are broadly used because of easy establishment process and low cost. Nevertheless, the sterile state created by antibiotics is often transient because some remaining bacteria will proliferate again under proper conditions. What is more, the mice that undergo such an antibiotic-involved microbiota depletion stage could suffer great harm to their physiological state (Leclercq et al, 2017; Sun et al, 2021; Wienhold et al, 2018), which would weaken the authenticity of the subsequent in vivo functional experiments implemented to investigate the immunomodulatory effects of the transplanted microbiota.
Therefore, to avoid unwanted changes to the physiology of conventional mice caused by the halfway sterilization process, it is highly advocated that the preclinical in vivo evaluation of the influences of specific microbiota on cancer immunotherapy should preferably be conducted on cancer cell line-inoculated GF mice (Gopalakrishnan et al, 2018; Matson et al, 2018; Routy et al, 2018).
GF animals and rederivation of mouse models
GF animals refer to animals without colonization of microorganisms and parasites in or on their body since birth. These animals are bred and raised under sterile conditions (usually in special isolators), being completely isolated from possible contaminations (Kamareddine et al, 2020; Li and Wei, 2019). The elimination of microbes makes GF animal models valuable tools for studies on the interplay between microbiome and hosts.
First, the huge amounts of commensal bacteria play a critical role in regulating the health of the human body throughout the development of various diseases. In vivo functional experiments based on GF animal models can effectively eliminate the complex background interference of nonhuman microorganisms that is ubiquitous in traditional experimental animals. Furthermore, the colonization of GF animals with bacteria of interest makes it possible to directly verify the potential causal relationship between certain commensal bacteria and physiological phenotypes in a more precise way than that of studies on conventional animal models.
Over the past decade, the academia has seen a spike in the application of GF animal models such as mouse, drosophila, zebrafish, and nematode (Al-Asmakh and Zadjali, 2015; Berg et al, 2016; Jia et al, 2021; Lee et al, 2019; Li and Wei, 2019; Melancon et al, 2017; Petkau et al, 2017). Since mouse is the most widely used animal model to study the human pathological processes and test the corresponding therapeutic strategies, its GF counterpart naturally becomes an indispensable tool to investigate the intervention effect of commensal microbiome on disease and treatment methods. Functional experiments based on GF mice have proved that the gut microbiota influence the development and therapeutic outcomes of a variety of major human diseases, including cancers, metabolic diseases, and mental disorders (Liao et al, 2019; Zhao et al, 2020; Zheng et al, 2019).
To rederive GF mice, a common practice in the first step is performing hysterectomy and isolating the pup from its conventional mother's uterus under sterile condition (Fig. 1). The pup is then fed and raised in a sterile isolator with artificial breastfeeding or a GF foster mother (Arvidsson et al, 2012). Because the mothers of the first generation of pups are not GF, it is important to monitor if a pup is contaminated through the placenta. Once a GF mouse strain is established using the above method, the progeny can be produced on a large scale for experiments.
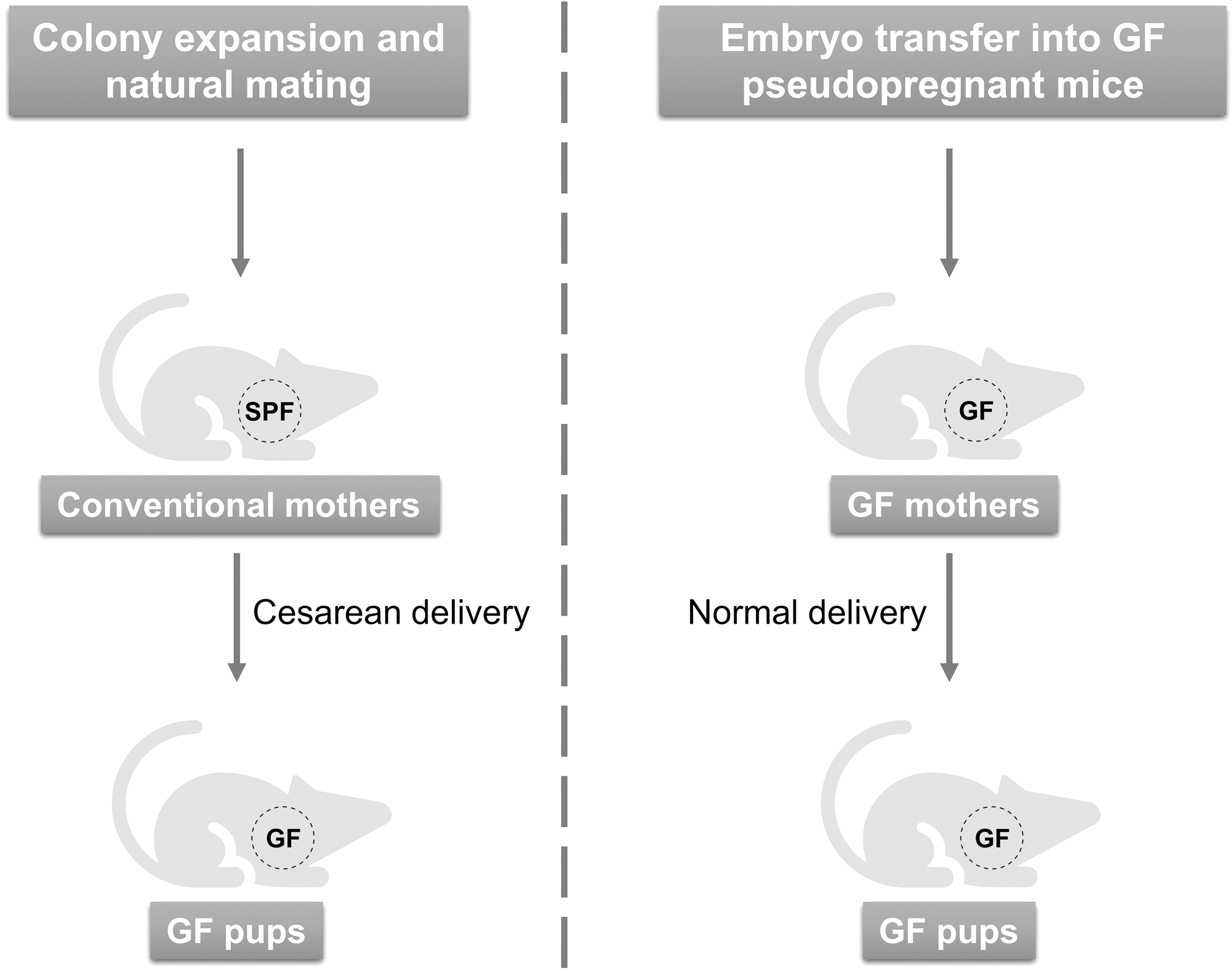
Certainly, in vitro fertilization (IVF) and embryo transfer under sterile conditions can also be used to rapidly establish a GF mouse colony provided that enough GF pseudopregnant recipients have been obtained in advance (Fig. 1) (Inzunza et al, 2005; Sztein et al, 2011). Of note, regular testing of bacteria, fungi, and viruses through serological testing, culture methods, and molecular biology techniques is essential to ensure the quality of GF mice. In addition to conventional strains, genetically modified mouse strains routinely used in cancer research are also worth rederiving as GF to underpin microbiome research.
Humanized Mouse Models in Immunotherapy Research
Due to species-specific differences, significant inconsistencies between in vivo experimental results based on conventional murine models and clinical outcomes are not uncommon in the context of cancer immunotherapy research (Fig. 2). This calls for predictive preclinical models that replicate the functional human immune system (HIS) and TME.
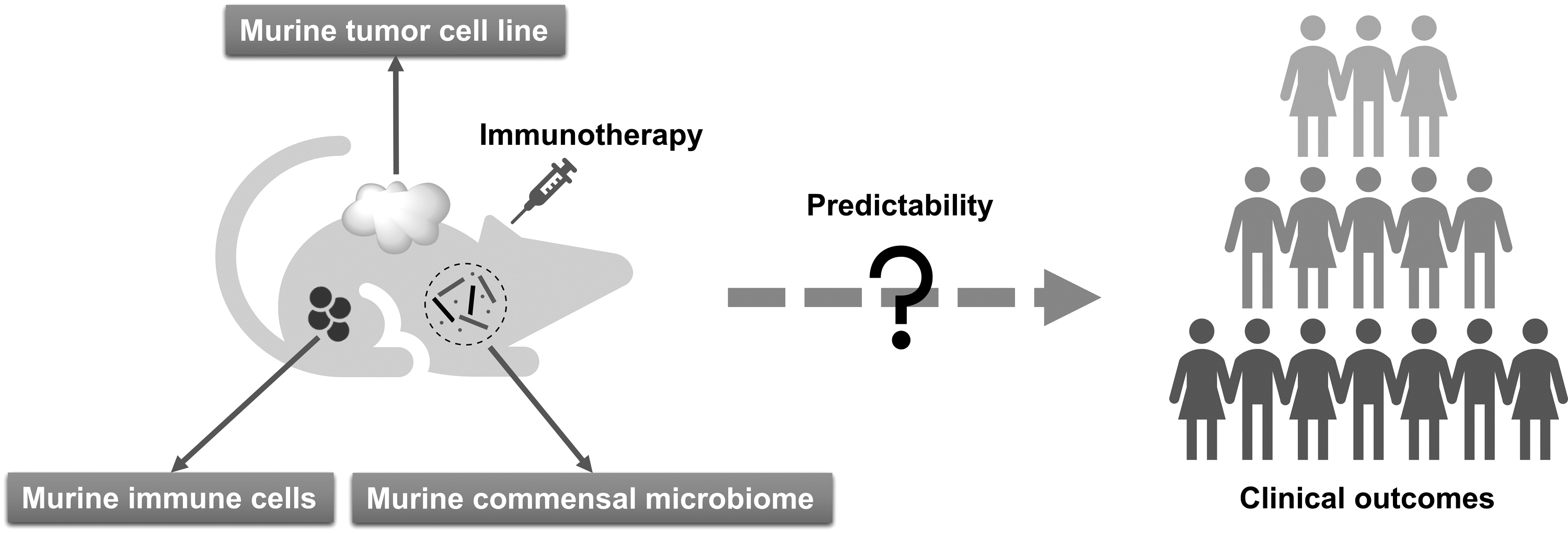
Accordingly, various mouse models xenotransplanted with human cells and/or expressing human gene products have been developed in recent years with the rise of cancer immunotherapy (Fig. 3) (Stripecke et al, 2020). As introduced in the Commensal Microbiome in Cancer Immunotherapy section, commensal microbiome is a key regulator of immunotherapy efficacy. Meanwhile, commensal microbiome has been demonstrated to impact host physiology and immunity (Clavel et al, 2017; McDermott and Huffnagle, 2014; Round and Palm, 2018). All of these facts imply that humanized mouse models harbor the potential to provide indispensable support for microbiome research in the field of immunotherapy.
The development of immunodeficient mice
To incorporate human immune cells into mice, the recipients should be preferably immunodeficient to facilitate the survival and function of the human graft. Since the discovery of nude mouse in the 1960s (Flanagan, 1966), this field has seen successive breakthroughs in the development of immunodeficient mice represented by scid, NOD-scid, NOG, NSG, BRGS, and B6RGS strains (Bosma et al, 1983; Ito et al, 2002; Legrand et al, 2011; Shultz et al, 2005; Shultz et al, 1995; Yamauchi et al, 2013), especially in the past 30 years.
Severely immunodeficient mice with defects in B, T, and natural killer (NK) cells are now being widely used (Shultz et al, 2019; Shultz et al, 2007). For example, based on the genetic background of NOD, the scid mutation in the Prkdc gene of the NOD-scid IL2Rgnull strains (e.g., NSG mice from The Jackson Laboratory) results in deficiencies in functional B and T cells.
Concurrently, the IL2Rgnull mutation blocks the corresponding cytokine signaling pathways, leading to loss of mature NK cells. Therefore, the NOD-scid IL2Rgnull mice are particularly suitable for reconstitution of functional human immunity. In recent years, many laboratories and companies around the world have developed their own NOD-scid IL2Rgnull strains to support the construction of humanized models for immunotherapy research.
Reconstitution of HIS in immunodeficient mice
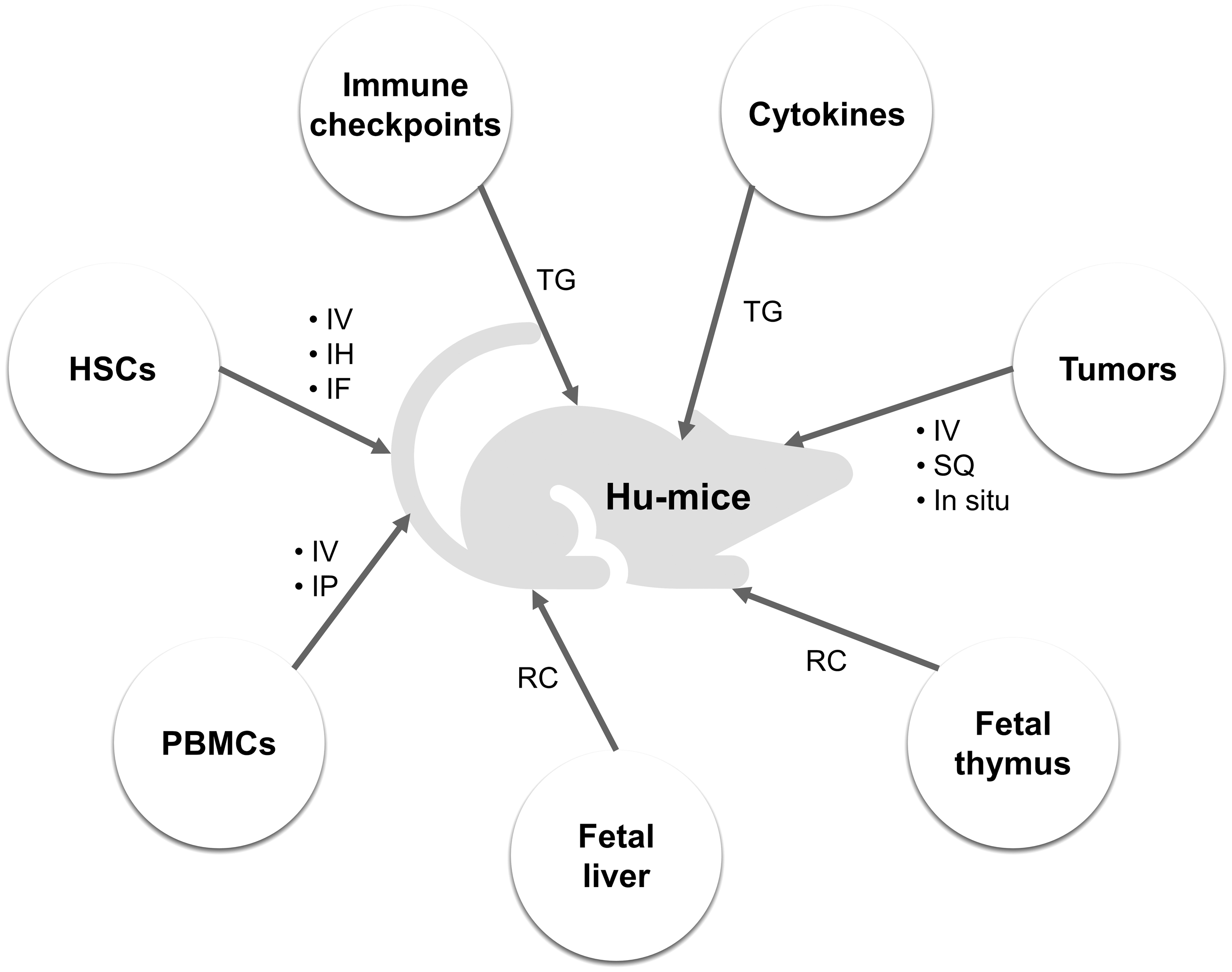
The reconstitution of HIS in immunodeficient mice can be realized through injection of human peripheral blood mononuclear cells (PBMCs). A distinct feature of PBMC-engrafted mice is the relatively short construction time, advancing preclinical immunotherapy research. However, the rapid development of graft-versus-host disease (GvHD) curtails the survival time of the models, hindering long-term studies (King et al, 2009).
Another approach to establish the functional HIS in immunodeficient mice is engraftment of CD34+ hematopoietic stem cells (HSCs) after irradiation. The HSC-engrafted mice support more diverse preclinical studies with optimized differentiation of human-derived immune cells although it takes months to achieve a successful reconstitution of HIS (Audige et al, 2017). The participation of cytokines in the development of immune system is crucial (Kondo et al, 2001). Due to species specificity, mouse cytokines limit the differentiation of human immune cells.
Human myeloid cell and NK cell development is particularly suppressed in the absence of compatible human cytokines. To improve the integrity of the reconstituted HIS in mice, derivatives of the abovementioned severely immunodeficient mice expressing human cytokines have been constructed. For example, NSG-SGM3 and NOG-EXL mice, both being capable of expressing human IL-3 and granulocyte-macrophage colony stimulating factor (GM-CSF), demonstrate enhanced development of human myeloid cells after transplantation of human HSCs (Ghosh et al, 2019; Jangalwe et al, 2016; Maser et al, 2020; Yoshimi et al, 2017). NSG-Tg(Hu-IL15) and NOG-Tg(Hu-IL15) mice, on the contrary, expressing human IL-15, can facilitate the development of human NK cells (Brehm et al, 2018; Katano et al, 2020; Katano et al, 2017; Yao et al, 2020).
Given that IL-6 is important in inducing the differentiation of B cells and the secretion of antibodies, human IL-6 gene knockin mice based on NOG, NOD, and BRGS strains were also generated to support studies on cancer immunotherapies targeting immunoregulatory myeloid cells (Hanazawa et al, 2018; Yu et al, 2017). In some cases, to develop a more robust HIS, another alternative is resorting to bone marrow-liver-thymus (BLT)-humanized mouse models. Just as the name implies, these models are transplanted with human HSCs, fetal liver, and thymus tissue (Brown et al, 2018; Lavender et al, 2014; McCune et al, 1988). BLT-humanized mice have shown great potential in predicting personalized immune response and adverse events of immunotherapy (Kalscheuer et al, 2012; Weaver et al, 2019; Yan et al, 2019).
Xenograft of human tumors based on immunodeficient mice
Another important aspect of constructing humanized mouse models is human tumor transplantation. For decades, the National Cancer Institute (NCI)-60 human cancer cell lines have been widely used around the world. After inoculating these human cancer cell lines into mice, the resulting cell line-derived xenograft (CDX) mouse models were established to support screening of numerous anticancer therapies.
Although the NCI-60 cell lines are human derived, they have been cultured for thousands of generations, which is adequate to substantially change their genetic background, leading to poor imitation of tumor characteristics of patients. Eventually, in 2016, the NCI decided to stop using the NCI-60 for anticancer drug screening and advocated developing patient-derived xenografts (PDXs) instead (Ledford, 2016). By implanting small fragments of a patient's tumor into an immunodeficient mouse, an environment that mimics the human body can be created.
A PDX model is capable of preserving tumor heterogeneity and TME to better predict the clinical outcomes of cancer treatment. The patient-derived tumor in the first generation of mice can be harvested and regrafted for the building of the next generation of PDX models. Consortium such as EurOPDX, The Jackson Laboratory, NCI's PDXNet, and Novartis have developed large PDX banks that aim at producing more translatable results from preclinical mouse models (Byrne et al, 2017; Evrard et al, 2020; Ledford, 2016; Risbridger et al, 2020). It is recommended that immunodeficient mice be coengrafted with human tumors and human stem cells to more accurately assess immunotherapeutic agents (Lin et al, 2018; Meraz et al, 2019; Zhao et al, 2018).
Humanized immune checkpoint mice
Considering the currently central status of ICB in cancer immunotherapy approaches and its expanding application scope, the need for appropriate humanized mouse models for preclinical evaluation of ICIs is growing since the recognition sites on checkpoint proteins for antibodies are different across the species. To address this problem and acquire more translatable preclinical results of human ICIs, humanized immune checkpoint mouse models have been developed. A common practice is replacing the extracellular binding sites of mouse immune checkpoints with human sequences using genetic engineering technologies while retaining the transmembrane and intracellular parts to ensure intact intracellular signal transduction (Barham et al, 2019; Choi et al, 2020).
Humanized immune checkpoint mice can be implanted with human cancer cell lines afterward, while PDXs are difficult to survive in these models as the mouse immune system is usually complete. To match the characteristics of multitarget antibodies, humanized mice expressing multiple human immune checkpoints simultaneously are also receiving increasing attention. Apart from the well-known negative molecules CTLA-4 and PD-1/PD-L1, more targets including both inhibitory (e.g., TIGHT, TIM-3, CD47, VISTA, and LAG-3) and costimulatory (e.g., GITR, OX40, 4-1BB, ICOS, CD27, CD28, and CD40) immune checkpoints are being investigated (Feng et al, 2019; He and Xu, 2020; Mayes et al, 2018). Thus, a wider variety of humanized immune checkpoint mice shall be developed to allow for assessing novel checkpoint-based immunotherapeutic approaches.
Considerations in construction and application of humanized mice for immunotherapy and microbiome research
Humanized mouse models have shown significant advantages over conventional models in imitating human physiology, pathology, and immune system, enabling in vivo anticancer research. Despite these advances, we found several limitations of the current humanized models. First, although delayed lethal GvHD and a longer life span can be achieved in HSC-engrafted or BLT humanized mice compared with PBMC-engrafted mice, the possible later onset of GvHD is unfavorable for long-term safety evaluation of therapeutics. Second, the lack of eligible source of human grafts such as stem cells and tumor tissues could have a significant impact on the reproducibility and translatability of preclinical research, especially increasing the studies' focus on individual medical needs.
Last but not least, mice transplanted with human fecal microbiota, known as humanized gnotobiotic mice, are widely used in commensal microbiome research. This calls for feasible and standardized protocols in preparing human fecal samples and the difference of basal microbiome between humans and mice must be taken into consideration.
Construction and Application of Humanized GF Mice
The current advances in the construction of GF mice and humanized models have laid the ground for explicit studies that investigate the causal relationship between commensal microbiome and cancer immunotherapy. Accordingly, we highly recommend integrating rederivation and humanization to generate humanized GF mouse models as more effective preclinical tools to facilitate studies in this field and develop novel clinical strategies.
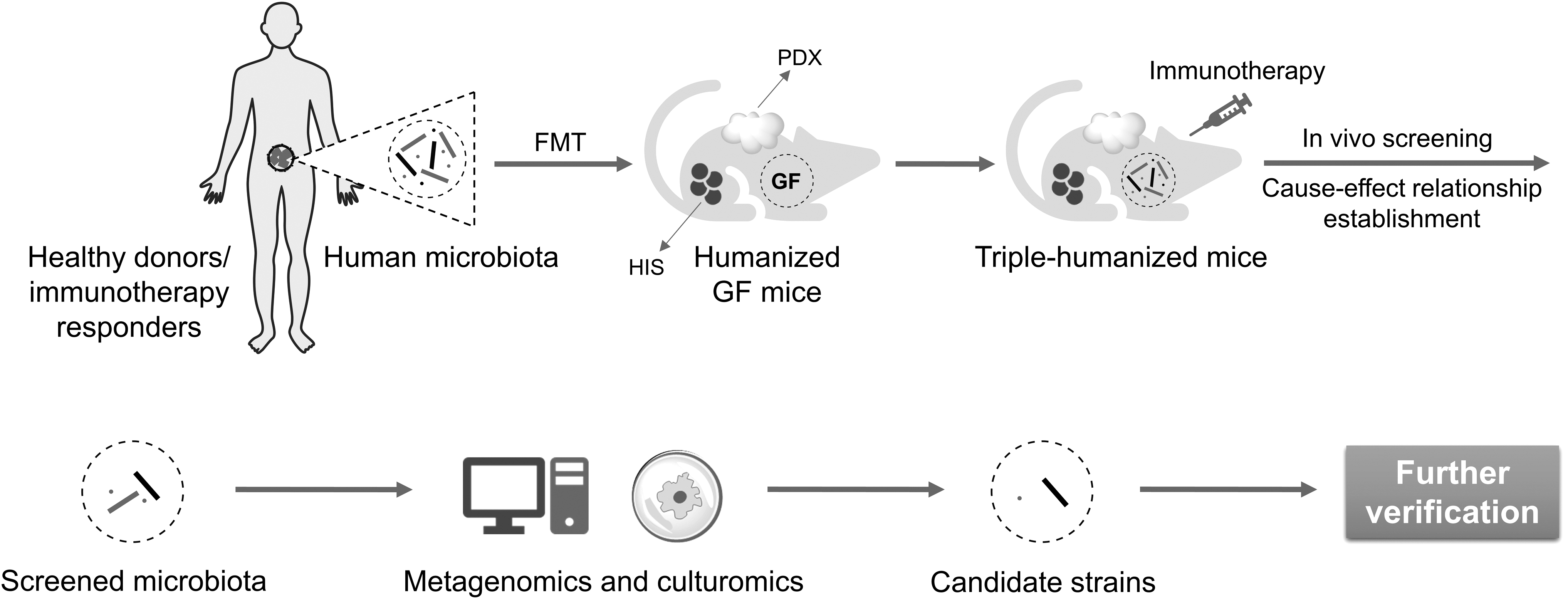
We have previously advocated the construction and application of humanized GF mouse models for tumor microbiome precision medicine (Fig. 4) (Wu et al, 2021a). First, a GF immunodeficient (e.g., NOD-scid IL2Rgnull ) mouse strain should be established. In general, the initial colony of GF immunodeficient mice is born through cesarean birth from the specific pathogen-free (SPF) mothers or embryo transfer into GF mothers. All the offspring are GF since birth and raised in sterile isolators. Next, patient-derived tumors and human PBMCs are transplanted into 4–7-week-old GF immunodeficient mice for humanization. After colonization of specific bacterial strains and administration of immunotherapy agents, the potential intervention effect of commensal microbiome on cancer immunotherapy can be evaluated.
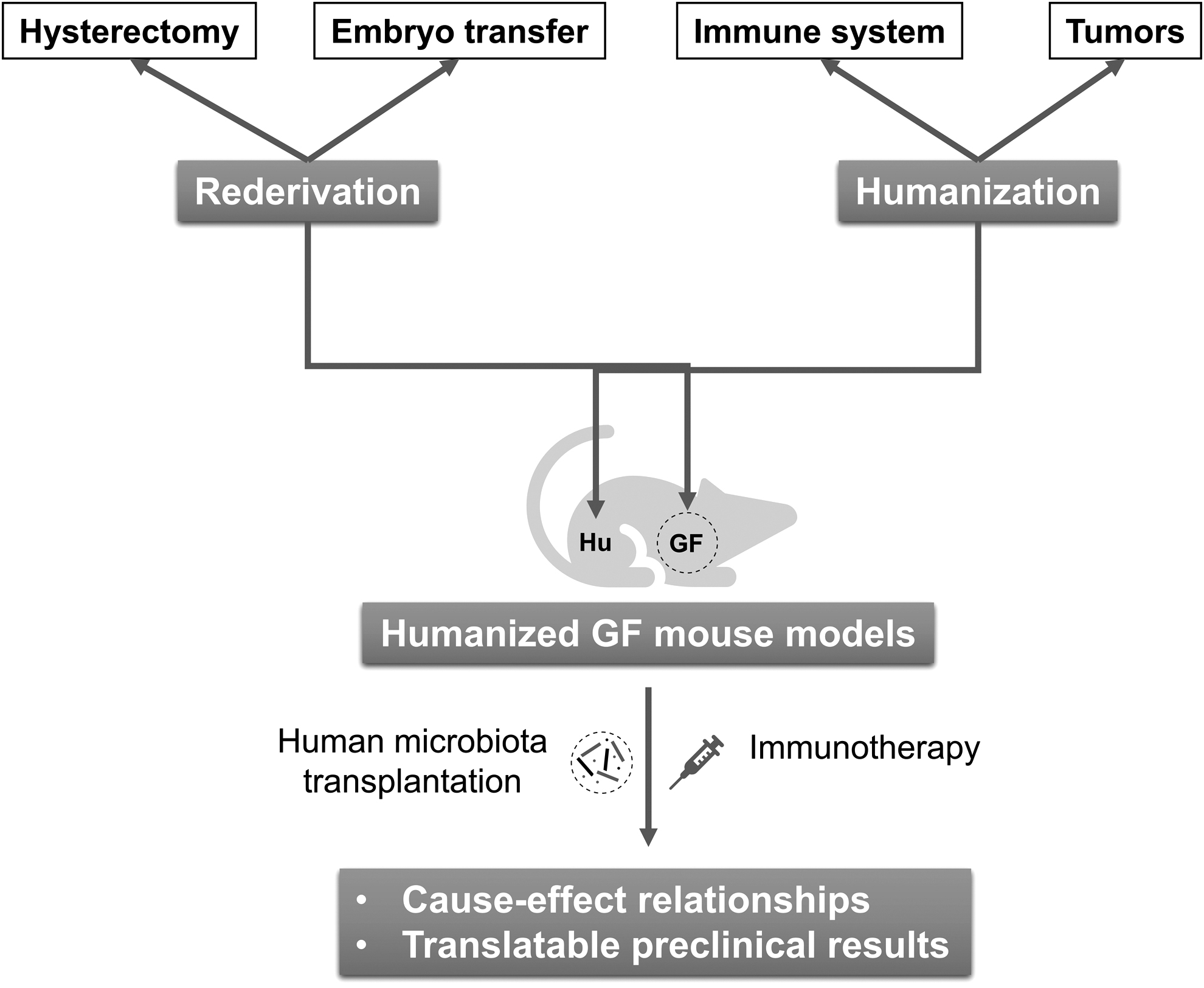
The order and timing of the above three humanization processes may vary according to the growth rate of tumors and colonization dynamics of bacteria. The key purpose is to ensure an appropriate therapeutic window for immunotherapy-associated commensal bacteria assessment. As discussed before, the life span of PBMC-humanized mice is limited by the quick onset of GvHD. If a longer time window is needed to evaluate the impact of microbiota on immunotherapy, HSC engraftment is also applicable in this model except that the construction time will increase dramatically.
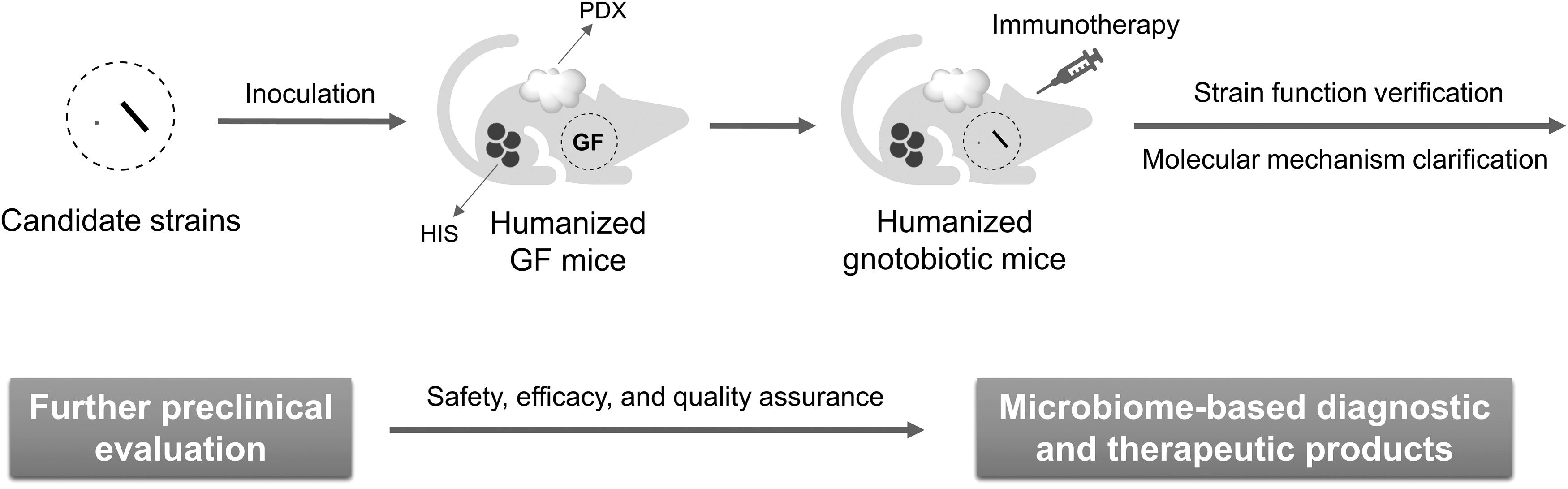
The above humanized GF mice are ideal tools for precise screening and evaluation of immunotherapy-related functional bacteria. By transplanting commensal microbiota from responders to immunotherapy or healthy donors into humanized GF mice, the cause–effect relationships could be established. Reasonably, negative regulators can also be found from the microbiota of nonresponders in the same way. Functional bacteria can be further obtained with the assistance of metagenomics and culturomics (Fig. 5) (Bilen et al, 2018; Kostrzewa et al, 2019; Lagier et al, 2018; Lagier et al, 2016; Martellacci et al, 2019). In turn, through inoculation of humanized GF mice with a candidate strain or a defined consortium of strains, one is capable of verifying the immunomodulatory function and clarifying the underlying molecular mechanism (Fig. 6).
Conclusion
Enormous efforts to explore the intervention factors of cancer immunotherapy have been made from various directions, in which commensal microbiome has emerged as a key regulator. Nevertheless, many association studies that tried to link microbiome and immunotherapy outcome based on superficial sequencing and unreliable murine models are far from sufficient to demonstrate causality and identify specific microbial strains as putative biomarkers or therapeutic targets (Walter et al, 2020; Wu et al, 2021b).
To verify the potential role of bacteria in modulating cancer immunotherapy, a prerequisite is bacteria culture, of which the feasibility has been greatly improved owing to recent advances in culturomics (Bilen et al, 2018; Kostrzewa et al, 2019; Tidjani Alou et al, 2020). Subsequently, dependable murine models shall play a vital role in elucidating the underlying mechanism and establishing the causal relationship between specific commensal bacteria and immunotherapy efficacy.
After decades of development, immunodeficient mice have been demonstrated to be powerful tools for reconstitution of HIS and implantation of human tumors, which are critical to better mimic the human immunotherapy environment (Lin et al, 2018; Meraz et al, 2019; Zhao et al, 2018). GF mice, on the contrary, have shown indispensable value in the context of human microbiome research in immunotherapy because of the blank microbial background and tightly controlled experimental environment (Huang et al, 2022; Matson et al, 2018; Tanoue et al, 2019).
Despite the fact that the number of new strains of both immunodeficient and GF mice is on the rise, respectively, the development of humanized GF mouse models is still in its infancy and their application in microbiome research can barely be seen in published articles. Given the increasing maturity of technologies in laboratory animal science and the urgent demand for translational research in cancer immunotherapy, now is the right time to positively develop humanized GF mouse models to facilitate precision research in clarifying the intervention effect of human microbiome on cancer immunotherapy.
Currently, we are focusing on optimizing the construction strategy of HIS-engrafted GF mouse models to elevate the throughput of in vivo experiments. Of note, we should fully realize there exist potential limitations in the humanized GF mice. One consideration is that the current humanization level should be further improved to meet the growing needs of precision medicine in immunotherapy. Besides, the breeding performance of some GF strains could be worse than their SPF counterpart, limiting the throughput of one experimental cohort. Furthermore, the characteristics of microbiota colonization between humanized GF mice and humans could sometimes be hugely divergent because of host-specific influences, which would cast doubt on the conclusions of corresponding studies.
Looking to the future, we anticipate more tangible evidences illustrating that the molecular mechanism of commensal microbiome influencing immunotherapy can be discovered by the agency of humanized GF mouse models. Moreover, the potential effect of a specific microbial strain or consortium, whether it plays a positive or negative role in cancer immunotherapy, can be better verified through in vivo experiments in humanized GF mice. Preclinical animal models are evolving and so are cancer treatments. The ultimate goal in this field is that novel diagnostic and therapeutic products drawn from commensal microbiome research based on advanced animal models will be developed and implemented into practice, benefiting the clinical outcome of cancer immunotherapy.
Footnotes
Authors' Contributions
X.W. and C.W. wrote the article and designed the figures. H.W. conceived the review and revised the article. All authors approved the final version of the article to be published.
Author Disclosure Statement
The authors have no conflict of interest.
Funding Information
No funding was received for this article.