
Abstract
Significance:
The need of cells to constantly respond to endogenous and exogenous stress has necessitated the evolution of pathways to counter the deleterious effects of stress and to restore cellular homeostasis. The inability to activate a timely and adequate response can lead to disease and is a hallmark of aging. Besides protein-coding genes, cells contain a plethora of noncoding regulatory elements that allow cells to respond rapidly and efficiently to external stimuli by activating highly specific and tightly controlled mechanisms. Many of these programs converge on the regulation of translation, one of the most energy-consuming processes in cells.
Recent Advances:
The noncoding dimension of translational regulation includes short and long noncoding ribonucleic acids (ncRNAs), as well as messenger RNA features, such as the sequence and modification status of the 5′ and 3′ untranslated regions (UTRs), that do not change the amino acid sequence of the produced protein.
Critical Issues:
In this review, we discuss the regulatory role of the nonprotein-coding components of translation under stress, particularly oxidative stress. We conclude that the regulation of translation through ncRNAs, UTRs, and nucleotide modifications is emerging as a critical component of the stress response.
Future Directions:
Further areas of study using long-read sequencing technologies will be discussed. Antioxid. Redox Signal. 39, 374–389.
Introduction
During oxidative stress, cells accumulate high levels of reactive oxygen species (ROS) that can originate from endogenous or external sources such as the mitochondria (Larosa and Remacle, 2018), cadmium, arsenite, and pollutants (Archer, 2011; Nemmiche, 2017). While ROS have several established roles in signaling (Archer, 2011; Nemmiche, 2017), their highly reactive nature and unregulated increase can cause oxidation of proteins (Holmstrom and Finkel, 2014; Reczek and Chandel, 2015; Schieber and Chandel, 2014), deoxyribonucleic acid (DNA) (Dizdaroglu and Jaruga, 2012), ribonucleic acid (RNA) (Li et al., 2014; Wilkinson et al., 2021), and lipids (Angelova et al., 2021). Oxidation of these fundamental molecules can lead to permanent damage and cellular dysfunction and, if unchecked, cell death. Cells have evolved with mechanisms to defend against oxidative stress by producing detoxifying enzymes, amino acids, and other molecules (Gorrini et al., 2013), as well as by limiting the amounts of endogenously produced ROS.
As the most energy-intensive process in the cell, translation requires high levels of mitochondrial respiration that generates a substantial amount of ROS (Leibovitch and Topisirovic, 2018), and is thus tightly regulated as part of an oxidative stress response.
While the factors that regulate translation are most readily associated with the coding region of messenger RNA (mRNA), as well as the protein components of the translation machinery, noncoding RNA (ncRNA) and the untranslated regions (UTRs) of mRNA are equally essential regulatory elements. ncRNAs are a diverse class of structural and functional RNAs with roles across all areas of biology (Goodall and Wickramasinghe, 2021). In translation, ncRNAs such as ribosomal RNAs (rRNAs) are responsible for the function and structure of the ribosome, the delivery of amino acids to the ribosome by transfer RNA (tRNA), and the modulation of translation efficiency through long ncRNA sponging of micro-RNAs (miRNAs) or direct binding to mRNA.
In contrast, UTRs are present on the 5′ and 3′ ends of coding mRNAs, and while they are not actively translated themselves, they are targets of regulatory pathways that govern translation. The UTRs are also frequent substrates for nucleotide modifications that can alter their function in translation. In this review, we discuss the mechanisms by which oxidative stress alters translation from the perspective of noncoding elements such as ncRNAs, UTRs, and RNA modifications. We describe how these noncoding elements participate in translation and serve as both critical modulators of translation efficiency and regions of vulnerability to oxidative stress. Finally, we discuss future outlooks for the study of noncoding elements in translation and advancement of new technologies applicable to the field.
ncRNA Elements in Translation
Protein translation, the synthesis of protein from the mRNA template, can be divided into three phases of ribosomal action: initiation, during which ribosomes are loaded onto the mRNA; elongation, during which the polypeptide chain is synthesized according to the mRNA coding region; and termination, during which the nascent peptide chain is released and ribosomal subunits are recycled. While the subject of translation is the open reading frame (ORF) corresponding to the mRNA region coding for the protein, the act of translation requires numerous ncRNA elements, including some within the ribosome itself (Fig. 1).

Translation begins with a mature mRNA that has completed splicing and that contains a modified guanine nucleotide N7-methylguanosine (m7G) cap at the 5′ end. Capped mRNAs are recognized in the cytoplasm by the eIF4F complex (eIF4E, eIF4G, and eIF4A), which serves both scaffolding and helicase functions, and eIF4B, which enhances the helicase function of eIF4F (Jackson et al., 2010). Part of the function of eIF4F is to facilitate ribosome loading by unwinding the secondary structure in the mRNA 5′ UTR (Pestova and Kolupaeva, 2002). After the mRNA is loaded with the eIF4F complex and the 5′ UTR is unwound, the 43S preinitiation complex, composed of the 40S small ribosomal subunit, the ternary complex (eIF2-GTP-tRNAiMet), and other initiation factors (Jackson et al., 2010), is recruited. This process is commonly referred to as cap-dependent or 5′-dependent translation and represents most translation events in cells.
A separate cap-independent form of translation also exists and uses internal ribosome entry sites (IRESs) to recruit the 43S complex, rather than an eIF4F-cap structure (Yang and Wang, 2019); this mechanism of translation is discussed later in this review. Even though mRNAs largely begin translation using a 5′-dependent mechanism, the efficiency by which ribosomes engage with mRNAs is not equal among all transcripts. Initiation efficiency is determined by sequence elements and structures in the 5′ UTR (Leppek et al., 2018), which can also act as a mechanism for selective translation during stress, as is the case for the mammalian target of rapamycin complex 1 (mTORC1) pathway discussed further below.
Once loaded, the 43S preinitiation complex begins “scanning” through the 5′ UTR in search of an AUG start site that the tRNAiMet of the loaded ternary complex can pair with. To ensure fidelity of start site selection, the intended start site that defines the expected ORF is often in an optimal context defined by the “Kozak Sequence” (GCCRCCAUGG in vertebrates), in which the most important nucleotides are a purine at −3 and a G at +4, relative to the A in the AUG codon (Hernandez et al., 2019). However, start sites are not exclusive to the expected ORF, and can appear elsewhere in the 5′ UTR where they can also act as alternative translation initiation sites and define upstream ORFs (uORFs). A growing number of mRNAs have been shown to contain uORFs (McGillivray et al., 2018), which can regulate translation through multiple mechanisms (Young and Wek, 2016).
Regardless, after start site selection, the guanosine triphosphate (GTP) of the ternary complex is consumed to guanosine diphosphate and a free 60S ribosomal subunit binds to the committed 40S subunit, displacing the remaining initiation factors (Jackson et al., 2010) and forming an 80S monosome.
Elongation begins with a formed 80S monosome loaded with a tRNAiMet in the P-site that is paired to the AUG start site in a codon–anticodon interaction. tRNAs are highly structured ncRNAs that are essential for elongation, as they are the source of amino acids during protein synthesis. Amino acids are covalently attached to tRNA by aminoacyl-tRNA synthetases and recognized by the elongation factor eEF1A that then brings the tRNA to the A-site of the ribosome (Dever and Green, 2012). In the A-site, the anticodon loop of the tRNA is paired to the codon of the mRNA, assisted by rRNA bases in the small subunit. tRNA pairing in the A-site is a rate-limiting step during elongation and is dependent on factors such as tRNA abundance, the strength of pairing between the codon and anticodon, and even neighboring codons (Brule and Grayhack, 2017; Hanson and Coller, 2018).
tRNAs are also regulated extensively during stress, and can be altered in abundance depending on cellular conditions (Gingold et al., 2014), modification status (Chan et al., 2010; Endres et al., 2015), and other types of regulation discussed below. After the tRNA is accommodated in the A-site and eEF1A is released, peptide bond formation occurs between the amino acid of the A-site tRNA and the P-site peptidyl chain in the peptidyl transfer center of the ribosome; the ribosome then translocates one codon, assisted by the elongation factor eEF2, and new tRNAs are accommodated into the A-site (Dever and Green, 2012).
The process of elongation continues until a stop codon (UAG, UGA, UAA) occupies the A-site and is recognized by the release factors eRF1 and eRF3 (Gerovac and Tampe, 2019). Recognition of the stop codon by eRF1 triggers the release of the nascent peptide and the splitting of the ribosomal subunits by ABCE1 into a 60S subunit and an mRNA-tRNA-bound 40S subunit that is recycled before translation initiation can begin again (Pisarev et al., 2010). As with start codons, the context of stop codons determines the efficiency of the stop, which is important during premature stop codon insertions and in ribosome read through (Lombardi et al., 2022).
rRNA Susceptibility to Oxidative Stress
The ribosome is a ribonucleoprotein complex with an ncRNA core composed of rRNA for both the 40S subunit (18S rRNA) and the 60S subunit (28S, 5.8S, 5S rRNAs), around which a multitude of ribosomal proteins (33 for the 40S and 47 proteins for the 60S) are arranged (Henras et al., 2015). The maturation of rRNAs is a highly complex process that varies even among eukaryotes. Briefly, a single precursor rRNA is transcribed by Pol I and extensively processed and modified to yield the 18S, 28S, and 5.8S segments, while the 5S rRNA is transcribed separately by Pol III (Henras et al., 2015); the resulting mature rRNAs, along with the ribosomal proteins, are assembled into the pre-40S and pre-60S subunits (Panse and Johnson, 2010).
rRNA is the most abundant RNA species in the cell, accounting for ∼80% of cellular RNA content (Milo et al., 2010), while ribosomes themselves exist at an order of magnitude of 107 copies per cell (Wisniewski et al., 2014). The high abundance of these components makes ribosome biogenesis a highly energy-consuming process (Warner, 1999), and thus a major contributor to the generation of endogenous ROS. Much of ribosome biogenesis, including ribosomal DNA transcription, rRNA processing and maturation, and subunit assembly, occurs within the nucleolus, a membraneless region in the nucleus (Fig. 2) (Boisvert et al., 2007). Since the nucleolus functions as the factory for ribosomes, it is also a sensor for stress (Boulon et al., 2010). During oxidative stress, ROS can induce nucleolar stress and negatively affect rRNA processing, thus decreasing the number of ribosomes produced (Chou and Lo, 2019).
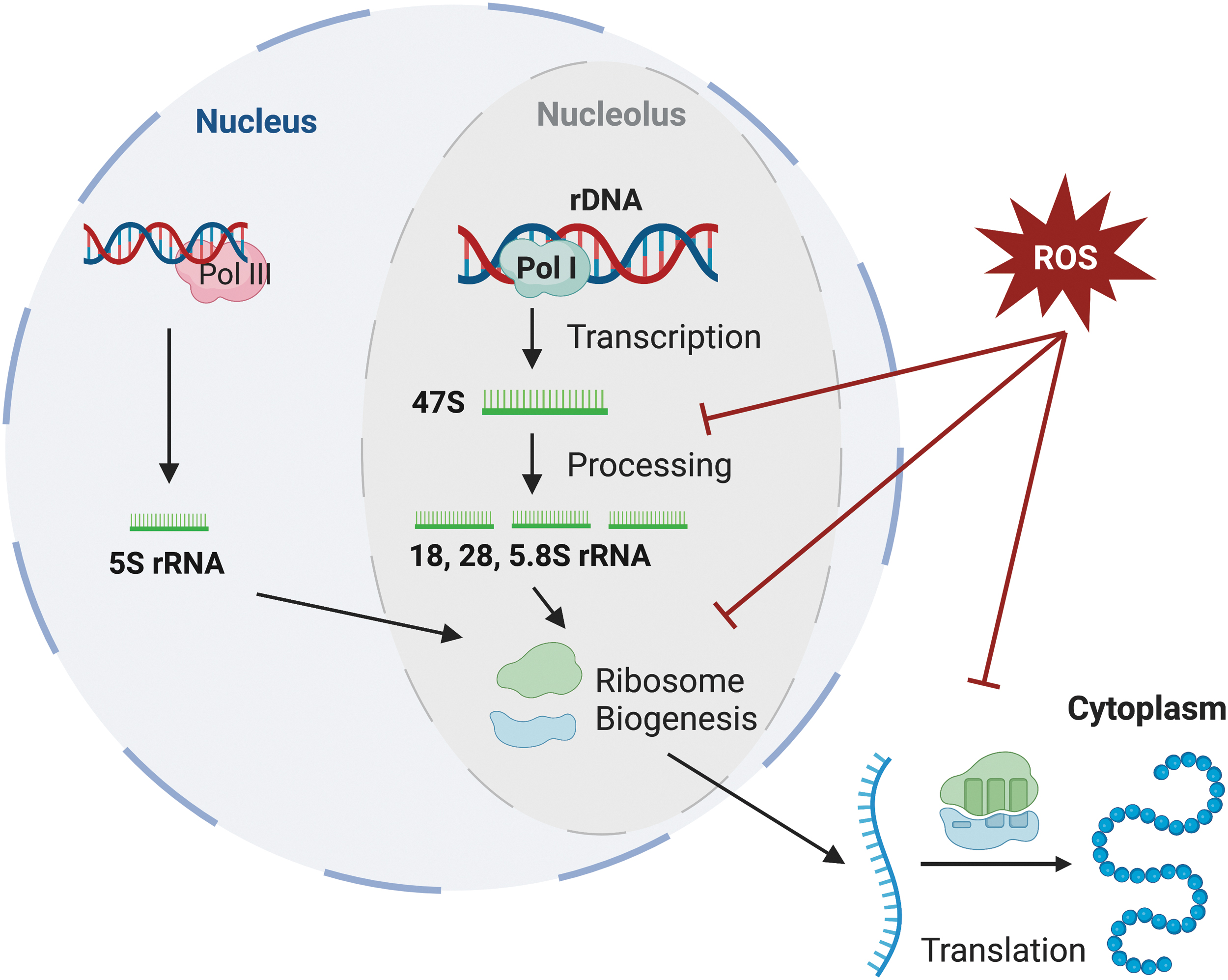
It has recently been shown that the first stages of rRNA processing are inhibited during oxidative stress (Szaflarski et al., 2022) and induce accumulation of precursor rRNAs (Sapio et al., 2021).
Besides inhibition of rRNA biogenesis, ROS can also directly damage rRNA and thus inhibit translation (Shcherbik and Pestov, 2019). rRNA has an extensive secondary structure that is essential to the formation of the ribosome. rRNA bases form critical components of the decoding and peptidyl transferase center of the ribosome, where codon–anticodon pairing and amino acid transfer to the nascent peptide chain occur, respectively (Khatter et al., 2015). ROS-mediated damage to rRNA is unevenly distributed across the rRNA. In bacterial ribosomes, oxidative stress is more pervasive in damaging the catalytic center of the large subunit than the decoding center (Willi et al., 2018). In yeast, endonucleolytic cleavage of a specific site in rRNA of the 60S subunit has been proposed to be a marker of oxidative stress (Shedlovskiy et al., 2017).
Furthermore, as with tRNA discussed later, oxidative stress can also induce rRNA cleavage (Thompson et al., 2008). Direct rRNA oxidation is associated with several neurodegenerative diseases, likely because of significant oxidative stress produced in the brain (Cobley et al., 2018). Increased rRNA oxidation is associated with early stages of Alzheimer's disease, and the resulting decrease in protein synthesis may be a critical stage in the development of the disease (Ding et al., 2012; Ding et al., 2006). Similarly, oxidative stress-induced dysfunctional ribosome biogenesis is also associated with Parkinson's disease (Parlato and Liss, 2014).
tRNAs Are Regulatory Targets of Oxidative Stress
tRNAs are short, stable, structured, and highly modified ncRNAs that are essential for translation (Phizicky and Hopper, 2010). Amino acids are covalently attached to the 3′ end of tRNAs, while the distal anticodon loop is used to decode mRNA codons during translation. While there are 20 canonical amino acids and 21 corresponding aminoacyl-tRNA synthetases (Rubio Gomez and Ibba, 2020), there are many more tRNA species owing to the degeneracy of the genetic code (61 codons for 20 amino acids), and an even greater number of tRNA genes, since a given tRNA species often exists with multiple DNA copies. In fact, 415 individual tRNA genes have been predicted in human cells (Chan and Lowe, 2016). This diversity of tRNA genes arises from the presence of tRNA “isoacceptors” and “isodecoders” (Schimmel, 2018).
tRNA species are considered isodecoders when the anticodon is identical but the tRNA body contains differences; for example, tRNAAla(AGC) has ∼15 variants (Chan and Lowe, 2016), although all of them decode the same alanine codon. The role of isodecoders in human biology and their effects on translation are poorly understood, although isodecoders have tissue specificity (Ishimura et al., 2014), indicating that this diversity may be important for cellular function.
In contrast, isoacceptors accept the same amino acid but recognize different mRNA codons owing to different anticodons; for example, the tRNASer isoacceptor family has four anticodons (AGA, CGA, UGA, GCU) that decode six serine codons in the mRNA (UCU, UCC, UCA, UCG, AGU, AGC). An important feature of isoacceptors is that codons and anticodons are not always exact matches, and both RNA modifications and non-Watson–Crick base pairing (termed wobble pairing) regulate the anticodon–codon pairing (Kubyshkin et al., 2018). As accommodation into the A-site is the rate-limiting step of translation elongation, this feature creates a substantial element of regulation referred to as codon optimality (Brule and Grayhack, 2017).
Aside from serving as critical components of translation, tRNA has also been shown to perform a regulatory role during oxidative stress in the form of tRNA-derived stress-induced RNAs (tiRNAs) (Fig. 3). tiRNA species are produced from endonucleolytic cleavage of tRNA in the anticodon loop by angiogenin (Thompson et al., 2008; Yamasaki et al., 2009), which is typically a progrowth protein involved in rRNA transcription when localized to the nucleus (Li and Hu, 2010; Tsuji et al., 2005). While the cleavage of tRNAs to produce tiRNAs might seem to regulate translation through tRNA reduction, this appears unlikely given that the amount of tRNA converted to tiRNAs is miniscule compared with the pool available for protein synthesis (Thompson et al., 2008). Instead, tiRNAs have been shown to have numerous other effects on biology, including gene expression, mRNA stability, and translation (George et al., 2022; Wang et al., 2022b).

Binding of tiRNAs to the ORF and 3′ UTR of mRNA encoding ribosomal proteins can enhance their translation, which has further downstream effects on ribosome biogenesis (Kim et al., 2019; Kim et al., 2017). tiRNAs can also act to decrease translation by displacing the eIF4G component of eIF4F from the 5′ UTR of mRNA, thus inhibiting translation initiation (Ivanov et al., 2011). Recently, it was shown that the potency of tiRNAs to displace eIF4G can be predicted by stretches of guanines in the tiRNA since eIF4G has a propensity to bind G-quadraplexes (Lyons et al., 2020). The function of tiRNAs themselves as biologically active regulators of translation is supported by experiments showing that transfection of tiRNAs into unstressed cells is sufficient to induce the integrated stress response (ISR) (Yamasaki et al., 2009).
Circular RNAs Are Substrates for Translational Regulation Under Stress
Circular RNAs (circRNAs) are a large class of RNAs produced by the back-splicing of exons of precursor RNAs, typically ncRNA (Kristensen et al., 2019), in a wide range of cell types (Hansen et al., 2013; Hsu and Coca-Prados, 1979; Wu et al., 2012). circRNAs are generally less abundant than linear RNAs, but the absence of a 5′ and 3′ end makes them resistant to decay by exonucleases. Although many of the functions of circRNAs remain unknown, certain abundant circRNAs can sequester miRNAs and RNA-binding proteins, in a mechanism known as “sponging,” thus modulating their biological function (Zheng et al., 2016). Disruption of circRNA expression programs has been found to be associated with aging and many human diseases, such as Alzheimer's disease, diabetes, and cancer (Cortés-López et al., 2018; Gruner et al., 2016; Long et al., 2021; Verduci et al., 2021; Vo et al., 2019).
Emerging evidence suggests that circRNAs play a role in the regulation of translation of linear RNAs, although direct interaction between circRNAs and the translation machinery has not been discovered thus far. It is therefore probable that their influence on translation is indirectly mediated through miRNA “sponging” (Chekulaeva and Rajewsky, 2019; Lin et al., 2019; Prats et al., 2020). Typically, the RNA-induced silencing complex is guided by miRNAs to trigger deadenylation followed by mRNA degradation or translational repression of target mRNAs (Bartel, 2018). Disruption of the miRNA-mRNA interaction through miRNA sponging by an abundant circRNA reduces miRNA availability and limits its ability to regulate translation (Hansen et al., 2013; Yang et al., 2018; Zhang et al., 2019; Zheng et al., 2016).
Similarly, abundant circRNA can act as sponge for translation-associated RNA binding proteins (Fig. 4). For example, circPABPN1 interacts with HuR, to inhibit its binding and function as a translational activator of the poly(A)-binding protein nuclear 1 (PABPN1) mRNA (Abdelmohsen et al., 2017; Li et al., 2020).

Recent studies have revealed that circRNAs can also be direct substrates for translation themselves, mediated via IRESs. Following ISR activation, cap-dependent translation is reduced under stress (Pakos-Zebrucka et al., 2016). It is therefore reasonable to hypothesize that circRNA translation, mediated via IRES, is likely favored in such conditions. In fact, Chen et al. (2021) showed that a circRNA in the fibroblast growth factor receptor 1 (FGFR1) gene produces a protein that partially overlaps the protein product of the linear counterpart. Interestingly, under stress conditions, the ratio of circular over linear protein production increases, thus enabling the protein product of circFGFR1 to act as a dominant-negative regulator of FGFR1 and induce cell proliferation (Chen et al., 2021).
Combined with other recent evidence, these findings highlight a tight connection between the stress response and translation of circRNAs (Chen et al., 2019; Cheng et al., 2019; Feng et al., 2020; Hanan et al., 2020; Li et al., 2021; Zhang and Sui, 2020).
The increase in usage of IRES translation under stress implies a distinct regulatory mechanism for IRES-dependent translation of linear versus circRNAs. Past studies have identified that the N6-methyladenosine (m6A) reader YTHDF3 can recognize m6A in circRNA to directly recruit translation initiation factors (Yang et al., 2017). A recent study used a library of random 10-nt sequences inserted before the start codon of green fluorescent protein coded by a circRNA to identify sequences that can drive circRNA translation (Fan et al., 2022). In total, 97 six-nucleotide-long sequences that drive circRNA translation were identified. Most of these sequences were found to be A-U rich and preferentially enriched in endogenous circular rather than linear RNAs. Similarly, Chen et al. (2021) used a high-throughput reporter assay to systematically screen and quantify IRES elements that can facilitate circRNA translation.
Interestingly complementarity between the 18S rRNA and short stem-loops on the IRES was found to facilitate cap-independent translation, specifically for circRNAs. In contrast, the number of AUG codons, the Kozak sequence, and DRACH motif did not have a similar effect (Chen et al., 2021). These results provide evidence of a bipartite mechanism that regulates circular and linear RNA translation via IRES, but the exact details of the mechanism remain unknown. An intriguing hypothesis arises regarding the utilization of various components of the translation machinery by cells in response to stress. Typically, cap-dependent translation surpasses IRES-dependent translation in terms of protein output under normal conditions. However, during times of stress, cells have the ability to modify their proteome by downregulating cap-dependent translation and, simultaneously, sustaining or even enhancing IRES-dependent translation specifically for circRNAs (Fig. 4).
The 5′ UTR: Translation Selectivity During Stress
The critical role of the 5′ UTR in ribosome recruitment and start site selection allows it to introduce selectivity to translation, adding an extra layer of regulatory control beyond mRNA abundance. Mechanisms of selective translation are most often activated in response to acute stressors and are notably involved in the two major pathways of translation regulation: the mTORC1 and the ISR pathway.
The mTORC1 pathway is a sensor for numerous growth signals, such as the presence of amino acids, energy availability, and growth factors, that regulates translation using the mTORC1 serine/threonine protein kinase (Sengupta et al., 2010). During optimal growth conditions, the mTORC1 pathway acts to increase protein synthesis through phosphorylation of two proteins with divergent regulatory roles: the inhibitory protein eIF4EBP1 and the protein kinase S6K. eIF4EBP1 negatively regulates translation by binding and sequestering eIF4E from the eIF4F cap-binding complex, thereby reducing translation initiation; in contrast, S6K is a protranslational kinase that phosphorylates components of the translation machinery to increase protein synthesis (Saxton and Sabatini, 2017). mTORC1 phosphorylation of eIF4EBP1 suppresses its binding to eIF4E (Gingras et al., 2001), while phosphorylation of S6K enhances its function as a kinase (Holz et al., 2005), and thus, both roles coordinate to increase protein synthesis (Fig. 5A).
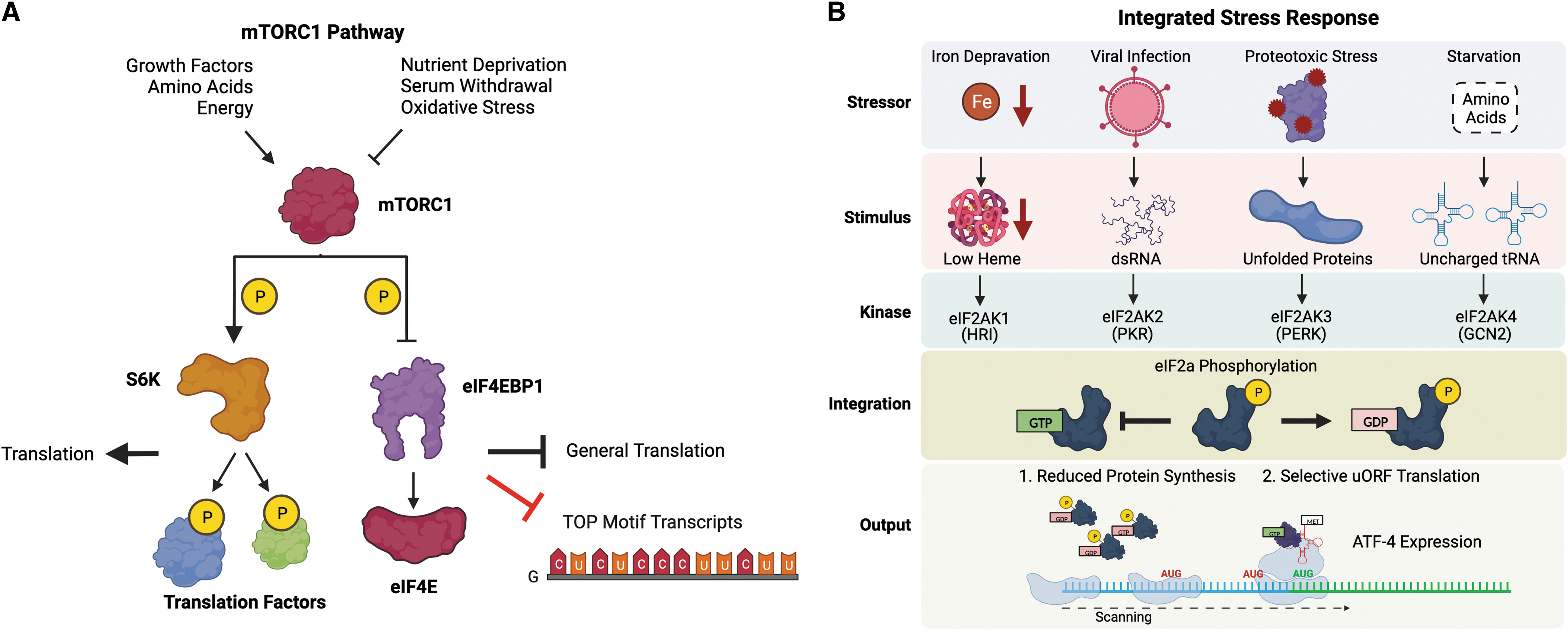
Although the mTORC1 pathway affects translation globally by activating S6K and suppressing eIF4EBP1, certain mRNAs are affected more than others. Experiments probing selective translation after mTORC1 inhibition identified a common 5′ terminal oligopyrimidine (TOP) motif among the most sensitive transcripts; this selectivity is largely due to the inhibitory action of eIF4EBP1, since 5′ TOP motif RNAs are more sensitive to disruptions of the eIF4E-eIF4G1 components of the eIF4F cap-binding complex (Thoreen et al., 2012). Interestingly, 5′ TOP motifs are overwhelmingly found on mRNAs encoding proteins that function in translation, strongly suggesting they are part of a coordinated mechanism to reduce protein synthesis during nonoptimal growth conditions that suppress mTORC1 activity (Hsieh et al., 2012). This mechanism is utilized as a survival mechanism in cancer, in which increased oxidative stress broadly reduces the translation of ribosomal proteins, since their transcripts are sensitive to mTORC1 inhibition (Tang et al., 2016).
In contrast to the mTOR pathway, the ISR pathway primarily functions to decrease translation in response to a diverse array of stressors including viral invasion, endoplasmic reticulum stress, amino acid starvation, oxidative stress, and others (Pakos-Zebrucka et al., 2016). The ISR pathway regulates translation initiation at the formation of the ternary complex (eIF2-GTP-tRNAiMet) necessary for start site selection by phosphorylating the eIF2α subunit of the eIF2 initiation factor. eIF2α phosphorylation is mediated through the activity of one of four kinases (eIF2AK1–4), commonly referred to as HRI, PKR, PERK, and GCN2 (Fig. 5B) (Donnelly et al., 2013). Due to their highly reactive nature, ROS can induce several different types of damage that will be recognized by the ISR.
In fact, HRI (Ill-Raga et al., 2015; Koncha et al., 2021; Suragani et al., 2012; Szwed et al., 2019), PERK (Farrukh et al., 2014; Harding et al., 2003; Verfaillie et al., 2013), GCN2 (Baker et al., 2012; Zhu et al., 2021), and PKR (Li et al., 2010; Zeng et al., 2022) have all been associated with activation of the ISR in response to oxidative stress. Phosphorylation of eIF2α globally reduces translation initiation, thus limiting protein expression. This reduction in protein expression also produces a selective increase in translation for certain transcripts with uORFs in their 5′ UTR, the most important of which is the multipurpose transcription factor activating transcription factor 4 (ATF4). Mechanistically, the decrease in ternary complex due to eIF2α phosphorylation increases the scanning time of ribosomes, allowing them to bypass an inhibitory uORF in the 5′ UTR of ATF4 (Young and Wek, 2016).
The use of translation selectivity by 5′ uORFs during oxidative stress extends beyond ATF4 as well (Andreev et al., 2015). Activation of the ISR during oxidative stress is also very important, as experiments show that disruption of the ISR increases sensitivity to ROS (Krishnamoorthy et al., 2018; Malin et al., 2021; McEwen et al., 2005; Rajesh et al., 2015).
The 3′ UTR: Selenocysteine Insertion Under Stress
Selenoproteins serve antioxidant roles and require noncanonical insertion of a selenocysteine residue at UGA stop codons. This event is mediated by a structured RNA element in the 3′ UTR called the selenocysteine insertion sequence (SECIS) that is expected to scaffold recoding factors, which ultimately enable insertion of selenocysteine at UGA stop codons (Vindry et al., 2018). The SECIS site is necessary for UGA recoding of selenoproteins, as well as sufficient to induce recoding when inserted into nonselenocysteine mRNAs (Shen et al., 1993). During oxidative stress, the recoding factors SBP2, L30, and EFsec are imported into the nucleus where they are recruited to the SECIS in selenoprotein mRNAs before their export into the cytoplasm for translation (Papp et al., 2006; Zahia et al., 2014). Thus, despite its noncoding nature, the 3′ UTR can function to dramatically alter translation during oxidative stress.
Epitranscriptomic Control of Translation Under Stress
Chemical modifications of RNA nucleotides were first discovered in 1957 (Davis and Allen, 1957) and currently, more than 330 modifications have been described (Boccaletto et al., 2022). Most of them localize on ncRNAs such as tRNAs, rRNAs, snRNAs, snoRNAs, and to a lesser extent on mRNAs (Boccaletto et al., 2022). Addition, removal, and sensing of chemical moieties on RNA bases are mediated by proteins known as writers, erasers, and readers, respectively (Esteve-Puig et al., 2020; Kumar and Mohapatra, 2021; Shi et al., 2019; Xuan et al., 2018).
RNA modifications can be reversible (e.g., m6A, 5-methylcytosine) or irreversible (e.g., A to I or C to U), add functional complexity in cellular regulation (Destefanis et al., 2021; Mathlin et al., 2020), and affect a range of cellular functions such as the immune response, cell death, DNA damage repair, stress response, and protein translation (Frye et al., 2018; Moradian et al., 2022; Wilkinson et al., 2022; Wilkinson et al., 2021).
m6A is among the most common reversible modification in mRNAs and has been involved in cellular response to oxidative stress and pathologies such as cancer, ischemic stroke, neurodegeneration, and aging (Chang et al., 2022; Condic et al., 2022; Desrosiers et al., 1974; Nunomura et al., 2012; Sun et al., 2022; Wu et al., 2022; Yang and Chen, 2021; Zhang et al., 2022a; Zhao et al., 2019). Recent reports have shown that m6A deposition on 5′ UTRs in response to oxidative stress provides a selective mechanism for sequestering mRNAs to stress granules, membraneless organelles formed upon stress, mediated by YTHDF3 (Anders et al., 2018). In addition, under hypoxic conditions, the m6A content increases for a subset of mRNAs and results in increased mRNA stability (Fry et al., 2017).
N4-acetylcytidine (ac4C), a reversible posttranscriptional RNA modification, is catalyzed by the N-acetyltransferase 10 and has recently been shown to be involved in position-dependent regulation of mRNA translation. While ac4C is primarily found in the 5′ UTR, in close proximity to the start codon, ac4C deposited within the coding region can promote translational efficiency by facilitating decoding of wobble sites and thus preventing mRNA decay (Arango et al., 2018). In contrast, ac4C deposited in the 5′ UTR impacts translation by promoting initiation at upstream sequences, thus competitively reducing initiation at canonical ORFs. Furthermore, acA4C within the Kozak sequence and immediately upstream of optimal start codons can disrupt an interaction between the modified C and the initiator tRNA, thus further inhibiting initiation (Arango et al., 2022).
These studies highlight the role of ac4C in modulating translation and RNA stability and raise the possibility that these processes are closely regulated in cells and can be hijacked by pathogens or be deregulated in disease. In fact, it was recently shown that HIV-1 RNA is modified with ac4C at multiple discrete sites that result in increased stability and thus viral gene expression (Tsai et al., 2020). Similarly, ac4C deposited within the 5′ UTR of enterovirus 71 enhances viral RNA translation via selective recruitment of PCBP2 to the viral IRES and boosts its genome stability. ac4C has also been shown to stabilize and increase translation of oncogenes, thus linking the modification to cancer (Feng et al., 2022; Wang et al., 2022a; Zhang et al., 2021).
Pseudouridine (Ψ) is typically deposited in rRNAs and tRNAs and to a lesser extent mRNAs (Cui et al., 2021; Guzzi et al., 2018; Jack et al., 2011; McMahon et al., 2019). Nevertheless, recent evidence show that deposition of Ψ on mRNAs is tightly regulated during stress and is implicated with human disease (Barbieri and Kouzarides, 2020; Cerneckis et al., 2022). Interestingly, while oxidative and heat shock stresses increase pseudouridylation, starvation modestly decreases it (Begik et al., 2021; Li et al., 2015).
The exact regulatory role of Ψ under stress remains unknown, but deletion of PUS7, a pseudouridine synthase, in yeast resulted in reduced mRNA stability under stress suggesting a possible role of Ψ in RNA protection (Schwartz et al., 2014). Bisulfite-induced deletion sequencing in human cells confirmed that TRUB1-deposited Ψ sites led to transcript stabilization and ribosome read through at Ψ sites within stop codons (Dai et al., 2023; Fernandez et al., 2013).
N1-methyladenosine (m1A), besides its role in tRNAs, is also found in the 5′ UTR of mRNAs close to the transcription start site. It is dynamically regulated under different stress conditions and has been associated with translation efficiency and initiation (Li et al., 2017; Li et al., 2016; Safra et al., 2017). Despite recent progress, more research is required to discover molecular mechanisms through which mRNA modifications regulate cell processes under stress (Fig. 6).

Besides mRNA, RNA modifications are found in ncRNAs such as rRNA and tRNA (Roundtree et al., 2017). The rRNA epitranscriptome is reprogrammed in response to the cellular environment as well as in development and disease (Sloan et al., 2017; Xue and Barna, 2012). 2′-O-methylation of rRNA defines the ribosome conformational status and is essential for its biogenesis and function (Khoshnevis et al., 2022; Natchiar et al., 2018). Also, several rRNA modifications are clustered in the peptidyl transferase center and other functional sites of the ribosome to promote translational fidelity (Sloan et al., 2017). In fact, single-molecule RNA modification profiling revealed that the eukaryotic rRNA is modified at more than 100 sites, particularly at highly conserved and functionally relevant nucleotides (Bailey et al., 2022).
Since cap-independent translation becomes more prominent under stress (Spriggs et al., 2008), the efficiency of these sites for ribosome recruitment and initiation is substantially influenced by modifications both on rRNA and on translation substrates such as mRNAs, circRNAs, or lincRNAs (Coots et al., 2017; Meyer et al., 2015; Qin et al., 2022; Yoon et al., 2006). Oxidative stress, in particular, results in the creation of abasic sites, strand breaks, and 8-oxo-7,8-dihydroguanine modifications on rRNAs and mRNAs that substantially alter their function (Shcherbik and Pestov, 2019).
Modifications on tRNA are also dynamically regulated under stress. Recent evidence shows that wobble modifications in tRNAs lead to selective translation of stress response proteins from codon-biased genes (Endres et al., 2015; Huber et al., 2022). Reprogramming of the epitranscriptome is reflected in changes in translation and its deregulation is involved in disease (Bednarova et al., 2017; Chujo and Tomizawa, 2021; Suzuki, 2021; Torres et al., 2014).
The role of the epitranscriptome in translation has not been fully appreciated until recently, partly due to the lack of scalable methods to probe modified nucleotides. Established technologies for the detection of modifications can generally be divided into quantification methods, locus-specific detection methods, and next-generation sequencing-based detection methods (reviewed in Zhang et al., 2022b). While the latter methods constitute a major improvement, they are limited by read length and the inability to probe multiple modifications simultaneously at single RNA molecules. Nanopore direct RNA sequencing is emerging as a technology that can bridge that gap and provide valuable modification information for distinct RNA classes, including mRNAs, tRNAs, and rRNAs (Gu et al., 2012; Wang et al., 2021; Zhang et al., 2020). However, while the information for modified nucleotides is likely contained in the raw sequencer output, the software to extract the information is currently under development (Table 1).
Software for Detection of Ribonucleic Acid Modifications from Nanopore Direct Ribonucleic Acid Sequencing Data
m6A, N6-methyladenosine.
Conclusion and Perspectives
In this review, we have discussed the role of ncRNA elements in translation during cellular stress. While translation has been studied extensively, much remains to be deciphered, particularly regarding the role of ncRNAs. Ribosome biogenesis and the maturation of rRNA continue to be a highly complex area of study with numerous idiosyncrasies between organisms and have now been further shown to change in a directed way during stress. Recent research has shown extensive contributions of the 5′ UTR to translation efficiency, in particular the realization that uORFs are far more abundant than previously thought and may mediate selective translational mechanisms not previously known. Similarly, tiRNAs generated during stress from tRNAs have gained increased recognition as important signaling molecules during stress, and the diversity of these molecules is further enhanced by the enormous diversity within tRNA sequences.
The identification of circRNAs has also opened new possibilities of decay-resistant RNAs that can be induced by stress to either tune the translation machinery by binding specific RNAs and proteins or be translated themselves through IRES sequences.
The epitranscriptome offers an added level of complexity to translation regulation, with RNA modifications having increased utility on mRNAs, rRNAs, and tRNAs. In particular, ac4C and m6A have recently been found to play roles in the regulation of translation for both mRNAs and circRNAs. The study of RNA modifications at scale has only recently become feasible with the development of new high-throughput, small-read sequencing approaches that reveal genome- and transcriptome-wide maps of modified nucleotides. Nevertheless, while these approaches provide an unprecedented view into the world of RNA modifications, they cannot capture long-range interactions or the combinatorial effect of different modifications on the same RNA molecule. Intriguingly, new technologies using nanopore sequencing are emerging as potentially revolutionary approaches to sequence RNA directly and deconvolve modified nucleotides and other RNA features at the single-molecule level.
Future studies will need to address current technology and software shortcomings to profile the complete RNome and expand upon the currently limited subset of identifiable modifications. This information will allow a transcriptome-wide exploration of the combinatorial role of RNA features, including modifications and 5′ UTR regulatory elements, on translation under stress.
Footnotes
Acknowledgments
We thank Dr. Myriam Gorospe for editing our article and providing feedback. We would also like to thank Yolanda L. Jones, NIH Library, for article editing assistance. Figures were created with BioRender.com
Author Disclosure Statement
No competing financial interests exist.
Funding Information
This work was supported by the Intramural Research Program of the National Institute on Aging, National Institutes of Health (ZIAAG000696).