
Abstract
Significance:
Peroxiredoxins (Prdxs) with a single peroxidative cysteine (CP) in a conserved motif PXXX(T/S)XXCP within its thioredoxin fold, have been classified as the peroxiredoxin 6 (Prdx6 ) family. All Prdxs can reduce H2O2 and short chain hydroperoxides while Prdx6 in addition, can reduce phospholipid hydroperoxides (PLOOH) due to its ability to interact with peroxidized phospholipid substrate. The single CP of Prdx6 uses various external electron donors including glutathione thioredoxin, and ascorbic acid for resolution of its peroxidized state and, therefore, its peroxidase activity. Prdx6 proteins also exhibit Ca2+-independent phospholipase A2 (PLA2), lysophosphatidylcholine acyltransferase (LPCAT), and chaperone activities that depend on cellular localization and the oxidation and oligomerisation states of the protein. Thus, Prdx6 is a “moonlighting” enzyme.
Recent Advance:
Physiologically, Prdx6s have been reported to play an important role in protection against oxidative stress, repair of peroxidized cell membranes, mammalian lung surfactant turnover, activation of some NADPH oxidases, the regulation of seed germination in plants, as an indicator of cellular levels of reactive O2 species through Nrf-Klf9 activation, and possibly in male fertility, regulation of cell death through ferroptosis, cancer metastasis, and oxidative stress-related signalling pathways.
Critical Issues:
This review outlines Prdx6 enzyme unique structural features and explores its wide range of physiological functions. Yet, existing structural data falls short of fully revealing all of human Prdx6 multifunctional roles. Further endeavour is required to bridge this gap in its understanding. Although there are wide variations in both the structure and function of Prdx6 family members in various organisms, all Prdx6 proteins show the unique a long C-terminal extension that is also seen in Prdx1, but not in other Prdxs.
Future Directions:
As research data continues to accumulate, the potential for detailed insights into the role of C-terminal of Prdx6 in its oligomerisation and activities. There is a need for thorough exploration of structural characteristics of the various biological functions. Additionally, uncovering the interacting partners of Prdx6 and understanding its involvement in signalling pathways will significantly contribute to a more profound comprehension of its role.
Introduction
Peroxiredoxin (Prdx) proteins are ancestral thiol-dependent but selenium- and heme-free peroxidases that are found ubiquitously throughout evolution in plants, bacteria, archaea, and eukaryotes (Rhee et al., 2005; Wood et al., 2003). They evolved from other peroxidases by replacing the prosthetic group with a conserved N-terminal peroxidatic cysteine (CP) residue acting as the site of oxidation by peroxides (Rouhier et al., 2001).
An additional conserved resolving cysteine residue (CR) may be present in the C-terminal region of Prdx protein. Prdx proteins found in both lower organisms, including archaebacteria, cyanobacteria, and prokaryotes, as well as in higher organisms and plants were originally categorized into three classes depending on the presence or absence of CR and its involvement in peroxidation: these classes were typical 2-Cys Prdx with CR found at the C-terminus to form an inter-subunit disulfide on oxidation; atypical 2-Cys Prdx with CR found at positions in the protein other than the C-terminus to form an intra-subunit disulfide upon oxidation; and 1-Cys Prdx with no CR present but requiring a thiol from another protein or molecule to complete the recycling process (Cui et al., 2012; Dietz, 2011; Fujii and Ikeda, 2002; Nelson et al., 2011; Rhee et al., 2005; Seo et al., 2000; Wood et al., 2003).
Under this classification, mammalian cells express six different Prdx proteins, that is, typical 2-Cys Prdx called Prdxs 1–4 (CR at the C-terminus), atypical 2-Cys Prdx called Prdx5 (CR elsewhere in the protein than the C-terminus), and 1-Cys Prdx (no CR present) called Prdx6. But the classification into either 1-Cys or 2-Cys Prdx was found to be inadequate as the existence and the location of the resolving cysteine can vary to a significant degree when taking both lower and higher organisms into account and some peroxidase proteins including AhpC/Prdx1 and Prdx6 exist within all the subfamilies (Nelson et al., 2011).
Use of the bioinformatic tool Deacon Active Site Profiling based on sequence comparisons among fragments plucked from a specific region of key active site residues in Prdxs of known structure has resulted in a more recent re-classification of Prdx, also into six subfamilies: (1) AhpC/Prdx1, (2) Prdx6, (3) Prdx5, (4) Tpx, (5) bacterioferritin co-migratory protein (BCP)/PrdxQ, and (6) alkyl hydroxyperoxide reductase E (AhpE) (Perkins et al., 2015; Perkins et al., 2016; Soito et al., 2011).
Prdx6 shows widespread presence in organisms of different kingdoms from lower prokaryotes to plants and higher mammals (Dietz, 2011; Loumaye et al., 2008). The presence of the conserved peroxidatic Cp in the motif PVCTTE is characteristic of all Prdx6 enzymes (Nevalainen, 2010). In the dimeric state at neutral pH, Prdx6 peroxidase activity can reduce hydrogen peroxide, peroxynitrite, and hydroperoxides (Fisher, 2011; Nevalainen, 2010; Pedrajas et al., 2000).
Besides its peroxidase activity, Prdx6 also shows phospholipase A2 (PLA2) and lysophosphatidylcholine acyltransferase (LPCAT) activities. PLA2 activity releases the fatty acyl group from the sn-2 position of a phospholipid, and LPCAT activity catalyzes the transfer of a fatty acyl CoA into the sn-2 position of lysophosphatidylcholine (LPC) to generate a phospholipid (Fisher, 2018).
Thus, these two reactions can combine within the same enzyme to serve a phospholipid remodeling function. Some members of the Prdx6 family in organisms such as the thermophilic archaeon have one or more cysteine residues besides Cp and show chaperone activity, in addition to their peroxidase activity (Lee et al., 2015); however, as these additional cysteines in Prdx6 do not function in the resolution of the oxidized protein, they are not a CR.
This review is focused on the variations in the primary and three-dimensional structures of Prdx6 family members, the reductants involved in their catalytic activity, their oligomeric state at different organismal levels, and their various physiologic and pathophysiologic activities and functions. It presents our current understanding of the Prdx6 family with primary reference to the structural and biophysical ramifications for its multi-functional properties.
The major headings used in this review of Prdx6 are: (1) primary properties of Prdx6 proteins; (2) structural features of Prdx6s; (3) mechanisms for oxidation-reduction of Prdx6 proteins; (4) oligomerization state of Prdx6 proteins; and (5) multi-functional activities of Prdx6.
Primary Properties of PRDX6 Proteins
Prdx6 motives for catalysis
Peroxidase activity
The alignment of the primary and secondary structures of eukaryotic Prdx1, Prdx2, Prdx3, Prdx4, and Prdx6 is depicted in Figure 1. Prdx6 family members have a unique catalytic motif, PXXX(T/S)XXCP, which is within 10 Å of key active site residues, resulting in a different mechanism for catalytic activity compared with other Prdxs. The catalytic triad, Cys47, Arg132, and His39 amino acids, is responsible for the peroxidase activity of Prdx6s and is expressed in a wide range of organisms from archaea to higher mammals (Fig. 2).


These 3 amino acids that are generally conserved throughout all phyla lower the pKa of the side chain of the Cp and stabilize the deprotonated form of thiol, which is important for catalytic activity. His39 is required for activation of Cp for peroxidase activity in Prdx6, but this amino acid is not required for activity in other Prdxs (Fig. 1). Pro40 and Ser44/Thr44 also are conserved in mammalian Prdx6s.
Phospholipase activity
The GXSXG consensus motif for phospholipase activity is conserved in all aligned sequences of the Prdx6 family (Fig. 2). The serine in the motif acts as a nucleophile and is typically part of a catalytic triad involving Ser, His, and either Asp or Glu residues. In addition to Prdx6, this motif is present in serine proteases, lipases, and esterases.
In most lipases other than Prdx6, this motif is located at the center of the sequence; however, in Prdx6, it occurs at the N-terminus of the sequence. The lipase motif that is responsible for human PLA2 activity consists of amino acids 30–34 and is composed of His26, Ser32, and Asp140 residues. The critical role of Ser32 is evident from its presence in most mammalian Prdx6 enzymes (Fisher, 2018; Kim et al., 1997; Manevich et al., 2007).
Asp140 is also conserved in most Prdx6 sequences but is replaced in Plasmodium falciparum Prdx6 by a Glu that maintains the acidic properties of the protein. Active site amino acids for phospholipase activity also are conserved in most non-mammalian Prdx6 sequences, but they may differ in some organisms including Azotobacter vinelandii, Triticum aestivum, Neurospora crassa, and Caenorhabditis elegans (Bannitz-Fernandes et al., 2019).
Analysis of sequences of 126 Prdx6 homologues revealed that out of the 67% of isoforms expressing the lipase motif, 38% had the classic lipase motif (GXSXG), 11% had an alternative lipase motif (GXSXA), and 18% had another variant lipase motif (Fisher et al., 1997; Kang et al., 1998; Nevalainen, 2010).
Subcellular localization
The primary structure of Prdx6 family members consists of a specific motif/pattern, which helps in directing the post-translational transport of the molecule to various intracellular compartments. The organellar localization of Prdx6 does not depend on binding of the protein to phospholipids but does depend on its binding to the chaperone molecule, 14-3-3 (Fisher et al., 2016).
The organelle targeting motif comprising 10 amino acids (31-DSWGILFSHP-41) is located in the N-terminal region of Prdx6 (Sorokina et al., 2009) for trafficking of the molecule into acidic organelles such as lysosomes and lung lysosomal-like structures (called lamellar bodies). The serine residue at position 32 (Ser32) of Prdx6 is essential for its organellar targeting as evident from the mutation of Ser32 to Thr32. This mutation abolished lamellar body targeting of human Prdx6, whereas the PLA2 activity of mutant Prdx6 was maintained (Sorokina et al., 2016).
The C-terminal extension
Prdx6 is unique in its possession of a C-terminal extension/arm region in addition to its N-terminal Trx fold; this feature is shared with Prdx1 but is absent in other Prdxs (Fig. 1) (Hall et al., 2011; Perkins et al., 2015; Phelan, 1999). Importantly, the length of the C-terminal arm region varies among different species (Fig. 2).
Structural Features of PRDX6s
Examination of the 3-D structure of Prdx6 and its homologs in the Protein Data Bank (PDB) helps the understanding of the structural roles of the various residues and motifs in Prdx6. The following five points indicate the important structure-function features of the Prdx6 family and contrast these features with those of the members of other Prdx families. These structure-function relationships of Prdx6 are described for the major motifs; additional investigation is required to confirm some of the less well-studied relationships.
Prdx6 domains
Prdx6 has a conserved spatial structure with two distinct domains. Domain I spans from 20 to 140 amino acids at its amino-terminus and comprises a conserved Trx fold, whereas domain II includes a smaller flanking region at its C-terminus spanning amino acids 150–224. In the crystal structure of Prdx6, two monomers of Prdx6 exist in a head-to-tail dimeric structure that are related by a non-crystallographic two-fold symmetry (Fig. 3A) (Choi et al., 1998).
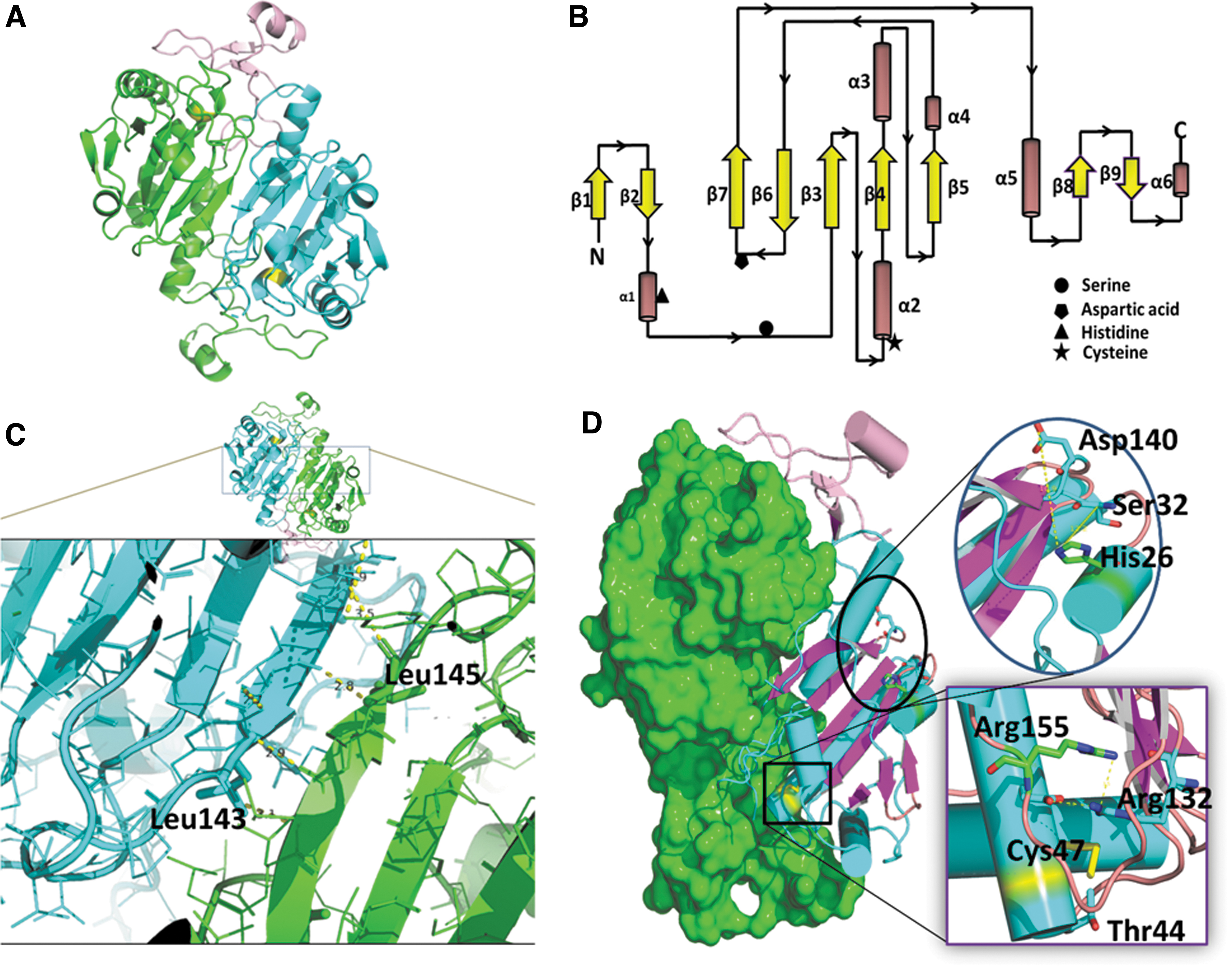
The dimer is stable and is facilitated by hydrophobic interaction of domain I of one monomer with domain II of the other monomer. The unique extended structure of the C-terminus of Prdx6 is stabilized by surface residues of the other monomer of the Prdx6 dimer. This extended domain II is also present in Prdx1, and it is involved in the interaction with sulfiredoxin for reactivating the hyperoxidized protein (Li et al., 2020). A possible role of the extended Prdx6 domain II in Prdx6 activity has not been described.
The amino acids L145 and L148 of the C-terminus play a role in the dimerization of protein monomers, and replacing these residues with Glu decreases Prdx6 peroxidase activity (Zhou et al., 2016).
Hydrophobic core
The spatially highly conserved Trx fold in the 3-D structure of Prdx6 has a common core of seven beta-strands (β1 through β7) and five alpha-helices (α1 through α5) (Fig. 3B). It has a twisted β-sheet at its center formed by five beta-strands (β5-β4-β3-β6-β7). It is notable that domain 1 contains both a βαβ motif (β strands in parallel) and a ββα motif (β strands in anti-parallel) situated at the amino and carboxy termini of the Trx fold. In Prdx6, the βαβ motif is represented by β3α2β4 and forms a hydrophobic core, which helps in binding of the substrate (Choi et al., 1998). A stretch of amino acids from 145 to 149 in the β7 strand of each monomer forms a 10-stranded β-sheet through hydrophobic interaction in an antiparallel manner.
This is termed a B-type interaction. A model of this interaction would be bringing the “palms” of the hands together side- by-side in a head-to-tail manner (Fig. 3C) (Perkins et al., 2015). It is likely that the interface of β7 is important for heterodimerization with πGST in the peroxidatic cycle of the Prdx6 enzymes, although this needs further investigation (Zhou et al., 2016).
In all Prdx structures, there is an additional β hairpin and a short helical motif at the N-terminus of the Trx fold. As mentioned earlier (primary structure section), the N-terminus contains both the organelle targeting and lipase motifs (Sorokina et al., 2016). The β hairpin of Prdx6 has a shorter β sheet than other Prdxs, as shown by analysis of the secondary structural elements (Fig. 1). There is also a motif of an α-β fold of varying length in different Prdxs before the start of the ββα motif. Domain II at the C-terminus of the molecule comprises three β strands and one α-helix. This domain II in Prdx6 is connected to the α5 helix of domain I by a loop that is followed by a beta strand, β8.
Phospholipid binding site
Crystal structural analysis of oxidized (sulfinic) human Prdx6 (PDB ID:5B6N) indicates almost 10 angstroms distance between the His26 and the Ser32 amino acids that have been proposed as the phospholipid binding site (Kim et al., 2016). Because of this relatively long distance, it has been suggested that phospholipids may bind from the flat surface across the PLA2 active site of one monomer to the peroxidatic active site of the other monomer (Kim et al., 2016).
Although confirming evidence has not been reported, we postulate that the conformation can change as the arm, possibly on binding of substrate, moves toward the active site. This movement would bring the H26 and S32 amino acids closer together, enabling them to interact with the bound phospholipid and, thereby, activate lipase activity.
Peroxidatic active site
The peroxidatic Cys (Cp) lies in the loop that connects the β3 strand and the α2 helix (Fig. 3B). The CP-loop that is part of the β3-α2-β4 motif of the Trx fold adopts an active conformation, thereby positioning CP at the bottom of the active site pocket. The peroxidatic Cp47 forms an active site triad with the guanidino group of Arg132 and the hydroxyl group of Thr44.
Two prolines in this motif help to shield the active site, whereas Asp42 and Phe43 lie above the indole ring of the highly conserved Trp82. The side chains of Glu50 form hydrogen bonds with the guanidinium moiety of conserved Arg132. Further, the nitrogen (N) of the guanidinium moiety of Arg132 can interact with the sulfur (S) of Cys47 in both the reduced and sulfenic acid forms (Kim et al., 2016).
The Cp is in the sulfenic acid form, Arg132 moves away from Cys47 to interact with Glu50. Even though there are minor structural changes of some residues with different states of oxidation, the active site pocket has sufficient room to perform its activity (Perkins et al., 2012). The α4-β6 loop in Prdx6 is believed to facilitate substrate access to the active site (Kim et al., 2016). The active site residue Arg132 is positioned through a hydrogen bonding network consisting of Arg-Glu-Arg (in human Prdx6, Arg155-Glu51-Arg132).
In Prdx5, BCP, and Tpx subfamily members, the second Arg is absent and the conserved Arg is shifted to another conformation, which is still able to form hydrogen bonds to the peroxide oxygen proximal to the cysteine thiolate as well as to the thiolate itself. The various side chain orientations near the active site contribute to the lowered pK of the peroxidatic Cys (Perkins et al., 2012).
Prdx6 family members have a conserved histidine residue (His39) directly preceding the β3-α2 loop, whereas other Prdxs have a Tyr or Trp at the corresponding position (Mizohata et al., 2005). Of note, the YF motif found in other Prdxs that helps overoxidized Cp to catalyze peroxidase activities is absent in Prdx6s.
Phospholipase active site
The active site for PLA2 activity is the catalytic triad of Ser32-His26-Asp140 (Fig. 3D). This site is unique and does not overlap the peroxidase active site. Thus, the active site for PLA2 activity is spatially and functionally independent from the active site for peroxidase activity. Of note, the distance between His26 and Asp140 is remarkably long in Prdx6 (∼9.7 Å) to form part of a catalytic triad; the distance is even longer in the known structures of oxidized human Prdx6 protein (PDB ID: 1PRX) (Choi et al., 1998; Kim et al., 2016). The crystal structure of oxidized Prdx6 shows that the active site for its binding (and PLA2-mediated catalysis) is on the protein surface and is connected to the peroxidatic site by a narrow pocket (Choi et al., 1998).
Although not confirmed by available crystal structures, conformation of the active site residues in Prdx6 may not be aligned to relay charges but their dynamic nature in solution may allow for plasticity of conformation and activation of the active site for hydrolytic properties. The oxidized phospholipid substrate can bind to the head group on the protein surface, and the oxidized sn-2 fatty acyl chain can insert into the 3D pocket, positioning it for hydrolytic activity (Manevich et al., 2007).
Mechanisms for Oxidation—Reduction of PRDX6 Proteins
Prdxs play a major role in protecting cells from oxidative stress owing to their abundance and relatively high catalytic activity (Hall et al., 2011). The reduction of peroxide oxidants such as hydrogen peroxide, organic hydroperoxides, and peroxynitrite by Prdx using the conserved Cp residue involves three main chemical steps of: (1) peroxidation, (2) resolution, and (3) recycling. While Prdx6 exhibits similar chemistry to other Prdx enzymes related to its peroxidation activity, the resolution and recycling steps of Prdx6 differ from that of other Prdxs.
Peroxidation
In the peroxidation step, the peroxide substrate binds to the reduced and fully folded Prdx6 enzyme. The conserved cysteine residue (CP) in the active site of Prdx6 then attacks the peroxide substrate, resulting in the formation of a covalent adduct between the cysteine residue and the peroxide. This attack by the nucleophilic cysteine residue is facilitated by a nearby proton-donating amino acid residue, such as histidine or arginine, which helps to protonate the leaving group of the peroxide in an SN2-type reaction.
The products of this reaction are a Cys-sulfenic acid (CP-SOH) intermediate plus water or alcohol, depending on the peroxide substrate that is being reduced (Hall et al., 2011). This peroxidation step is crucial to the Prdx6 catalytic cycle and allows the enzyme to scavenge and reduce harmful peroxide oxidants.
Resolution
Resolution (reduction of oxidized Prdx6) is necessary to complete the Prdx6 oxidation/reduction cycle. In competition for resolution, the CP-SOH product generated by peroxidation of Prdx6 is prone to irreversible oxidation to a sulfinic (CP-SOOH) or a sulfonic (CP-SO3H) acid derivative that makes Prdx6 peroxidatically inactive. As protection from its oxidation, CP-SOH may be: (1) dehydrated reversibly to form a sulfenylamide (CP-SN), (2) reduced by the accompanying cysteine residue in the C-terminus to form an intramolecular disulfide, or (3) glutathionylated by πGST to form a heterodimeric disulfide (Fisher, 2017; Forman et al., 2017; Zhou et al., 2016).
Since Prdx6s lack a resolving cysteine, an external electron donor is required to reduce the sulfenic acid intermediate. Potential external electron donors (described in greater detail below) include Trx, ascorbic acid, the GRX-GSH couple, and the GST-GSH system (Manevich et al., 2004; Rouhier et al., 2001).
Recycling
The formation of CP-SOH leads to conformational change from the FF active state to a locally unfolded conformation to allow interaction of the sulfenic acid intermediate with piGST (Fig. 4) to form a heterodimeric disulfide (Prdx-SP–S-G) with the release of H2O. This recycling step also involves the reduction of the disulfide by another protein or small thiol molecule to regenerate the free thiols, SPH and GST.

Crystal structure analysis of human Prdx6 shows that the peroxidatic Cys47 is situated at the base of a relatively narrow pocket (Choi et al., 1998), but the pocket has accessibility for a thiol of appropriate size to reduce the oxidized Cys47 to its functional native conformation (Peshenko and Shichi, 2001).
This suggests that the reduction of the disulfide may occur through a thiol-disulfide exchange reaction, with the reducing thiol interacting with the peroxidatic Cys residue in the narrow pocket. There is wide variation in which small thiol molecules can generate a free thiol (SPH) to interact with Prdx6 in various organisms. Thus, diverse agents act as the reductant to reduce oxidized Prdx6, thereby preventing oxidative stress.
GSH has been proposed as the probable primary physiological reductant in mammalian cells due to both its high intracellular concentration and the inability to identify any other reductant as the primary one (Deponte and Becker, 2005; Pedrajas et al., 2010; Peshenko et al., 1996; Shichi and Demar, 1990). But studies with recombinant Prdx6 protein have failed to show universal effectiveness of GSH (Fisher et al., 1999; Singh and Shichi, 1998). This suggests that other reductants can be involved in the reduction of oxidized Prdx6.
Potential physiologic reductants besides GSH include cysteine, homocysteine, N-acetylcysteine, mercaptosuccinic acid, NADPH, Trx, ascorbate, and cyclofilin A (Table 1). Non-physiologic reductants that can act as electron donors to Prdx6 in vitro include β-mercaptoethanol and dithiothreitol (DTT), although they are not effective in vivo.
Physico-Chemical and Functional Characteristics of Prdx6 Enzyme Family in Different Organisms
Unlike DTT that supports Prdx6 peroxidase activity, β-mercaptoethanol can form a mixed disulfide with Prdx6 in vitro but does not result in peroxidase activity. Bovine Prdx6, which has 87% identity to the human Prdx6 amino acid sequence, showed peroxidase activity in the presence of glutathione, and could be reduced by DTT as well as by dihydrolipoic acid and hydrogen sulfide (Pedrajas et al., 2010; Peshenko and Shichi, 2001; Zhou et al., 2016).
But oxidized bovine Prdx6 was not reduced by cysteine, homocysteine, N-acetyl-cysteine, mercaptosuccinic acid, or NADPH confirming that none of these latter agents are effective physiologically as a co-factor for the peroxidase activity of mammalian Prdx6 (Fisher et al., 1999; Kang et al., 1998).
Ascorbic acid is another physiological compound that can reduce oxidized Prdx6 in vitro and has been demonstrated to protect glutamine synthetase from oxidative inactivation in an assay containing Prdx6, Fe, and O2 (Monteiro et al., 2007). The K m for reduction of human Prdx6 by ascorbate in association with H2O2 is 760 μM and the ascorbate-dependent efficiency for H2O2 reduction (kcat/Km) is only 0.9 × 106 mol−1 s−1 (Manevich and Fisher, 2005).
A comparison of seven Prdx6 proteins from different organisms showed peroxidase activity with 500 μM ascorbate at 93% as compared with the same concentration of DTT (Monteiro et al., 2007). Thus, reduction mediated by ascorbate is less efficient than glutathione-dependent reduction. Despite this, ascorbate as a reductant may have an important physiological function in plants as described next.
A variety of Prdx6 reductants are active in non-mammalian organisms. Trx can play a major role; it is an efficient reductant of oxidized Prdx6 and is important physiologically in many organisms. For example, yeast Prdx6 has peroxide reductase activity using the mitochondrial Trx system as an electron donor (Li et al., 2020; Pedrajas et al., 2000). The role of NADPH thioredoxin reductase (NTR) in this process supports its critical role in maintaining the function of cells under oxidative stress (Pulido et al., 2009).
In the Cyanobacterium Synechocystis sp. PCC 6803, thioredoxin B, which is encoded by the slr1198 gene, is a highly efficient electron donor for Prdx6 (Hisabori et al., 2005; Perez-Perez et al., 2009). Similarly, Trx is the reductant during the catalytic cycle of Prdx6 in the aerobic hyperthermophilic archaeon Aeropyrum pernix (Kawakami et al., 2004).
Other compounds with an oxidized state that is reduced by Trx are Thermococcus kodakaraensis and the anaerobic hyperthermophilic archaeon Pyrococcus shorikoshii; both contain a consensus sequence PVCTTE and a primary structure that shows high homology with the human Prdx6 sequence. In all of these examples with Trx serving as the reductase, the Prdx6 activity from a functional standpoint resembles that of a 2-Cys Prdx as both conserved CP and CR are involved in the peroxidatic cycle with Trx serving as the reductant of the inter-subunit disulfide (Kawakami et al., 2004; Lee et al., 2015).
Other mechanisms for the reduction of oxidized Prdx6 protein also exist. Both Brome grass and Arabidopsis thaliana utilize cysteine from another protein or a small thiol molecule for the physiological reduction of oxidized Prdx6. The AhpE expressed in Mycobacterium tuberculosis, a Prdx6 family member, can be reduced through a mixed disulfide with either mycoredoxin-1 or mycothiol (Hugo et al., 2014; Kumar et al., 2016). NADPH can regenerate (reduce) Prdx6 in the nuclei of wheat seed cells exposed to oxidative stress.
Finally, there would appear to be other reductants of oxidized Prdx6 yet to be identified. As a basis for this supposition, P. falciparum Prdx6 (PfTPx-1) that is present in protozoan parasites has only low peroxidase activity with physiologically available reducing agents, including Trx1, GSH, and Grx (Nickel et al., 2006). Thus, the probable physiological reductant of PfTPx-1 remains unknown.
Oligomerization State of PRDX6 Proteins
Both redox regulation and oligomerization of Prdx proteins are complex processes that are important for their functions in maintaining cellular redox homeostasis. Homodimerization is common within all Prdx subfamilies including Prdx6 (Perkins et al., 2015) and the existence of mammalian Prdx6 in dimers and higher oligomers has been observed under various physiological conditions. The formation of dimers or higher oligomers of Prdx6 depends on the oxidative state of the protein, pH of the solvent, and the presence/absence of substrate (Table 1) (Bystrova et al., 2007; Chowhan et al., 2021; Shahnaj et al., 2019; Shahnaj et al., 2020; Wu et al., 2006).
Significantly, all oligomerizations of Prdx6 are multiple forms of dimers (Lee et al., 2015; Pedrajas et al., 2000). Oligomerization does impact the peroxidase activity of Prdx6 since this function is redox based and is conformation- and oligomer-driven (Chowhan et al., 2020). However, there is great variability among various species in the oligomers formed within Prdx6s. The dimer-higher oligomer interconversion seems to play a role during the catalytic cycle and influences the catalytic efficiency (Wood et al., 2002), but the functional effects of different oligomers have not been well studied.
The following describes some of the conditions that are important for oligomerization of Prdx6:
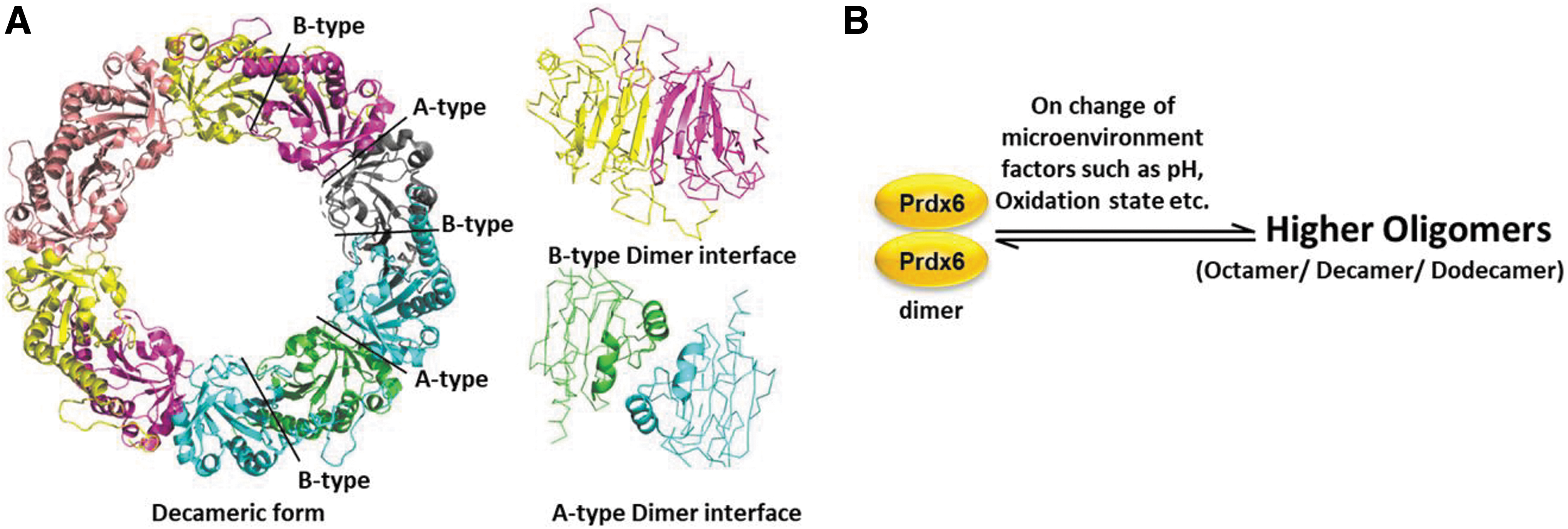
Toroidal complexes
The B-type dimer of Prdx6 can interact to form toroidal complexes, coming together through interactions at their A interfaces to form (α2)5 decamers and rare (α2)6 dodecamers (Fig. 5).
Overoxidation
The hyperoxidation of the CP cysteine of Prdx6 to sulfinic and sulfonic acid forms irreversibly inactivates its peroxidase activity. This hyperoxidation also occurs in Prdx1 and can be reversed by ATP-driven sulfiredoxin, indicating that different Prdx proteins may have different redox regulatory mechanisms (Chuang et al., 2006; Jang et al., 2004; Moon et al., 2005).
It is reported that hyperoxidation of Prdx6 induces cell cycle arrest at the G2/M transition through up-regulation of its PLA2 activity (Kim et al., 2008). Biophysical studies have shown that hyperoxidation of Prdx6 induces the formation of higher oligomers, such as decamers, that expose its catalytic triad and enhance its aiPLA2 activity (Shahnaj et al., 2019).
Similarly, the Prdx6 proteins from several archaea, including Aeropyrum pernix (ApPrdx), Pyrococcus horikoshii (PhPrdx), and Thermophilic archaeon (TkPrdx), can form decamers or dodecamers under non-reducing conditions. In the case of PhPrdx, a redox-dependent conversion of its quaternary structure was observed and a point mutation at the sixth position made it similar to human Prdx6 and resulted in a dimer in solution but a dodecamer in crystal structure (Nakamura et al., 2017).
TkPrdx, a Prdx6 from the anaerobic archaeon T. kodakaraensis KOD1, has been found to switch its function from a peroxidase to a molecular chaperone on hyperoxidation-induced formation of higher oligomers (Lee et al., 2015). This chaperone function is disrupted by mutation of its Cys moiety, indicating that it requires the formation of higher oligomers. SiPrdx, another Prdx6 from the archaeon S. islandicus, behaves mainly as a decamer, suggesting a more specialized chaperone function than that of TkPrdx (Stroobants et al., 2019).
C-terminal extension
The Prdx6 family is characterized by a unique C-terminal extension/arm region following the Trx fold. This extension contains a conserved “CXDWWFC” motif in most archaeal Prdx6 proteins, where the cysteine residues in the motif form a disulfide bond. This disulfide bond helps the dimers to establish higher toroid-shaped oligomers including octamers and hexadecamers with two-fold symmetry (Lee et al., 2015; Nakamura et al., 2017).
The Prdx6 protein of A. pernix contains three cysteine residues, namely Cys50, Cys207, and Cys213; this Prdx6 can form either two-fold toroid-shaped octamers and hexadecamers associated with the formation of intermolecular disulfide bridges between Cys50 and Cys213 in the normal reaction mechanism in the fully-folded conformation or intramolecular disulfide bonds between Cys207 and Cys213 in the locally unfolded state at high H2O2 concentration (Jeon and Ishikawa, 2003).
The formation of these disulfide bonds plays a critical role in regulating the activity of Prdx6s, namely the peroxidase activity that protects cells from oxidative stress. The oligomeric state of the protein and the location of the disulfide bond formation determine the activity of the protein, as well as its ability to interact with other proteins or cellular components.
Presence of cysteines
Some Prdx proteins functionally are 2-Cys proteins but have some characteristics of a 1-Cys Prdx (i.e., Prdx6) and vice versa. Prdx in the marine annelid (Arenicola marina) can be classified as a Prdx6 based on similarity (65%) of its amino acid sequence to human Prdx6, but it has five cysteines, two of which function as peroxidatic and resolving cysteines, similar to a typical 2-CysPrdx (Loumaye et al., 2008; Smeets et al., 2008). Further, unlike Prdx6, the structure of region IV corresponding to the loop between strands b6 and b7 in A. marina prevents the formation of toroid-shaped decamers (Wood et al., 2002).
Prdx from Toxoplasma gondii (Tg-Prdx2) belongs to the Prdx6 family even though, like A. marina, it possesses 5 cysteine residues (Deponte and Becker, 2004). This Prdx shows a similar reaction mechanism as described for typical 2-Cys Prdx and it exists as a homodimer under physiological conditions (Deponte and Becker, 2004) but, unlike Prdx6, the addition of H2O2 and high salt (1.7 M NaCl) induce the formation of multimers (Barranco-Medina et al., 2009).
AhpE from Mycobacterium tuberculosis has sequence and atomic structure similar to typical 2-Cys Prdx proteins but functionally belongs to the Prdx6 family based on the absence of a resolving cysteine residue (Barranco-Medina et al., 2009); size exclusion chromatography and light scattering analysis have detected both dimers and octamers in AhpE (Li et al., 2005).
Synechoccus elongatus PCC7942 expresses six different Prdxs; one is a Prdx6 that shows 56% amino acid homology and a similar catalytic mechanism to human Prdx6 and exists as a dimer as observed through size exclusion chromatography (Stork et al., 2005, 2009).
Multi-Functional Activities of PRDX6
All Prdx6 proteins examined thus far express both peroxidase and phospholipase activities that have important physiological roles. But some Prdx6 proteins may express additional physiological activities or may participate in cell signaling that can, in turn, regulate other pathways. The following are physiological functions of Prdx6s (Fig. 6). Some of these functions were described based on the effects of mutations of Prdx6 in mouse models (Feinstein, 2019).
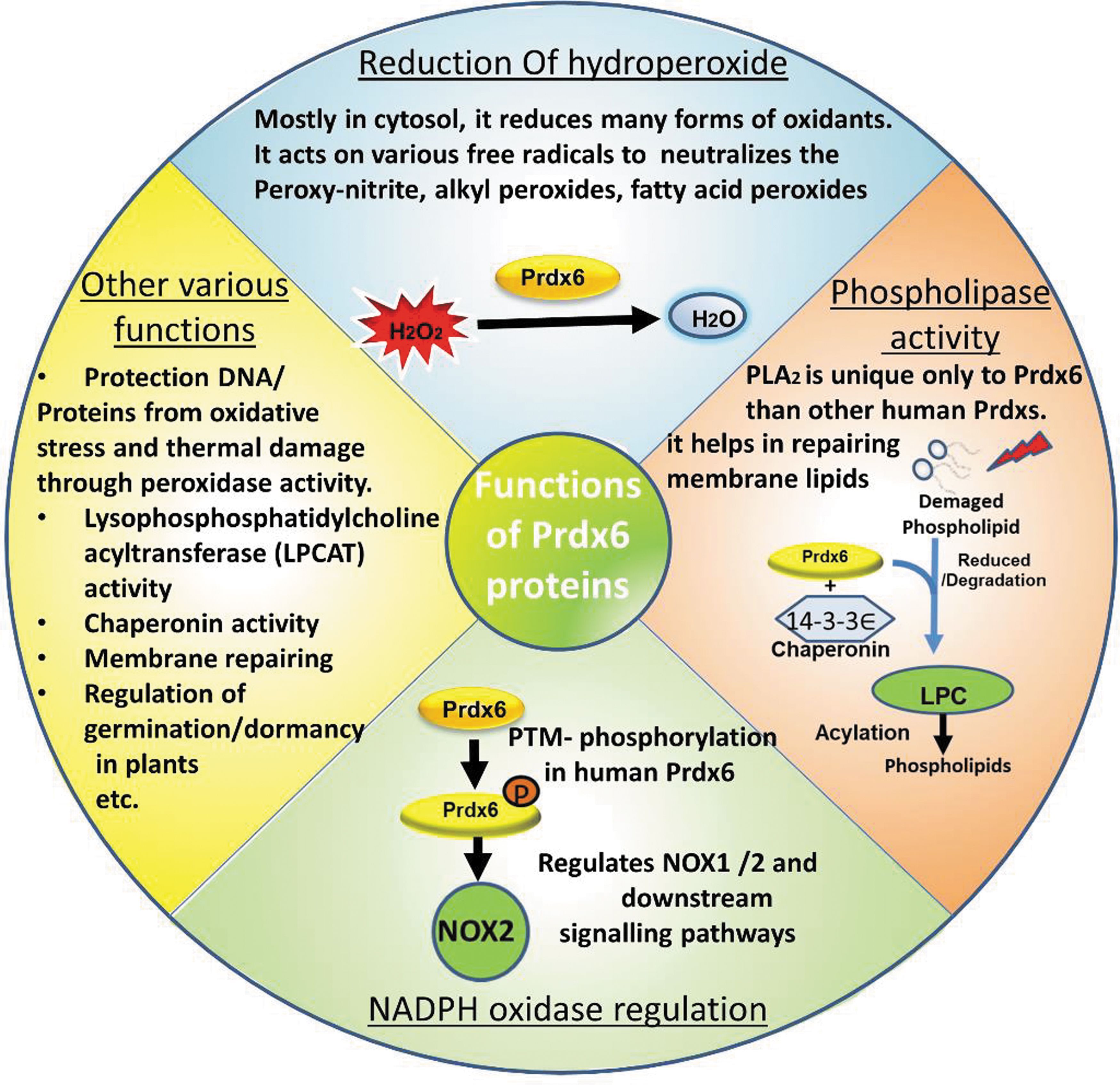
These mutations were originally characterized biochemically and subsequently; several knock-in mouse strains have been produced, each containing one mutation. These mice include the S32T knock-in that affects protein transport, the C47S knock-in that inactivates the peroxidase enzymatic activity, the D140A knock-in that inactivates the PLA₂ enzymatic activity, and the H26A knock-in that inactivates the peroxidase and blocks binding to phospholipids.
Chaperone activity of Prdx6
Some Prdx6 family proteins, including TkPrdx and SiPrdx, possess chaperone activity, in addition to their peroxidase activity. Chaperone proteins help in the proper folding of newly synthesized proteins and prevent protein aggregation and misfolding under stress conditions. The ability of TkPrdx and SiPrdx to act as chaperones provides a mechanism for these proteins to protect other cellular proteins from damage caused by stress conditions.
For example, TkPrdx has been shown to form a dodecamer structure on hyperoxidation, enabling it to exhibit chaperone activity that protects against protein aggregation induced by thermal stress, that is, heat shock (Lee et al., 2015). The mutant Cp46S of TkPrdx also exhibits chaperone activity, indicating that the formation of the oligomer is independent of the oxidized cysteine in the protein.
Instead, the C-terminal cysteines are responsible for a redox-dependent switch between a dimeric and a higher oligomeric state. SiPrdx, on the other hand, exists mainly as a decamer and exhibits a more specialized chaperone function than TkPrdx (Stroobants et al., 2019). Its decameric structure suggests a specific role in protein folding under certain conditions, but this function is presently not well understood.
Reduction of hydroperoxides and antioxidant defense
The peroxidase activity of Prdx6s can reduce hydrogen peroxide, alkyl hydroperoxides, and other reactive peroxides. This antioxidant function of Prdx6 can vary depending on the oligomeric state of the protein, which, in turn, can depend on the presence of certain Cys residues and its long C-terminal extension. In Cyanobacterium synechocystis sp., the peroxidase activity of Prdx6 (slr1198) is involved in the Trx-dependent reduction of hydrogen peroxide and alkyl hydroperoxides (Perez-Perez et al., 2009).
The Prdx6 of archaea protects its proteins and DNA from oxidative stress and thermal damage through its peroxidase activity (Lee et al., 2015; Limauro et al., 2006). In Mycobacterium tuberculosis, the Prdx6 homolog, AhpE, catalyzes the reduction of peroxynitrite and fatty acyl hydroperoxides. The conformation of AhpE is dependent on the redox state of the catalytic cysteine, as evidenced by changes in tryptophan fluorescence (Hugo et al., 2009).
In addition to reduction of H2O2 and other short chain peroxides, mammalian Prdx6 can reduce phospholipid hydroperoxides (Fisher, 2017). The role of Prdx6 in repair of peroxidized phospholipids is due to its ability to interact with peroxidized phospholipid substrate (Fisher, 2011). This interaction was shown by studies with pulmonary microvascular endothelial cells in primary culture and with isolated lungs in which His26, the main binding site of Prdx6 for hydroperoxidase activity, was specifically mutated to Ala26, thereby preventing specific binding of Prdx6 (Fisher et al., 2018).
Unlike the effect of wild-type Prdx6 where oxidized proteins were efficiently reduced, the mutant Prdx6 failed to support reduction of oxidized phospholipids (Fisher et al., 2018). Thus, it seems clear that the peroxidase activity of Prdx6 enzymes can play an important role in antioxidant defense by reducing oxidized phospholipids and thereby reverse the effects of oxidative stress.
Phospholipid metabolism and cell membrane repair
Prdx6 also has a role in cellular phospholipid metabolism based on its PLA2 and LPCAT activities. These activities are involved in the metabolism of phospholipids in cells and cellular membranes. The PLA2 activity of Prdx6 is Ca2+-independent and optimal at either acidic pH or neutral pH following its phosphorylation at Thr177 via MAPK kinase activity (Fisher, 2018; Fisher, 2017; Rahaman et al., 2012).
Several roles for the PLA2 actvity of Prdx6 have been described (Fisher, 2018): (1) the liberation of free fatty acids in vitro from the sn-2 position of phospholipids; (2) the presence of the lipase motif GXSXG in mammalian Prdx6 (the latter is also conserved in homologues of some lower organisms, although no specific function for this PLA2 activity in lower organisms has yet been described); (3) inhibition of the PLA2 activity by a designed substrate transition state analogue (called MJ33); (4) the presence of Prdx6 in intracellular acidic organelles (lysosomes and lysosomal-related organelles) in the lung epithelium and alveolar macrophages where the PLA2 activity is involved in lung surfactant metabolism; (5) inhibition of the PLA2 activity by surfactant protein A; and (6) physiologic effects from the genetic manipulation of Prdx6 (Nevalainen, 2010).
The PLA2 activity of Prdx6 is affected by the oxidation state of the protein (Jia and Dong, 2023; Kim et al., 2008); the interactions of reduced Prdx6 with oxidized substrate or oxidized Prdx6 with reduced substrate enhance both protein-substrate binding and enzymatic activity (Fisher, 2018; Fisher et al., 1999). The gain of PLA2 activity at cytosolic pH with oxidized lipids as substrate is important for understanding the role of this enzyme in the repair of peroxidized cell membranes.
At neutral pH, Prdx6 also shows LPCAT activity, that is, the transfer of a fatty acyl CoA into the sn-2 position of LPC. If this latter activity is coupled with its PLA2 activity, the combined PLA2 and acyl transferase activities allows Prdx6 to serve as a remodeling enzyme by removing one fatty acyl group and substituting another (Fisher, 2018).
An interesting example of the role of Prdx6 in phospholipid metabolism is shown by a recent study of malaria parasites. Infection with the parasites and their intravascular growth in the target organism results in significant oxidative damage to the parasite cell membranes.
This study has demonstrated that the repair of the malarial cell membrane damage requires Prdx6 that is taken up by the parasites from the host blood; inhibition of the PLA2 activity of Prdx6 prevents cell membrane damage and promotes death of the parasite (Wagner et al., 2022). Thus, an inhibitor of Prdx6 activity could function to protect against the effects of malarial infection.
Metabolism of lung surfactant
The PLA2 and LPCAT activities of Prdx6 play an important role in overall cellular phospholipid metabolism. This role in phospholipid metabolism is highlighted by examination of phospholipids involved in lung surfactant turnover. Thus, Prdx6 activity is important for the synthesis of pulmonary surfactant (Manevich and Fisher; 2005, Fisher et al., 2016).
Lung surfactant is a lipid-protein complex that plays an important role in the lung by maintaining a low surface tension, thereby enabling efficient breathing. PLA2 activity catalyzes the hydrolysis of phospholipids responsible for the bulk of lung surfactant structure to produce free fatty acids and lysophospholipids. These products are then further metabolized and recycled by ancillary enzymes and pathways to thereby maintain the composition and functional properties of the surfactant.
So, both the peroxidase and phospholipase activities of Prdx6 are important for surfactant synthesis and turnover while also protecting surfactant lipids from oxidative damage. This regulation of the metabolism and turnover of pulmonary surfactant is crucial for normal lung function and respiration (Fisher and Dodia, 2001).
Metabolism of lipids and glucose
Recently, it has been suggested that Prdx6, in addition to its role in phospholipid metabolism, also may play a role in controlling fatty acid oxidation (Shen et al., 2023). Rats fed a high fat diet showed increased liver Prdx6, whereas Prdx6 “knock-out” exacerbated hepatic steatosis.
These rats with absent Prdx6 showed decreased RNA and protein levels of carnitine palmitoyltransferase-1 alpha and acyl-CoA oxidase 1, the target genes of peroxisome proliferator-activated receptor alpha. Further, Prdx6 promoted the hydrolysis of oxidized fatty acids through hydrolysis of oxidized low-density lipoprotein, resulting in the generation of oxidized fatty acids (Shen et al., 2023).
In addition to its important roles in lipid metabolism, it has been postulated that Prdx6 plays a role in glucose metabolism (Pacifici et al., 2022). Mice that are null for Prdx6 develop a mild form of diabetes mellitus with reduced glucose-stimulated insulin secretion and decreased peripheral insulin sensitivity. Studies with isolated pancreatic beta cells confirmed that knockdown of Prdx6 resulted in a reduction of glucose-stimulated insulin secretion (Pacifici et al., 2022). The mechanism for the possible effects of Prdx6 on glucose metabolism is unknown.
Activation of NADPH oxidase enzymes and reactive oxygen species generation
The PLA2 activity of Prdx6 is important for the activation of nicotinamide adenine dinucleotide phosphate (NADPH) oxidases types 1 and 2 (NOXes 1 and 2) and their subsequent production of reactive oxygen species (ROS) (Ambruso et al., 2012; Chatterjee et al., 2011; Kwon et al., 2016). NOXes are widely expressed, especially in phagocytes and microvascular endothelial cells.
The pathway for activation of NOXes 1 and 2 is through the Prdx6-mediated generation of lysophosphatidic acid (LPA) and the subsequent interaction of LPA with its receptor (LPAR) followed by the release and translocation of Rac protein to the cell membrane (Vázquez-Medina et al., 2016). Rac is one of the four cytosolic proteins that are required for the activation of cell membrane-bound NOX 1 or 2. Thus, Prdx6 has an important role in promoting the multiple physiological roles of these NOX enzymes as discussed next.
Anti-bacterial defense
The generation of ROS by NOX enzymes in mammalian organisms has an important role in anti-bacterial defense and bactericidal activity (Vermot et al., 2021). Thus, Prdx6 through its role in the generation of ROS is important for both the prevention and treatment of bacterial infection.
Sperm fertilization
Loss of the PLA2 activity of Prdx6 and the subsequent decrease in NOX-generated ROS impairs the fertilizing competence of mouse sperm in vitro, indicating an important role for Prdx6 in male fertility (Moawad et al., 2017). Both the PLA2 and peroxidase activities of Prdx6 appear to be important for sperm viability (Bumenlag et al., 2022).
Cell death (ferroptosis)
Another physiologic role for Prdx6-mediated ROS generation is the regulation of ferroptosis, an iron-dependent form of cell death triggered by oxidized lipids (Zou et al., 2023). Prdx6 can regulate ferroptotic cell death, and decreased Prdx6 leads to increased cell death in the presence of ferroptosis inducers such as erastin (Lu et al., 2019). The increased ferroptotic-mediated cell death is associated with an increase in cellular lipid hydroperoxides that can be regulated, at least in part, by Prdx6 activity (Jia and Dong, 2023).
Other functions for Prdx6-mediated ROS generation
Additional roles for NOX-generated ROS in cellular physiology and pathology that are possibly mediated through Prdx6 activation are still under active investigation. These include the post-translational processing of proteins, cellular signaling, regulation of gene expression, and cell differentiation (Vermot et al., 2021).
Tissue injury by ROS
While Prdx6-mediated activation of ROS production can have important physiologic functions in the organism as described earlier, excessive ROS production from NOX enzymes may cause tissue injury, primarily through oxidation of cellular lipids. This represents a “toxic” effect of Prdx6-mediated NOX1,2 activation. The effect of excessive ROS from NOX activation is demonstrated by the protective effect of inhibition of the activation of NOXes 1 and or 2.
The consequent decrease in ROS generation is protective in various models of mouse lung injury, including: (1) inflammatory lung injury associated with lipopolysaccharide administration (Fisher et al., 2019; Lee et al., 2014); (2) hyperoxia-induced lung injury (Benipal et al., 2015); (3) lung injury associated with lung overventilation (called VILI) (Fisher et al., 2021a); and (4) lung injury following cecal ligation and puncture (Fisher et al., 2021b).
Thus, the regulation of ROS production by the activation of NOX enzymes, which requires Prdx6 PLA2 activity for the release of Rac protein, may have both physiological and pathological consequences for the organism. Clearly, there appears to be a thin line between the beneficial and the detrimental effects of NOX 1,2 activation. Thus, considerable additional research will be required to determine the extent of Prdx6/ROS effects and to clarify the mechanisms and parameters for differentiation between the beneficial and damaging effects of the Prdx6-mediated activation of NOX2.
Carcinogenesis and metastasis
Increased expression of Prdx6 has been demonstrated to be associated with various neoplasias and, in some cases, increased Prdx6 expression has correlated with tumor invasiveness (Falidas et al., 2021; Li et al., 2022). Examples of Prdx6 expression correlating with increased malignant progression include skin cancers (Rolfs et al., 2013) and human cervical cancer (Hu et al., 2020).
An in vitro study with a gastric cancer cell line showed that knockdown of Prdx6 gene expression inhibited cell proliferation, migration, and invasiveness (Mu et al., 2022). Histologic examination of excised colon cancer tissue showed Prdx6 protein overexpression in about 40% of specimens (Falidas et al., 2021). Examination of intrahepatic cholangiocarcinoma (ICC) from patients showed that Prdx6 expression was significantly increased in the tumors compared with the peritumoral tissues. Experimentally, Prdx6 genetic knockout in rats significantly inhibited ICC progression.
The increased Prdx6 expression that has been detected in many types of cancer has correlated with increased ROS production (Li et al., 2022). The ROS can be involved in both pro-tumorigenic and anti-tumorigenic signaling (Moloney and Cotter, 2017). As an example, excessive ROS production appears to contribute to the metastasis and invasiveness of lung cancer cells, although the mechanism is not yet understood (Han et al., 2016).
In a human hepatocarcinoma cell line with Prdx6 knockout, there was diminished respiratory capacity, downregulation of mitochondrial proteins, and alteration of mitochondrial morphology (Lopez-Grueso et al., 2020). At present, it is unclear which of these effects of Prdx6 on carcinoma growth and invasion are specifically secondary to increased ROS production (possibly through NOX1 or 2) or are due to some other activity related to Prdx6.
Function of Prdx6 in plants
Seed germination
The Prdx6 gene in plants was first identified in Brome grass, and subsequent studies identified the gene's expression in hydrated dormant embryos (Goldmark et al., 1992). Prdx6 expression increases in response to abscisic acid (ABA), which arrests germination, suggesting that Prdx6 expression has a dormancy-related function (Stacy et al., 1996).
Prdx6 protein is localized mainly to the nucleolus of cereal seed cells, including arabidopsis, rice, buckwheat, and wheat; it plays a crucial role in the protection of nucleic acid DNA from oxidative damage during the period of drying and reswelling of seeds (Stacy et al., 1996). A. thaliana Prdx6 (AtPER1) on interaction with ABA plays a crucial role in regulating seed germination under increased osmotic pressure, increased salt concentration, and oxidative stress.
Specifically, decreased levels of AtPER1 were associated with earlier seed germination under stress. Conversely, plants overexpressing AtPER1 exhibited reduced efficiency in germination compared with wild-type plants. These observations suggest that AtPER1 serves as a stress sensor and is involved in delaying germination and seedling establishment in unsuitable or suboptimal environments (Haslekas et al., 1998).
Gene expression in plants
Nuclear Prdx6 in plant seeds, such as Triticum aestivium, shows antioxidant nitroreductase activity (NTR) in the presence of NADPH (Pulido et al., 2009). This antioxidant activity of the NTR/Prdx6 redox system in the nucleus may have a relevant physiological function, since nuclei are a major source of H2O2 in plants as shown in tobacco bright-yellow cells (Ashtamker et al., 2007).
The nuclear level of hydrogen peroxide may control the redox status of nuclear proteins that are important for gene expression. These nuclear proteins include transcription factors that are subject to redox regulation as, for example, the R2R3 Myb factor from maize (Heine et al., 2004). However, the Prdx6 found in mitochondria of Saccharomyces cerevisiae shows only Trx peroxidase activity to protect it from oxidative stress (Li et al., 2020). Recombinant expression of AtPER1 decreased the sensitivity of Escherichia coli cells to cumene hydroperoxide, compatible with its role in antioxidant defense (Haslekas et al., 2003).
Plant aging
Metabolizing and photosynthesizing plant cells contain ascorbic acid in mM concentrations, a level compatible with a significant role in antioxidant defense (Monteiro et al., 2007). Thus, ascorbate may be important in the reduction of oxidized Prdx6 in plants. Both the ascorbate concentration and the reduction state of plants decrease during aging and also with environmental stress, suggesting that ascorbate is a major reductant of the sulfenic acid form of Prdx6 in plants (Foyer and Noctor, 2009).
Summary
Prdx proteins containing one or more cysteine residues are present in many organisms of essentially all kingdoms of life from lower prokaryotes to higher mammals. Prdx6 is a unique member of the Prdx family of proteins and exhibits a diverse range of biochemical activities, including glutathione peroxidase, PLA2, LPCAT, and chaperone activities (Table 1).
Since Prdx6 expresses multiple and essentially unrelated biochemical functions, it can be classified as a moonlighting protein (Jeffery, 2018). The distinguishing features of Prdx6 include: (1) a single conserved cysteine residue; (2) a conserved motif responsible for its peroxidase activity; (3) a lipase motif responsible for its PLA2 activity; and (4) a long C-terminal extension that is not present in other Prdxs. Eukaryotic Prdx6s contain a motif for organellar (lysosomal) targeting but lack the conserved GGLG and YF motifs found in other mammalian Prdx proteins.
These unique characteristics of Prdx6 suggest that the enzyme has evolved to perform various functions in response to different cellular stresses. Its peroxidase activity protects against oxidative stress, and its PLA2 and LPCAT activities play a role in membrane repair and cell signaling. Although studies of Prdx6 have focused mainly on eukaryotes, the protein is present in organisms from prokaryotes and plants to higher mammals.
Further studies of eukaryotic Prdx6 will provide further insights into its physiologic and pathophysiologic roles, whereas studies in lower organisms will help to shed light on the evolutionary and functional adaptation of this versatile protein family.
Footnotes
Author Disclosure Statement
No competing financial interests exist.
Funding Information
The assembly of this review article was supported in part by grant BT/359/NE/TBP/2012 (HR and LRS) from the Department of Biotechnology, Govt. of India. The authors acknowledge the Department of Science and Technology (DST), India, for a National Post-Doctoral fellowship to K. Herojit Singh (PDF/2020/000760).