
Abstract
Significance:
Pyroptosis is a discovered programmed cell death that is mainly executed by the gasdermin protein family. Cell swelling and membrane perforation are observed when pyroptosis occurs, and is accompanied by the liberation of cell contents.
Recent Advances:
As the study of pyroptosis continues to progress, there is increasing evidence that pyroptosis influences the development of tumors. In addition, the relationship between pyroptosis and tumor is diverse for different tissues and cells.
Critical Issues:
In this review, we first introduce the research history and molecular mechanisms of pyroptosis. Then we specifically discuss the link between pyroptosis and metabolic and oxidation in tumorigenesis. In the subsequent sections, we focus on the induction of pyroptosis in cancer and its potential role as a promising target for cancer therapy, and discuss the implications of pyroptosis in tumor treatment. In addition, we further summarize the therapeutic value of pyroptosis in tumor treatment.
Future Directions:
A detailed understanding of the role played by pyroptosis in tumors will help us to further explore tumor formation and progression and provide ideas for the development of new pyroptosis-based therapeutic approaches for patients. Antioxid. Redox Signal. 39, 512–530.
Introduction
Cancer has become the most important public health burden due to its high morbidity and mortality rate (Torre et al., 2016). Although diagnostic and therapeutic aspects have been optimized, the survival rate of cancer patients remains unsatisfactory. Cell death plays a crucial role in regulating homeostasis and development in organisms (Galluzzi et al., 2018), and the induction of tumor cell death remains the primary strategy of cancer treatment. Several cell death pathways have been identified, including necrosis, apoptosis, autophagy, necrosis, ferroptosis, and pyroptosis, which are divided into two categories: accidental cell death and regulated cell death (RCD). As a form of RCD, pyroptosis is characterized by the formation of cell membrane pores, cytoplasmic vacuolation, and release of intracellular contents, such as interleukin (IL)-18 or IL-1β (Broz and Dixit, 2016; Fink and Cookson, 2007; Fink et al., 2008).
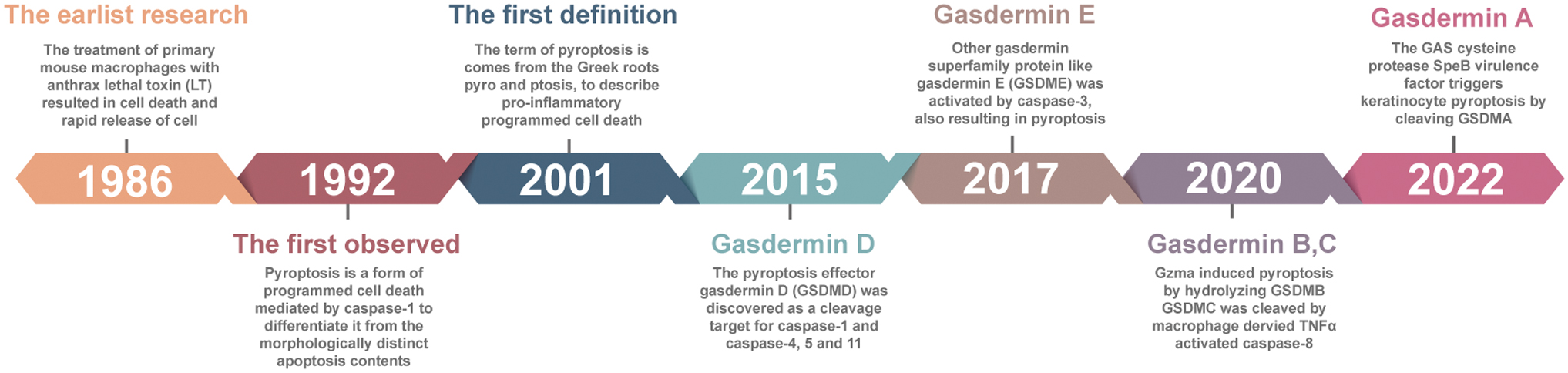
The earliest research on pyroptosis was reported in 1986. Friedlander (1986) showed that exposure to lethal toxin caused cell death of primary mouse mammalian cells, leading to the release of cell contents. Pyroptosis occurs due to caspase-1 activation, which was first observed in 1992, and this feature distinguished it from morphologically distinct apoptosis (Zychlinsky et al., 1992). In 1997, Hilbi et al. discovered that Shigella dysenteries activate caspase-1 in host cells (Hilbi et al., 1997). In 1999, a further study demonstrated that the inhibition of caspase-1 can prevent cell death from Salmonella (Hersh et al., 1999).
In 2001, D'Souza and Heitman (2001) coined the term “pyroptosis,” which is derived from the Greek roots”pyro” (fire/fever) and “ptosis” (to-sis, falling), to describe a proinflammatory cell death program. Since then, even though there have been many reports of pyroptosis, the effector of pyroptosis remained unknown until 2015, when gasdermin (GSDM) D (GSDMD) was discovered as a cleavage target for caspase-1, caspase-4, caspase-5, and caspase-11 (Kayagaki et al., 2015; Shi et al., 2015). More recently, accumulating evidence has shown that other proteins of the gasdermin superfamily, such as GSDME, are activated by caspase-3, resulting in the formation of pyroptosis (Wang et al., 2017). In 2020, it was reported that granzyme A (GZMA) in cytotoxic T lymphocytes (CTLs) can enter target cells by perforation and induce pyroptosis by hydrolysis of GSDMB at the Lys 229/Lys 244 site (Zhou et al., 2020).
In the same year, Hou et al. (2020) discovered that the activation of p-STAT3 in a hypoxic environment promotes the nuclear translocation of programmed cell death protein 1 (PD-L1). Nuclear PD-L1 and p-STAT3 promote the synergetic expression of GSDMC, while caspase-8 activated by macrophages can cleave GSDMC into N-GSDMC (Hou et al., 2020). More recently, Deng et al. (2022) indicated that the Group A Streptococcus (GAS) cysteine protease Streptococcal pyrogenic exotoxin B (SpeB) virulence factor triggers keratinocyte pyroptosis by cleaving GSDMA after Gln246, unleashing an active N-terminal fragment that initiates pyroptosis (Fig. 1). In the present review, we summarize the role and mechanisms of pyroptosis in cancer progression and briefly demonstrate how oxidative stress and metabolism trigger pyroptosis based on existing relevant studies, further affecting tumorigenesis and progression.
Molecular Mechanisms of Pyroptosis
In 2015, pyroptosis was defined as programmed cell death mediated by a gasdermin family member, and GSDMD was identified as the central executor of pyroptosis (Shi et al., 2015). In addition, there are five other members of the gasdermin family, such as GSDMA/B/C/D/E and pejvakin (PJVK), in humans. The mouse gasdermin family includes GSDMA, GSDMC, GSDMD, GSDME, and PJVK, but not GSDMB (Broz et al., 2020; Feng et al., 2018). Except for PJVK, all gasdermin proteins have two conserved domains, the N-terminal effector domain and the C-terminal inhibitory domain (Ding et al., 2016). In the presence of endogenous stimuli or various microbes, gasdermin is cleaved by active caspases or granzymes. The liberated N-terminal domain forms large pores on the membrane to release cell contents and execute pyroptosis (Table 1) (Aglietti and Dueber, 2017). Among the proteins described above, GSDMD and GSDME are the most thoroughly studied proteins in pyroptosis.
Gasdermin Cleavage by Various Proteases Triggers Pyroptosis
CTL, cytotoxic T lymphocyte; DAMPs, damage-associated molecular patterns; GAS, group A Streptococcus; GZMA, granzyme A; GZMB, granzyme B; LPS, lipopolysaccharide; NK, natural killer; PAMP, pathogen-associated molecular pattern; ROS, reactive oxygen species; SpeB, streptococcal pyrogenic exotoxin B; TNFα, tumor necrosis factor-alpha.
Currently, seven different pathways have been identified to induce pyroptosis. In general, pyroptosis is mainly executed via two pathways: inflammasome-dependent or inflammasome-independent. The inflammasome-dependent pathways are primarily classified as canonical and noncanonical, while the independent pathways contain caspase-3/-8-mediated pathways, granzyme-mediated pathways, and streptococcal pyrogenic exotoxin-mediated pathways (Fig. 2).

Canonical inflammasome pathway
Several inflammasomes have been identified in the past few years, and the composition of each inflammasome is determined by the activation of a unique pattern recognition receptor (PRR) in response to pathogen-associated molecular patterns or damage-associated molecular patterns (DAMPs) (Liston and Masters, 2017; Shi et al., 2014). The canonical inflammasome pathway-mediated pyroptosis mainly occurs in immune cells, serving as a host defense mechanism against pathogen infection. Activated caspase-1 cleaves the GSDMD protein and releases the N-terminal GSDMD (GSDMD-N) fragment, triggering pyroptosis through the nonselective pore of the cell membrane. In addition, activated caspase-1 directly stimulates the synthesis of IL-1β and IL-18, which can be released into the extracellular space through the pyroptosis pore (Bergsbaken et al., 2009).
Upon stimulation of PRRs, procaspase-1 is recruited directly by caspase recruitment domain (CARD)-carrying PRRs or indirectly via apoptosis-associated speck-like protein to assemble caspase-1-dependent inflammasomes, which is followed by caspase-1 activation through self-cleavage. Active caspase-1 not only cleaves inactive IL-1β and IL-18 precursors but also cleaves GSDMD to release GSDMD-N for pore-formation, eventually leading to inflammatory responses and pyroptosis (Gao et al., 2022; Miao et al., 2010). NLRP3 plays a critical role in the canonical inflammasome pathway (Arumugam et al., 2022; Hsu et al., 2022; Xu et al., 2022; Zhu et al., 2023).
Noncanonical inflammasome pathway
In the noncanonical pathway, accumulating evidence indicates that intracellular lipopolysaccharide from gram-negative bacteria is recognized by caspase-4/-5/-11 via the structural CARD, leading to activation of caspases, and then caspase-4/-5/-11 directly cleaves GSDMD into a C-terminal fragment and an N-terminal fragment (Shi et al., 2014). The N-terminal fragment forms a pore in the cell membrane, causing pyroptosis (Aglietti et al., 2016). Furthermore, activated caspase-4/-5/-11 does not mediate pro-IL-1β/pro-IL-18 maturation through direct cleavage, but indirectly regulates cytokine maturation and release by regulating the NLRP3/caspase-1 pathway (Shi et al., 2017). Notably, GSDMD-N also leads to K+ efflux and ultimately induces pyroptosis through a mechanism dependent on the activation of the NLRP3 inflammasome (Fernandez-Duran et al., 2022; Ruhl and Broz, 2015; Schmid-Burgk et al., 2015). Except for inflammatory caspase-1/-4/-5/-11, some apoptotic caspases can also trigger pyroptosis. For example, chemotherapeutic drugs can induce apoptosis mediated by caspase-3.
If target cells express GSDME, activated caspase-3 can lyse GSDME and induce pyroptosis, thus switching the cell death pattern. Wang et al. (2017) found that both cisplatin and other conventional chemotherapeutic drugs can cause pyroptosis by caspase-3-mediated cleavage of GSDME. In addition, caspase-8 cleaves GSDMC, releasing the N-terminal fragment of GSDMC and forming pores in the cancer cell membrane (Hou et al., 2020). Caspase-4 or caspase-6 also orchestrates the noncanonical inflammasome pathway (Fernandez-Duran et al., 2022; Flores et al., 2022; Hou et al., 2023; Li et al., 2021; Xu et al., 2021; Zheng et al., 2020).
Recently, it has been shown that granzymes derived from natural killer (NK) cells, CTLs, or chimeric antigen receptor (CAR)-T cells, transmitted to target cells via perforin, can cleave specific members of the gasdermin family and induce pyrolysis of cancer cells. For example, lymphocyte-derived GZMA cleaves GSDMB at the junction and liberates the cytotoxic N-terminal fragment, forming a pore in the membrane and leading to pyroptosis in GSDMB-expressing cells (Zhou et al., 2020). In 2020, Zhang et al. also demonstrated that granzyme B (GZMB) from CTLs can directly cleave GSDME to trigger caspase-3-mediated pyroptosis in GSDME-expressing cells (Zhang et al., 2020c). The cysteine protease SpeB virulence factor of GAS can cleave GSDMA, releasing an active N-terminal fragment that triggers the pyroptosis of keratinocytes (Deng et al., 2022).
Metabolism, Reactive Oxygen Species, and Pyroptosis
Metabolism and oxidation play a crucial role in the development and progression of cancer. These processes can contribute to cancer by disrupting cellular homeostasis, leading to oxidative stress and triggering the activation of cell death pathways (Miller et al., 2022; Morgan and Liu, 2011; Siegel et al., 2022). One such cell death pathway is pyroptosis, a highly inflammatory form of programmed cell death that plays a significant role in cancer. The interplay between metabolism, oxidation, and pyroptosis in cancer can be seen in various types of cancer and highlights the importance of understanding these processes in the development of new therapeutic strategies.
Cancer cells often exhibit a unique metabolic phenotype characterized by high glucose uptake and lactate production, which contributes to oxidative stress (Warburg, 1956). This metabolic reprogramming can lead to increased levels of reactive oxygen species (ROS) production, which in turn triggers the activation of pyroptotic cell death pathways. ROS can directly target cellular components, leading to damage and eventual cell death. For instance, oxidative stress can cause damage to mitochondrial DNA, leading to the release of mitochondrial DAMPs, which further activate the pyroptotic pathway (Krysko et al., 2012). In addition, oxidative stress can also activate caspase-1, a key effector of pyroptosis, leading to the release of proinflammatory cytokines such as IL-1β. This contributes to the inflammatory response seen in many cancers, which can facilitate cancer progression and tumorigenesis.
The activation of pyroptosis by oxidative stress can also be seen in pancreatic cancer, where increased ROS production has been shown to activate caspase-1 and trigger pyroptotic cell death (Fucikova et al., 2020). Metabolic reprogramming can also affect the availability of nutrients and energy, leading to cellular stress and triggering pyroptosis. For example, alterations in the availability of amino acids such as arginine and glutamine, which are essential for cancer cell survival, can lead to the activation of the pyroptotic pathway. This has been observed in various types of cancer, including prostate cancer and breast cancer. In prostate cancer, arginine depletion has been shown to trigger pyroptotic cell death and contribute to cancer progression. In breast cancer, alterations in glutamine metabolism have been linked to the activation of pyroptosis and the development of resistance to chemotherapy (Yan et al., 2021).
The interplay between metabolic and oxidative stress and pyroptosis can also be seen in melanoma, a type of skin cancer. Melanoma cells have been shown to exhibit increased glucose uptake and lactate production, leading to oxidative stress and the activation of pyroptotic cell death pathways. This has been linked to the development of resistance to chemotherapy in melanoma and highlights the importance of understanding the role of metabolic and oxidative stress in cancer progression (Baatarjav et al., 2022; Zhou et al., 2018).
Association between pyroptosis and metabolism
Cancer metabolism can be traced back to Otto Warburg, who observed that even under aerobic conditions, cancer tissue sections in vitro use large amounts of glucose to produce lactate compared with normal tissues, and this phenomenon is referred to as aerobic glycolysis or the Warburg effect (Koppenol et al., 2011; Warburg, 1956). However, many studies have demonstrated that most cancer cells require respiration to promote the tricarboxylic acid (TCA) cycle for tumor growth (Krall et al., 2021; Martínez-Reyes et al., 2020; Vasan et al., 2020). Moreover, some tumors need to use the TCA cycle to provide the necessary metabolites for their growth even though they do not respire (Cardaci et al., 2015). The Warburg effect of cancer cells was gradually understood until the resurgence of cancer metabolism (Vander Heiden et al., 2009).
Shim et al. (1997) have shown that the transcriptional target of the oncogene MYC, the glycolytic enzyme lactate dehydrogenase A, is required for increased glycolysis and tumorigenic potential of cancer cells. In addition, it has been demonstrated that protein kinase B (PKB)/AKT, Mammalian target of rapamycin (mTOR), and hypoxia-inducible factors, which are indispensable for tumor survival and growth, can increase glycolysis through the upregulation of glucose transporters and phosphorylation of glycolytic enzymes alone by transcription (Hoxhaj and Manning, 2020; Sabatini, 2017). Therefore, these studies reveal that both glycolysis and the TCA cycle can support tumor growth via the biosynthesis of metabolites (DeBerardinis and Chandel, 2020).
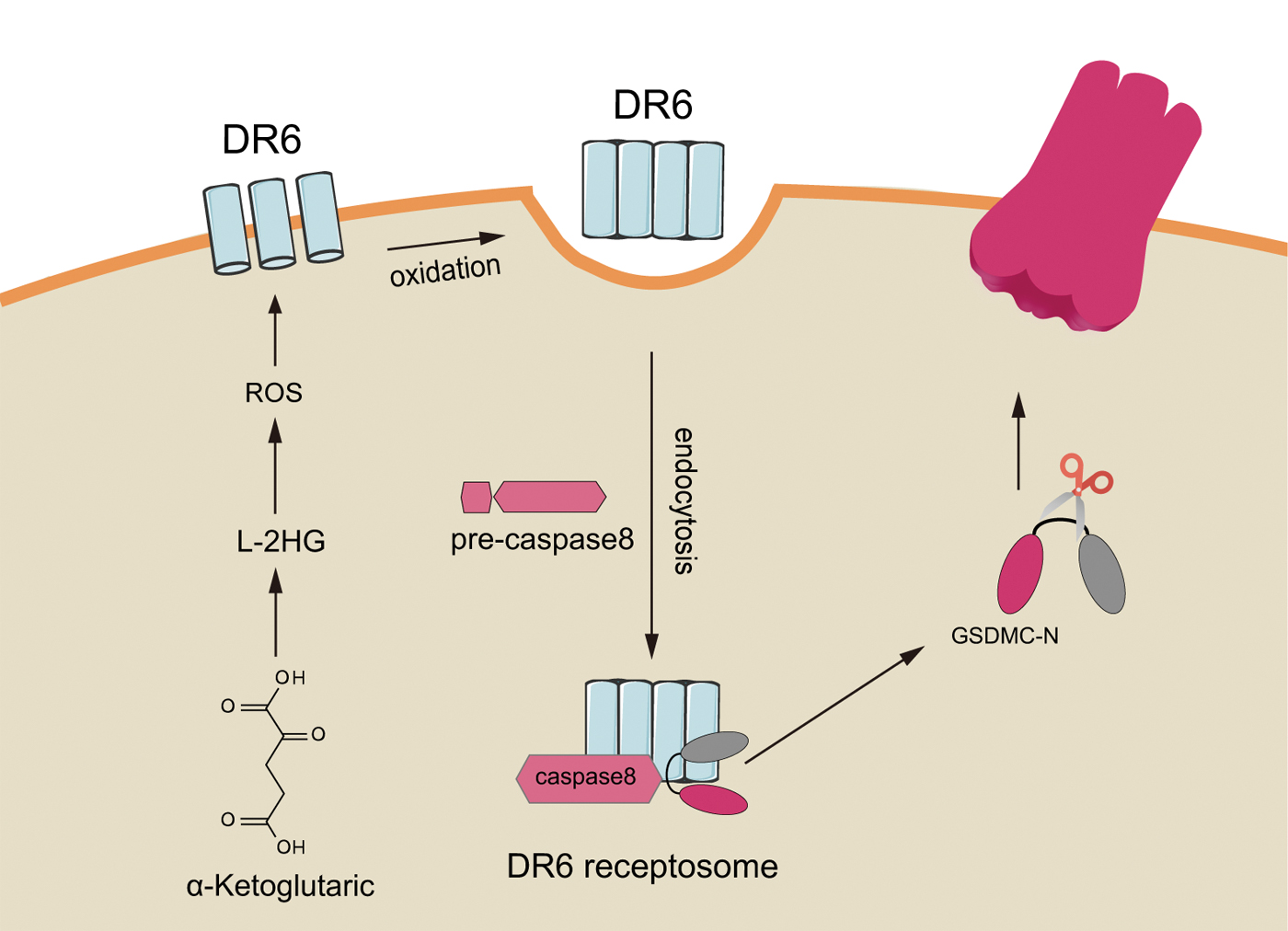
Alpha-ketoglutarate (α-KG) is an essential metabolite in the TCA cycle, playing a vital role in various physiological processes, including lipid biosynthesis, protein modification, and cell death. Morris et al. (2019) found that α-KG has some antitumor effects. For example, α-KG inhibits breast cancer by switching cellular respiration from glycolysis to oxidative phosphorylation. Zhang et al. (2021a) first showed the ability of the metabolite α-KG to trigger pyroptosis and explained the molecular mechanism in detail, confirming that pyroptosis induced by the metabolite α-KG also inhibits tumor growth and metastasis in mice.
In briefly, α-KG in an acidic environment is reduced to L-2HG by the metabolic enzyme MDH1 to further increase the concentration of intracellular ROS. The plasma membrane-localized death receptor DR6 is oxidized and internalized into the cell membrane, and internalized DR6 further recruits caspase-8 and GSDMC, allowing activated caspase-8 to cleave GSDMC and induce pyroptosis. Further studies have found that xenograft nude mice injected with dimethyl-α-ketoglutarate (a derivative of α-KG) have smaller tumors than controls (Zhang et al., 2021a). This recent study not only illustrates a metabolite-related pyroptosis pathway but also provides an important research direction for clinical oncology treatment through pyroptosis induction (Fig. 3).
Association between pyroptosis and ROS
ROS, also known as reactive oxygen intermediates, include highly reactive free oxygen radicals and stable nonradical oxidants. ROS are a product of normal cellular metabolism and are highly active in oxidation, accounting for ∼2%–3% of the oxygen consumption of the respiratory chain. When ROS levels are high in cells, they can act as destructive molecules, leading to cell death or tumorigenesis (Moloney and Cotter, 2018). In fact, it has been proven that cancer cells carry more ROS than normal cells (Szatrowski and Nathan, 1991). H2O2 from membranes and mitochondria can reversibly oxidize cysteine residues in proteins and control their activities by phosphorylation or other modifications (Tonks, 2005).
Further assays have revealed thousands of proteins for reversible oxidation, including those signaling pathways involved in mediators of cancer cell survival, proliferation, invasion, and metastasis (van der Reest et al., 2018; Weerapana et al., 2010). A deficiency of antioxidant stress also increases the occurrence of tumorigenesis. Many studies have demonstrated that mice with depletion of Prdx1, Sod1, and Sod2 have a higher risk of developing malignant tumors than normal mice (Elchuri et al., 2005; Rani et al., 2012; Van Remmen et al., 2003). NLRP3 is a very representative inflammasome that functions as a cellular stress sensor. The crystal structure of NLRP3 contains a highly conserved pyrin domain disulfide-linked structure and a nucleotide-binding site structure susceptible REDOX to, which mediates the activation of ROS (Hu et al., 2018).
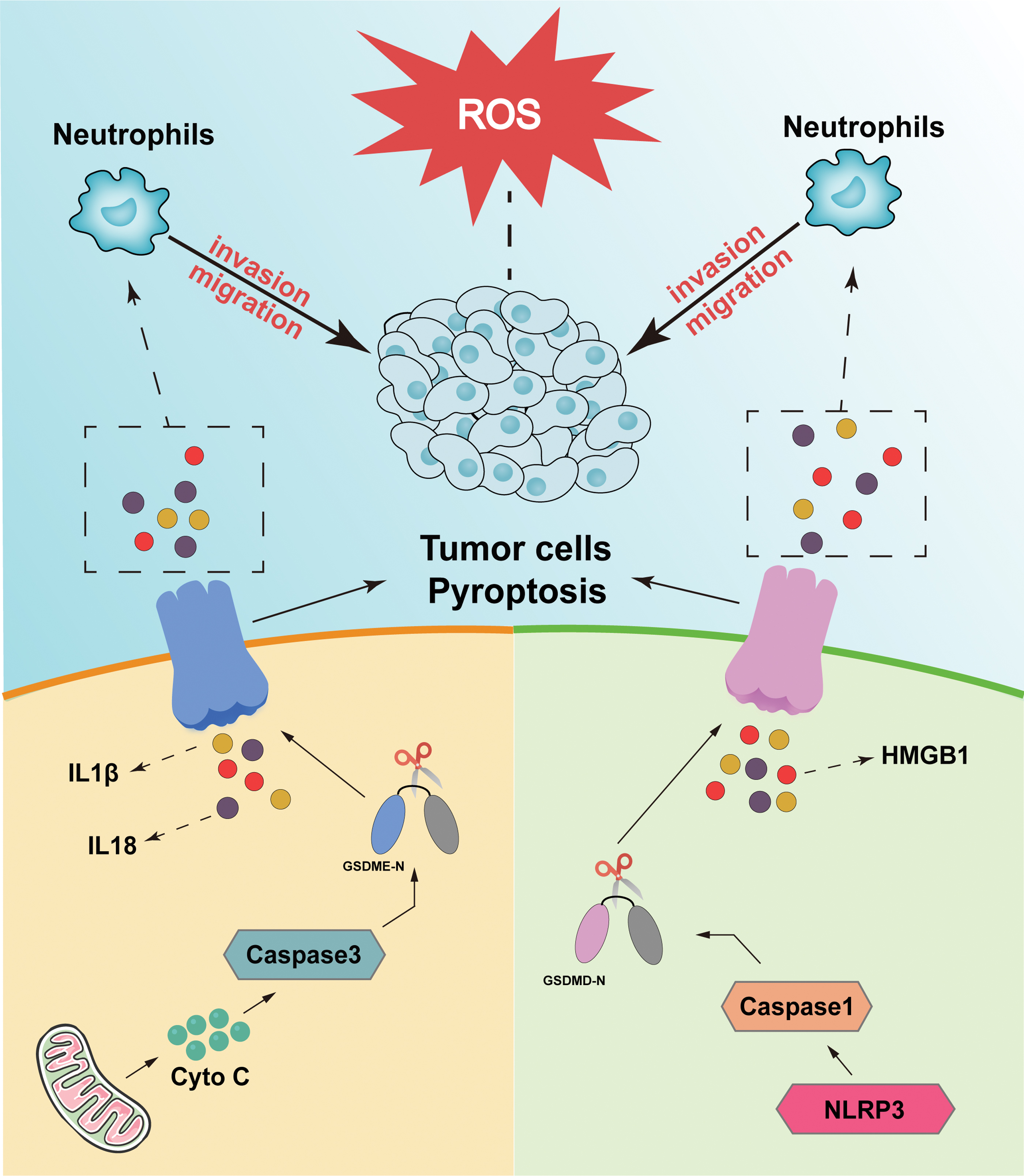
Excessive ROS in cells will cause an imbalance between oxidant and antioxidant responses, which can lead to oxidative stress. Such an imbalance is associated with the pathogenesis of several malignancies, including lung, breast, and liver cancers (Adcock et al., 2011; Sakaguchi et al., 2011; Vera-Ramirez et al., 2011). It has been reported that ROS are also associated with the occurrence of pyroptosis (Chen et al., 2020; Yang et al., 2020). In melanoma cells, ROS signaling is enhanced by iron ions and activates caspase-3 to cleave GSDME, resulting in pyroptosis (Zhou et al., 2018). Moreover, another study reported that MST1 partly inhibits the migration and invasion of pancreatic ductal adenocarcinoma cells through ROS-induced pyroptosis (Cui et al., 2019).
These results suggest that high levels of ROS exert their tumor suppressive effect, at least partly, through the induction of pyroptosis to inhibit the migration of cancer cells. On the contrary, ROS can affect the signaling pathway of NF-κB (Morgan and Liu, 2011). Furthermore, ROS can inhibit the NF-κB signaling pathway to trigger pyroptosis (Li et al., 2019). The effect of pyroptosis on cancer cells may be more complex, as IL-1β (Rebe and Ghiringhelli, 2020), IL-18 (Li et al., 2016), and high-mobility histone box protein 1 (HMGB1) (Jiao et al., 2021; Wu et al., 2020), inflammatory factors that are liberated after pyroptosis, can promote the invasion and migration of cancer cells in various ways (Fig. 4). Overall, the above studies suggest that ROS can influence the generation of pyroptosis and further affect cancer cell invasion and migration through pyroptosis. However, the detailed mechanism remains to be explored.
Interestingly, various natural drug components act as anticancer agents through ROS-mediated pyroptosis. For example, Teng et al. (2020) found that saponin polythene VI can play an anticancer role in nonsmall-cell lung cancer (NSCLC) by inducing caspase-1-mediated pyroptosis by triggering the ROS/NF-κB/NLRP3/GSDMD signaling axis. Furthermore, the anthraquinone derivative multitone isolated from the Salvia divinorum root can effectively induce ROS accumulation and then stimulate the BAX-caspase-3-GSDME pathway to trigger pyroptosis and induce hepatocellular carcinoma (HCC) cell death (Zhang et al., 2020b). On the contrary, many lead compounds, such as acetone analogs and perondine analogs, can trigger pyroptosis by modulating ROS, thus producing anticancer effects (Li et al., 2019; Zhu et al., 2018). In conclusion, the discovery of these drugs confirms that it may be feasible to regulate pyroptosis by modulating the degree of oxidative stress and provides new ideas and targets for future cancer therapy.
Role of Pyroptosis in Tumors
Pyroptosis has been implicated in a number of diseases, including cancer. In cancer cells, pyroptosis can either suppress or promote the progression of the disease, depending on the context and the stage of the cancer (Fang et al., 2020). For example, in early-stage cancers, pyroptosis can act as a tumor suppressor mechanism by eliminating abnormal cells. In later stage cancers, however, pyroptosis can contribute to tumor progression by promoting inflammation and tissue damage.
The molecular mechanisms of pyroptosis in cancer are complex and involve multiple signaling pathways and cell types. Key components of the pyroptosis pathway include inflammasomes, which are multiprotein complexes that sense cellular stress and activate caspases, as well as other signaling molecules such as cytokines and oxidative stress pathways (Feng et al., 2018). In terms of downstream effects, pyroptosis can have a significant impact on the tumor microenvironment, leading to increased inflammation and oxidative stress, as well as changes in the tumor vasculature and immune system. For example, pyroptosis can activate immune cells, such as macrophages and NK cells, and promote the recruitment of immune cells to the site of the tumor. This can result in increased inflammation and oxidative stress, further promoting cancer progression (Zhang et al., 2020c).
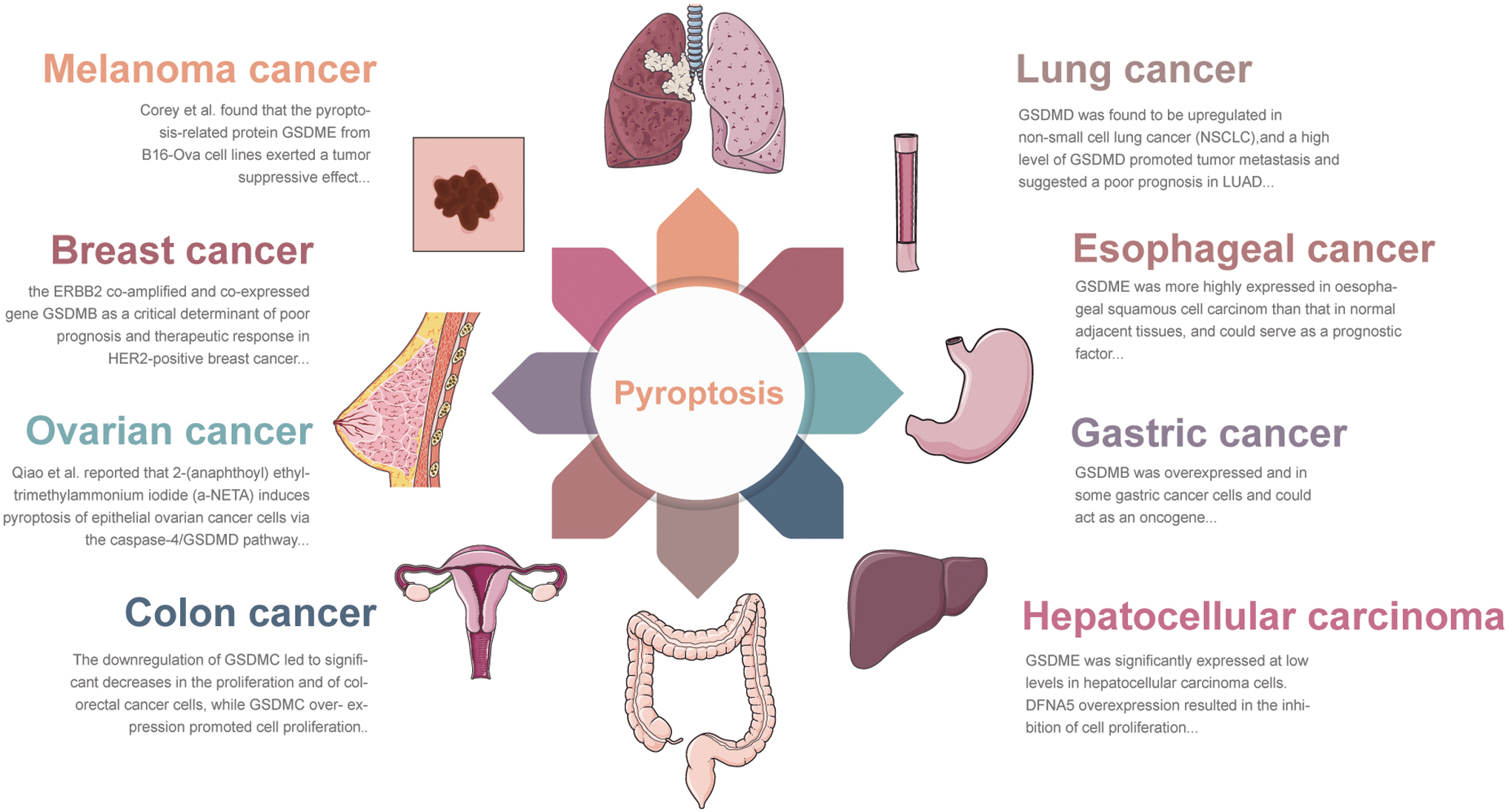
Pyroptosis may play a vital role in various tumors, and the mechanism of pyroptosis is different in cancer cells and immune cells. Pyroptosis plays a crucial role in tumor development and antitumor immunity, acting as a double-edged sword that can exhibit both tumor-promoting and tumor-suppressing functions. As research progresses, there is a growing understanding of the role played by pyroptosis in tumor and antitumor immunity, providing valuable insights into clinical treatment. Increasing evidence suggests that gasdermin cleaved by diverse caspases can trigger cancer cell pyroptosis (CCP) (Wang et al., 2020). Wang et al. (2017) reported that chemotherapy-induced activation of caspase-3 can trigger thermal denaturation by cleaving GSDME at the conserved site Asp270, leading to toxicity in normal tissues. Soon after, GSDME/caspase-3-mediated thermalization was reported in chemotherapeutic drug-treated gastric cancer (GC) cells (Wang et al., 2018b).
This finding suggests that gasdermin-mediated pyroptosis can occur not only in immune cells but also in healthy tissues and tumor cells. Since then, there has been a dramatic increase in research on gasdermin-induced cell death in cancer pyroptosis (Fig. 5).
Pyroptosis is a complex and dynamic process that has both positive and negative effects on cancer progression. While further research is needed to fully understand the molecular mechanisms and downstream effects of pyroptosis in cancer, it is clear that this process plays an important role in the development and progression of the disease. To effectively target and treat cancer progression, it is important to consider the role of pyroptosis and the interplay between this process and other cellular processes such as apoptosis, autophagy, and oxidative stress. This may involve developing new drugs or therapeutic strategies that specifically target the components of the pyroptosis pathway or manipulating the tumor microenvironment to promote or suppress this process as needed. In general, inflammasome-triggered pyroptosis mediated by caspase-1 and the release of pyroptosis-dependent cytokines, such as IL-1β and IL-18, can create a chronic inflammatory microenvironment.
This microenvironment may help tumor cells evade the immune response of host cells, leading to local immunosuppression and subversion of antigen-specific immune reactions and thus protecting tumor development.
The interaction between pyropotsis, metabolism, and oxidative stress in cancer
The interactions between pyroptosis, metabolism, and oxidative stress are critical in determining cancer progression in various types of cancer. For instance, several studies have demonstrated that pyroptosis plays a significant role in the development of colon cancer (Gan et al., 2020; Wu et al., 2019a). A study demonstrated that the activation of the inflammasome, a protein complex that triggers pyroptosis, can lead to the progression of colon cancer by promoting oxidative stress and cellular metabolism. In addition, the study showed that oxidative stress contributes to inflammasome activation by increasing the levels of ROS, which then results in the activation of the NLRP3 inflammasome and the subsequent progression of colon cancer.
Similarly, another research showed that pyroptosis plays a crucial role in the development of HCC, the most common type of liver cancer. The study found that the activation of the NLRP3 inflammasome can trigger pyroptosis and lead to the progression of HCC. In addition, a study demonstrated that pyroptosis contributes to oxidative stress and metabolic alterations in liver cancer cells, further contributing to the progression of HCC (Zhang et al., 2020b). Breast cancer is another type of cancer in which the interactions between pyroptosis, metabolism, and oxidative stress play a critical role. Knott et al. (2018) and Huang et al. (1999) found that pyroptosis can induce oxidative stress and alter cellular metabolism, leading to the progression of breast cancer. Moreover, they showed that pyroptosis-induced oxidative stress can activate specific signaling pathways that drive the progression of breast cancer.
In lung cancer, the activation of caspase-1 by oxidative stress has been linked to the initiation and progression of lung tumorigenesis (Gao et al., 2018). In prostate cancer, Cui et al. (2019) showed that oxidative stress caused by the buildup of ROS could lead to the activation of caspase-1 and the subsequent induction of pyroptosis. This study found that increased oxidative stress could contribute to the progression of prostate cancer by promoting cell death and inflammation (Cui et al., 2019). In glioblastoma, a type of brain cancer, a research showed that the activation of caspase-1 can lead to the induction of pyroptosis and contribute to the progression of glioblastoma. This study found that the upregulation of caspase-1 in glioblastoma cells was associated with increased oxidative stress and the release of proinflammatory cytokines, which could contribute to tumor growth and spread (Kim et al., 2017).
In conclusion, the interactions between pyroptosis, metabolism, and oxidative stress are critical in determining the progression of different types of cancer (Table 2) (Kim et al., 2017; Liu et al., 2019; Yuan et al., 2021). Further research is needed to fully understand the mechanisms by which pyroptosis, metabolism, and oxidative stress interact to drive cancer progression and to identify potential therapeutic targets. However, due to the heterogeneity of the different cancers, we discuss the role of pyroptosis in these different cancers in the next section.
Interaction Between Pyroptosis, Oxidative Stress, and Metabolism in Cancer
α-KG, alpha-ketoglutarate; DM-α-KG, dimethyl-α-ketoglutarate; ESCC, esophageal squamous cell carcinoma; GBM, glioblastoma; HMGB1, high-mobility histone box protein 1; NAC, N-acetylcysteine; PKM2, pyruvate kinase M2.
Pyroptosis and lung cancer
Lung cancer is one of the significant causes of death and the most common cancer globally (Hong et al., 2015; Villalobos and Wistuba, 2017). Chronic inflammation is one of the most critical factors causing the exacerbation of lung cancer (Siegel et al., 2019). GSDMD is considered a tumor-promoting gene in NSCLC (Gao et al., 2018). High expression of GSDMD is associated with larger tumor size, later tumor-node-metastasis stage, and lower survival rates (Gong et al., 2019). Moreover, high expression of GSDMD suggests a poorer prognosis in lung adenocarcinoma (LUAD), whereas this is not true for squamous cell carcinoma, indicating that GSDMD may serve as an independent prognostic biomarker for LUAD. More importantly, researchers have further demonstrated that in GSDMD-deficient NSCLC cells, activated NLRP3 inflammasome signaling triggers apoptosis but not thermal apoptosis (Gao et al., 2018).
Moreover, Zhang et al. (2019b) found that the tumor suppressor gene p53 directly interacts with NLPR3 to promote pyroptosis in NSCLC. Further study confirmed that GSDMD directly regulates EGFR/Akt signaling in NSCLC and promotes tumor proliferation by inhibiting apoptosis (Gao et al., 2018). In contrast to GDSMD-mediated pyroptosis, GSDME-mediated pyroptosis is usually triggered by chemotherapeutic agents. The classical chemotherapeutic agents 5-fluorouracil (5-FU), cisplatin, paclitaxel, and doxorubicin trigger pyroptosis in various cancer cells through caspase-3/GSDME action (Yu et al., 2019a; Yu et al., 2019b; Zhang et al., 2019a). Zhu et al. (2018) synthesized an analog of chalcone with antiproliferative effects against NCI-H460 cells. Its effects were time- and concentration-dependent, and it triggered caspase-3-mediated pyroptosis by regulating ROS. In addition, it had a superior safety profile when tested in vivo (Zhu et al., 2018).
These results suggested that cisplatin induced a higher level of pyroptosis than paclitaxel in lung cancer cells, indicating that cisplatin may provide an additional advantage in treating lung cancer with high GSDME expression.
Pyroptosis and esophageal cancer
Esophageal cancer is one of the most dangerous malignancies in the world because of its progressive dysphagia, esophageal stenosis, and high resistance to radiotherapy and chemotherapy, which often prevent it from achieving good treatment results (Gan et al., 2020). Metformin, the most widely used antidiabetic drug, has abundant evidence of anticancer properties (Janzer et al., 2014). In contrast, the exact mechanism underlying the anticancer effect of metformin remains incompletely understood. In a recent study, Wang et al. (2019) found that metformin induces GSDMD-mediated pyroptosis in esophageal squamous cell carcinoma (ESCC). Further mechanistic studies have shown that metformin activates ESCC pyroptosis by targeting the miR-497/proline, glutamate-, and leucine-rich protein-1 pathway (Wang et al., 2019).
These data reveal the exact mechanism of metformin-induced CCP, providing an opportunity to develop new drugs to control ESCC. Moreover, it has also been found that BI2536, an inhibitor of the ATP-binding domain of Polo-like kinase-1, in combination with cisplatin in esophageal cancer cells, enhances caspase-3 activity, causing pyroptosis and enhanced DNA damage. Wang et al. (2019) found that higher expression of GSDME in ESCC is related to a better prognosis, indicating that pyroptosis may soon significantly impact clinical practice (Wu et al., 2019b). Another study found that STAT3β increases chemosensitivity by activating caspase-3 and GSDME and ultimately triggers cellular pyroptosis in ESCC (Zheng et al., 2021). Other studies have found that the expression of GSDMC is inhibited in some esophageal cancer cell lines, suggesting its potential role as an antitumor agent (Saeki et al., 2009).
Pyroptosis and GC
GC is one of the most common malignancies. Its rising incidence has been classified as a significant health problem worldwide (Siegel et al., 2019). GC is a multifactorial disease characterized by a high recurrence rate, a poor prognosis, and a low overall survival rate (Hu et al., 2017). Notably, recent studies have revealed the role of pyroptosis in the pathogenesis of GC and chemotherapy, highlighting the clinical significance of low or no expression of the pyroptosis death executive protein gasdermin family in gene-targeting therapeutic approaches for malignant gastric diseases.
A recent study demonstrated that GSDMD is associated with GC, and its reduced expression has been shown to induce cancer cell proliferation. In vitro, the expression of GSDMD at the mRNA and protein levels was significantly downregulated in both GC cell lines and clinical GC tissues. In addition, the downregulation of GSDMD expression could activate signal transducers and activators of the STAT3 and PI3K/PKB signaling pathways and regulate cell cycle-related proteins, suggesting that the downregulation of GSDMD expression is beneficial for the treatment of GC (Wang et al., 2018a). In contrast, the potential role of GSDMD in GC still needs to be investigated in depth. GSDMD is believed to prevent the proliferation of GC cells.
Recent studies provide a new perspective suggesting that GSDMD can be a diagnostic marker and therapeutic target for GC. Treatment with chemotherapeutic agents, such as 5-FU, induces pyroptosis rather than apoptosis in GC cell lines expressing high levels of GSDME (Wang et al., 2018b). Pyroptosis is also caused by microbial infections (Bergsbaken et al., 2009; Frantz et al., 2003). Semper et al. (2014) reported that the production of IL-1β by H. pylori in innate immune cells was primarily associated with the activation of the NLRP3 inflammasome. High expression of NLRP3 in GC accelerated NLRP3 inflammasome activation and the liberation of macrophage IL-1β. Furthermore, H. pylori infection significantly promoted the expression of NLRP3 but reduced the expression of the NLRP3 inhibitor miR-22 (Chen et al., 2018b). In conclusion, the mechanism underlying activation of the NLRP3 inflammasome and its role in GC still need further clarification.
Pyroptosis and HCC
HCC accounts for ∼85%–90% of all primary liver cancers, with ∼841,000 new cases and 782,000 deaths annually. It is the second leading cause of cancer-related deaths worldwide, and its incidence is expected to further increase (Bray et al., 2018; De Toni et al., 2020). Berberine has been shown to inhibit the viability of HepG2 cells through caspase-1-mediated pyroptosis (Chu et al., 2016). Moreover, GSDME has been reported to be significantly expressed at low levels in HCC cells (Wang et al., 2013). Based on available statistics, men have a higher risk of developing HCC than women, and 17β-estradiol (E2) contributes to a large extent to this risk. Therefore, E2 and its receptors protect against HCC incidence (Yeh and Chen, 2010).
Wei et al. (2019) showed that E2 treatment induces the upregulation of caspase-1 and IL-1β in the HCC cell line HepG2, leading to NLRP3 inflammasome-mediated pyroptosis. Targeting the NLRP3 inflammasome may have an inhibitory effect on the proliferation, metastasis, and invasion of HCC, and NLRP3 may play an inhibitory role in cancer progression (Wei et al., 2019). Evidence has confirmed that the NLRP3 inflammasome is associated with liver failure and liver disease (Ganz et al., 2011; Kim and Lee, 2013). Hepatic cirrhosis or steatosis is strongly related to the development of HCC (Lee et al., 2019). Emerging evidence indicates that inflammasomes play a vital role in nonalcoholic fatty liver (NAFL), a series of metabolic disorders ranging from steatosis (NAFL) to nonalcoholic steatohepatitis to cirrhosis.
Another study evaluated the anticancer activity of cannabidiol (CBD) derived from cannabis in HCC. Shangguan et al. studied the effects of CBD on HCC cells (HepG2, HUH7, MHCC97H, and HCCLM3) (Shangguan et al., 2021). They found that CBD effectively inhibited the growth of HCC cells both in vitro and in vivo, and induced the pyroptosis of HCC cells in a caspase-3/GSDME-dependent pathway. These results suggest that CBD can inhibit glycolysis through the integrative stress response-dependent ATF4/IGFBP1/AKT/GSK3β axis. All the observations described here reveal a new mechanism of CBD antitumor activity in HCC cells and indicate that CBD can be considered a promising compound for the treatment of HCC. Furthermore, inflammasome activity has been reported to inhibit hepatic steatosis (Zhang et al., 2016). Activation of the inflammasome and endoplasmic reticulum stress-induced IL-1β production creates a positive feedback loop that amplifies the inflammatory response, leading to hepatic steatosis and injury.
Pyroptosis and colorectal cancer
A few decades ago, colorectal cancer (CRC) was not often diagnosed. However, CRC currently accounts for ∼10% of all cancers diagnosed and cancer-related deaths worldwide, and it is the fourth deadliest cancer in the world, killing nearly 900,000 people each year (Bray et al., 2018; Dekker et al., 2019). GSDMC has been reported to act as an oncogene that promotes tumorigenesis in CRC. Upregulation of GSDMC is accompanied by mutations in adenomatous polyposis coli and TGFBR2, and these characteristics predispose patients to develop CRC (Miguchi et al., 2016). Another report found that GSDME-mediated pyroptosis promotes CRC development by releasing HMGB1, inducing tumor cell proliferation, and enhancing the expression of proliferating nuclear antigens via the ERK1/2 pathway (Tan et al., 2020).
In addition, the development of CRC has been strongly associated with IL-18. Allen et al. (2010) found that injection of recombinant IL-18 into Caspase-1 KO mice can reduce disease progression or outcome in response to dextran sodium sulfate and azoxymethane. Therefore, local production of IL-18 by intestinal epithelial cells in the early stages may be beneficial for epithelial repair after injury. In contrast, the overproduction of IL-18 in the advanced stages of chronic inflammation may promote tumor development, suggesting that this pyroptosis-related cytokine can be a potential drug candidate for treating CRC. In another study, the inhibitory role of apoptin on the viability of HCT116 cells was explored. Apoptin was discovered to play a time-dependent role in inducing pyroptosis in cells. This was achieved through the cleavage of caspase-3 and GSDME instead of GSDMD.
The results showed that GSDME knockdown led to a switch in apoptin-induced cell death from pyroptosis to apoptosis in vitro. Furthermore, it was observed that the effect of apoptin on GSDME-dependent pyroptosis could be reduced by the knockdown of caspase-3 and caspase-9 by siRNA. In addition, apoptin was found to increase intracellular ROS, leading to the aggregation of the mitochondrial membrane protein Tom20. This, in turn, caused the release of bax and cytochrome c, triggering the activation of caspase-9 and ultimately resulting in pyroptosis. Therefore, it was concluded that GSDME mediates apoptin-induced pyroptosis through the mitochondrial apoptotic pathway. Based on these findings, apoptin may offer a new strategy for CRC therapy (Liu et al., 2022).
Pyroptosis and ovarian cancer
As a heterogeneous disease with genetic variations and complex molecular mechanisms, ovarian cancer (OC) is a common disease of the female reproductive system with extremely high recurrence and lethality rates (Siegel et al., 2019). Due to the absence of practical screening tools and credible markers for early diagnosis, most OC patients are already advanced at the time of diagnosis. In contrast, 50%–70% of patients experience relapse within just 2 years of treatment, and 5-year survival rates are as low as 30% (Kuroki and Guntupalli, 2020). Therefore, it is critical to understand the resistance mechanisms involved and identify potential targets for rational targeted therapy. Chemotherapy resistance is a primary challenge for OC patients. Recently, Qiao et al. (2019) found that a reversible choline acetylcholinesterase inhibitor, α-NETA, can inhibit the proliferation of epithelial OC cells both in vitro and in vivo through the induction of GSDMD/caspase-4-mediated pyroptosis.
Li et al. (2018) investigated the suppressive role of lncRNA GAS5 in OC and found that lncRNA GAS5 in OC was associated with the formation of inflammasomes and pyroptosis. The overexpression of lncRNA GAS5 was associated with the upregulation of caspase-1, which induced the liberation of IL-18 and IL-1β in a time-dependent manner, whereas the depletion of lncRNA GAS5 resulted in the downregulation of IL-18 and IL-1β (Li et al., 2018). These data suggest that lncRNA GAS5 in OC cells can arrest tumor progression and is a potential target for diagnosis and treatment in clinical applications. Tan et al. found that HOTTIP was upregulated in OC tissues. Consistently, HOTTIP expression was also significantly increased in OC cell lines (CAOV-3, A2780, OVCAR3, and SKOV3), especially in the A2780 and OVCAR3 cell lines. (Tan et al., 2021a).
An additional study discovered that chrysin, a phytochemical extracted from citrus fruits, effectively suppressed the growth of human ovarian cancer cells (HOCCs) and caused DNA damage. Further investigations revealed that chrysin induced the generation of ROS and autophagy in HOCCs by reducing the mitochondrial membrane potential, thereby promoting pyroptosis mediated by GSDMD and GSDME. Cinchonin, as a functional food component, holds great potential as a novel anti-OC agent that induces apoptosis and triggers ROS-mediated pyroptosis through modulation of autophagy in OC cells (Zhang et al., 2020a). These findings suggest that pyroptosis is a new strategy to fight cancer in treating epithelial OC.
Pyroptosis and breast cancer
Breast cancer is the most common cancer in women and the leading cause of cancer-related deaths in females. Although the 5-year relative survival rate of breast cancer has been improved due to the development of mammography, recurrent and metastatic breast cancer remains a significant challenge in clinical practice (Miller et al., 2022; Siegel et al., 2022). For this reason, it is critical to elucidate the molecular mechanisms of breast cancer and identify new effective prognostic biomarkers or therapeutic targets.
Some studies have proposed that GSDMB has critical functions, such as promoting metastasis and resistance, in HER2 breast cancer (Molina-Crespo et al., 2019). The activation of pyroptosis in breast cancer cells by docosahexaenoic acid (DHA) was investigated by Pizato et al. (2018). The experimental conditions involved the addition of DHA to MDA-MB-231 breast cancer cells, which resulted in cell death. Unlike untreated breast cancer cells, the DHA-treated cells exhibited heightened activation of caspase-1 and GSDMD, elevated secretion of IL-1β and HMGB1, and the formation of membrane pores in conjunction with lysosomes and ROS. These findings indicate that DHA elicits pyroptosis in breast cancer cells. Furthermore, the caspase-1 inhibitor (YVAD) protected breast cancer cells from the pyroptosis induced by DHA. This is the first study to demonstrate that DHA triggers pyroptosis in breast cancer cells, and provides valuable insight into the significance of incorporating omega-3 supplementation during breast cancer treatment (Pizato et al., 2018).
Interestingly, another study found that the expression of GSDME is increased in decitabine-treated tumor cells, which can enhance the effect of chemotherapy in breast tumors (Fan et al., 2019). However, GSDME is not consistently expressed in different breast cancer cell lines. For example, the expression of GSDME is higher in EMT6 TNBC, while its expression is lower in 4TIE TNBC (Zhang et al., 2020c). Yang et al. (2020) first reported that factors secreted from human umbilical cord mesenchymal stem cells significantly increase the expression of caspase-4 and NLRP1 in MCF7 cells, inducing MCF7 cell pyroptosis through the NLRP1-dependent canonical pathway and caspase-4-dependent noncanonical pathway (Jiao et al., 2020; Jiao et al., 2018). In addition, obesity-related activation of the IL-1a signaling pathway by the NLRC4 inflammasome promotes breast cancer progression, indicating that obesity enables breast cancer progression (Kolb et al., 2016).
Pyroptosis and melanoma cancer
Melanoma is considered one of the most aggressive human cancers (Lo and Fisher, 2014). Due to genetic or epigenetic alterations, melanoma cells are not very responsive to the induction of apoptosis (Gray-Schopfer et al., 2007; Soengas and Lowe, 2003). Therefore, it is crucial to develop new strategies for melanoma. Studies have revealed that GSDME-deficient melanoma cells can form larger tumors than wild-type cells (Rogers et al., 2019). Furthermore, a recent study also demonstrated a positive correlation between the expression of GSDME at the mRNA level and the chemosensitivity of melanoma cells (Lage et al., 2001). In contrast, GSDMC is rarely expressed in normal epithelial cells and is upregulated in malignant melanoma, which may be related to the invasion and metastasis of melanoma (Xia et al., 2019).
On the contrary, Yu et al. (2019b) demonstrated that inhibition of calmodulin-dependent protein kinase III (eukaryotic elongation factor-2 kinase, eEF-2K) in melanoma cell lines with high GSDME expression enhanced the efficacy of doxorubicin by increasing pyroptosis. Moreover, HMGB1 released by pyroptosis enhances melanoma cancer progression, and its upregulation is negatively correlated with melanoma survival (Ellerman et al., 2007). This finding also uncovers a potential iron-based therapeutic strategy for melanoma patients. Zhang et al. (2021a) established a B16 cell xenograft model in nude mice and discovered that intratumoral injection of 500 mg/kg DM-α-KG daily for 1 week caused a significant accumulation of ROS, activating DR6 and leading to a substantial reduction in xenograft tumor growth through a GSDMC-dependent pathway. Furthermore, the same treatment notably hindered the lung metastasis of B16 cells in mice. However, when GSDMC or DR6 was knocked down, the inhibitory effect of DM-α-KG on lung metastasis of B16 cells was lost.
To summarize, α-KG operates as an antitumor metabolite that restrains both tumor growth and metastasis through an ROS-DR6-GSDMC-mediated pyroptotic pathway (Zhang et al., 2021a).
Pyroptosis as a Target for Antitumor Immunotherapies
Current cancer immunotherapies are very successful in some patients. However, at the same time, they are ineffective in treating most patients. This fact suggests that further research is needed to promote antitumor immunity. Cancer cells, either spontaneously or in response to therapeutic intervention, have a tremendous impact on the type of immune response that is activated (Zhang et al., 2021b). Remarkably, the ability to induce an adaptive immune response, known as immunogenic cell death (ICD), is determined by their antigenic and adjuvant characteristics, such as both the presence of tumor-associated antigens and the liberation of endogenous DAMPs (Fucikova et al., 2020; Tan et al., 2021b).
Pyroptosis can cause a strong inflammatory response and is thought to be a form of ICD in some situations (Zhang et al., 2020c). Although the connection between pyroptosis and antitumor immunity is still under investigation, many studies suggest that pyroptosis can mediate tumor clearance through amplified immune activation. In addition, some immune cells can directly induce pyroptosis of tumor cells, indicating that pyroptosis may be involved in a positive feedback loop of antitumor immunity.
Wang et al. (2020) showed that successful delivery of the GSDMA protein isoform Gsdma3 into mouse 4T1 (mammary) cancer cells induced pyroptosis. Moreover, in BALB/c mice, an increase in CD3+ T cell infiltration and a decrease in CD4+FOXP3+ T regulatory cells were observed only in 4T1 tumors delivered with GSDMA3. More importantly, exhaustion of CD4+ and CD8+ cells prevented tumor regression in this treatment model, implying that CTLs and CD4+ T helper cells play an essential role in pyroptosis-induced tumor clearance (Wang et al., 2020). Xi et al. (2019) investigated the relationship between gasdermin gene expression in CTLs and LUAD, lung squamous carcinoma (LUSC), and melanoma tumor samples using data from The Cancer Genome Atlas.
Among these five gasdermin gene members, only the expression of GSDMD is positively correlated with CD8+ cell markers (such as CD8A, CD8B, GZMA, GZMB, and IFNG) in all three tumors (Xi et al., 2019). Upregulation of GSDMD is also observed in human-activated CD8+ T cells. Similarly, high levels of GSDMD protein are observed in tumor-infiltrating lymphocytes in LUAD and LUSC tissue samples (Xi et al., 2019) Subsequently, other studies have clarified the mechanism of NK- and CTL-induced tumor cell pyroptosis via the release of granzyme (Liu et al., 2020; Zhang et al., 2020c; Zhou et al., 2020). In contrast, Zhou et al. (2020) found that in the human embryonic kidney cell line, when 293T cells lacking endogenous expression of the gasdermin gene were forced to express only GSDMB but not the other gasdermin members, it led to the pyroptotic death of 293T cells from cocultured human NK cells (Fig. 6).
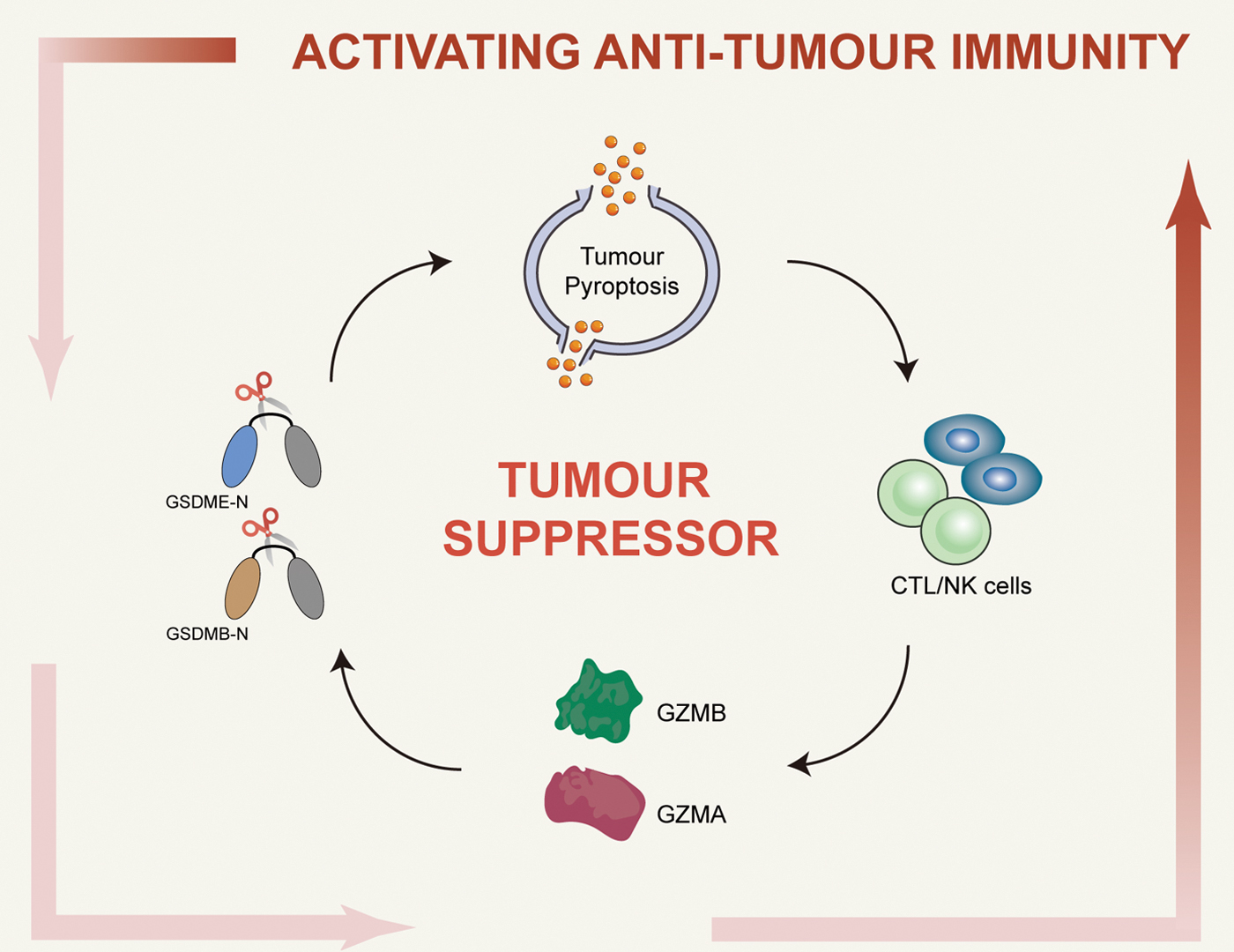
Interestingly, treatment with a pancaspase inhibitor had no effect, implying that GSDMB-mediated pyroptosis by NK cells is caspase-independent. However, inhibition of granzyme or NK cell perforin and degranulation blocked NK cell-induced pyroptosis and the cleavage of GSDMB in 293T cells. Further studies have revealed that among the five human granzymes, only GZMA can rapidly cleave GSDMB, which is similar to the model observed in NK cells (Zhou et al., 2020). The authors have also found that GSDMB cleavage and pyroptosis occured in 293 T cells expressing CD19 and GSDMB when incubated with human anti-CD19 CAR-T cells (Zhou et al., 2020). In conclusion, Zhou et al. (2020) demonstrated that GSDMB-mediated pyroptosis functions downstream of GZMA, and that CTLs can deliver GZMA to GSDMB-expressing cancer cells to promote antitumor immunity.
Furthermore, Zhang et al. (2020c) found that transplantation of mouse 4T1E breast cancer cells that ectopically express mouse GSDME into immunocompetent mice significantly inhibits tumor growth and leads to increased infiltration of tumor-associated macrophages and NK cells. Moreover, the antitumor effect of GSDME requires both lymphocytes and perforin and involves the involvement of NK and CD8+ T cells.
Further investigations have revealed that the NK cell line can activate the pyroptosis of GSDME-expressing HeLa cells via GZMB, which cleaves GSDME while indirectly activating caspase-3 (Zhang et al., 2020c). Similarly, Liu et al. (2020) found that CAR-T cells can induce GSDME-mediated tumor cell pyroptosis via the liberation of perforin and GZMB in B leukemia and solid tumor cells. In addition, ATP and HMGB1 in the supernatant of cocultured CD19 CAR-T cells and cancer cells were sufficient to promote the secretion of IL-1β and upregulation of IL-6 in macrophages. To a large extent, these findings are consistent with those seen in mouse models of leukemic CAR-T cell-induced cytokine release syndrome (CRS), suggesting that CAR-T cell treatment induces CRS through GSDME-mediated pyroptosis.
This finding was further supported when analyzing primary B leukemia cells from patients before CD19-CAR-T cell treatment. Higher levels of GSDME expression were correlated with more serious CRS (Liu et al., 2020). In a previous study, Erkes et al. (2020) found that pyroptosis of melanoma cells induced by GSDME and caspase-3 promotes the release of HMGB1, and its release activates dendritic cells, which in turn causes the proliferation and maturation of T cells and promotes antitumor immune responses. The above study shows that pyroptosis is a form of inflammatory death with an immune effect, providing a new perspective for cancer immunotherapy.
Furthermore, metabolism and oxidative stress can significantly affect pyroptosis of immune cells and play a crucial role in cancer progression. The interplay between pyroptosis, metabolism, and oxidative stress creates a complex network of pathways that contribute to the progression of cancer. A substantial number of compounds have been discovered to modulate pyroptosis through oxidative stress or metabolic pathways (Noguchi et al., 2021; Wu et al., 2019a; Zhu et al., 2018), ultimately enhancing the sensitivity of cancer cells to chemotherapy or elevating survival rates (Table 3). One example of this interplay can be seen in CRC, where oxidative stress and metabolic changes can activate the NLRP3 inflammasome, leading to pyroptosis of immune cells (Wu et al., 2019a).
Induction of Pyroptosis in Cancer by Various Therapeutic Compounds
IL, interleukin; PDAC, pancreatic ductal adenocarcinoma.
For instance, studies have shown that glucose metabolism and oxidative stress activate the NLRP3 inflammasome in macrophages, leading to the release of proinflammatory cytokines and pyroptosis. These events create a favorable microenvironment for the growth and progression of cancer cells (Chen et al., 2018a).
Therefore, inhibiting the interaction between metabolism and oxidative stress in pyroptosis could be a promising strategy for cancer treatment. By targeting specific metabolic enzymes such as NOX4 and mTOR, oxidative stress can be potentially reduced and pyroptosis can be induced, which in turn may prevent the progression of cancer. However, more research is needed to fully understand the mechanisms underlying the relationship and to develop more effective pyroptosis-inducing therapies. Currently, the majority of research on cancer and immune cell pyroptosis is focused on two types of pyroptosis, namely, GSDMD- and GSDME-mediated pyroptosis. This is because the discovery of GSDMD and GSDME occurred relatively early on, and thus, more research has been conducted on these two types.
However, as an increasing number of studies on GSDMA, GSDMB, and GSDMC are published, there is increasing evidence that these proteins mediate cancer cell death and immune cell pyroptosis. For instance, recent studies have shown that GSDMA and GSDMB play important roles in cancer cell death and that these types of pyroptosis may have different targets and therapeutic directions compared with GSDME- and GSDMD-mediated pyroptosis.
Although GSDME- and GSDMD-mediated pyroptosis has been widely studied, more research is needed to determine the extent to which other types of pyroptosis, namely GSDMA, GSDMB, and GSDMC, contribute to cancer cell death and immune cell pyroptosis. As the field advances, it is likely that our understanding of pyroptosis in cancer will continue to expand, leading to new and more effective therapies for the treatment of cancer.
Conclusion and Perspectives
As a proinflammatory form of cell death, the function and mechanism of pyroptosis have gradually become better understood. However, many questions still need further investigation, such as whether there are inhibitors or agonists of pyroptosis and whether other proteins of the gasdermin family play different roles in pyroptosis. Currently, increasing evidence has confirmed the vital role of pyroptosis in tumorigenesis. Although the dual mechanisms of tumor promotion and suppression by pyroptosis remain to be explored, it is undeniable that existing studies have shown the feasibility of combined pyroptosis and immune-based therapeutic approaches. Established tumors have a specific immunosuppressive microenvironment, which reduces T cell infiltration and limits the antitumor activity of CTLs. In contrast, induction of tumor cell pyroptosis has been shown to increase tumor immunogenicity and kill tumors by attracting more antitumor lymphocytes.
However, no studies have yet confirmed the involvement of the gasdermin family in tumorigenesis or as tumor suppressors. Further elucidation of the role of gasdermin in the tumor immune response is needed, which may also provide a strategy for the precision treatment of tumor immunity. Specifically, can gasdermin be considered a biomarker that classifies patients as being sensitive or insensitive to targeted drug treatment? Therefore, how to regulate the pyroptosis of tumor cells to achieve the desired therapeutic effect or how to avoid the pyroptosis of normal cells during tumor treatment still needs to be explored. In addition, NLRP3 inflammasome-mediated pyroptosis plays critical roles in the vasculature (Chi et al., 2022; Lai et al., 2022; Nurmohamed et al., 2022; Schunk et al., 2021), leading to the question of whether vascular pyroptosis affects tumorigenesis and turnover.
In fact, immune checkpoint inhibitors for tumor treatment also may trigger ROS and metabolism dysregulation, especially in the cardiovascular system (D'Souza et al., 2021a; D'Souza et al., 2021b; Dolladille et al., 2021; Kondapalli and Neilan, 2021; Michel et al., 2022; Thuny et al., 2022). Overall, it is foreseeable that research on pyroptosis and tumors will continue to improve and provide new ideas for clinical research.
The relationship between metabolism, oxidative stress, and pyroptosis in cancer is an area that has gained increased attention in recent years. There is a growing body of evidence indicating that targeting the metabolic and oxidative stress pathways may be a promising strategy for treating cancer. The concept of using metabolic and oxidative stress to target pyroptosis in cancer cells has been demonstrated in several studies, including those using drugs such as metformin and vitamin E to modulate the activity of key metabolic and oxidative stress pathways.
However, further research is needed to fully understand the underlying mechanisms involved in the relationship between metabolism, oxidative stress, and pyroptosis in cancer cells. For example, while some studies have found that drugs that modulate metabolism and oxidative stress can lead to increased pyroptosis in cancer cells, others have shown no such effect. In addition, while some cancer types may respond well to drugs that modulate metabolism and oxidative stress, others may not. Further research is needed to identify the specific metabolic and oxidative stress pathways involved in pyroptosis in different cancer types, and to determine the most effective strategies for targeting these pathways in a personalized manner.
In conclusion, the relationship between metabolism, oxidative stress, and pyroptosis in cancer is complex and multifaceted, and much more research is needed to fully understand this relationship and to develop effective strategies for treating cancer. However, existing evidence suggests that targeting the metabolic and oxidative stress pathways in cancer cells has the potential to be a promising strategy for cancer treatment, and could lead to improved outcomes for patients with cancer.
Footnotes
Authors' Contributions
C.Q., D.J.L., and P.W. designed and conceived the review. C.Q., J.B.Z., D.Y.S., J.T.F., W.B.W. X.F.C., D.J.L., and P.W. wrote the article. C.Q., D.J.L., and P.W. revised, extended, and provided the final proof of the article. All the authors reviewed the article.
Author Disclosure Statement
No competing financial interests exist.
Funding Information
This work was supported by grants from the National Natural Science Foundation of China (82073915, 81673485, 81773719, 81973312, and 81971306), the National Key Research and Development Project (2018YFA0108301), and the Shanghai Science and Technology Commission (21XD1424900).