
Abstract
Aims:
Reactive oxygen species (ROS) play a vital role in conveying the cytotoxicity and resistance of most chemotherapy drugs. Therefore, gaining a comprehensive understanding of the intricate activities against oxidative stress in cancer cells may provide valuable insights into the discovery of common mechanisms underlying chemoresistance.
Results:
We identified a novel long noncoding RNA (lncRNA), designated fluorouracil-associated transcript-1 (FUAT1), as a key nongenetic player involved in ROS-mediated intrinsic chemoresistance by employing a unique screening strategy based on transcriptome sequencing (RNA-Seq) technology. To investigate the precise role of the FUAT1 regulatory axis in chemoresistance, we conducted a series of in vitro and in vivo assays including gain/loss-of-function and rescue experiments. Mechanistically, our findings revealed that FUAT1 upregulates Tensin 4 (TNS4) by sponging miR-140-5p, which allows gastric cancer cells to survive chemotherapy by inhibiting ROS-mediated apoptosis. Clinically, we observed that the FUAT1/TNS4 regulatory axis is negatively associated with overall survival and progression-free survival among gastric and colon cancer patients treated with 5-fluorouracil adjuvant chemotherapy.
Innovation:
We devised a novel screening strategy distinct from conventional approaches using drug-resistant strains. Through this approach, we identified the previously unrecognized lncRNA FUAT1/TNS4 axis that plays a critical role in ROS-mediated intrinsic chemoresistance.
Conclusions:
Our findings shed light on fundamental adaptive mechanisms employed by cancer cells to respond to chemotherapy and provide new insights into developing strategies aiming at overcoming chemoresistance.

Introduction
Currently, 5
Therefore, it is crucial to investigate the specific damage-tolerant behavior of cancer cells after multidrug treatment to reveal common mechanisms underlying multidrug chemoresistance and optimize clinical outcomes. Recent studies have highlighted the essential role of intracellular reactive oxygen species (ROS) both in conveying cytotoxicity and mediating resistance against most chemotherapy drugs (Fruehauf and Meyskens, 2007; Kim et al., 2019; Pervaiz and Clement, 2004; Trachootham et al., 2009); however, a comprehensive understanding of how ROS contribute to multidrug chemoresistance remains elusive.
As byproducts of oxidative metabolism, ROS play important roles in cellular signaling transduction and oxidative stress responses. Under normal physiological conditions, an appropriate increase in ROS levels is essential for cell proliferation and differentiation; nevertheless, excessive production can trigger cell death once surpassing the cell's tolerance threshold (Kong et al., 2000; Schumacker, 2006). Cancer cells typically possess more efficient antioxidative systems compared with their normal counterparts, enabling them to better tolerate the elevated oxidative stress resulting from their rapid proliferation and metabolism (Halliwell, 2000; Sosa et al., 2013).
Innovation
The enhanced resistance of cancer cells to oxidative stress poses a significant barrier to effective chemotherapy. In this study, we devised a unique screening strategy aimed at identifying the key non-genetic drivers underlying intrinsic chemoresistance. This approach led us to uncover a novel lncRNA FUAT1/TNS4 axis that plays a pivotal role in ROS-mediated apoptosis of cancer cells in response to chemo-drugs. Mechanistically, by functioning as a miR-140-5p sponge, FUAT1 facilitates the upregulation of TNS4 expression, thereby conferring cancer cells with the ability to evade apoptosis induced by ROS (Graphical Abstract). Furthermore, we have substantiated the clinical significance of this epigenetic regulatory axis in patients diagnosed with gastric cancer and colorectal adenocarcinoma, underscoring its potential as both a promising therapeutic target and a prognostic marker for individuals who have undergone chemotherapy based on 5-FU. Thus, our findings offer valuable insights into the development of strategies aimed at overcoming chemoresistance through crucial non-genetic mechanisms.
Consequently, this augmented defensive system employed by cancer cells poses a significant impediment toward improving chemotherapy. Therefore, gaining deeper insights into the molecular mechanisms governing activities against the oxidative stress in cancer cells will offer novel perspectives for developing therapeutic strategies aimed at overcoming chemoresistance.
In this study, we developed an ROS-mediated chemoresistance screening system, through which we identified a novel long noncoding RNA (lncRNA) called fluorouracil-associated transcript-1 (FUAT1) and its downstream target Tensin 4 (TNS4). Furthermore, we demonstrated the critical roles of FUAT1 and TNS4 in ROS-mediated chemoresistance both in vitro and in vivo, and provided clear evidence of the clinical implications of these molecules based on our analysis of clinical specimens.
Mechanistically, we found that FUAT1 acts as a competitive endogenous RNA (ceRNA) for miR-140-5p, thereby modulating the expression of TNS4. Our findings unveil a previously unrecognized role of lncRNA in ROS-mediated chemoresistance, providing valuable insights into the nongenetic adaptive mechanism involved in chemotherapy.
Results
Establishment of a novel ROS-mediated chemoresistance screening system in gastric cancer cells
As a nonradical ROS, H2O2 is widely used to induce oxidative stress in cultured cells in vitro. Upon treatment with H2O2, the molecules exhibiting altered expression patterns are considered to be implicated in the cellular antioxidative response. Thus, the overlapping molecules that display differential expression in cells treated with either H2O2 or a chemotherapy drug are likely participated in ROS-mediated chemoresistance. However, for this hypothesis to hold true, two prerequisites must be met. First, the levels of ROS induced by either H2O2 or the chemotherapy drug need to be comparable so as to eliminate any potential systematic error of the experiment.
Second, it is imperative to maintain a reasonable cell survival rate after either treatment to obtain an adequate number of cells for subsequent processing and analysis. The experiment design illustrating the identification of key molecules involved in ROS-mediated chemoresistance is schematically depicted in Figure 1A.
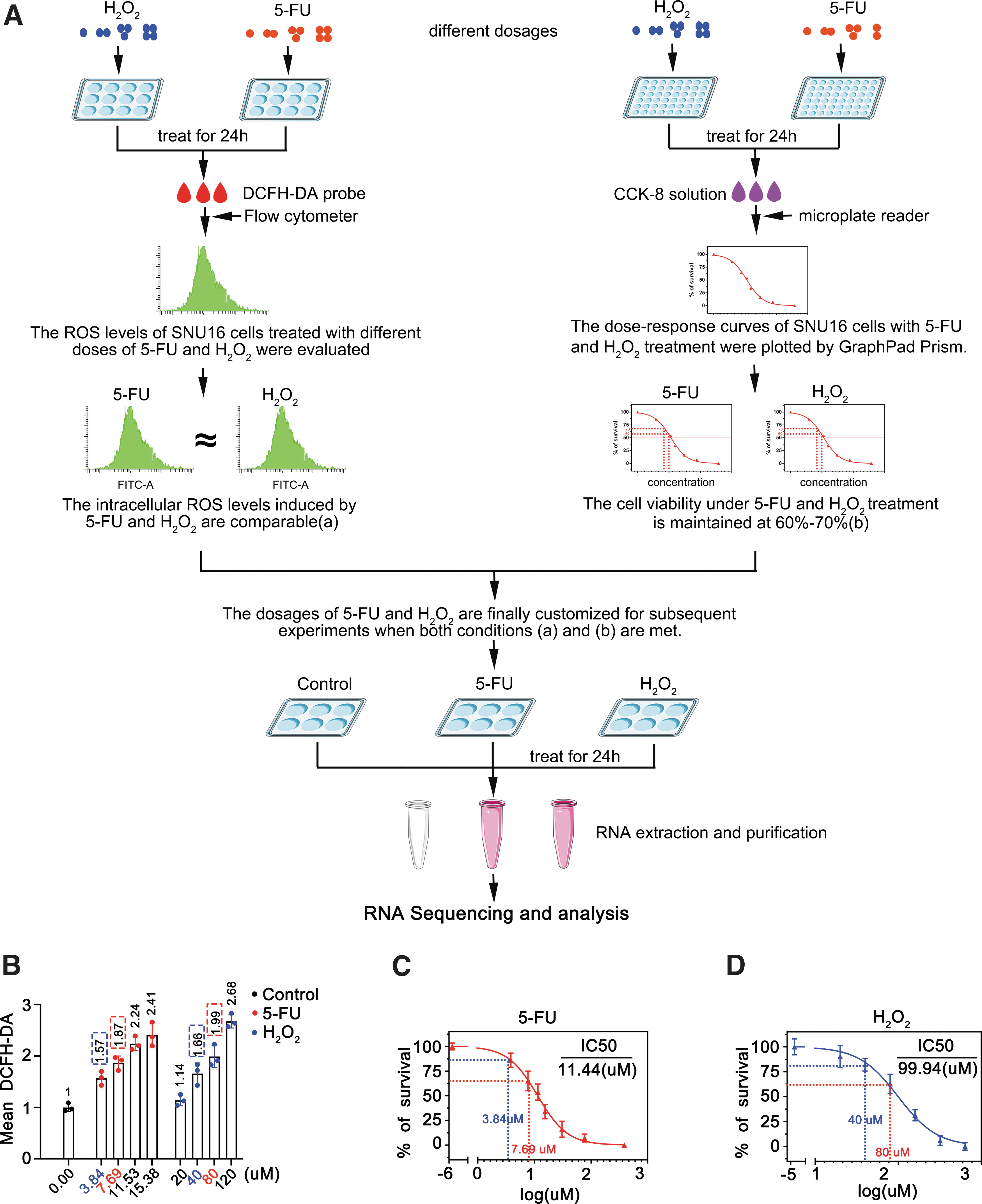
To customize this screening system, we initially determined the optimal dosage of H2O2 or the chemotherapy drug required to induce an equivalent amount of ROS in SNU16 gastric cancer (GC) cells. The corresponding intracellular ROS levels were measured after treatment with different doses of H2O2 or 5-FU for 24 h. Flow cytometer results showed that the ROS levels in the cells exposed to 5-FU at concentrations of 3.84 and 7.69 μM were approximately equal to those induced by H2O2 at concentrations of 40 and 80 μM, respectively (Fig. 1B).
Subsequently, to evaluate the viability of SNU16 cells after treatment with either 5-FU or H2O2 for a duration of 24 h, dose-dependent curves were plotted by cell counting kit-8 (CCK-8) assays. Based on these curve diagrams, the survival rates ranged from ∼80%–90% and 60%–70% upon 5-FU treatment with respective concentrations of 3.84 and 7.69 μM (Fig. 1C). Similarly, cell survival rates also roughly ranged from 80% to 90% and 60%–70%, after exposed to H2O2 at concentrations of 40 and 80 μM, respectively (Fig. 1D).
Based on these results, we optimized our screening system for ROS-mediated chemoresistance by treating SNU16 cells with either 5-FU or H2O2 at final concentrations of 7.69 or 80 μM for 24 h. Subsequently, the cells were harvested and subjected to whole transcriptome sequencing to determine alterations in gene expression profile.
Identification of a novel lncRNA through the ROS-mediated chemoresistance screening system
To identify the critical player in ROS-mediated chemoresistance, we conducted a comprehensive analysis of gene expression profile in SNU16 cells treated with either 5-FU or H2O2. The lncRNA-seq results revealed that 72 and 121 lncRNAs were upregulated in SNU16 cells after treatment with 5-FU and H2O2, respectively, with an overlap of 9 lncRNAs between the 2 treatments. Meanwhile, 35 and 185 lncRNAs were downregulated by 5-FU and H2O2, respectively, with only two overlapping lncRNAs observed (Fig. 2A). Thus, a total of 11 differentially expressed lncRNAs were identified under both treatments.

These lncRNAs are listed along with their fold changes in Table 1. Among them, our focus was on ENST00000623381.1 (GRCh38.p13), which exhibited the most significant change in expression level. This particular transcript is located on the reverse strand of human chromosome 17:40,473,554–40,475,289 and spans a length of 1736 bp; however, it lacks protein-coding potential (Supplementary Fig. S1A–D and Fig. 2B). The putative secondary structure analysis indicated desirable base pairing patterns as well as stable hairpin structures for this transcript (Fig. S1E). Given its upregulation after treatment with 5-FU, we have named it as FUAT1 (lncRNA FUAT1).
The Differentially Expressed lncRNAs in Both 5-Fluorouracil and H2O2 Treatments, Compared with Those in Untreated SNU16 Cells
5-FU, 5-fluorouracil; FC, fold change; lncRNA, long noncoding RNA.
To validate the above lncRNA-seq findings, we determined the expression level of lncRNA FUAT1 in a variety of GC cell lines. The quantitative real-time polymerase chain reaction (qRT-PCR) results demonstrated a significant increase in lncRNA FUAT1 levels upon treatment with 5-FU or H2O2 treatment across all tested cell lines. Among them, SNU16 cells exhibited the highest level followed by MKN45 cells. Consequently, these cell lines were selected for further experiments (Fig. 2C, D). Furthermore, our findings showed that both 5-FU and H2O2 induced upregulation of lncRNA FUAT1 in a dose-dependent manner (Fig. 2E, F).
Notably, NAC (the ROS scavenger N-acetyl-cysteine) abrogated the upregulation of lncRNA FUAT1 by 5-FU, indicating that ROS mediated this effect (Fig. 2G). Furthermore, platinum drugs such as cisplatin (CDDP) and oxaliplatin (L-OHP) also dose-dependently and ROS-dependently upregulated the expression of FUAT1 (Fig. 2H–K). Collectively, these results demonstrate that chemotherapy drugs treatments can significantly enhance the expression of FUAT1 through the ROS pathway, suggesting a critical role it plays in coping with chemotherapy-induced insults.
lncRNA FUAT1 confers chemoresistance in GC cells by elevating the threshold of tolerance to ROS
To investigate the potential role of FUAT1 in ROS-mediated chemoresistance, we assessed its effect on 5-FU sensitivity in GC cells. The cell viability of SNU16 and MKN45 cells was determined after exposure to different doses of 5-FU for 24 h. Our result showed that siRNA-mediated knockdown of FUAT1 significantly increased the sensitivity of these cells to 5-FU (Fig. 3A, B). Notably, silence of FUAT1 reduced the cell survival rate from 70% to 40% when treated with 5-FU at a concentration of 7.69 μM.
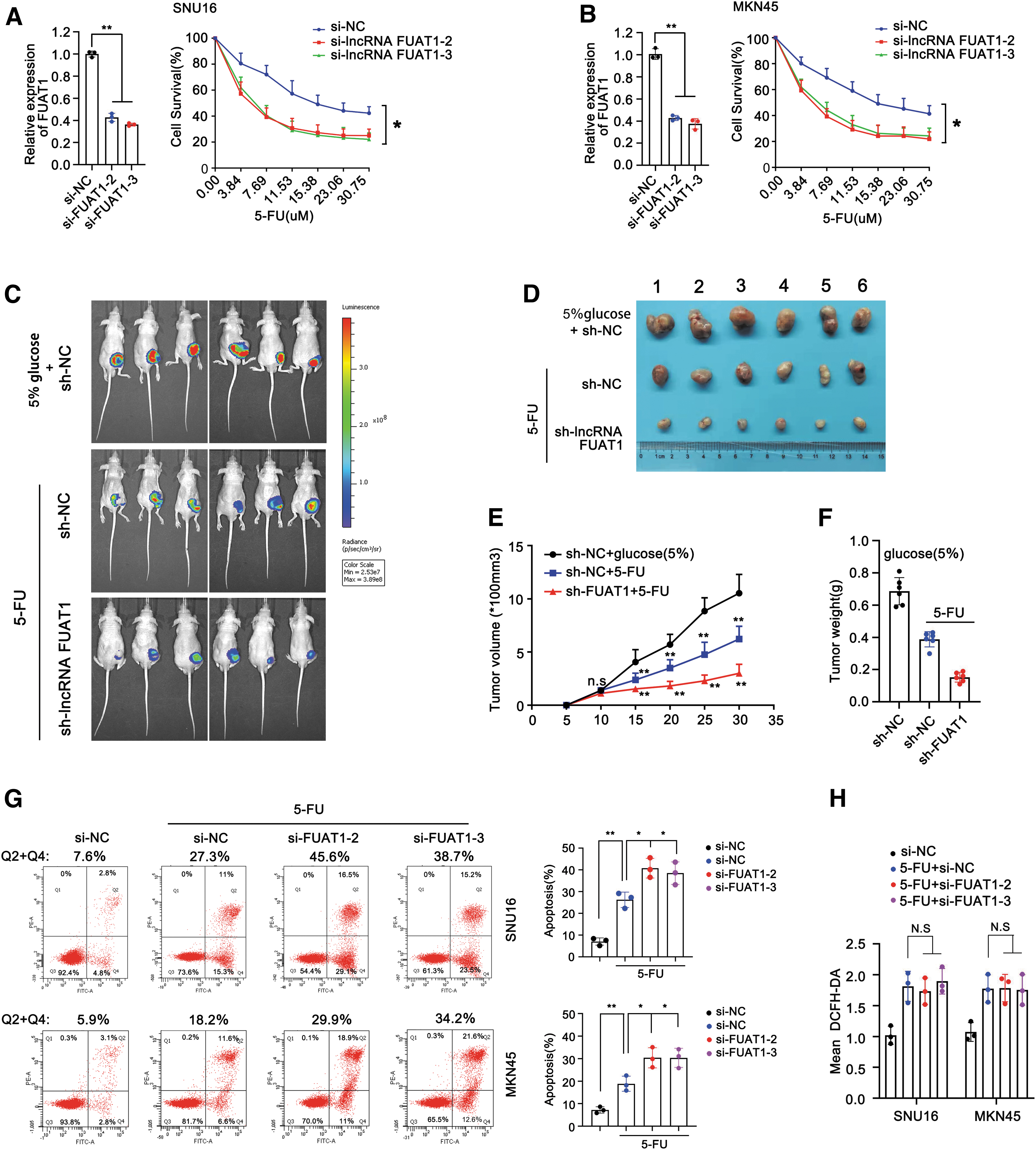
Furthermore, we evaluated the impact of suppressing FUAT1 on xenografted GC tumors in response to 5-FU therapy. Nude mice bearing xenografted tumors derived from MKN45 cells transfected with either sh-negative control (NC) or sh-FUAT1 were treated with 5-FU for 4 weeks. As expected, the 5-FU treatment groups showed slower tumor growth rates that the control group receiving 5% glucose. Strikingly, knockdown of FUAT1 further enhanced the suppressive effect of 5-FU on tumor growth (Fig. 3C). Meanwhile, both tumor size and average weight were significantly smaller and lighter in the sh-FUAT1 group compared with control groups (Fig. 3D–F). Thus, these data demonstrate that suppression of FUAT1 can enhance the sensitivity of GC cells to 5-FU both in vitro and in vivo.
Given that many cancer cells have the ability to activate their ROS scavenging machinery to protect themselves against ROS insults and escape drug-induced apoptosis, we then evaluated the effect of FUAT1 on both apoptosis rate and ROS levels in GC cells after 5-FU treatment. The Annexin V-FITC assay revealed that knockdown of FUAT1 significantly augmented the rates of 5-FU-induced apoptosis from 27.3% and 18.2% to 45.6% and 29.9% in SNU16 and MKN45 cells, respectively, indicating an evident antiapoptotic role for FUAT1 (Fig. 3G).
However, subsequent result showed that knockdown of FUAT1 had little, if not null, suppressive effect on the surge of ROS induced by 5-FU in these GC cells, suggesting that FUAT1 had no direct impact on ROS levels (Fig. 3H). Together, these findings suggest that FUAT1 may confer chemoresistance to GC cells by increasing their tolerance threshold toward ROS insults, thereby enabling them to evade ROS-mediated apoptosis.
lncRNA FUAT1 upregulates its downstream target TNS4 in response to the surge of ROS induced by 5-FU treatment
To identify potential downstream target of FUAT1, we analyzed the mRNA-seq data to look for changes in gene expression profile in SNU16 cells after treatment with either 5-FU or H2O2. Our results revealed 36 upregulated genes and another 36 downregulated genes in both treatment groups (Fig. 4A and Supplementary Fig. S2A). Growing evidence has shown that lncRNAs can exert their functions by modulating neighboring genes in cis (17). Intriguingly, among the 36 upregulated genes, TNS4 was found to be located immediately downstream of FUAT1 (Fig. 4A and Supplementary Fig. S2B).
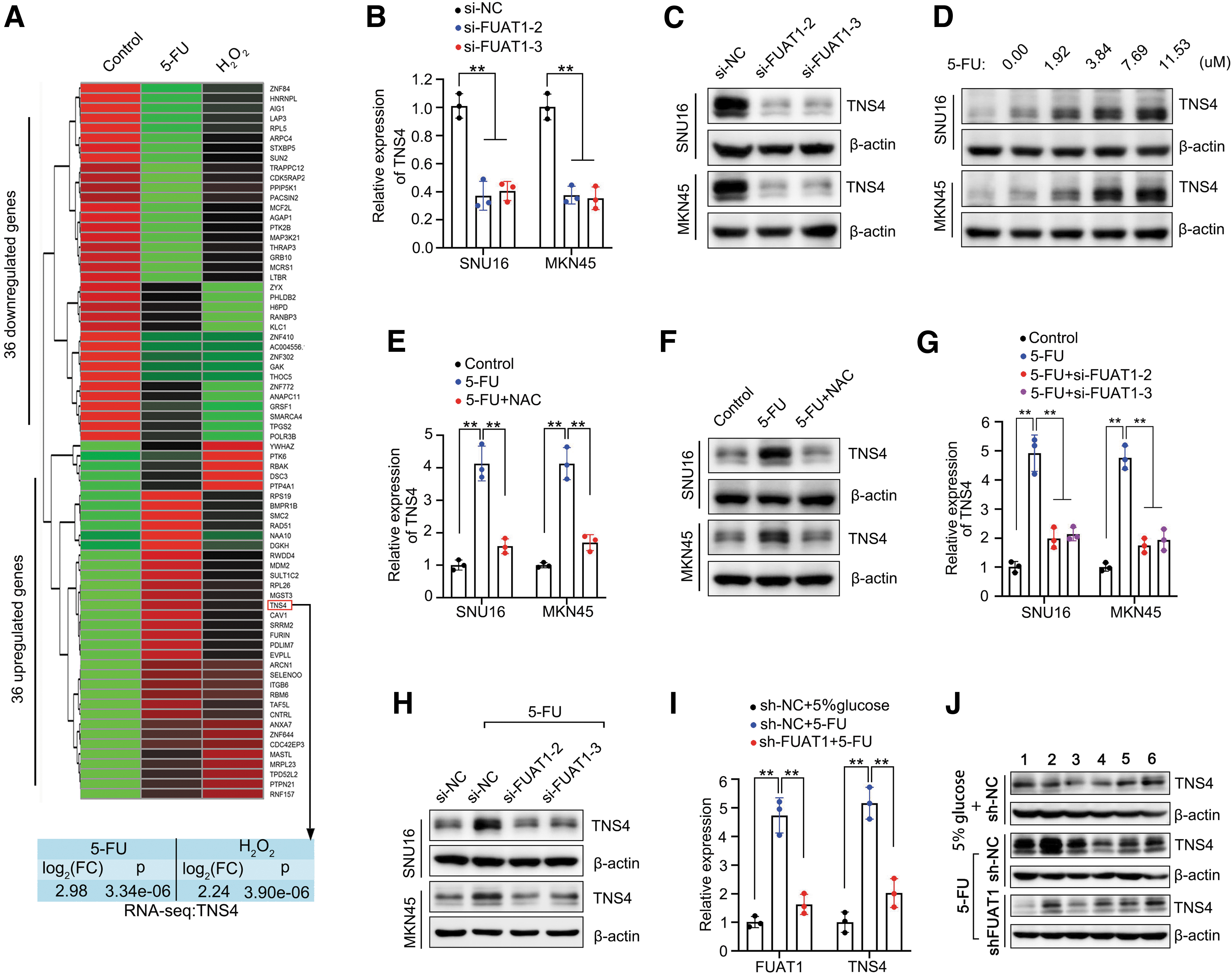
To assess whether TNS4 is a target gene of FUAT1, we examined its expression levels in SNU16 and MKN45 cells upon silencing FUAT1. The results demonstrated a significant suppression of TNS4 expression at both protein and mRNA levels after knockdown of FUAT1, indicating a critical regulatory role for FUAT1 in TNS4 expression (Fig. 4B, C).
Considering that 5-FU could induce the expression of FUAT1, we further investigated the effect of different doses of 5-FU on TNS4 expression. Remarkably, it was observed that TNS4 expression was also upregulated by 5-FU in a dose-dependent manner (Fig. 4D). Notably, this induction effect on TNS4 by 5-FU was dependent on ROS since pretreatment with NAC blocked the upregulation of TNS4 (Fig. 4E, F). Furthermore, our results showed that silencing FUAT1 abolished the upregulation of both mRNA and protein levels of TNS4 induced by 5-FU (Fig. 4G, H).
Consistently, knockdown of FUAT1 resulted in a significant reduction in elevated TNS4 levels in the xenografted GC after treatment with 5-FU, comparable with those observed in the controls (Fig. 4I, J). These results underline an indispensable role of FUAT1 in modulating the upregulation of TNS4 induced by treatment with 5-FU.
TNS4 is essential for FUAT1-mediated 5-FU resistance in GC cells
After identifying TNS4 as the downstream target of FUAT1, we conducted further investigation into the role of TNS4 in mediating 5-FU resistance. Our results showed that knockdown of TNS4 significantly affected the viability of GC cells treated with 5-FU. Specifically, silencing TNS4 resulted in a notable decrease in cell survival rate by ∼30%–40% across different doses of 5-FU (ranging from 3.84 to 30.75 μM) in SNU16 or MKN45 cells (Fig. 5A, B). Moreover, depletion of TNS4 exhibited an apparent proapoptotic effect on 5-FU-treated cells, leading to an increase in apoptotic population from ∼20% to ∼40% (Fig. 5C).
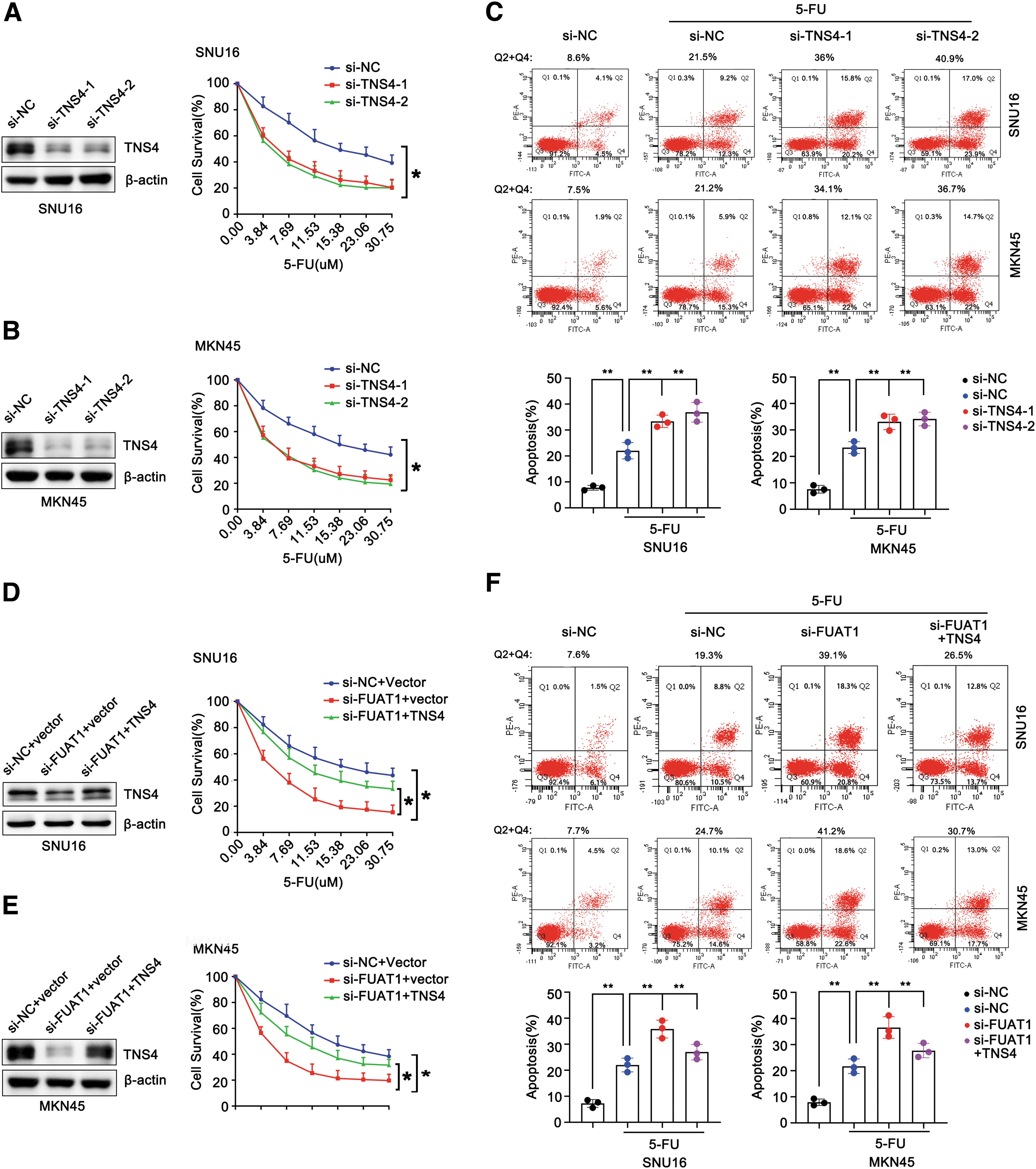
Intriguingly, exogenous expression of TNS4 partially rescued the reduced cell viability caused by silencing lncRNA FUAT1 in response to treatment with 5-FU (Fig. 5D, E). Furthermore, exogenous expression of TNS4 could attenuate the proapoptotic effect induced by silencing FUAT1 on cells treated with 5-FU, as evidenced by a discernible decline in apoptotic population from ∼40% to ∼30% (Fig. 5F). These data provide compelling evidence for the key role of TNS4 in conveying 5-FU resistance in GC cells as dictated by FUAT1.
lncRNA FUAT1 regulates TNS4 expression by competitive binding with miR-140-5p
The subcellular localization of lncRNAs plays a crucial role in determining their functions and distinguishing between cytoplasmic and nuclear regulatory mechanisms (Liang et al., 2020). To investigate how FUAT1 regulates TNS4 expression, we initially examined the subcellular localization of FUAT1 in GC cells. Fluorescence in situ hybridization (FISH) revealed that FUAT1 was predominantly located in the cytoplasm of SNU16 and MKN45 cells (Fig. 6A). As cytoplasmic lncRNAs can act as competing endogenous RNAs by competing for shared miRNAs to regulate target RNA transcripts, we subsequently screened out the potential miRNAs that could directly bind to both FUAT1 and TNS4 using bioinformatics tools (RNAhybrid and TargetScanHuman7.2).

Eleven miRNAs were found to putatively bind to FUAT1 (Supplementary Table S1), among which miR-140-5p, miR-149-5p, and miR-204-5p could also bind to TNS4 (Fig. 6B). We then transfected mimics of these three miRNAs into SNU16 and MKN45 cells to check their effects on TNS4 expression levels. Only the mimic of miR-140-5p significantly inhibited transcriptional as well as protein levels of TNS4 while its inhibitor greatly enhanced the expression levels of TNS4 (Fig. 6C–F).
To confirm the pivotal role of miR-140-5p in bridging FUAT1/TNS4 regulatory axis, we conducted a dual-luciferase reporter assay. Cotransfection of miR-140-5p mimics significantly suppressed luciferase activity in the wild-type (WT) FUAT1 reporter, whereas no change was observed in the mutant reporter (Fig. 6G). Since miRNAs exert their effects by binding to mRNA targets and causing translational suppression or RNA degradation in an Ago2-dependent manner, we performed anti-Ago2 RNA immunoprecipitation (RIP) in SNU16 and MKN45 cells transiently overexpressing miR-140-5p.
Both lncRNA FUAT1 and miR-140-5p were specifically enriched in AGO2 antibody-associated complex, with minimal enrichment observed in the control IgG (Fig. 6H, I). The FISH assay further demonstrated colocalization of lncRNA FUAT1 and miR-140-5p in the cytoplasm (Fig. 6J), providing evidence that lncRNA FUAT1 directly targets miR-140-5p. In addition, ectopic expression of an miR-140-5p mimic significantly reduced the luciferase activity driven by WT TNS4 3′-untranslated region (3′UTR) but had no effect on the mutant counterpart (Fig. 6K).
Strikingly, silencing of lncRNA FUAT1 restored inhibited luciferase activity of TNS4 3′UTR when combined with ectopic expression of an miR-140-5p inhibitor (Fig. 6L). Furthermore, knockdown of lncRNA FUAT1 led to downregulation of TNS4 levels in GC cells that was rescued by treatment with an miR‑140‑5p inhibitor (Fig. 6M), indicating that lncRNA FUAT1 functions as a ceRNA sponge for modulating TNS4 expression via sequestering miR‑140‑5p. Moreover, we observed that the introduction of miR-140-5p mimic partially counteracted the upregulation of TNS4 induced by 5-FU. Conversely, miR-140-5p inhibitor could overcome the blockage of NAC on 5-FU-induced upregulation of TNS4, suggesting an autonomous role of miR-140-5p in modulating TNS4 levels (Fig. 6N, O).
lncRNA FUAT1/TNS4 axis harbors clinical significance especially in patients receiving 5-FU-based chemotherapy
To assess the clinical significance of FUAT1/TNS4 regulatory axis, we first determined the expression levels of FUAT1 and TNS4 in GC patients. A total of 33-paired tumor and normal tissue specimens were collected from the GC patients who underwent surgical tumor resection. The subsequent qRT-PCR analysis revealed significantly higher levels of FUAT1 and TNS4 in GC tissue than normal control (Fig. 7A, B). In addition, we confirmed the overexpression of FUAT1 and TNS4 in GC tissues by analyzing the Gene Expression Profiling Interactive Analysis (GEPIA) data set, which included 211 normal and 408 GC profiles (Fig. 7C, D).
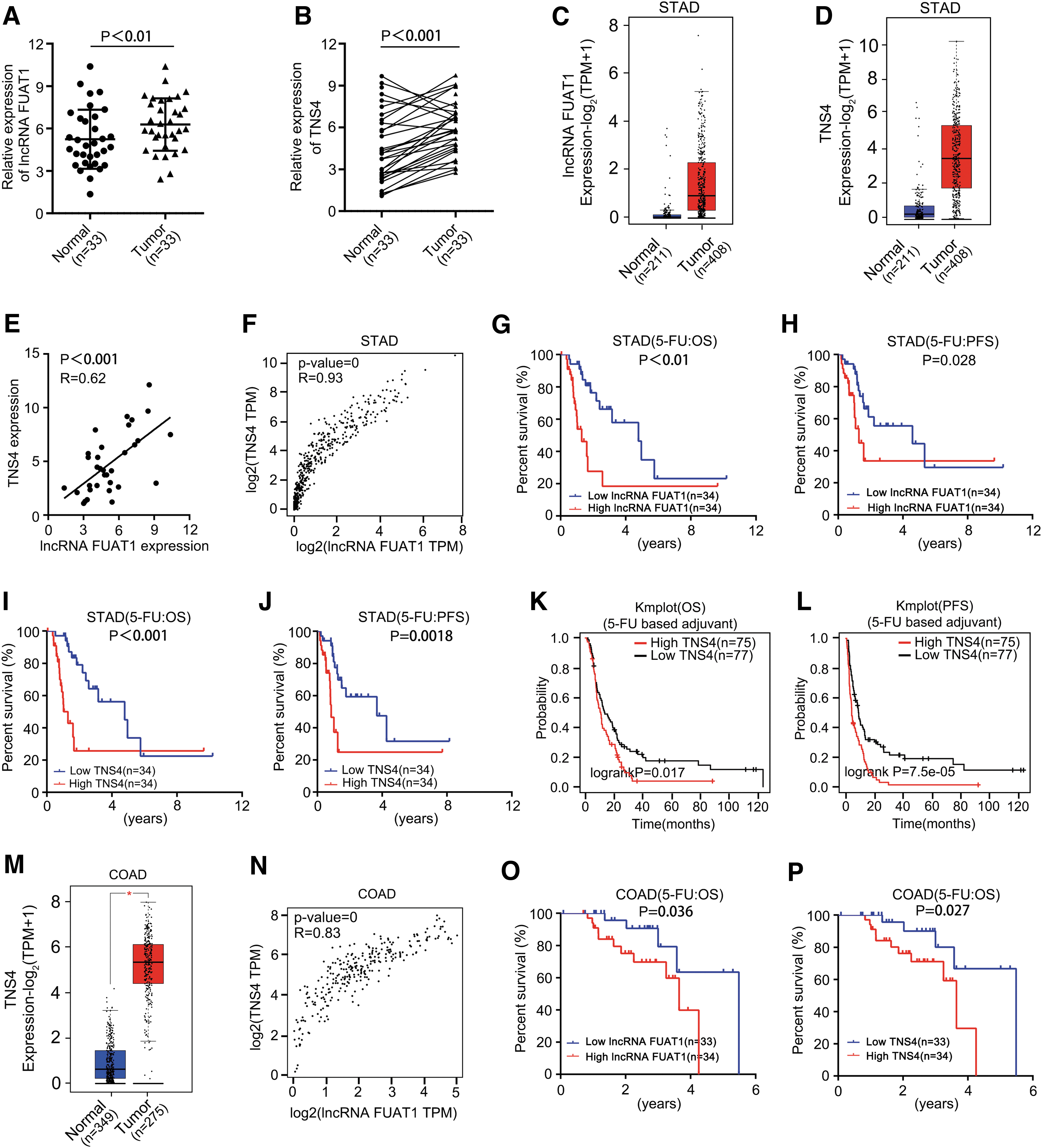
Notably, there was a positive correlation between FUAT1 expression level and TNS4 level, based on our cohort as well as the above GEPIA data set (Fig. 7E, F). These findings strongly suggest the existence of a regulatory axis involving FUAT1/TNS4 in GC, potentially playing a role in managing oxidative stress within cancerous cells.
Furthermore, Kaplan–Meier analysis conducted on our cohort revealed that the 17 patients with high expression levels of FUAT1 and TNS4 showed a worse overall survival (OS) than the other 16 patients (Supplementary Fig. S3A, B). Analysis using the “stomach adenomas and adenocarcinomas (STAD)” The Cancer Genome Atlas (TCGA) data set consisting of 216 patients further validated that patients with high expression levels of both FUAT1 and TNS4 were associated with poor OS rates (Supplementary Fig. S3C, D).
Subsequently, detailed clinical–pathological characteristics analyses were performed on our cohort as well as the TCGA-STAD cohort. Our multivariate Cox regression analysis results indicated that both FUAT1 and TNS4 could serve as independent prognostic factors for GC patients (Supplementary Tables S2–S4).
Importantly, among the 68 patients who received adjuvant therapy based on 5-FU from the aforementioned TCGA-STAD cohort, those with the higher levels of FUAT1 and TNS4 showed poorer progression-free survival (PFS) and OS outcomes (Fig. 7G–J). Furthermore, analysis results from Kaplan Meier plotter database, which included a total of 152 GC patients receiving adjuvant chemotherapy based on 5-FU, also demonstrated that high expression levels of TNS4 were correlated with inferior OS and PFS (Fig. 7K, L).
Considering that adjuvant therapy based on 5-FU is also the first-line treatment for colon adenocarcinoma (COAD) patients, we further evaluated the clinical significance of the FUAT1/TNS4 axis in these individuals. Consistent with the findings in GC cohorts, we observed significant overexpression of FUAT1 and TNS4 in COAD within a panel of profiles consisting of 349 normal samples and 275 COAD samples from the GEPIA data set. Moreover, the expression levels of FUAT1 and TNS4 were positively correlated in COAD samples (Fig. 7L, M and Supplementary Fig. S4).
Most notably, among the “colon adenocarcinoma (COAD)” TCGA data set comprising data from 67 COAD patients who underwent adjuvant therapy based on 5-FU administration, unfavorable OS was observed in those with higher levels of lncRNA FUAT1 and TNS4 expression (Fig. 7O, P). All together, these statistical analyses based on clinical data suggest that the FUAT1/TNS4 axis plays an important role in managing oxidative stress across various cancers while potentially serving as a prognostic indicator for individuals who have undergone chemotherapy involving use of 5-FU.
Discussion
The prevailing perspective on genetic evolution as the primary driver of drug resistance has recently encountered challenges (Fong et al., 2015; Shlush et al., 2017). Growing evidence suggests that genetic alterations in cancer cells are not the sole or the most common mechanism for chemoresistance (Kim et al., 2018; Marine et al., 2020). The recognition of chemoresistance without well-defined genetic causes has increasingly brought attention to an array of nongenetic adaptive mechanisms, including modulation by lncRNAs (Wei et al., 2020). However, the true prevalence and the molecular mechanisms underlying nongenetic-driven chemoresistance remain largely elusive.
In this study, we uncover a novel axis involving lncRNA FUAT1/TNS4 that plays a pivotal role in ROS-mediated apoptosis in response to chemotherapy drugs. Our results show that upon exposure to chemotherapy, cancer cells rapidly upregulate lncRNA FUAT1 to modulate the expression of TNS4 at the post-transcriptional level via mir-140-5p sponging, thereby enabling their survival under therapeutic pressure. Importantly, overexpression of the lncRNA FUAT1/TNS4 axis is negatively correlated with OS and PFS in patients treated with regimens based on 5-FU. Thus, our results support the notion that lncRNA FUAT1 acts as a critical nongenetic driver conferring inherent cellular plasticity and/or an immediate adaptive mechanism for cancer cells to evade chemotherapy.
The residual cancer cells remaining after chemotherapy, known as “minimal residual disease” (MRD), serve as a reservoir from which cancer relapse inevitably arises (Wei et al., 2020). Chemoresistance can be broadly categorized into intrinsic or acquired resistance. Although acquired resistance undeniably contributes to cancer relapse and has garnered more attention in the field, it is actually intrinsic resistance that plays a pivotal role in MRD. Therefore, accurately characterizing intrinsic resistance becomes critical for the development of therapeutic strategies aimed at preventing and circumventing disease relapse.
However, the tangled interplay between intrinsic and acquired resistances, along with the diverse array of mechanisms underlying intrinsic resistance, poses a big challenge that requires novel approaches for precisely delineating the inherent cellular plasticity of cancer cell after chemotherapy.
In this study, we employed a unique screening system to identify the key nongenetic drivers of intrinsic chemoresistance by analyzing cells at a specific time point (24 h) after chemotherapy, when establishing acquired chemoresistance is not feasible. In addition, distinct from conventional screening approaches that utilize drug-resistant strains, our delicate screening system avoids confounding factor caused by accumulated genetic alterations under persistent survival pressure. This contributes to the discovery of a panel of lncRNAs that do not overlap with those reported in previous studies (Izadirad et al., 2021; Raziq et al., 2020; Wang et al., 2021; Wang et al., 2020; Wei et al., 2019, 2020; Xia et al., 2021; Xie et al., 2022).
Moreover, instead of focusing solely on drug- or cancer type-specific chemoresistance, our screening system places greater emphasis on identifying the common mechanism of ROS-dependent chemoresistance. Indeed, our results show that both 5-FU and platinum drugs such as CDDP and L-OHP can dose-dependently up-regulate the expression level of lncRNA FUAT1/TNS4 axis (Supplementary Fig. S5). Furthermore, this axis is present and modulated upon chemotherapy not only in GC but also in other cancer types such as COAD, suggesting its potential as an attractive approach to get away from the “whack-a-mole game” caused by targeting drug- or cancer type-specific mechanism in cancer therapy.
The nuclear factor erythroid 2-related factor 2 (NRF2) has been widely recognized as the master regulator in response to ROS insults (Rojo de la Vega et al., 2018; Xue et al., 2020). Therefore, considering the crucial role of the lncRNA FUAT1/TNS4 axis in ROS-mediated chemoresistance, we aimed to investigate whether NRF2 is functionally involved in this regulatory axis. Notably, although potential NRF2 binding sites were identified within the TNS4 promoter region using the JASPA database (Supplementary Fig. S6A, B), our loss-of-function experiments provided evidence that NRF2 does not participate in the transcriptional regulation of TNS4 nor is it regulated by lncRNA FUAT1 (Supplementary Fig. S6C–E).
Consistent with these findings, our results indicate that chemoresistance conferred by lncRNA FUAT1/TNS4 axis is likely attributed to enhanced ROS tolerance rather than ROS scavenging in cancer cells. However, the precise role of TNS4 in modulating ROS tolerance remains unknown.
TNS4, a member of the Tensin focal adhesion family, predominantly localizes to cellular focal adhesions (Lo and Lo, 2002). It plays a critical role in various cellular processes including cell motility, differentiation, and survival. Aberrant expression of TNS4 has been observed in many cancers (Chan et al., 2015; Chang et al., 2021; Lo, 2014; Lo et al., 2005; Muharram et al., 2014; Qi et al., 2020; Sawazaki et al., 2017). A recent study has reported an upregulation of TNS4 expression and its nuclear localization in colon cancer, where it interacts with nuclear β-catenin to promote tumorigenicity (Liao et al., 2009).
Given the well-established role of Wnt signaling in regulating cell death machinery, it is reasonable to speculate that TNS4 may confer antiapoptotic capabilities by interacting with frequently hyperactivated β-catenin, thereby potentially contributing to drug resistance in various cancers (Chen et al., 2001; Han et al., 2013; Luo et al., 2012; Martinez-Font et al., 2017; Senthivinayagam et al., 2009; Tao et al., 2013; Yuan et al., 2005; Zhang et al., 2010). Our findings consistently demonstrate that TNS4 significantly influences the survival of GC cells after 5-FU treatment primarily through its antiapoptosis effect.
Other factors such as β-catenin-induced reprogramming event known as epithelial-to-mesenchymal transition have long been associated with chemoresistance and could potentially coexist alongside the effects mediated by TNS4. Further investigation is warranted to elucidate how exactly TNS4 confers chemoresistance to cancer cells.
The identification of a common mechanism underlying chemoresistance holds immense potential for the development of curative cancer treatment. By employing a customized screening strategy, here we discovered a novel lncRNA FUAT1/TNS4 axis that confers inherent cellular plasticity to cancer cells, enabling them to escape from ROS-mediated apoptosis induced by chemotherapy. Furthermore, we have validated the clinical significance of this epigenetic regulatory axis in patients with GC and COAD, highlighting its potential as both a promising therapeutic target and a prognostic marker for those who have undergone 5-FU-based chemotherapy.
Thus, our findings provide valuable insights into the development of strategies aimed at overcoming chemoresistance by targeting crucial nongenetic mechanisms. However, due to its complexity, this study did not explore the initial mechanism through which chemotherapy drugs/H2O2 induce expression of lncRNA FUAT1. The regulation of lncRNA FUAT1 expression may occur at multiple levels including transcriptional regulation, post-transcriptional modification, and epigenetic histone modifications involved in chromatin remodeling.
It is well established that ROS activate several key transcription factors that modulate cellular redox status (Hayes et al., 2020; Srinivas et al., 2019). Notably, when aligning sequences with well-known ROS-associated transcription factors using the JASPAR website, putative binding regions for HIF-1α, HSF1, NF-Κb, and NRF2 were found within a 2000 base-pair stretch of the 5′-promoter region of lncRNA FUAT1 except for TP53 (Supplementary Fig. S7). This finding suggests that these ROS-associated transcription factors may play a role in regulating lncRNA FUAT1 expression.
In addition, further investigations are warranted to explore the potential contributions of epigenetic modifications on histones and m6A modification on lncRNA FUAT1 itself toward regulating and stabilizing this long noncoding RNA (Li et al., 2022; Mele et al., 2017; Ni et al., 2023; Zhang et al., 2017).
Materials and Methods
RNA sequencing
RNA contamination and degradation were assessed by using 1% agarose gel electrophoresis. RNA purity and concentration were examined using NanoPhotometer® spectrophotometer. RNA integrity and quantity were measured using RNA Nano 6000 Assay Kit of the Bioanalyzer 2100 system. RNA library for lncRNA-seq was prepared as rRNA depletion and stranded method. The insert size and accurate concentration of the acquired library were examined by Agilent 2100 Bioanalyzer and qPCR, respectively. The samples were then subjected for sequencing when the insert size and concentration of the library were identical.
The PE150 (paired-end 150 nt) sequencing was performed for lncRNA-seq. All the transcripts were merged using Cuffmerge software and lncRNAs were identified from the assembled transcripts. Quantification of the transcripts and genes was performed using StringTie software and Reads Per Kilobase of transcript per Million mapped reads was obtained. Cuffdiff or edgeR was used for differential expression analysis. The resulting p-values were adjusted using the Benjamini and Hochberg's approach for controlling the false discovery rate. Genes with |log2 (Fold Change) | > 0 and p < 0.05 were assigned as differentially expressed. The RNA sequencing raw and processed data have been submitted to the GEO database and the GEO accession numbers is GSE236987.
Clinical samples and processing
Thirty-three paired GC and normal tissues were obtained from Tianjin Medical University Cancer Institute and Hospital (TMUCIH), China. Informed consents were signed by all patients. The study was approved by the ethics committee of TMUCIH. Tissue samples were stored at liquid nitrogen immediately after surgery. The total RNA was extracted for qRT-PCR assay.
Cell lines and cell culture
SNU16 cells were purchased from American Type Culture Collection (ATCC; Cat. no.: CRL-5974). MKN45 (Cat. no.: 1101HUM-PUMC000229) and HEK293T (Cat. no.: 1101HUM-PUMC000091) cells were obtained from the National Infrastructure of Cell Line Resources (Beijing, China). All the above cells were grown in the medium according to the instructions of the providers in a 37°C incubator (Thermo Scientific) with 5% CO2. The cells had been authenticated for STR profiling and tested for mycoplasma by the vendor. 5-Fluorouracil (Cat. no.: 51-21-8; MCE) was dissolved in UltraPure™ Distilled Water (Cat. no.: 10977015; Invitrogen). The 3% hydrogen-peroxide solution (3%H2O2) was bought from Boster (Cat. no.: AR1108, China).
Cell transfection
Plasmid vector PLKO.1-puro was purchased from Synbio Technologies (China). The constructs of lncRNA FUAT1-shRNAs were chemically synthesized from RiboBio (China). The siRNAs specifically targeting lncRNA FUAT1, TNS4, NRF2, and miRNA mimics/inhibitors used in this study were also synthesized from RiboBio. The transfection experiments were performed by using Lipofectamine 3000 Transfection Reagent (Invitrogen) following the manufacturer's instructions.
RNA extraction and qRT-PCR
Total RNA was extracted from cells and tissue samples with Trizol reagent (Invitrogen) following the manufacturer's instructions. PrimeScript™ RT reagent kit (Perfect Real Time) (TaKaRa) was used for reverse transcription. Quantitative real-time PCR (qRT-PCR) was conducted with TB Green® Premix Ex Taq™ II (Tli RNaseH Plus) (TaKaRa). GAPDH and U6 were used as the endogenous controls. The primers were synthesized from Sangon Biotech (China) and the sequences information is listed in Supplementary Table S5.
Western blot assay
Cells were lysed in cell lysis buffer (Beyotime, China) for Western blot. The same quantity of total proteins was subjected to Western blot analysis. The antibodies used in this study include anti-TNS4 (11580-1-AP, Proteintech), anti-NRF2 (66504-1-Ig, Proteintech), and anti-β-actin (A5441, Merck).
ROS detection
SNU16 cells treated with different doses of 5-FU or H2O2 for 24 h were exposed to 5 μM 2′,7′-dichlorodihydrofluorescein diacetate (DCFH-DA) probe (Beyotime) for 20 min, then washed with serum-free 1640 medium for three times. Nonfluorescent DCFH is oxidized by intracellular ROS into fluorescent DCF. The fluorescence signals from DCF were detected by flow cytometer. Totally 1 × 104 cells were analyzed per sample.
Cell viability assay
The cell viability was conducted by using Counting Kit-8 (Japan). In brief, after the indicated siRNAs transfection for 16 h, cells (5 × 103 cells/well) were seeded into 96-well plates. After culturing for 12 h, cells were treated with 5-FU at the indicated concentration for another 24 h. Then the cell viability was assessed (10 μL/well) based on the absorbance at 450 nm with the SynergyTM H1 multimode microplate reader (BioTek) after adding CCK-8 solution. The dose–response curve was plotted using GraphPad Prism 6.0.
Apoptosis assay
After indicated treatments, cells were washed twice with cold PBS and then resuspended in 1X Binding Buffer (BD Pharmingen) at a concentration of 1 × 106 cells/mL. The cell suspension (100 μL, 1 × 106 cells/mL) was incubated with 5 μL of FITC Annexin V and 5 μL PI (BD Pharmingen) for 20 min at room temperature in the dark, when 200 μL of 1X Binding Buffer was added. Flow cytometry was employed to detect the apoptosis rate, and 1 × 104 cells were analyzed per sample.
RNA FISH
Spread 10 μL cell suspension (1 × 107/mL) evenly on lysine-coated glass slides and place it at room temperature for 5–10 min until the liquid is dry. lncRNA FUAT1 fluorescent probes labeled with Cy3 dye (RiboBio) and miR-140-5p fluorescent probes labeled with Cy5 dye were used according to the fluorescent in situ hybridization kit's instructions (C10910; RiboBio). Fluorescence signal is detected by a laser scanning microscopy (ZEISS LSM 880).
Dual-luciferase reporter assay
To evaluate the regulatory effects of miR-140-5p on FUAT1 and TNS4, dual-luciferase reporter assay was performed. In brief, the lncRNA FUAT1 sequence containing 776–787 bp and the 3′UTR of TNS4 containing 1051–1058 bp were synthesized by PCR and cloned between the SacI and SalI sites of the pmirGLO luciferase reporter plasmid (Synbio Technologies). The primer sequences specific to FUAT1 for the luciferase reporter assay: Forward, 5′-TTGA GCTCCTCAGCAAGCGATGCAAGAGAGGCTATGGC-3′, Reverse, 5′-TTGTCG ACCAGTTCCCTTCTGCTGCATGTCCAGCCACC-3′. The TNS4 3′-UTR primer sequences: Forward, 5′-TTGAGCTCGCTTCGGCCTCCCAAAGTGCTGGGATTA CAG-3′, Reverse, 5′-TTGTCGACACTGGGGCCTGTGACCTTGAGAACCTCATC-3′.
The mutant (miR140-5p targeting) luciferase reporter constructs of lncRNA FUAT1 and TNS4 3′UTR were generated by the Fast MultiSite Mutagenesis System Kit (TransGen). All obtained constructs were confirmed by DNA sequencing. lncRNA FUAT1/TNS4-WT/MUT was transfected into 293T cells with miR-140-5p mimics/NC for 48 h. The luciferase activity was detected using Dual Luciferase Reporter Gene Assay Kit (Yeasen, China) and the luciferase luminescence was normalized to Renilla luciferase luminescence.
RNA immunoprecipitation
The RIP assay was performed using a Magna RIP RNA-Binding Protein Immunoprecipitation Kit (Millipore) according to the manufacturer's instructions. In brief, the cell lysates were incubated with magnetic beads conjugated with human anti-Ago2 antibody or normal rabbit IgG control (Millipore). The immunoprecipitated RNAs were then extracted and detected by qRT-PCR.
In vivo xenograft mouse study
Four-week-old male BALB/c nude mice were purchased from Beijing Vital River Laboratory Animal Technology Co., Ltd. (China) and raised in SPF barrier facilities. Lentivirus-infected MKN45 cells (sh-NC/sh-lncRNAFUAT1) that stably expressed firefly luciferase were injected subcutaneously into the dorsum flank region of the mice (mice were divided into three groups randomly, 3 × 105 cells/100 μL per flank, n = 6 per group). One week later, the mice were treated with 5-FU at 25 mg/kg body weight via intraperitoneal injection twice a week.
The tumor volumes (V) were measured every 5 days using the formula: V = (length × width 2 )/2. At the 30th day postimplantation, each mouse was injected with luciferin (PerkinElmer) at 150 mg/kg body weight via the abdominal cavity, and 10 min later, tumor volumes were monitored and imaged by the bioluminescence imaging system (PerkinElmer). Subsequently, the mice were euthanized and the tumors were removed and weighed. All the procedures involving animals were complied with relevant ethical regulations, and the protocol was approved by the animal ethics committee of TMUCIH.
Bioinformatics
The study cohort consisted of stomach adenocarcinoma (GC) and COAD (colon cancer) patients selected and retrieved from TCGA (https://portal. gdc.cancer.gov/) for comprehensive integrated analysis. The patients with incomplete clinical information are not included in the study. This study was deemed exempt from institutional board approval and patient informed consent was waived, due to the retrospective nature of the study.
Statistical analysis
Statistical analyses and graphical presentation were performed by using GraphPad Prism 6.0 software and SPSS (version 22.0). Data are presented as the mean ± standard error of three or more independent biological replicates. Student's t test and χ 2 test were applied to evaluate the statistical significance. The correlation analysis was performed by Spearman's rank test and survival analysis was assessed with the log-rank test. p Value <0.05 was considered statistically significant.
Electronic laboratory notebook was not used.
Footnotes
Authors' Contributions
L.L. conceived and supervised the study and finalized the article. M.L. and X.L. participated in the study design and drafted the article. M.L. designed and performed most of the experiments and analyzed the data. X.L. and H.L. participated in analysis and interpretation of the clinical data. H.L. engaged in the mouse work. B.P., J.Z., Y.P., and M.S. contributed to the in vitro experiments. All authors read and approved the final article.
Ethical Approval and Consent to Participate
All procedures for the animal experiments were approved by animal ethics committee of TMUCIH (LLSP2019-170) and conducted according to the Basel Declaration. The study with human GC tissues was approved by the ethics committee of TMUCIH (bc2021092) and has been performed in accordance with the Declaration of Helsinki. Informed consents were signed by all involved patients.
Consent for Publication
Informed consents were signed by all involved patients.
Availability of Data and Materials
The data sets supporting the conclusions of this article are included within the supplementary files and other related data are available from the corresponding author (L.L.) on reasonable request.
Author Disclosure Statement
No competing financial interests exist.
Funding Information
This study was funded by the National Natural Science Foundation of China (no. 81572416), Haihe Laboratory of Cell Ecosystem Innovation Fund, the National Key Technologies R&D Program of China (no. 2016YFC1303200), and the TMUCIH Cancer Translational Medicine Seed Funds (no. 1701-1).
Supplementary Material
Supplementary Table S1
Supplementary Table S2
Supplementary Table S3
Supplementary Table S4
Supplementary Table S5
Supplementary Figure S1
Supplementary Figure S2
Supplementary Figure S3
Supplementary Figure S4
Supplementary Figure S5
Supplementary Figure S6
Supplementary Figure S7
Abbreviations Used
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.