
Abstract
Aims:
Radiation-induced sensorineural hearing loss (RISNHL) is one of the major side effects of radiotherapy for head and neck cancers. At present, no effective clinical treatment or prevention is available for RISNHL. This study thus aimed to investigate the cochlear pathology so that the underlying mechanisms of RISNHL may be elucidated, consequently paving the way for potential protective strategies to be developed.
Results:
Functional and morphological impairment in the stria vascularis (SV) was observed after irradiation (IR), as indicated by endocochlear potential (EP) reduction, hyperpermeability, and SV atrophy. The expression of zonulae occludins-1 was found to have decreased after IR. The loss of outer hair cells (OHCs) occurred later than SV damage. The disruption to the SV and OHCs could be attributed to reactive oxygen species (ROS)–related damage. In addition, EP shifts and the loss of OHCs were reduced when ROS was reduced by N-acetylcysteine (NAC) in C57BL/6 mice, attenuating auditory threshold shifts.
Innovation:
The damage to the SV was found to occur before OHC loss. ROS-related damage accounted for SV damage and OHC loss. The incidences of SV damage and OHC loss were decreased through ROS modulation by NAC, subsequently preventing RISNHL, suggesting the possible role of NAC as a possible protective agent against RISNHL.
Conclusion:
The findings from this study suggest oxidative stress–induced early SV injury and late OHC loss to be the key factors leading to RISNHL. NAC prevents IR-induced OHC loss, and attenuates auditory brainstem response and EP shifts by regulating the level of oxidative stress. Antioxid. Redox Signal. 40, 470–491.
Introduction
Radiotherapy (RT) is mainly used for treating head and neck cancers (Strojan et al., 2017), especially nasopharyngeal carcinoma (De Ruysscher et al., 2019). Although maximum tumor control is hoped to be achieved with minimal risk of damage to normal tissues as RT technology advances, it is sometimes unavoidable. Unfortunately, patients who receive RT are at a 40%–50% risk of developing sensorineural hearing loss (SNHL), which is known as radiation-induced sensorineural hearing loss (RISNHL) when it occurs in this context (Gupta et al., 2020; Tan et al., 2013). RISNHL is irremediable (Gupta et al., 2020) and may progress to complete deafness (Bhandare et al., 2010). Therefore, not only does RISNHL pose a communication barrier to the patients, it also greatly affects the quality of life of a patient (Strojan et al., 2017). At present, no safe and effective treatment or prevention strategy is available for RISNHL in clinical practice. There is thus a pressing need for the mechanisms of radiation-associated ototoxicity to be elucidated, so that effective strategies against RISNHL can be subsequently developed.
Researchers face a huge challenge in acquiring cochlear samples from RISNHL patients; the target for such studies is therefore shifted onto the establishment of an appropriate animal model. Previous studies focused on hair cell (HC) death in the organ of Corti, which was thought to be the key reason for RISNHL (Tan et al., 2013). However, hearing loss may be owing to a combination of damage in the stria vascularis (SV), loss of HCs or neuronal demyelination (Wang and Tepper, 2021). Henceforth, this study aims to clarify the differences in terms of sensitivity to irradiation (IR) between different cochlear structures by observing pathological changes after RT, and to analyze the patterns of major pathological changes that accompany SNHL after IR.
Innovation
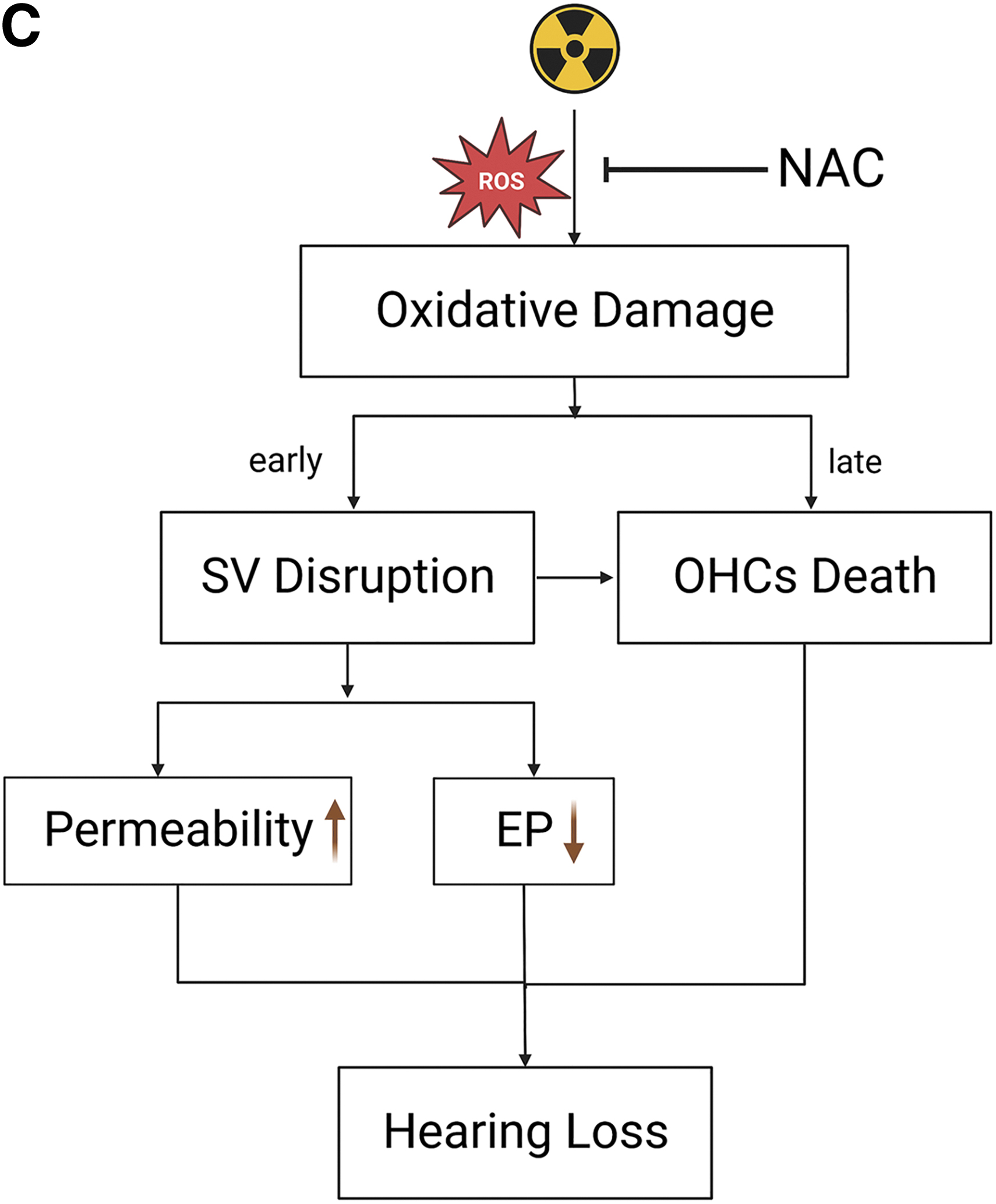
The damage to the stria vascularis (SV) was found to have occurred before hair cell loss, contrasting some findings from previous reports. Damage to the SV was associated with early hearing loss after irradiation. In this case, reactive oxygen species (ROS)–related damage was identified to account for the SV damage and outer hair cell (OHC) loss. Therefore, the modulation of ROS by N-acetylcysteine (NAC) reduces SV damage and OHC loss, consequently prevents radiation-induced sensorineural hearing loss (RISNHL) and sheds light on the possible role of NAC as a protective agent against RISNHL.
IR classically kills cells in a nonselective manner by damaging DNA and cellular components in the tumor and bystander cells via cytotoxic reactive oxygen species (ROS) generated from water molecules or direct ionization (Bruckbauer and Cox, 2021; Dou et al., 2018). ROS is also thought to play a crucial role in HC death in both noise- and drug-induced hearing loss (Pyun et al., 2011). As IR exposure leads to increased production of intracellular ROS, hearing loss after RT is therefore speculated to be related to oxidative stress damage.
The antioxidant N-acetylcysteine (NAC) was found to reduce ROS levels and minimize oxidative stress induced by excessive ROS production (Lu et al., 2014). An in vitro study showed a significant increase in cell viability and decrease in ROS levels after irradiated mouse auditory cell line HEI-OC1 was treated with NAC (Duzenli et al., 2019). Although NAC has not been tested in any in vivo experiments after IR yet, this study deduced the possible roles of NAC in delaying RISNHL progression via oxidative stress regulation based on the pathological damage to the inner ear after IR. This suggests that NAC, or drugs that regulate the level of oxidative stress, may have the potential to become a clinical radioprotective agent.
The study of the mechanism of RISNHL along with the development of prevention strategies have therefore become increasingly important as RISNHL is one of the most common complications of RT for head and neck cancer. This study aims to investigate the functional and pathological characteristics of cochlear subregions after IR using an animal model, and simultaneously elucidates the relationship between RISNHL and oxidative stress damage. We were also interested in finding out if NAC can protect a patient against RISNHL in vivo by modulating the level of oxidative stress (Fig. 1).
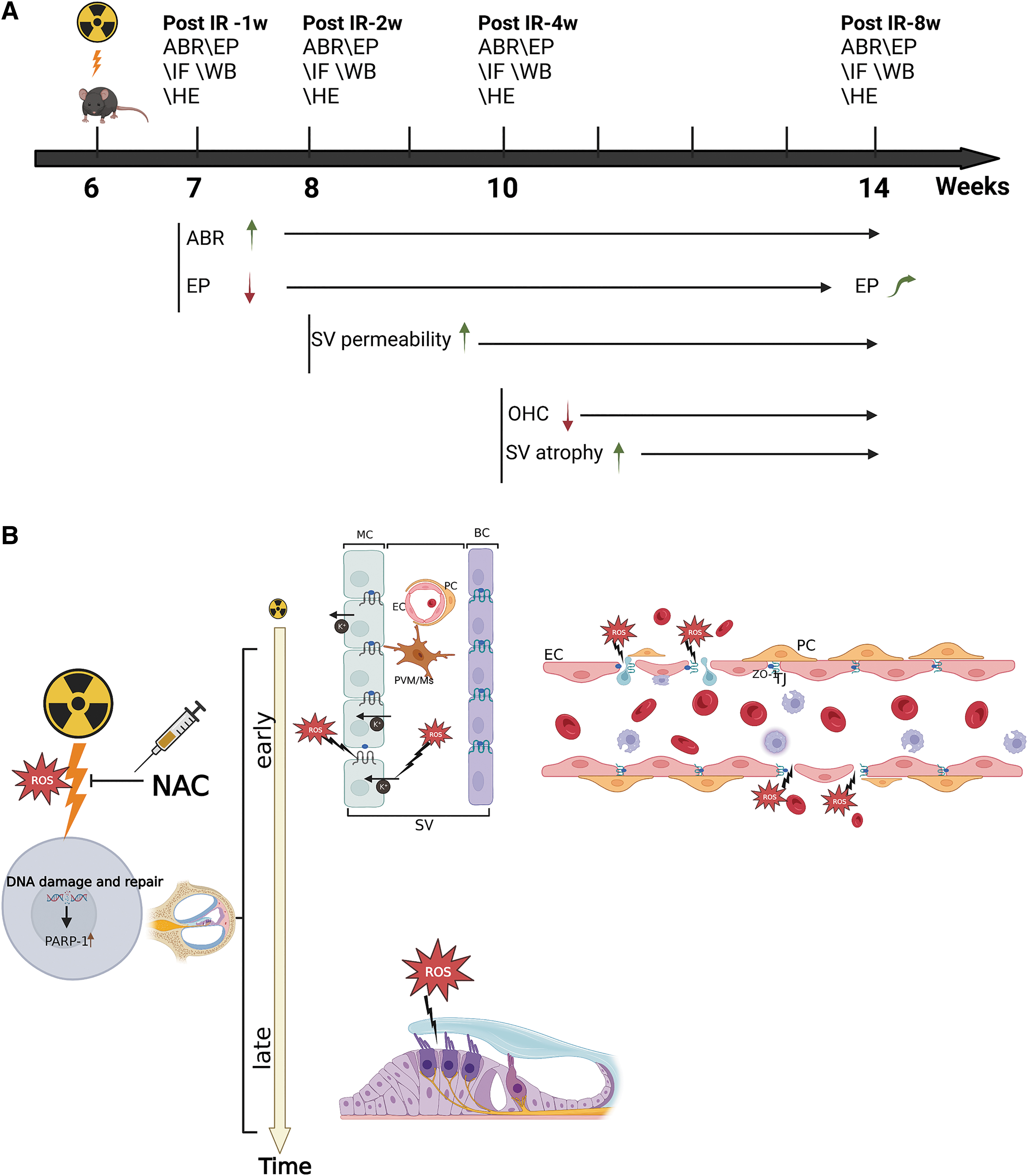
Results
Time-dependent progressive hearing threshold changes after IR
Six-week-old mice were exposed to RT. These mice were killed for surface preparations for Western blot (WB), immunofluorescence, and hematoxylin–eosin (HE) staining after final auditory brainstem response (ABR) and endocochlear potential (EP) measurements to evaluate the injury levels at 1, 2, 4, and 8 weeks after IR exposure (corresponding to 7, 8, 10, and 14 weeks of age, respectively). The timeline for the C57BL/6 mouse experiment is given in Figure 2A. A C57BL/6 mouse model was established, with a radiation dose of 15 Gy being delivered to both ears in the IR group (Fig. 2C). The mice in the control group received no treatment. This radiation dose was selected based on literature review (Liu et al., 2022) and preliminary experiments, and no IR-induced death occurred in mice at this dose.
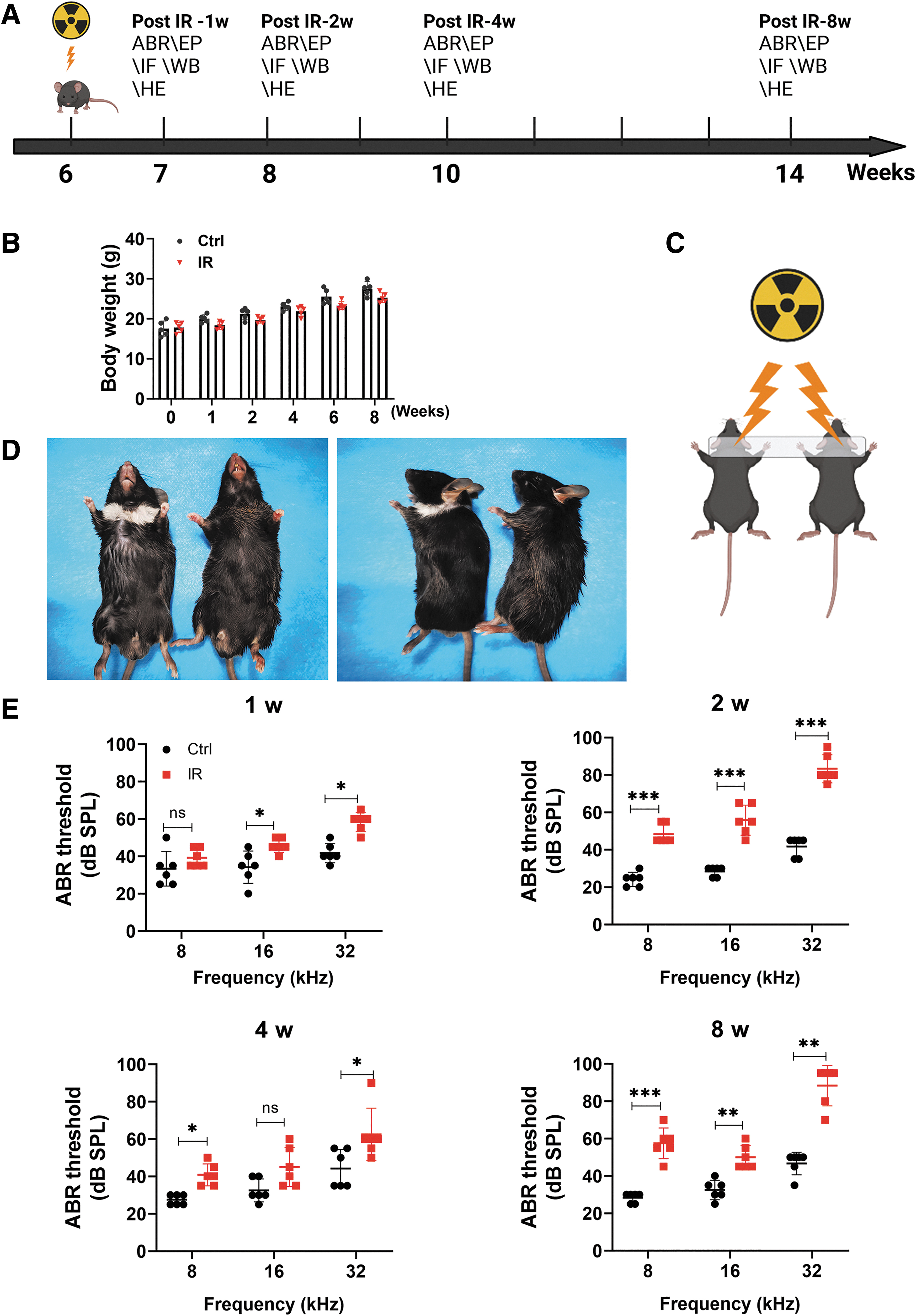
Changes in body weight and fur condition were monitored each week for 8 weeks. There was no significant difference in body weight between mice in the control and IR groups (Fig. 2B). In the IR group, skin damage occurred at 2 weeks after radiation exposure, and self-healing wounds with scabs occurred at 1–2 weeks after IR without intervention. The local skin area was rough, thickened, and hardened after healing, whereas the regenerated hair was white in color (Fig. 2D; IR group on the left and control group on the right). It is possible that this occurrence was related to the hair follicle, sweat gland, and sebaceous gland being damaged (Rosenthal et al., 2019).
The ABR is an objective electrophysiological test of the hearing function. The ABR was used to record the progression of RISNHL. At 1 week, the ABR thresholds of mice at 16 kHz (45.83 ± 3.764 dB) and 32 kHz (58.33 ± 5.164 dB) in the IR group were significantly higher than those in the control group. At 2 weeks, IR was found to cause more significant elevations in ABR thresholds at 8 kHz (48.33 ± 5.164 dB), 16 kHz (55.83 ± 8.010 dB), and 32 kHz (83.33 ± 7.528 dB). The average thresholds obtained for mice in the control group at 8, 16, and 32 kHz were significantly lower than those obtained by the radiation-exposed mice.
At 4 weeks, the ABR thresholds were significantly increased at 8 kHz (40.33 ± 5.845 dB), 16 kHz (45.00 ± 10.488 dB), and 32 kHz (62.50 ± 14.053 dB) for mice in the IR group, with those at 8 and 32 kHz being significantly higher than those in the control group. The increase in the ABR threshold at 2 weeks was even more significant than that at 4 weeks; however, this effect was temporary, likely indicating acute injury. The specific reasons for this effect need to be further studied. At 8 weeks, the ABR threshold was found to gradually increase over time; with the ABR thresholds being significantly increased at 8 kHz (57.50 ± 8.216 dB), 16 kHz (50.00 ± 6.325 dB), and 32 kHz (88.33 ± 10.801 dB). The average ABR thresholds obtained for the 8-week IR group at 8, 16, and 32 kHz were significantly higher than those obtained for the control group (Fig. 2E).
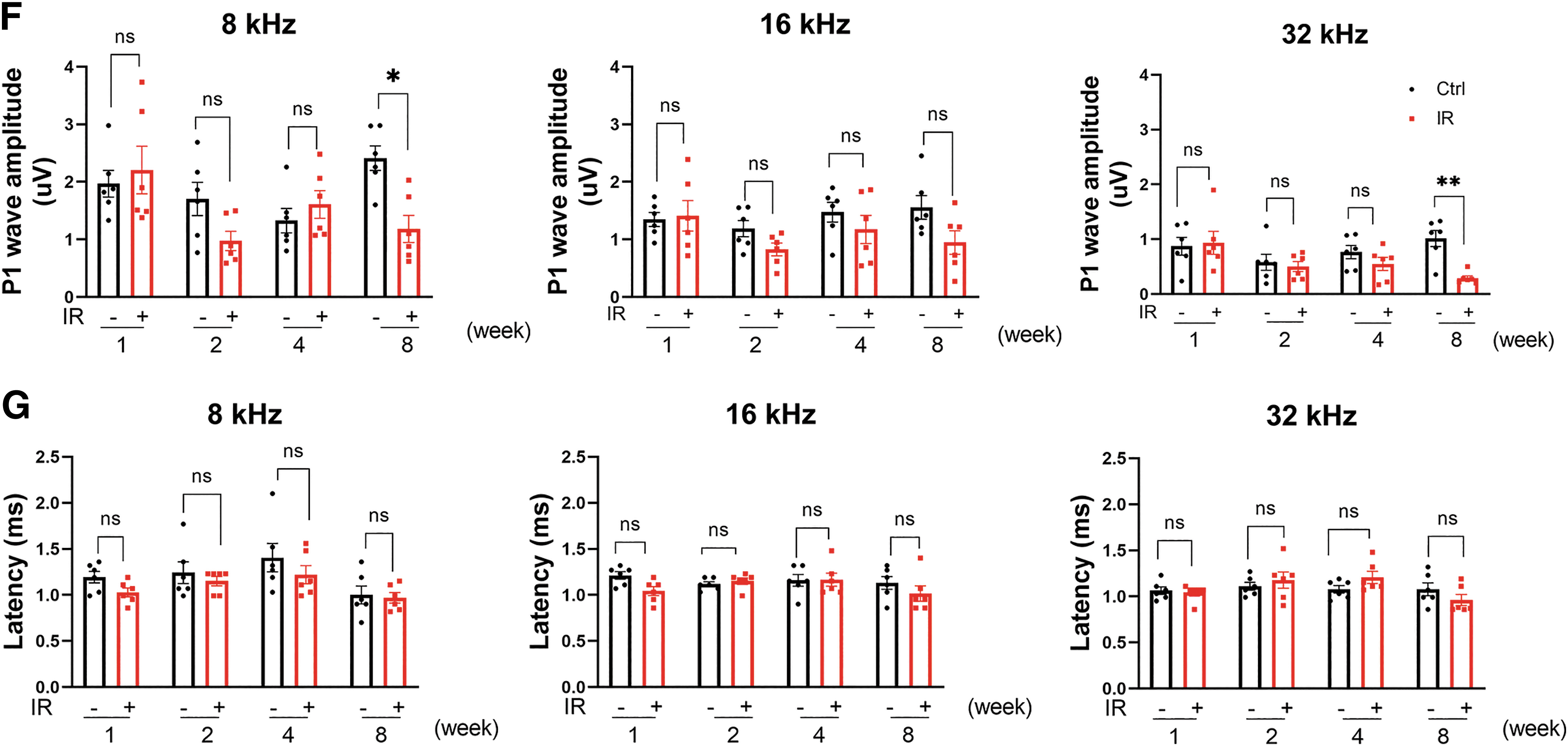
Further explorations were carried out to determine the impact of IR on hearing function at the suprathreshold level. Changes in the amplitudes and latencies of the P1 wave were analyzed. For P1 wave amplitudes, statistically significant differences were not observed until 8 weeks after IR, whereas the IR group yielded significantly reduced amplitudes at 8 and 32 kHz compared with the control group at 8 weeks. Finally, no significant differences were found in the latencies of the P1 wave between the IR group and the control group (Fig. 2F, G).
Outer HC loss after IR
To assess whether the functional deficits after IR correspond to the amount and localization of HC loss, cochlear HCs were evaluated using a surface preparation. Myosin-VIIa (red) staining showed three rows of outer hair cell (OHC) and one row of inner hair cell (IHCs) arrays in all turns (Fig. 3A). No HC loss was observed in the control group. There was no apparent IHC loss from the apical to basal region in the IR-treated mice. In contrast to the ABR results, there was no apparent OHC loss or IHC loss at 1 and 2 weeks after IR. The loss of OHCs was significantly increased only from 4 to 8 weeks after IR. By 4 weeks, there was <10% loss of OHCs, 1.714% ± 0.543% in the apical turn, 2.571% ± 0.543% in the middle turn, and 6.857% ± 2.347% in the basal turn. At 8 weeks, there was 1.714% ± 0.797% loss of OHCs in the apical turn, 2.952% ± 1.321% in the middle turn, and 8.00% ± 1.730% in the basal turn (Fig. 3B, C).
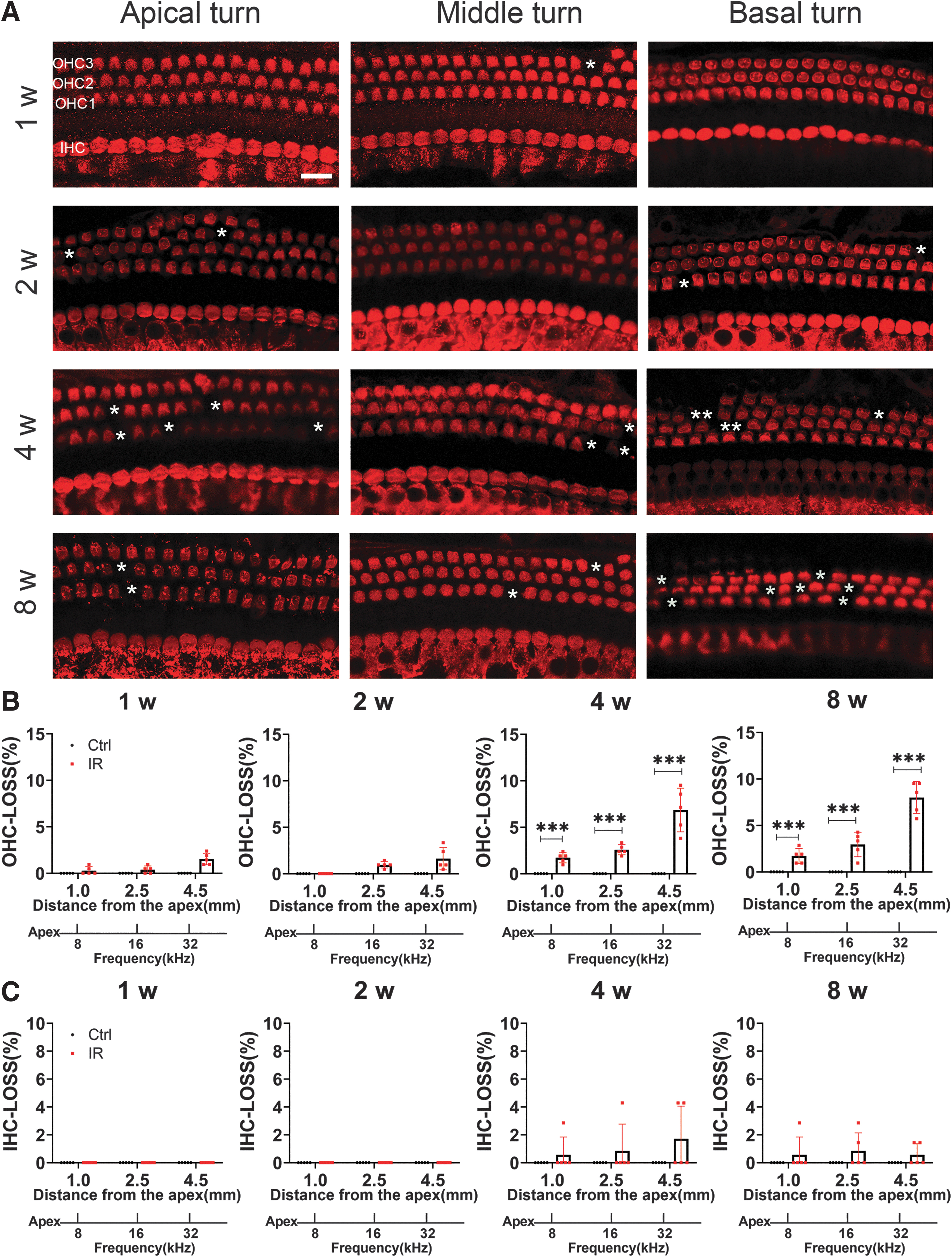
Synaptic ribbons were not lost
Cochlear ribbon synapses are known to connect IHCs with auditory nerve fibers as the main targets of external insults. In this study, immunolabeling was performed to stain the C-terminal binding protein 2 (CtBP2) (red) as a marker of presynaptic ribbons, GluA2 protein (green) as a postsynaptic marker, and myosin-VIIA protein (blue) as a marker of IHCs 1–8 weeks after IR. To visualize the binding of pre- and postsynaptic ribbons, the y–z axis was added. No obvious dissociation of the pre- and postsynaptic ribbons immunolabeled with CtBP2 and GluA2 was found after IR. In the control group, the mean number of binding synaptic ribbons was 12.25 ± 2.217/IHC in the apical turn, 15.75 ± 1.258/IHC in the middle turn, and 13.75 ± 0.50/IHC in the basal turn.
At 1 week after IR, the mean number of binding synaptic ribbons was 11.25 ± 1.5/IHC in the apical turn, 15.25 ± 2.22/IHC in the middle turn, and 13.25 ± 1.71/IHC in the basal turn. At 2 weeks after IR, the mean number of binding synaptic ribbons was 11.75 ± 1.7/IHC in the apical turn, 14.75 ± 3.59/IHC in the middle turn, and 11.75 ± 1.71/IHC in the basal turn. At 4 weeks after IR, the mean number of binding synaptic ribbons was 11.75 ± 0.96/IHC in the apical turn, 14.25 ± 2.22/IHC in the middle turn, and 11.250 ± 1.50/IHC in the basal turn. At 8 weeks after IR, the mean number of binding synaptic ribbons was 11.25 ± 0.96/IHC in the apical turn, 15.0 ± 2.45/IHC in the middle turn, and 12.75 ± 0.957/IHC in the basal turn. There was no apparent inner binding synaptic ribbon loss from the apical to basal region when comparing between mice from the IR group and the control group (Fig. 4).

Functional and morphological impairments induced by IR in the SV
The sensitive function of hearing is highly dependent on the integrity of SV. The HCs are immersed in the endolymph fluid. The EP and the high K+ concentration (∼150 mM) provided by the endolymph are generated and maintained by the SV. Thus, the integrity of the blood–labyrinth barrier (BLB) is critical for maintaining inner ear homeostasis and the EP, which is an important driver of auditory function. Normal EP values range from 80 to 120 mV. There were significant changes in the EP between mice in the control group (104.15 ± 2.51 mV) and mice in the 1-week (58.37 ± 24.59 mV), 2-week (59.69 ± 8.37 mV), and 4-week (57.95 ± 22.52 mV) IR-treated groups.
This result suggests the reduction of EP after IR at an early stage. EP decreased significantly at 1–4 weeks compared with that in the control group, indicating the impairment of SV function after IR. However, EP returned to and was maintained at nearly normal levels at 8 weeks after IR (Fig. 5A).
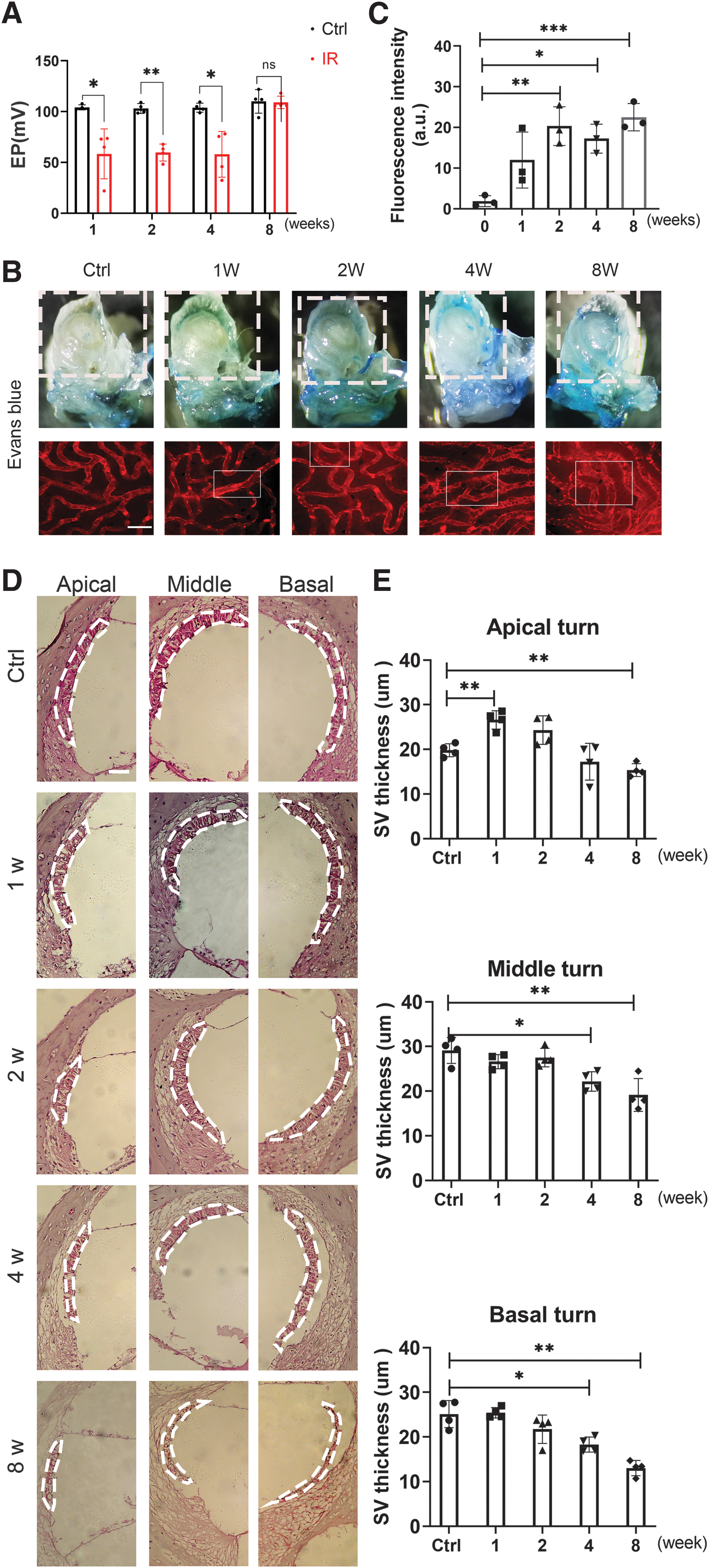
Evans blue (EB; MW: 961 Da) is commonly used to test the permeability of the blood–brain barrier and is useful in checking the permeability of the SV in the inner ear. Permeability was measured by quantifying EB extravasation and observing EB dye staining. EB has a molecular weight similar to that of plasma albumin and has a high affinity for plasma albumin in the blood. When the permeability of blood vessels increases, the albumin bound to EB leaks out, and the amount of EB exudation can be measured to reflect the degree of permeability. The fluorescence wavelength has a strong peak at 470 and 540 nm and a weak peak at 680 nm. EB dye was extravasated in the lateral cochlear wall of the IR group at a greater level than that in the control group.
We observed the leakage of EB from the microvessels in the SV with an excitation wavelength of 540 nm and quantified the fluorescence intensity. The fluorescence leakage of EB from the SV was enhanced in the IR group compared with that in the control group at 2–8 weeks. The results of the permeability assay using fluorescent tracer EB are given in Figure 5B, C. In addition, HE staining of the cochlear lateral walls showed atrophy of SV in the middle and basal turns at 4 weeks, whereas significant decreases in thickness of SV at the apical, middle, and basal turns were observed at 8 weeks. The thickness of SV only increased in the apical turn at 1 week after IR treatment (Fig. 5D, E).
A summary of the functional and morphological changes is given in Figure 1A. In week 1, the increase in ABR threshold was accompanied by a decrease in EP after IR. After week 2, the IR-treated mice demonstrated higher SV permeability than those in the control group. OHC loss and SV atrophy occurred 4 weeks after IR and became more significant by 8 weeks, whereas EP recovered at week 8.
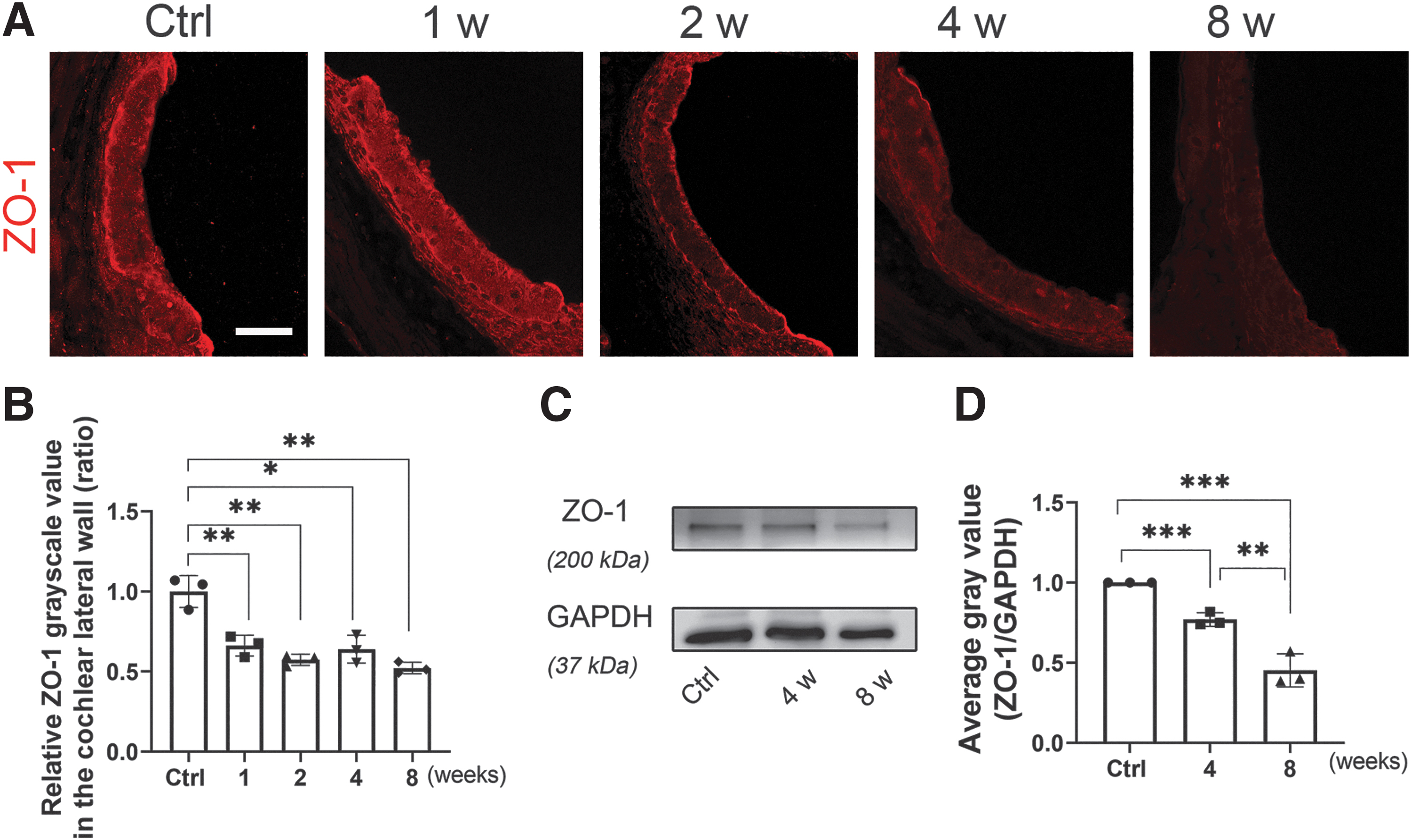
Decreased zonulae occludins-1 expression was associated with IR-induced damage to the tight junction networks of the SV
The barrier function of tight junctions (TJs) in the BLB is essential for sensory transmission. The protein zonulae occludins-1 (ZO-1), which is associated with TJs, was found in the SV (Gow et al., 2004). In this study, ZO-1 (red) immunolabeling was observed in the lateral wall, especially in the SV, 1–8 weeks after IR, and specimens from the IR group demonstrated weaker staining than the control group. Quantification of ZO-1 immunolabeling in the SV showed significant time-dependent decreases. Immunoblotting with a ZO-1 antibody revealed that the ratio of the ZO-1 band density to that of glyceraldehyde-3-phosphate dehydrogenase (GAPDH) was significantly different between mice from the control group and the IR-treated group. The IR group displayed reduced fluorescence intensity of ZO-1, consistent with the protein amounts verified by WB analysis (Fig. 6).
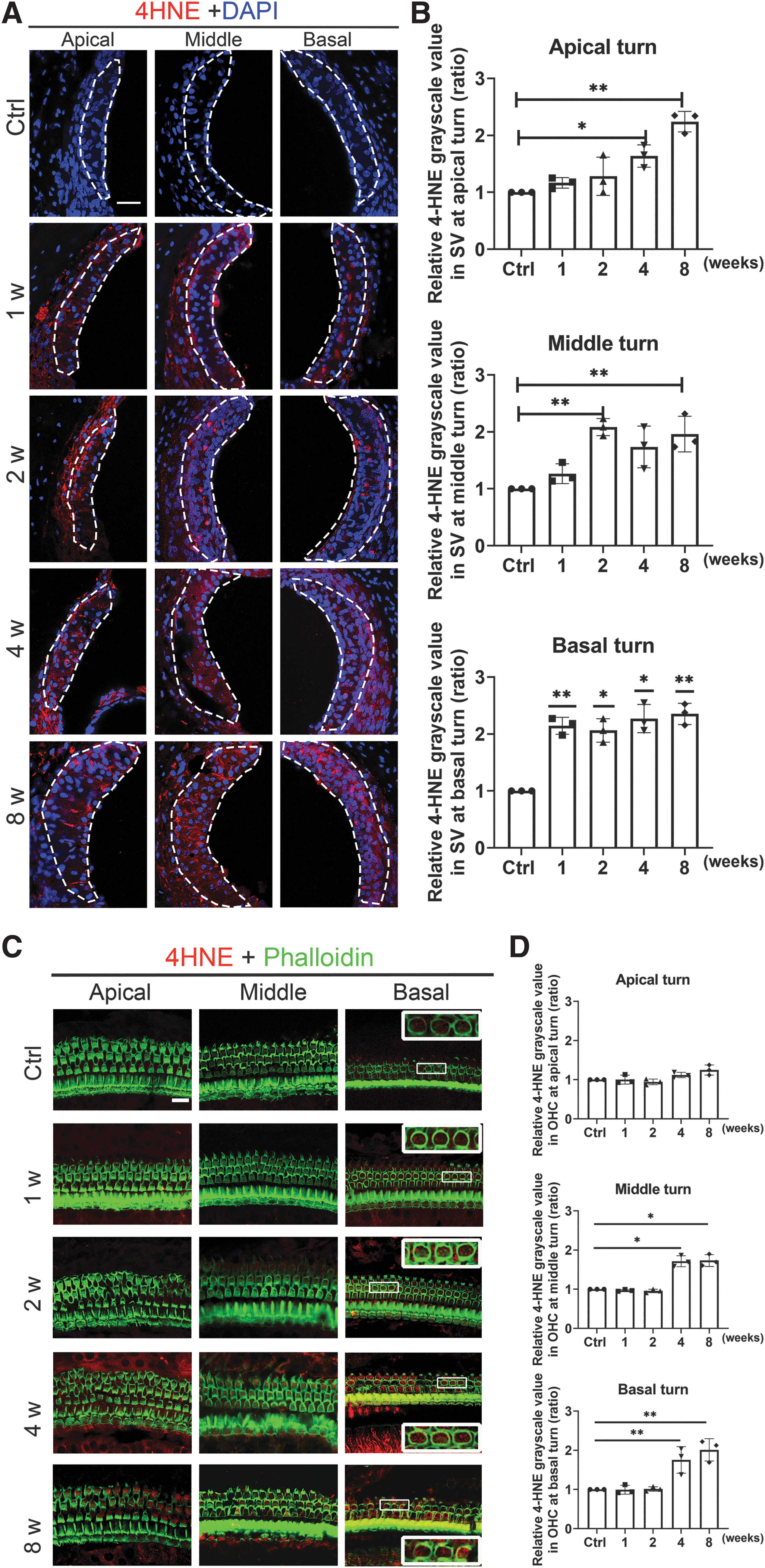
Increased 4-hydroxynonenal expression is associated with IR-induced ROS damage to the cochlear lateral wall and OHCs
Increased oxidative stress, as defined by enhanced 4-hydroxynonenal (4-HNE) levels, has been well documented after noise exposure. The intensity of 4-HNE was hence measured to evaluate the oxidative stress level in the cochlea. Immunolabeling of 4-HNE (red) combined with DAPI staining (blue) was noticed to have increased in the cochlear lateral wall, especially the SV, at 1–8 weeks after IR, with the IR-treated group exhibiting stronger fluorescence than the control group. Quantification of 4-HNE immunolabeling in the SV showed time-dependent enhancement. Immunostaining for 4-HNE did not appear in OHCs until 4 weeks after IR. At week 4, 4-HNE levels were more significantly enhanced at the apical and basal turns in the OHCs. Meanwhile, the 4-HNE levels became more significantly enhanced at the apical, middle, and basal turns in week 8. Quantification of 4-HNE immunolabeling in the OHCs also showed time-dependent enhancement, peaking at week 8 in all turns (Fig. 7).
DNA damage in the cochlear SV and OHC after IR
Poly (ADP-ribose) polymerase-1 (PARP-1) is activated in response to DNA damage and repair induced by oxidative stress and inflammatory damage (Cai et al., 2019; Salech et al., 2020). Immunoblots labeled with PARP-1 antibody revealed a significant increase in the PARP-1 to GAPDH band density ratio in the IR-treated group from week 2 to 8. Immunolabeling of PARP-1 (red) combined with DAPI staining (blue) was observed in the cochlear lateral wall, especially the SV, from week 2 to 8 after IR, with the IR-treated groups showing stronger staining output than the control group in the basal turn. Immunolabeling of PARP-1 (red) was observed alongside phalloidin (green) staining in the OHCs after IR, with the IR-treated groups yielding stronger staining than the control group from week 4 to 8. Quantification of PARP-1 immunolabeling showed time-dependent enhancement in both the SV and the OHCs (Fig. 8).

Modulation of ROS by NAC prevents IR-induced loss of OHCs and attenuates ABR and EP shifts
NAC is a naturally occurring compound that has been described as safe for humans (Jaudoin et al., 2021). The protective effect of NAC on HEI-OC1 cells via suppression of the ROS pathway has been previously studied in RISNHL (Duzenli et al., 2019). However, no in vivo test has been carried out to investigate the implications of NAC on the side effects of IR. NAC was injected intraperitoneally 24 h before and 2 h before IR (Fig. 9A). No changes in ABR were observed after the injection of either sodium chloride (NaCl) or NAC alone. Because NAC is a ROS scavenger, the ROS levels in the NAC+IR and IR groups were compared. ABR thresholds at 8, 16, and 32 kHz changed with time in the C57BL/6 mice that had received NAC+IR treatment.


Upon ROS modulation by NAC before IR, the threshold shift at 16 and 32 kHz were reduced at 1 and 2 weeks after NAC+IR treatment compared with that of the IR-treated groups, whereas the threshold shift at 8, 16, and 32 kHz were all reduced at 4 weeks, and the threshold shift at 8 and 32 kHz were reduced at 8 weeks after NAC+IR treatment compared with that of the IR-treated groups (Fig. 9B). Nevertheless, the P1 wave amplitude was noticed to decrease after IR and did not recover even after NAC injection (Fig. 9D). Modulating ROS by NAC via IP injections alleviated IR-induced OHC loss in all turns when assessed at 4–8 weeks after IR (Fig. 9E, F). These results combined show the ability of NAC treatment in attenuating auditory threshold shifts and preventing IR-induced loss of OHCs to some extent.
Because it was found out earlier that IR leads to functional and morphological impairments in the SV, the ability of NAC treatment in improving SV function after IR was subsequently assessed. No changes were observed in the EP of both the control and the NAC-only groups, but changes were observed when EP of mice in the NAC+IR group was compared with that of IR-treated mice at 1, 2, and 4 weeks after IR treatment, signifying the effective alleviation of IR-induced EP changes when ROS was modulated by NAC. Nonetheless, no significant change was observed at week 8 (Fig. 9C).
NAC inhibits IR-induced ROS formation and damage
Data obtained from this study show that IR significantly increased the level of ROS in the SV and OHCs (Fig. 7). Because the loss of OHCs and the level of ROS in the basal turn were observed to be significantly increased after IR and because such damage follows a base-to-apex gradient, 4-HNE immunolabeling in the basal turn of the SV and OHCs was quantified. It was noticed that the IR-induced enhancement of 4-HNE in the SV at 1 and 8 weeks after treatment was blocked to some extent after ROS was modulated by NAC (Fig. 10A, C), whereas IR-induced increase of immunolabeling for 4-HNE (red) and PARP-1 (red) in the basal turn of the OHCs was also prevented at 8 weeks after treatment owing to NAC (Fig. 10E–H). On the contrary, IR-induced decrease of ZO-1 (red) immunolabeling in the SV was noticed to be inhibited by NAC treatment at 2 weeks after IR (Fig. 10B, D).


Discussion
With the rapid development in medical radiology techniques and the widespread application of traditional and modern three-dimensional conformal RT technology, hearing loss after head and neck RT has become increasingly prominent (Wang and Tepper, 2021; Young, 2019). It is challenging to directly acquire inner ear tissue from patients undergoing head and neck RT, hence a mouse IR model was established instead. With that, the hearing threshold was found to change progressively over time after IR, typically starting at a high frequency and progressed to lower frequencies over time (Gupta et al., 2020; Schuette et al., 2020; Zhu et al., 2019), a finding that is in good agreement with clinical findings (Cohen-Cutler et al., 2021).
The C57BL/6 mice RISNHL model revealed the pathological characteristics of the inner ear after IR. One of the typical characteristics is the marked absence of HCs, which are thought to be the part of the cochlear structure that is most vulnerable to damage at the cellular level (Kurabi et al., 2017). Therefore, past studies on RISNHL tended to focus on OHCs. However, some researchers have a different opinion (Tan et al., 2013). A single dose of 10–15 Gy IR resulted in the loss of IHC and the disorder of OHC arrangement but no loss of OHCs (Kim and Shin, 1994). This trend was reversed on day 30 when a large number of OHCs were lost without any IHC loss (Chen et al., 2021; Dinh et al., 2017; Liu et al., 2022).
This finding confirms the results of this study as no damage to the IHC was noticed after IR treatment, whereas significant loss of OHCs occurred at week 4 (28 days). However, the increase in the hearing threshold at 1–2 weeks after IR could not be explained by the loss of HCs; thus, the synaptic ribbons and SV were considered instead because these parts are known to impact hearing as well. The ABR threshold increased at 1 week after IR, and EP changes were recorded at a low level at the same time, whereas the permeability of the SV began to increase at 2 weeks. The failure of the SV to maintain a normal cochlear internal environment may consequently lead to functional damage of the cochlear epithelial sensory nerve cells, resulting in hearing threshold shifts.
If the function of the SV is not recovered, then the sustained damage to epithelial sensory cells such as HCs may further deteriorate to pathological changes. Although it is difficult to accurately detect synaptic function and morphology in vivo, changes in the ABR P1 wave amplitudes induced by a suprathreshold stimulus and quantification of the binding or overlapping of Ctbp2 and GluA2 immunolabeling (Liberman and Kujawa, 2017) may reflect the number, shape, and structural abnormalities of synapses to some extent. Results obtained from this study showed that the amplitudes of the P1 wave in the ABR threshold stimulation decreased significantly compared with those in the control group only at 8 weeks after IR, but the number of binding synaptic ribbons did not decrease significantly at the corresponding positions at week 1–8 after IR.
This finding suggests that the change in ABR hearing threshold after IR exposure may not be related to the decreased amplitude of the P1 wave or that it is not a “synaptic disease.” Several explanations were considered for the decrease in the amplitude of the P1 wave and the absence of a change in the number of binding synapses at 8 weeks after exposure. First, the nervous system is thought to be somewhat resistant to radiation (Mujica-Mota et al., 2014). Second, the amplitude of the P1 waves only reflects synaptic ribbons in an indirect manner and does not reflect true differences in terms of synaptic ribbon conditions, which are also affected by other changes in the auditory pathway (Braude et al., 2015; Kujawa and Liberman, 2009), such as nerve fiber density or the number of spiral ganglion cells.
Of interest, the EP was reduced at 1 week after IR, which seems consistent with hearing threshold shifts. However, the EP recovered at 8 weeks after IR without treatment. This effect may be owing to the adaptive nature of human body regulation, causing K+ to reach a new equilibrium point in the transfer process and ensuring homeostasis of the EP. Although EP remained normal after 8 weeks, irreversible pathological changes in the cochlea had already occurred. The BLB in the SV is critical for ion exchange, so that a positive EP could be maintained as an electrophoretic driving force for HC depolarization (Zhang et al., 2012) and for the stability of the cochlear microenvironment. The ABR threshold increased at 1 week after IR, and EP changes were recorded at the same time.
EP remained at a low level at 2 weeks, whereas the permeability of the SV began to increase at this time, although the histopathologic changes in the lateral wall of the cochlea were not obvious. The failure of the SV to maintain a normal cochlear internal environment may consequently lead to functional damage of the cochlear epithelial sensory nerve cells, resulting in hearing threshold shifts. If the function of the SV is not recovered, then the sustained damage to epithelial sensory cells such as HCs may further deteriorate to pathological changes. It was therefore hypothesized that disruption of the SV after IR coincides with the onset of hearing loss. It is therefore crucial for intervention and protection measures to be applied when only functional changes in the SV have occurred, because then further irreversible pathological changes in the HCs and the SV may be delayed or reversed.
Data collected from this study demonstrated that ROS levels in the SV were increased by IR. A previous study discovered that overactivation of PARP-1 implies DNA damage by ROS production (Zhou et al., 2022). Results from this study pointed out that IR increases the expression of PARP-1 in the SV and the OHCs, in line with IR-induced ROS, implying the damage of DNA in the SV. As reported in the literature, capillary thinning, destruction, extravasation, petechiae formation, and inflammation are more obvious with microvascular IR (Venkatesulu et al., 2018). Radiation-induced vascular endothelial injury may alter the balance of pro- and anti-inflammatory cytokines and induce the release of large levels of ROS (De Ruysscher et al., 2019), resulting in leaky and disorganized vascular networks that alter normal vascular homeostasis.
It can therefore be hypothesized that ROS may not only act on the HCs but it may also be an important cause of early SV damage. Therefore, dysfunction and structural damage of the SV in the cochlea are important causes of hearing loss after IR despite the absence of OHC loss in the early stages.
Besides being known to reduce ROS levels, the antioxidant NAC also protects HEL-OC1 cells after a single radiation dose of 20 Gy (Duzenli et al., 2019). However, insufficient in vivo studies have been conducted to confirm the correlation and validity of these effects. In vivo experiments were therefore performed in this study to confirm the effectiveness of NAC, and the animal models confirmed that the ROS scavenger NAC can be used to reduce cochlear SV and OHC ROS levels. EP and ZO-1 levels were also improved to some extent upon treatment with NAC, ultimately protecting hearing after IR. It is thus believed that the dynamic balance of ROS in the SV affects the function of the SV and may regulate cochlear homeostasis, which is an important factor affecting hearing.
More importantly, NAC acts on the SV and protects hearing in mice after IR. At present, no ideal radiation protectors are available for clinical use, although several potential candidates have been reported. NAC antioxidants were chosen because they are safe and can cross the labyrinth barrier. In this study, NAC was administered via intraperitoneal (i.p.) injection, which poses a limitation. In the future, it is hoped that NAC could be administered locally through the inner ear to avoid the effect of systemic administration on the tumor.
In summary, results from this study collectively address functional and morphological impairment in the SV during the early stage of hearing loss induced by IR, as indicated by EP reduction and hyperpermeability. The expression of ZO-1 also decreases after IR, impacting the ability of TJs in regulating vascular integrity and permeability. Disruption of the SV and loss of the OHCs were subsequently verified to be attributed by ROS-related increases in 4-HNE level and DNA damage-related increases in the level of PARP-1. ROS-related damage to cochlear HC and SV is supported by the effects of NAC treatment in mice after IR. Finally, the modulation of ROS by NAC in C57BL/6 mice reduced radiation-induced ROS-related damage and hearing loss, indicating the potential protective effect of NAC against RISNHL.
Materials and Methods
An electronic laboratory notebook was not used.
Animals and study groups
Healthy 6-week-old C57BL/6 male mice weighing 15–22 g were obtained from the Laboratory Animal Center of Sun Yat-sen University. The experiments were performed only on male mice to avoid the influence of sex on the results. The C57BL/6 mouse has been used as an animal model for RISNHL in several articles (Liu et al., 2022; Zhang et al., 2020). A characteristic of the C57BL/6 mouse strain is moderate tolerance to radioactive substances, which could therefore avoid additional mouse mortality. C57BL/6 is also the most widely used model in the study of adverse reactions caused by radiation in multiple sites, such as brain injury, intestinal injury, and lung injury (Dong et al., 2015; Wilson et al., 2009; Zhang et al., 2023).
According to the literature, the commonly used C57BL/6 strains develop progressive high-frequency hearing loss as early as 3 months of age and severe low-frequency hearing loss by 12–13 months of age, although no progressive EP decline is observed at >24 months of age (Bowl and Dawson, 2019; Francis et al., 2003; Hequembourg and Liberman, 2001). However, most age-related hearing loss is apparent only 5 or 6 months after birth (Ohlemiller, 2006; Shin et al., 2021; Zheng et al., 1999). In this study, radiation was administered to mice at the age of 6 weeks when they were sexually mature, and the changes were observed at 8 weeks after IR exposure. By this time, the mice were no more than 14 weeks (3.5 months) old.
Moreover, the ABR results at 8, 16, and 32 kHz from 1 to 8 weeks after IR were compared with that of the control group and no significant difference was found in terms of ABR thresholds. According to the ABR test results of the control group at the same age, the C57BL/6 mice had normal hearing without age-related hearing loss. The establishment of age-matched controls allowed us to safely proclaim that age did not affect the C57BL/6 mice used in the experiments of this study, although it is recognized that age may have an effect on the mice. Detailed results are given in Supplementary Figure S4.
All mice used in the experiments had free access to water and a regular mouse diet. These mice were acclimatized in a standard animal room at 22°C for at least 7 days. All C57BL/6 mice with an abnormal baseline ABR or middle ear pathology were excluded at the beginning of the study.
A total of 284 mice were randomly divided into 5 groups: The control group (Ctrl) received no treatment (n = 88); The IR group was treated with 15 Gy IR (n = 88); The NAC group only received NAC (i.p.) (n = 36); The NAC+IR group received NAC (i.p.) 24 h and 2 h before IR via 15 Gy RT (n = 36); The NaCl+IR group (NaCl+IR) received saline (i.p.) 24 h and 2 h before IR via 15 Gy RT (n = 36).
Results from both the NaCl+IR group and the IR group were combined because the outcome was the same for both groups, whereas the group receiving only the vehicle (NaCl) served as the control group (Ctrl). The sample preparations procedure is listed in Supplementary Table S2.
In each group, the mice were monitored 1, 2, 4, and 8 weeks after IR. All mice were also monitored for weight loss and fur condition. Animal care and use, as well as the experimental protocol, were reviewed and approved by the Animal Research Committee of Sun Yat-sen University and by the Animal Research: Reporting In Vivo Experiments guidelines. Figure 1A illustrates the general experimental timeline.
Drug administration via the i.p. route
Throughout the experiment, mice were anesthetized with sodium pentobarbital (1%; 50 mg/kg) via the i.p. route.
NAC protects HC structure against cisplatin-induced ototoxicity (NAC, 400 mg/kg) (Thomas Dickey et al., 2004) and noise-induced ototoxicity (NAC, 325 mg/kg or 300 mg/kg) (Lu et al., 2014; Wu et al., 2020; Yuan et al., 2015). With reference to the above literature on animal experiments, the concentration of 325 mg/kg was chosen for a preliminary test as a safety measure. This concentration did not cause death in the animals but hearing changes were observed, so it was used as the standard concentration of NAC throughout this study. NAC (A9165; Sigma-Aldrich) was dissolved in saline to make a concentrated stock solution (60 mg/mL) and stored at −20°C.
Before use, the final pH of the solution was adjusted to 7.4 with sodium hydroxide. NAC at a dose of 325 mg/kg was administered 24 h before and 2 h before IR via the i.p. route. Mice in the blank group received only the same volume of NaCl. The control group and the blank group were eventually combined into one single control group. Figure 9A illustrates the NAC experimental timeline.
Local radiation
Before IR, the animals were anesthetized with sodium pentobarbital (1%, 50 mL/kg, i.p.) and kept in a prone position. Mice were irradiated with an RS 2000pro irradiator (Rad Source Technologies, Inc.). A lead plate with a size of 150 mm × 150 mm × 1.0 cm was constructed to protect other organs, such as the eyes, respiratory tract, and digestive tract, from exposure. A rectangular opening of 8 × 35 mm was drilled to expose only the area near the ear. The auricular–temporal region was chosen as the IR field, and the nonirradiated body parts were covered with a lead plate. Each mouse from the IR groups was exposed to radiation at a calculated dosage of 15 Gy at 160 kV and 25 mA. Supplementary Figure S1 illustrates the instrument.
Marks et al. (2010) estimated a dose–volume limit for hearing at 14–15 Gy in a single dose clinically. To reduce mortality while achieving sensorineural hearing loss, a single radiation treatment was considered for this study. According to previous reports, C57BL/6 mice demonstrated sensorineural deafness upon the administration of 10–20 Gy in therapy doses (Chen et al., 2021; Liu et al., 2022; Zhang et al., 2020), which is consistent with the results of our preliminary test. The dosage of 15 Gy was ultimately chosen for the experiments in this study as preliminary test results showed that a 15 Gy dose yielded almost no mortality yet with sensorineural hearing loss occurring.
ABR measurements
ABR measurement was randomly performed in groups of six mice that were anesthetized with sodium pentobarbital (1%; 50 mg/kg, i.p.). Body temperature of these mice was maintained at 37°C using a heating pad. The ABR threshold was recorded in a double-walled soundproof room. The hypodermic needle electrodes were inserted into the top of the skull (active), in the back near the tail (grounded), and under the left ear (reference). Hardware and software from Tucker-Davis Technologies (TDT System III, Alachua, FL) were used to generate the acoustic signal and process the response. The stimulus level was calibrated, and the stimuli were applied in a free field.
Up to 1024 responses were collected and averaged for each stimulus level at 8, 16, and 32 kHz, by reducing the sound intensity in 5-dB steps until the threshold was reached. The threshold was defined as the lowest stimulus level at which a repeatable waveform could be identified. The ABR was evaluated at 1, 2, 4, and 8 weeks after IR treatment.
The P1 wave of ABR was interpreted to represent the action potential of auditory nerve fibers. Suprathreshold ABR P1 wave amplitude was defined as the value (uV) from the baseline to the first spike, in response to stimuli at 90 dB sound pressure level. P1 wave amplitude is often thought to correlate with synapses. Latency was the time (ms) at which the peak occurred. Therefore, the amplitude and latency of the P1 wave evoked by a 90 dB sound pressure level stimulus were analyzed. Supplementary Figure S2 illustrates representative waveforms of ABR and P1 waves.
EP measurement
After ABR recording, EP measurements were randomly performed on groups of four mice at 1, 2, 4, and 8 weeks after IR treatment. After the mice were anesthetized, the bulla of the left ear was exposed through the ventral approach for easy operation. After the bulla was opened, a small hole was carefully drilled with a polishing drill in the basal turn cochlear bone. A glass capillary pipette was equipped with 150 mM KCl and installed on the micromanipulator. To record the EP from the scala media, the micromanipulator was rotated gently until the electrode came into contact with the basement membrane, and the baseline was recorded. The glass electrode was then inserted into the scala media and the reading was recorded. The reading typically ranges from +80 to +120 mV as the electrode enters the scala media.
Histological preparation
All mice were killed with a lethal dose of sodium pentobarbital (i.p.) under deep anesthesia, and their cochleae were harvested.
Basement membranes were prepared for immunofluorescent staining
The extracted cochleae were immersed in 4% paraformaldehyde (dissolved in phosphate-buffered saline [PBS], pH 7.4), the stapes were excised, and a drill hole was made in the apex of the cochlea using a 1 mL syringe needle under a microscope. Cochleae were perfused with 4% paraformaldehyde until no air bubbles were observed. Then, the cochleae were fixed overnight at 4°C by immersing in 4% paraformaldehyde. After decalcification with 4% sodium ethylenediaminetetraacetic acid (EDTA) solution (pH 7.4) at 4°C for 48 h, the cochleae were microdissected into three turns (apex, middle, and base). The specimens were subsequently incubated with 3% Triton X-100 for 30 min. After washing three times in PBS, the specimens were incubated for another 30 min at room temperature in 10% goat serum to block nonspecific antibody binding.
Next, the specimens were incubated at 4°C for 36 h with the following primary antibodies at a concentration of 1:200: rabbit polyclonal anti-4-HNE (ab46545; Abcam), rabbit polyclonal anti-PARP-1 (ab227244; Abcam), and rabbit polyclonal ZO-1 antibody (40-2200; Thermo Fisher). The specimens were then incubated with the secondary antibody anti-rabbit (Alexa Fluor 594) at a concentration of 1:500 and phalloidin (Alexa Fluor 488) at a concentration of 1:500 for 1 h at 37°C in the dark. After the specimens were washed three times, DAPI (10 mg/mL; Sigma Aldrich) was added for 10 min. After being washed, each sample was fixed on a glass slide in glycerol, and the edges were sealed with nail polish. Control incubations were routinely processed without primary antibody treatments. The sample was imaged with a confocal microscope (Carl Zeiss, Germany) and analyzed with the ImageJ software.
Basement membranes were prepared for IHC synapse immunofluorescence staining
The extracted cochleae were immersed in 4% paraformaldehyde (dissolved in PBS, pH 7.4), the stapes were excised, and a drill hole was made in the apex of the cochlea using a 1 mL syringe needle under a microscope. Cochleae were perfused with 4% paraformaldehyde for 1.5 h at room temperature in a fast shaker. This step differs from those used in the other immunofluorescence staining protocols. After decalcification with 4% sodium EDTA solution (pH 7.4) at 4°C for 24 h, the cochleae were microdissected into three turns (apex, middle, and base). The specimens were subsequently incubated in 3% Triton X-100 for 30 min at room temperature. After washing three times with PBS for 5 min per wash, the tissues were incubated for another 30 min in 10% goat serum to block nonspecific antibody binding.
Next, the specimens were incubated at 37°C in the dark overnight with the following primary antibodies at a concentration of 1:200: rabbit anti-myosin VIIa polyclonal antibody (25-6790; Proteus Biosciences), mouse anti-GluA2 IgG2a (MAB397; Millipore), and mouse anti-CtBP2 IgG1 (612044; BD Biosciences). The specimens were then incubated with the secondary antibodies anti-rabbit IgG (Alexa Fluor 647), anti-mouse IgG1 (y1) (Alexa Fluor 594), and anti-mouse IgG2a (Alexa Fluor 488) at a concentration of 1:200 for 1 h at 37°C in the dark. Control incubations were routinely processed without primary antibody treatments.
Frozen sections were used for immunofluorescence staining
The extracted cochleae were perfused with 4% paraformaldehyde and kept in this fixative overnight at 4°C. The cochleae were decalcified in 4% sodium EDTA solution for 48 h. Then, the specimens were incubated in 15% sucrose for 36 h followed by 30% sucrose for 48 h at 4°C. Next, the cochleae were embedded in O.C.T. compound (Sakura) and processed as resin-embedded specimens. Before use, frozen cochlear sections were mounted on glass slides and stored at −20°C for performing immunohistochemistry at 12 μm. The frozen sections were incubated in 3% Triton X-100 for 30 min and blocked with 10% goat serum for another 30 min at room temperature.
Then, the frozen sections were incubated overnight at 4°C with the following primary antibodies: rabbit polyclonal anti-4-HNE, rabbit polyclonal anti-PARP, and rabbit polyclonal ZO-1 (40-2200; Thermo Fisher). The specimens were then incubated with the anti-rabbit (Alexa Fluor 594) secondary antibody at a concentration of 1:500 for 2 h at 4°C in the dark. After being washed, the specimens were treated with DAPI for 10 min. After the specimens were washed again, the plate was sealed using glycerol. Control incubations were routinely processed without primary antibody treatments.
Paraffin-embedded tissue sections were used for HE staining
The cochleae were dissected, fixed, and decalcified as described for frozen section immunostaining. After successive dehydration in ethyl alcohol at 50%–70%–80%–90%–95%–100%–100%, the inner ears were incubated twice in xylene for 10 min at room temperature. The xylene in the inner ear was then replaced with paraffin wax at 70°C for 3 h. Each cochlea was then embedded in a paraffin block and sliced into 3–5 μm slides. Paraffin sections were stained with HE according to the manufacturer's protocol. After HE staining, images were acquired with a light microscope.
Detailed information on the antibodies is given in Supplementary Table S1.
Quantifying HCs and synaptic ribbons
The cochleae were microdissected into three parts from the apical turn to the basal turn. Both inner and OHCs were identified through their immunoreactivity for myosin VIIa. Phalloidin-fluorescein conjugate was used to label F-actin so that the presence or absence of IHCs and OHCs can be observed. HCs were immunolabeled with myosin VIIa or phalloidin. Images were captured using a 40 × lens and a 63 × lens on a Zeiss confocal microscope. The percentage of OHC and IHC loss per 0.5 mm of epithelial cells was recorded. The distances between the apex and the base were 10%–20%, 40%–60%, and 60%–80% of the total cochlea, roughly corresponding to the frequency ranges of 8, 16, and 32 kHz (Viberg and Canlon, 2004), respectively.
Myosin VIIa was used to stain IHCs, CtBP2 was used to stain presynaptic ribbons, and GluA2 was used to stain postsynaptic ribbons. Images were captured using a 63 × lens on a Zeiss confocal microscope from the 8, 16, and 32 kHz regions in each sample under identical Z-stack conditions with a z-step size of 0.3 μm. Each stack spanned the entire synaptic ribbon of the HCs in the z-dimensions. The number of synaptic ribbons for each IHC was counted using the ImageJ software 3D object counter. For synapse counting, colocalized bound pre- and postsynaptic immunofluorescent spots were counted.
Assessment of SV permeability and SV thickness
EB (E2129, MW: 961 Da; Sigma Aldrich) was used to compare the vascular permeability of mice in the control group and the IR-treated group by the quantitation of EB extravasation and observation of EB dye staining. When the permeability of blood vessels increases, the albumin bound to EB leaks, so the level of EB exudation can be measured to reflect the degree of permeability. The fluorescence wavelength has a strong peak at 470 and 540 nm and a weak peak at 680 nm (Gu et al., 2022; Radu and Chernoff, 2013). After the mice were anesthetized, EB was injected through the intercanthal vein and the limbs and tail were blue for ∼10 min. Cochleae were immediately harvested and fixed with 4% paraformaldehyde for 2 h. Cochlear SV tissue was carefully dissected and visualized under a laser-scanning confocal microscope.
Frozen sections were used for immunohistochemical analyses, and paraffin sections were used for HE analyses. Apical, middle, and basal regions of the HE-stained cochlear sections were captured using a 40 × objective lens under the light microscope (Leica Microsystems), so that SV thickness could be measured and calculated by the ImageJ software. SV thickness was measured in the midpoint between the attachment of Reissner's membrane and the spiral prominence (Gu et al., 2022). The mean value from four cochleae was used to define the width of the SV.
Western blotting analysis
Cochlear tissues were homogenized in ice-cold radioimmunoprecipitation assay lysis buffer (CW2333; CWBIO) with 1% protease inhibitor cocktail (100 × ) (CW2200S; CWBIO) for 30 min. The sample was centrifuged at 12,000 g and 4°C for 30 min, and the supernatant was collected. The total protein concentration was determined using the BCA Protein Assay Kit (P0012S; Beyotime). Equal amounts of protein were electrophoresed on 12.5% sodium dodecyl sulfate– polyacrylamide gel electrophoresis (SDS‒PAGE) and transferred to a polyvinylidene fluoride (PVDF) membrane (Millipore, MA). The PVDF membrane was blocked with 5% nonfat dry milk in TBS with 0.1% Tween-20 (FD0020-500), incubated with anti-ZO-1 and anti-PARP-1 antibodies at a concentration of 1:1000 overnight, washed three times (at 10-min intervals) using the TBST buffer and incubated for 1 h with the appropriate horseradish peroxidase–conjugated secondary antibody (1:10,000) at room temperature. After the membrane was washed, the immunoreactive bands were visualized by enhanced chemiluminescence (Millipore). GAPDH was used as an internal control.
Statistical analysis
All data and statistical analyses were performed using the SPSS 22.0 and GraphPad software. Data are presented as mean ± standard deviation based on sample size and variability within groups. One-way analysis of variance with Fisher's post hoc test, two-sample t-test, and one-sample t-tests were performed for statistical analyses. p Values of <0.05 were considered as statistically significant.
Institutional Review Board Statement
This study followed the guidelines for the care and use of laboratory animals. The Animal Care and Use Institutional Committee of Sun Yat-sen University approved all procedures in this study (Approval No. SYSU-IACUC-2022-000802).
Footnotes
Acknowledgments
The authors acknowledge Chinese Academy of Sciences Institute of Guangdong Provincial Key Laboratory of Malignant Tumor Epigenetics and Gene Regulation and Medical Research Center in Sun Yat-Sen Memorial Hospital of Sun Yat-Sen University for providing support and assistance for this article. The authors thank Prof. Yamei Tang (Department of Neurology, Sun Yat-Sen Memorial Hospital, Sun Yat-Sen University), Prof. Hao Xiong and Dr. Junbo Zeng (Department of Otolaryngology, Sun Yat-Sen Memorial Hospital, Sun Yat-Sen University) for help to experimental design. The authors thank Prof. Jie Tang (Department of Physiology, School of Basic Medical Sciences, Southern Medical University) group for helping us with the EP measurement. The authors express their gratitude to Prof. Peie Cai and AJE for the expert linguistic services provided.
Authors' Contributions
G.Y., P.J. and Z.Y. conceived and designed the experiments; W.F. and H.W. provided experimental guidance; G.Y. and H.W. performed the experiments; G.Y., P.J., W.F. and C.Z. analyzed the data; G.Y. wrote the article; P.J. and Z.Y. reviewed the article. All authors read and approved the final version of the article.
Author Disclosure Statement
No competing financial interests exist.
Funding Information
This work was supported by the National Natural Science Foundation of China (No. 82171138) and Guangdong Natural Science Foundation (Nos. 2020A1515011321 and 2022A1515012608).
Supplementary Material
Supplementary Table S1
Supplementary Table S2
Supplementary Figure S1
Supplementary Figure S2
Supplementary Figure S3
Supplementary Figure S4
Abbreviations Used
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.