
Abstract
Significance:
Cardiovascular disease is a major contributor to human mortality and morbidity. The cardiac tissue undergoes fibrotic healing after injury because of the limited regenerative capacity of adult mammalian cardiomyocyte (CM). Extensive research has been performed to identify therapeutic targets for CM regeneration, as the success of promoting adult human CM regeneration to repair the injured heart is considered the Holy Grail in the field.
Recent Advances:
To date, more than 30 target genes have been shown to regulate adult mammalian CM proliferation. More than 20 targets have been validated in adult mouse myocardial infarction (MI) model in a therapeutic setting. In this review, the translational efficacy readouts from 17 selected pharmaceutical targets are summarized, among which the Hippo-yes-associated protein (Yap) pathway is the most extensively investigated and fits the criteria for a promising target for pro-CM-regeneration therapy development.
Critical Issues and Future Directions:
As the pro-CM-regeneration potential of current drug treatment for cardiovascular patients is limited, to help identify and fill the gap between basic research and drug discovery in this specific field, details regarding target identification, validation in mouse MI models, high-throughput screening assay development, and preclinical in vivo efficacy model optimization are discussed. Finally, suggestions and recommendations are also provided to help establish a common guideline for in vivo translational studies for drug discovery focusing on CM regeneration. Antioxid. Redox Signal. 39, 1070–1087.
Introduction
Cardiovascular diseases (CVDs) have been the top killer worldwide for decades. Despite the advances in medicine, the survival rate of CVD patients who developed heart failure (HF) is only 50% after 5 years, worse than some cancers (Rodriguez-Artalejo et al., 2004). The enormous health care burden resulting from CVDs is expected to significantly increase with the expansion of the aging population, if no new breakthrough in effective therapies could be developed.
Cardiomyocyte (CM) loss is an important pathological feature of CVDs, especially in myocardial infarction (MI) (Tan et al., 2022). The adult heart in mammals has very limited regeneration capacity, therefore the injured heart undergoes fibrotic repair to replace the dead CMs (Bergmann et al., 2009; Prabhu and Frangogiannis, 2016). Effective regeneration of functional adult CM to repair the damaged myocardium and preserve cardiac function is considered the Holy Grail in the cardiovascular research field.
Several strategies have been investigated to promote new adult CM generation: adult cardiac stem cell differentiation into new CMs (Afjeh-Dana et al., 2022), endogenous proliferation from pre-existing CMs (Senyo et al., 2013), induced pluripotent stem cell derived CM (iPSC-CM) cell therapy (Shiba et al., 2016), and cardiac fibroblast to CM trans-differentiation (Cao et al., 2016; Ieda et al., 2010; Liu et al., 2017; Qian et al., 2012; Song et al., 2012) (Fig. 1).
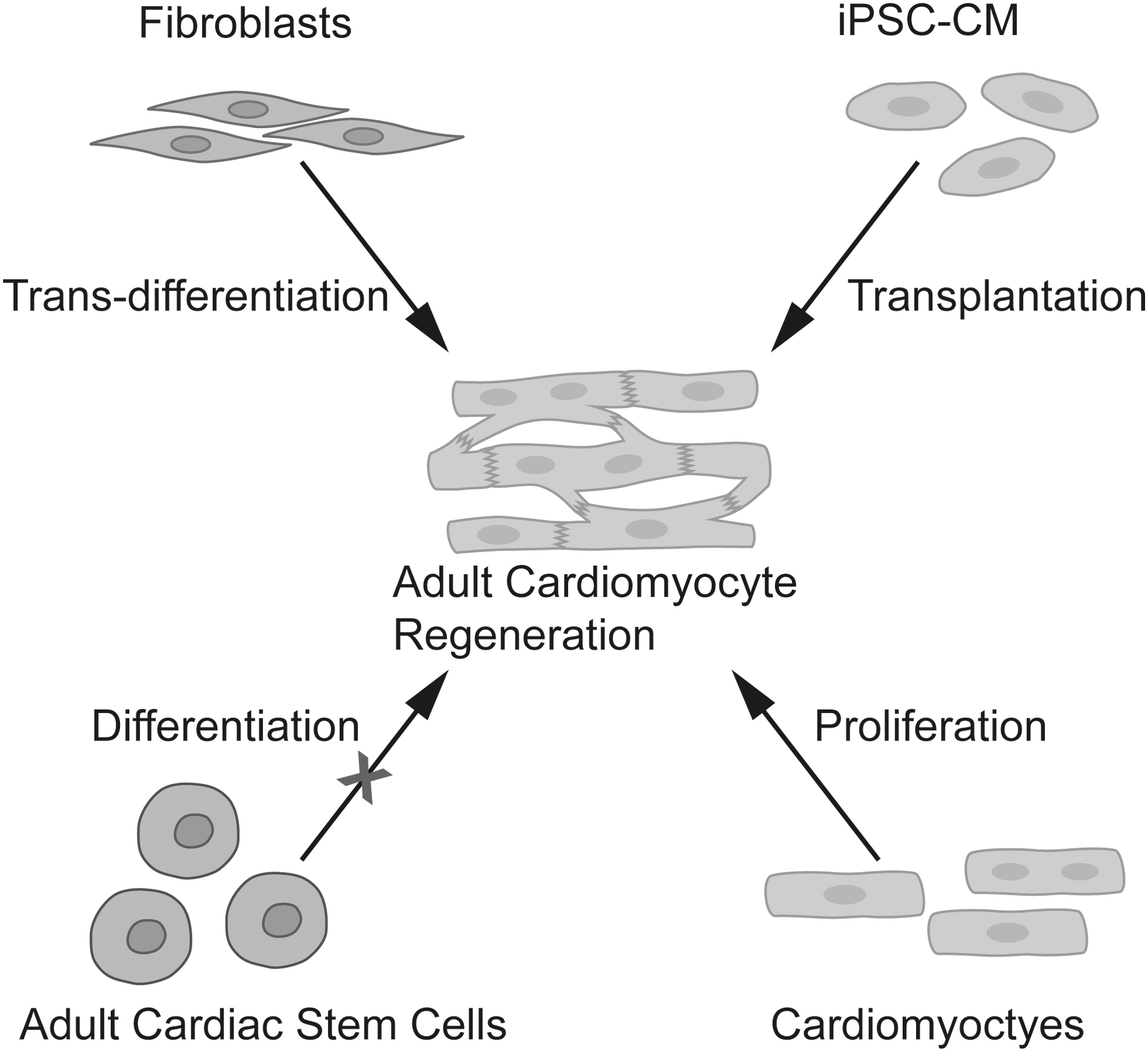
Summary of research strategies for adult CM proliferation. CM, cardiomyocyte. Color images are available online.
After more than two decades' basic research and clinic investigation, the field has now reached a common understanding that endogenous adult cardiac stem cells do not differentiate into CMs in vivo (Monroe et al., 2019). Cell therapy using iPSC-CM in non-human primates has shown cardiac regeneration potential post-MI but with severe barrier of life-threatening arrythmia waiting to be overcome (Shiba et al., 2016). Induction of endogenous cardiac fibroblast trans-differentiation into CM has been investigated majorly by either virus mediated genetic modification (Ieda et al., 2010; Qian et al., 2012; Song et al., 2012) or low molecular weight (LMW) compounds treatment (Cao et al., 2016). Promising trans-differentiation efficiency in vitro has been achieved, whereas further optimization of in vivo cell trans-differentiation efficiency and efficacy in cardiac regeneration are needed for translational purpose (Ricketts and Qian, 2022; Yamakawa and Ieda, 2021). In the current review, we will focus on the pro-CM-proliferation strategy that has been shown to be responsible for the naturally happened cardiac regeneration in zebra fish (Jopling et al., 2010) and the neonatal mouse heart (Porrello et al., 2011) after injury.
Before the 1990s, adult mammalian CM was considered post-mitosis and unable to generate new CM through proliferation. This prevailing dogma changed after the accumulation of evidence showing that adult mammalian CM can proliferate, although at a very low rate (0.5%–1% turnover per year in adult human hearts) (Bergmann et al., 2009). Although a low turnover capacity as such is not enough to counter the massive CM loss after MI, it is hoped that a deep understanding of the regulatory mechanisms of adult mammalian CM proliferation would help to develop new therapies driving the adult CM regeneration at a controllable, higher level for sufficient cardiac repair.
Successful new therapies, especially new drug development in CVDs, need to be based on solid preclinical research data, which includes target identification and validation, robust high-throughput screening assay for lead compound selection, favorable pharmacokinetic (PK) profile, approachable pharmacodynamic (PD) marker, delivery route selection, reliable efficacy studies in rodent and large animals, strict safety test, etc.
Valuable information from basic research publications on target identification, target validation in in vivo MI or ischemia-reperfusion (I/R) models, high-throughput CM screening assay development, and in vivo efficacy model optimization would significantly facilitate the often lengthy and costly drug development process in pharmaceutical companies. In this review, we will mainly discuss how the current knowledge on adult CM proliferation could be translated into potential clinical therapy. We will focus on the targets that have been evaluated in a therapeutic setting via adenovirus-based delivery or direct treatment in the adult mammalian MI model and discuss the translational gaps and possible ways to fill them.
Potential Therapeutic Targets for Adult CM Proliferation and Cardiac Function Preservation
New drug development for CVDs is well known to be extremely time- and resource-consuming, more so than most cancer drugs (Fordyce et al., 2015). With the handful potential targets for CM proliferation, how to choose the most promising one is the first key thing that drug hunters need to carefully consider. To date, more than 20 potential targets for CM regeneration have been shown in a therapeutic setting to be able to promote adult mouse CM proliferation and preserve cardiac function post-MI (Li et al., 2021; Naqvi et al., 2021; Neininger et al., 2019).
These targets cover a wide range of molecular categories: cell cycle regulators, transcription factors, microRNAs (miRs), soluble factors, extracellular matrix components, and signaling pathway elements (Fig. 2). Due to the safety concern of cell cycle regulators (Abouleisa et al., 2022) and the low feasibility to pharmaceutically targeting transcription factors, targets from those two categories are not discussed in the current review. According to our searching results in Pubmed after excluding studies using the criteria mentioned above, to date, about 17 targets for adult CM proliferation have been investigated in preclinical translational studies (treatment after ischemia injury) by adenovirus-based gene therapy or direct treatment.

Summary of published targets and pathways that could promote adult mammalian CM proliferation. Color images are available online.
Here, we summarized the therapeutic features of these targets based on the following features: (1) potency to promote adult rodent CM proliferation post-MI in vivo (Table 1); (2) efficacy in the preservation of cardiac function in the MI or I/R model (Table 1); (3) long-term and short-term safety concern; and (4) validation in large animals.
Therapeutic Efficacies of Different Targets for Cardiomyocyte Proliferation in Adult Myocardial Infarction Mice
Most of the results were estimated from the bar plots in the original studies.
WGA staining was used for cell counting in this study.
The CMs in the MI border zone were used for cell counting.
The CMs in the infarct area were used for cell counting.
CM, cardiomyocyte; MI, myocardial infarction; Yap, yes-associated protein; EdU, 5-ethynyl-2′-deoxyuridine; AAV, adeno-associated virus; WGA, wheat germ agglutinin; TT-10, the pharmaceutical product TT-10 (C11H10FN3OS2); GFP, green fluorescent protein; Sav, Salvador; pHH3, phosphohistone H3; Pi3kcb, phosphatidylinositol-4,5-bisphosphate 3-kinase; shRNA, short hairpin RNA; Sdh, succinate dehydrogenase; BrdU, bromodeoxyuridine; Mapk, mitogen-activated protein kinase; PBS, phosphate-buffered saline; miR, miRNA; Fgf1, fibroblast growth factor 1; Lrp6, low-density lipoprotein receptor-related protein 6; Mydgf: myeloid-derived growth factor; Nrg1, neuregulin 1; Erbb, Erb-B2 receptor tyrosine kinase; Osm, oncostatin M; gp130, glycoprotein 130.
In vivo potency to promote adult CM proliferation
When ranking the targets with potency based on the in vivo CM proliferation rate in the border zone of rodent MI hearts, inconsistencies caused by different histology standard and readout for proliferating CM identification and quantification make it a very challenging task. As highlighted in a recent review by Achampach et al. (2022), it would be greatly appreciated if the field of heart regeneration could apply a common experimental standard for CM proliferation or cell cycle re-entry identification and quantification.
In our previous study, we have used a combination of four fluorescent staining [Hoechst, alpha-actinin, phosphohistone H3 (pHH3), wheat germ agglutinin (WGA)] together with Z stack imaging to identify cell cycle marker positive CM in the adult mouse heart (Xiang et al., 2016). The WGA membrane staining is critical to distinguish non-CM adjacent to or vertically overlapping a CM. Ang et al. (2010) performed an analysis to evaluate how much WGA staining affects CM identification when combined with sarcomere markers. They showed that the use of WGA improved the overall positive predictive power (the probability of a positive CM nuclear identification) from a range of 42%–58% to a range of 85%–88% (Ang et al., 2010). Given that there are normally just a few cell-cycle marker positive CMs within each observation field of myocardium tissue, this would make WGA a key experimental factor to accurately identify CM. Other studies have also used pericentriolar material 1 (Pcm1) to specifically mark CM nuclei as this protein localizing at the perinuclear space of adult CM (Bergmann and Jovinge, 2012; Ng et al., 2020). It is still not clear whether Pcm1 will remain perinuclei in a mitotic CM, so there might be a risk of underestimation of true mitotic CM.
In the translational studies listed in Table 1, we summarized the in vivo CM proliferation rate quantified by cell cycle marker labeling. Notably only several targets, including yes-associated protein (Yap), p38 mitogen-activated protein kinase (p38 Mapk), fibroblast growth factor 1 (Fgf1), miR-34a, miR-302-367, and miR-199a, were validated via methodology involving sarcomere marker together with cell membrane staining (WGA or Caveolin3) to distinguish CM and non-CM for proliferating CM quantification.
Various cell cycle markers such as bromodeoxyuridine (BrdU), 5-ethynyl-2′-deoxyuridine (EdU), Ki67, pHH3, and Aurora B kinase, which represent different stages of the cell cycle, have been used. Although true CM dividing could not be fully evidenced by these markers, increased activation of cell cycle activity in adult CM represents a trend for cell division and might be a reparative cascade trigger regulating the following healing process, as almost all the targets that can promote adult CM cell cycle activity also showed enhanced cardiac repair and preserved function (Table 1).
All the targets listed in Table 1 have been reported to significantly increase CM cell cycle activity in the MI rodent heart and preserve cardiac function (Table 1). The evaluation time for CM cell cycle activity ranges from 5 days to 12 weeks post-MI. Most target treatments showed a significant increase of two-to-nine-fold increased CM cell cycle activities.
Periostin therapy (Kühn et al., 2007) was reported to induce a 60-fold increase (0.01% to 0.6%) of BrdU-positive CM in rat hearts 1-week post-MI. The readout from their control group seems much lower compared with the baseline level reported by all other studies (0.01% vs. 0.04%–0.8%), although data were quantified at different time points or by EdU instead of BrdU. Further, the effect of periostin on regulating CM proliferation remains debated. A genetic study performed by Lorts et al. (2009) showed that periostin overexpression did not increase CM proliferation after MI in mice. Another target that has shown conflicting results from different therapeutic studies is neuregulin 1 (Nrg1).
A study conducted by Reuter et al. (2014) did not find increased CM proliferation with 7 daily injections of recombinant Nrg1β1 post-MI in mice. Zurek et al. (2020) also reported that Nrg1 treatment induces hypertrophy and worsens cardiac performance in a rat MI model. On the other hand, the Hippo-Yap pathway has shown consistent efficacy in promoting adult mammalian CM cell cycle activation post-MI in multiple studies from different laboratories (Chen et al., 2021; Leach et al., 2017; Lin et al., 2015; Lin et al., 2014; Liu et al., 2021; Monroe et al., 2019; von Gise et al., 2012; Xin et al., 2013).
Efficacy in cardiac function preservation
Cardiac function is one of the major readouts for clinical study related to ischemic heart disease (IHD), especially the change of ejection fraction (EF) in response to the treatment. Interestingly, all the targets listed in Table 1 that can promote adult CM cell cycle activity also showed an improvement of cardiac function about 5%–15% unit increase of fraction shortening (FS) post-MI measured by M-mode echocardiography (echo) when treatments were performed right after MI. These findings suggest that pro-CM-regeneration therapy might be beneficial in preserving cardiac function in the sub-acute stage post-MI via promoting CM cell cycle activation during the endogenous wound-healing process.
Interestingly, Leach et al. (2017) showed significant improvement in cardiac function when starting the adeno-associated virus 9-Salvador-short hairpin RNA (AAV9-Sav-shRNA) treatment 3 weeks post-MI when the infarct area has normally matured into a stiff fibrotic scar. The efficacy of AAV9-Sav-shRNA was further evaluated in the pig I/R model. AAV9-Sav-shRNA was injected at 2 weeks after I/R in pigs and showed significant preservation of cardiac function measured by magnetic resonance imaging (MRI) 3 months after I/R.
Salvador homolog (Sav) is one of the critical components in the Hippo-Yap signaling pathway. These exciting findings have revealed the potential of therapies targeting the Hippo-Yap pathway in treating both sub-acute and chronic MI patients.
Safety concern
Two potential safety concerns related to the pro-CM-proliferation treatments have been discussed: pathological cardiac remodeling and on-target oncogenesis. Long-term safety evaluation is only carried out on two targets in genetic models and treatment studies. For the Hippo-Yap pathway, Lin et al. (2014) found that long-term Yap overexpression did not cause cardiac pathological hypertrophy but induced a slight, statistically significant increase in myocardial fibrosis.
Myocardial injection of AAV9-Yap did not lead to liver tumors up to 23 weeks as described by Lin et al. (2014) but transgenic expression of YAP5SA, which genetically maximizes the Yap activation by mutation of all Last1/2 phosphorylation sites from S to A, caused CM hyperplasia and early death in mice (Monroe et al., 2019). Moreover, virus-mediated expression of miR-199a in mice and pigs caused arrhythmia and sudden death (Eulalio et al., 2012; Gabisonia et al., 2019). Thus, transient therapy with adjustable administration doses and treatment windows is needed to avoid potential deleterious or tumorigenic side effects from pro-CM-regeneration therapy.
Validation in large animals
Efficacy of therapeutic strategies targeting the Hippo-Yap signaling pathway and miR-199a has been validated in the pig I/R model. Liu et al. (2021) showed that inactivation of Sav via AAV9 delivery 2 weeks after I/R increased CM cell cycle activation and preserved cardiac function 3 months after I/R in 3-month-old pigs. Moreover, compared with the low-dose AAV9-Sav-shRNA, the high-dose AAV9-Sav-shRNA gene therapy showed a greater number of EdU+ or pHH3+ CMs but no difference in cardiac function preservation (Liu et al., 2021).
miR-199a gene therapy is also validated in the pig I/R model. Gabisonia et al. (2019) showed that AAV6-miR-199a treatment increased the percentages of Ki-67+, BrdU+, and pHH3+ CMs and preserved EF in adult pigs after I/R. However, 7 out of 10 pigs died around 7–8 weeks after I/R due to tachyarrhythmia.
In summary, among the 17 potential therapeutic targets for adult CM regeneration, the Hippo-Yap signaling pathway is currently the most promising target for pro-CM-regeneration therapy according to the following characterizations: (1) good efficacies in increasing adult mammalian CM cell cycle activation; (2) preserving cardiac function post-MI in rodent, as reported by multiple laboratories; (3) limit long-term side effect in genetic studies; and (4) efficacy validated in large animal models.
Moreover, the molecular function connection between these 17 targets is investigated and shown in Figure 3. The protein-protein interaction network was generated using the STRING platform. All druggable targets were clustered based on the Markov Cluster Algorithm (with an inflation parameter of 3). Different colors in this figure refer to different clusters. Three major connection clusters are found: cell cycle regulators, Nrg1/Erb-B2 receptor tyrosine kinase 2 (Erbb2) network, and the Hippo-Yap pathway.

Interaction of functional connection between published therapeutic targets that could promote adult mammalian CM proliferation. Three major connection clusters are found: cell cycle regulators, Nrg1–p38 network and Hippo/Yap pathway. Nrg1, neuregulin 1; Yap, yes-associated protein. Color images are available online.
Interestingly, the Hippo-Yap pathway is involved in the pro-CM proliferation mechanism of other six listed targets, including miR-199a (Gabisonia et al., 2019), miR-302-367 (Tian et al., 2015), progesterone (Lan et al., 2020), phosphatidylinositol-4,5-bisphosphate 3-kinase (Pi3kcb) (Lin et al., 2015), Agrin (Bassat et al., 2017), and oncostatin M (Osm) (Li et al., 2020).
More importantly, there are plenty of druggable kinases in the classic Hippo-Yap pathway, such as macrophage stimulating (Mst) 1/2, Sav, Mps one binder kinase activator 1 (Mob1), and large tumor suppressor kinase (Lats) 1/2, which is suitable for LMW drug development. With all the published data and characterization, the Hippo-Yap pathway has emerged as an intriguingly attractive therapeutic target for drug development of pro-CM-regeneration therapy.
Hippo pathway in heart regeneration
The Hippo-Yap pathway is an evolutionarily conserved pathway involved in various physiological and pathological processes (Edgar, 2006; Varelas, 2014). The Hippo is an STE20 protein kinase family member first studied in Drosophila melanogaster. In mammals, the following components consist of the core Hippo signaling: Mst1/Mst2, Sav1, Lats1/Lats2, and Mob1 (Wang et al., 2018). Mst1, Mst2, and Sav1 form a complex that phosphorylates Lats1 and Lats2 kinases interacting with the cofactor Mob1, which further phosphorylate and degrade the transcriptional coactivators Yap and PDZ-binding motif (Taz). However, if the Hippo signaling is repressed, Yap and Taz can translocate into the nucleus, where it can bind to different transcription factors to regulate the transcription of downstream genes (Wang et al., 2018).
The Hippo-Yap pathway plays a crucial role in regulating cardiac development, growth, homeostasis, disease, and regeneration as nicely reviewed by Wang et al. (2018) and Zheng et al. (2020). As discussed earlier, this pathway is currently the most potential target for CM regeneration. Blocking the Hippo signaling or activating Yap in CMs can promote cell renewal. A thorough understanding of the downstream genes or biology processes regulated by the pathway in the heart is essential for cardiac regeneration drug development.
After Yap is activated, it will translocate into the nucleus and bind with its cofactors [TEA domain transcription factor family members (Zhao et al., 2008)] or partner transcription factors [such as pituitary homeobox 2 (Tao et al., 2016), forkhead box protein O1 (Shao et al., 2014)], to regulate the transcription of target genes that can encode proteins regulating cell proliferation, survival, differentiation, migration, and metabolism (Liu and Martin, 2019).
The reported target genes for cell cycle induction or cell proliferation include: Aurka, Aurkb, Ccna2, Cdc2, Cdc20, Cdk1, Lin9, Ccna2, Ccnb1, Ccnd1, Ccnd2, E2f1, E2f2, Plk1, Myc, and p38 (Lin et al., 2014; Monroe et al., 2019; Wang et al., 2018). Meanwhile, microRNAs have also been identified as a downstream target of Yap (Yang et al., 2015b).
In summary, the Hippo-Yap pathway plays a pivotal role in CM regeneration. The components of Hippo and Yap protein can be regulated by different factors, which affect the translocation of Yap in nucleus and ultimately change the expression of target genes to promote cell regeneration (Fig. 4).

The Hippo-Yap pathway participates in various biological processes in the heart and plays a pivotal role in cardiomyocyte regeneration. The components of Hippo and Yap protein can be regulated by different factors, which affect the translocation of Yap in the nucleus and ultimately change the expression of target genes. Color images are available online.
Potential Effects of Current Pharmaceutical Treatment for IHD Patients on CM Proliferation
The current pharmaceutical therapy for IHD patients includes antiplatelet, anticoagulation, vasodilation, antihyperlipidemic, antidiabetic, and antihypertension. We did a scientific search on the majority of the drugs used for IHD patients and found that several have been shown to increase CM proliferation in animal models (Fig. 5).
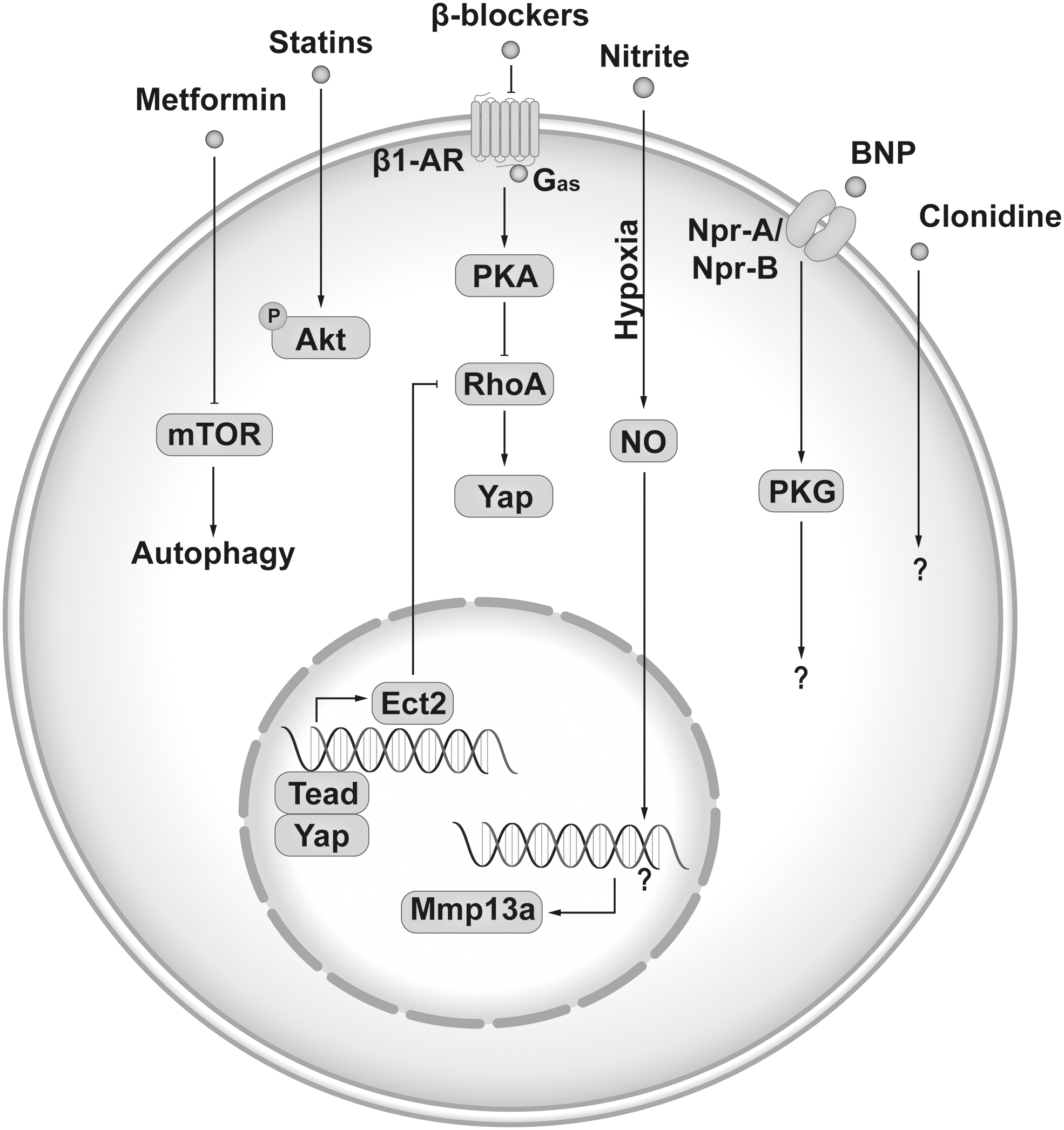
Summary of pharmacotherapy used in IHD treatment that could promote CM proliferation. IHD, ischemic heart disease. Color images are available online.
Nitrates are a front-line treatment for angina. They are a group of medications that cause vasodilation, including venous vessels, coronary arteries, and small arterioles by donating nitric oxide (NO). In zebrafish hearts, physiological levels of nitrite treatment at hypoxia condition induced about 50% increase of proliferating cell nuclear antigen (PCNA) positive CM at day 5 after cryoinjury or apex amputation but showed no effects at normoxia condition (Rochon et al., 2020). The underlying mechanisms are related to NO-dependent immune cell recruitment. But the role of nitrite in mammal CM proliferation was not explored. Another drug that could induce zebrafish CM proliferation is metformin, which is the main first-line medication for type 2 diabetes. In the zebrafish cardiac cryoinjury model, metformin treatment has been shown to induce a three-fold increase of PCNA-positive CM 4 days after injury via enhanced autophagy (Xie et al., 2021). Whether metformin could induce a similar effect in mammal CM proliferation is unknown.
Among the four drugs that showed the potential to increase mammalian CM cell cycle activities, statin is the only one that had effects on adult rat CM proliferation in the MI heart. Treatment with 20 mg/kg/d Rosuvastatin in drinking water starting 2 weeks before MI induced a three-fold increase of BrdU-positive CM (0.95%–3.4%) at border zone 28 days post-MI in female rats (Cianflone et al., 2020). To note, the identification of CM in this study is only based on α-sarcomeric actin staining without WGA co-stain, which might cause a false positive. Further in vivo testing with therapeutic design (start treatment after MI) and more convincing evidence of CM cell cycle activity or division would help to validate the potential of statin on mammalian CM proliferation.
Further, clonidine, brain natriuretic peptide (BNP), and propranolol have been shown to increase CM cell cycle activity or population when starting the treatment within 1 week after birth in mice (Bielmann et al., 2015; Liu et al., 2019; Murganti et al., 2022). Clonidine is an α2-adrenergic agonist medication used to treat high blood pressure, attention deficit hyperactivity disorder, drug withdrawal, menopausal flushing, diarrhea, spasticity, and certain pain conditions. In a troponin T2-fluorescent ubiquitination-based cell cycle indicator (TNNT2-FUCCI) human iPSC-CM based high-throughput screening with an autophagy compound library, clonidine was identified to increase proliferation in human iPSC-CMs (Murganti et al., 2022). The effects of clonidine on CM proliferation were further tested in vivo. When given along with EdU daily to mice from postnatal day 1 to 5, clonidine induced about 40% increase of EdU+ CM nuclei (EdU+/Pcm1+) but the percentage of mononucleated CM decreased, suggesting mitotic activity without cytokinesis. The role of clonidine in adult mammal CM proliferation was not explored.
BNP, a hormone secreted by cardiac cells, is used in hospitalized patients with acutely decompensated congestive HF with dyspnea at rest. It causes potent vasodilation, resulting in significant reductions in venous and ventricular filling pressures and mild increases in cardiac output. It was reported to induce a 34% increase of CM total number after 2-week (1 μg/mouse) treatment starting from postnatal day 3 in mice (Bielmann et al., 2015), whereas whether BNP treatment in adult mice could also induce CM population increase remains unknown.
Intriguingly, β-blocker propranolol, which is used to treat tremors, angina, hypertension, heart rhythm disorders, and other heart or circulatory conditions, has been shown to increase CM division through promoting cytokinesis in neonatal mice (Liu et al., 2019). Propranolol daily administration starting from postnatal day 1 for 1 week in mice induced a 30% increase of CM population by the activation of downstream cytokinesis gene ect2. A recent study also showed that metoprolol, a cardio-selective β blocker for β1-adrenergic receptor (β1-AR), promotes cardiomyocyte proliferation in juvenile mice (< 1 month old) through activation of an RhoA-Yap signaling axisin (Sakabe et al., 2022). Moreover, analysis of heart tissue from an infant with tetralogy of Fallot with pulmonary stenosis labeled with isotope-tagged thymidine demonstrated that CM cytokinesis failure is increased and is associated with ect2 repression. Based on this exciting finding, a phase I trial of propranolol hydrochloride NCT04713657 has been registered to evaluate the potential effects of propranolol on human CM division in infants with tetralogy of Fallot (El Khoudary et al., 2021).
In summary, the potential of current pharmacological therapy on driving CM proliferation still needs to be further investigated. Most of the drugs listed in Figure 5 have already been widely used in HF patients and the beneficial effects if including CM proliferation have contributed to the overall outcome. Currently, the 5-year survival rate of HF patients is only about 50%, and the huge unmet medical needs urgently call for novel therapy discovery and development.
Pro-CM-Regeneration Target Discovery and Drug Development via High-Throughput CM Screening
High-throughput screen (HTS) is the cornerstone of modern drug discovery. With the capability to test a large number of small molecules, peptides, and antibodies in a reasonable period, it has demonstrated its crucial utility and immense power of identifying lead molecular entities for further optimization, therefore laying down a solid foundation for drug discovery and development projects.
In recent decades, we are seeing the increased use of robotics and automation to aid workflow and speed up the process, the shrinking of assay dimensions to fit in denser plates (384-well or 1536-well plates), automated data collection and curation systems, as well as artificial intelligence assisted data analysis pipelines. All these technological advances pushed the sophistication and efficiency of HTS to an unprecedented level.
This general trend has continued in the CM regeneration field. In the past decade, several reports emerged using HTS to search for small molecules and biologics that modulate the CM cell cycle and regulate their proliferation. Researchers from academic institutions and pharmaceutical companies utilized a variety of cells as their model to design, validate, and execute the screen, including induced human pluripotent stem cell-derived CM, and primary CMs from mice and rats.
The readout chosen by the individual teams ranges from rather basic CM nuclei counting, EdU/Ki67/pHH3 staining to sophisticated genetic labeling systems such as FUCCI and mosaic analysis with double markers (MADM). The size of library varies from tens to tens of thousands, and molecular entities sampled range from LMW compounds and miRNAs to secreted proteins. A summary of the primary screenings reported, detailing the experiment design, assay window, and the top selected molecules can be found in Table 2.
Summary of the Reported Primary High-Throughput Screenings for Pro-Cardiomyocyte-Regeneration Target Identification
α-MHC, alpha myosin heavy chain; Ampk, adenosine 5′-monophosphate-activated protein kinase; Cfse, carboxyfluorescein succinimidyl ester; Erk, extracellular regulated protein kinases; FUCCI, fluorescent ubiquitination-based cell cycle indicator; Gsk, glycogen synthase kinase; iPSC-CM, induced pluripotent stem cell-derived cardiomyocyte; Mapk, mitogen-activated protein kinase; mTori, mammalian target of rapamycin inhibitor; NC, negative control; PC, positive control; Raf, rapidly accelerated fibrosarcoma; TNNT2, troponin T2; Jak1, Janus Kinase 1; Dyrk, dual-specificity tyrosine-regulated kinases; Pten, phosphatase and tensin homolog; Mct1, monocarboxylate transporter 1; FBS, fetal bovine serum; DMSO, dimethylsulfoxide.
With the development of iPSC and reprogramming techniques, human iPSC-CMs become a favorable cell type for high-throughput screening for cardiovascular drug development. In recent years, human iPSC-CMs are widely used for the safety analysis of compounds in cardiac electrophysiology. Several clinical translatable assays were developed, which makes it a suitable counter-screening assay to predict the potential arrhythmia induction risk of lead candidates (Gintant and Traebert, 2020).
As human iPSC-CMs exhibited an immature phenotype and consisted of a mixed population of cardiac cell types (ventricle, atria, and nodule) (Ma et al., 2011), the clinical translatability of this cellular system for CM proliferation drug discovery is not yet fully established. However, the advantage of human origin makes it a necessary counter-screening assay to characterize pro-CM-regeneration hits from primary CM-based screening.
Notably, miR-199 was discovered as a potent hit for CM proliferation from high-throughput neonatal rat CM (NRCM) phenotypic screening. The five- to six-fold increase of EdU/Ki67-labeled NRCM proliferation induced by miR-199 in vitro (Eulalio et al., 2012) (Table 2) was successfully translated into a three- to four-fold increase of pHH3 positive adult CM (Table 1) in pig I/R hearts with cardiac function preservation (Gabisonia et al., 2019).
Success validation of the microRNA hits from phenotypic screening based on primary NRCM proliferation in in vivo mouse and pig cardiac ischemia model has provided valuable insights into the feasibility and translatability of pro-CM-regeneration target discovery via high-throughput phenotypic screening. Recently, Carrillo Garcia et al. (2022) developed a postnatal day 1 and day 6 rat/mouse cardiac cell based high-throughput assay for compound screening in a 384-well plate format and screened a library of 26 small molecular kinase inhibitors at three doses. A series of med-chem studies were performed based on the primary hits. However, the dose reaction curve generated from the top hits seems to be challenging for half maximal effective concentration (EC50) curve fitting with such a small assay window and big variability. Like what we found during the optimization of the NRCM high-throughput screening assay at 384- or 1546-well format, the small assay window of CM proliferation (highest readout is about three- to four-fold increase) makes the following EC50 analysis and structure activity relationship inventory very difficult but is valuable for target identification.
Intriguingly, Du et al. (2022) recently reported the synergistic effects of candidate small molecules from an NRCM high-throughput assay based on FUCCI cell cycle labeling system. A cocktail of five molecules targeting α1 adrenergic receptor, Janus Kinase 1 (Jak1), dual-specificity tyrosine-regulated kinases (Dyrk), phosphatase and tensin homolog (Pten), and monocarboxylate transporter 1 (Mct1) has shown a 30-fold assay window. A MADM system was also integrated as a secondary assay to visualize true CM dividing. Several counter assays, including human atrial CM and fibroblast, have been applied to demonstrate the selectivity and translatability of the top hits. Importantly, the 5-compound cocktail has been validated in an vivo mouse MI model and showed about a 2–3.6-fold increase in CM with cell cycle activation. The underlying mechanism is associated with lactate-LacRS2 signaling (Du et al., 2022). This is the first academic report of developing a compound cocktail from primary CM culture-based high-throughput screening for pro-CM-proliferation LMW drug discovery. The big difference between in vitro assay readout and in vivo results might be due to the unoptimized PK profile. Also, the combination treatment of LMW compounds would require a much more complicated med-chem optimization process and has a much higher chance of safety concern.
In summary, primary NRCM-based high-throughput screening has been shown to be suitable for pro-CM-regeneration target identification. The translatability could be evaluated in human CM-based counter-assays. However, based on the published data as well as our experience, for LMW drug development, more robust assays of NRCM and human iPSC-CMs proliferation with larger assay windows need to be further explored.
Preclinical In Vivo Efficacy Model for IHD Targeting CM Regeneration and Repair
There has been an increasing awareness and recognition of the necessity of establishing a standard guideline for in vivo mouse MI studies to allow interpretation and comparative examination across published studies. Among an extensive pool of published studies, there are significant variabilities in experimental design, often with unclear exclusion criteria (Lindsey et al., 2021). This makes it very difficult to compare the targets on a level playing field and evaluate the relative efficacy of the proposed therapies.
A comprehensive and detailed guideline for in vivo mouse MI experimental design has been presented by Lindsey et al. (2021; 2018), aiming at improving the reproducibility and translatability of cardiovascular research. Many lessons were learned from the cardio-protection research field, as none of the potential targets, which showed significant effects in infarct size limiting in animal models, successfully translated into clinics (Heusch, 2017; Heusch and Rassaf, 2016) majorly due to a lack of common standard methodology and shared guidelines for experimental design.
The importance of selecting a suitable and reliable in vivo model for IHD has been emphasized in several recent reviews (Lindsey et al., 2021; Lindsey et al., 2018). For early stage target validation or lead compound optimization, mouse models are still the best choice mainly due to the ease of genetic manipulation and relatively low cost compared with large animals. Mouse cardiac I/R and MI models are the two most widely used models in the field. For preclinical data collection to evaluate a potential new CM regeneration therapy, major readouts would include animal survival, cardiac function preservation, CM proliferation rate, and cardiac repair index (vessel density, level of pathological hypertrophy, non-infarct area fibrosis severity, etc.).
According to the differences between I/R and MI models in mice (De Villiers and Riley, 2020), I/R model normally results in variable small infarct size, high survival rate, and minimum cardiac function impairment. For efficacy evaluation in cardiac repair after myocardial ischemia, the MI model induced by permanent ligation of the left anterior descending (LAD) coronary artery would provide more consistent results in infarct size and cardiac function, which makes it more suitable for initial target validation and lead compound optimization. However, the evaluation of a compound's effect on cardio-protection in acute I/R settings will eventually be required to provide comprehensive information of the potential cardiovascular effects from the drug candidate.
Although target validation in MI model is the foundation of a CM regeneration drug discovery project, the lead compound identification and optimization stage after target validation normally requires effort of an order of magnitude more, as each round of optimization would need to be done in the same model with different doses and proper controls. Given the significant time and resources required by using the mouse MI model for target validation and drug optimization, a rigorous experimental design must be employed to ensure reliable and replicable results and efficient progression of drug discovery projects.
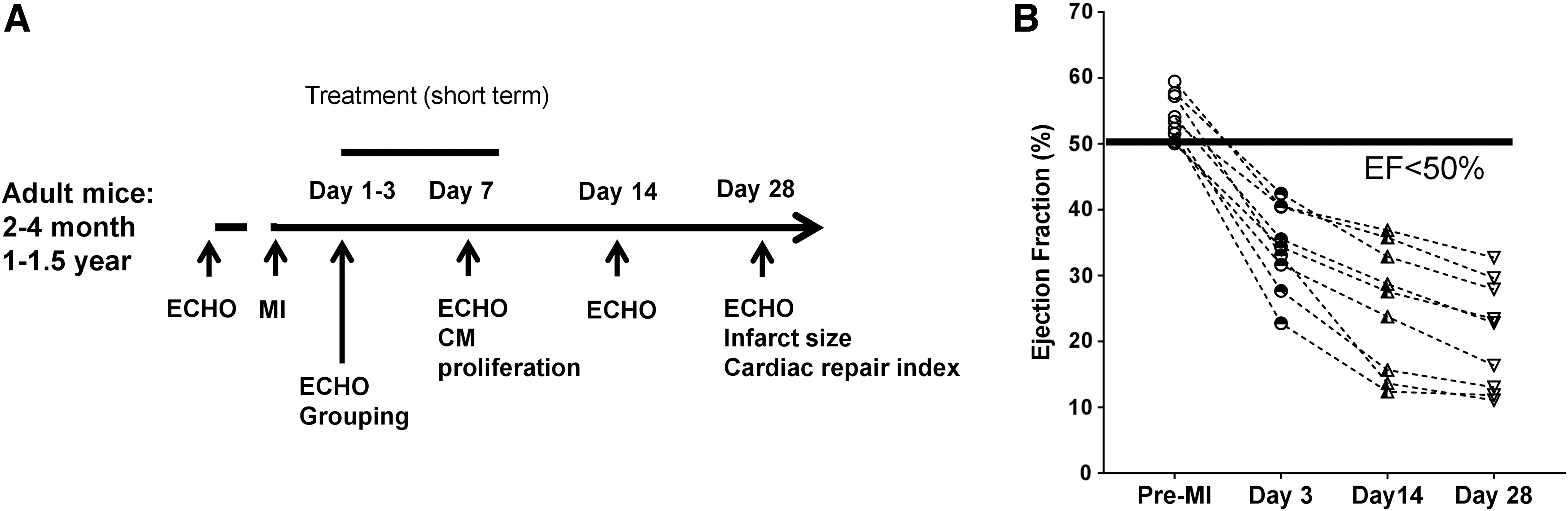
From our experience as well as highlights from several recent reviews (Lindsey et al., 2021; Lindsey et al., 2018), the following elements or recommendations could be considered in MI experiment design to validate the potential translatability of a target for CM regeneration: Mouse selection Most published studies have used young adult male mice for assessment of pro-proliferation potency. It is known that cell regenerative capacity decreases with aging (Bergmann et al., 2009). The majority of the patients with CVDs are from the aged population, so validation of efficacy in aged mice is necessary to provide additional insight in CM regeneration therapy development. Usage of both young adult (2–4-month-old) and aged mice (1–1.5-year-old) (Fig. 6A) would allow evaluation of efficacy differences in age or discovery of age-dependent regulators for CM regeneration. Gender differentiation in disease outcome has gained a lot of attention in the past decade (Arosio et al., 2022). Despite the social, cultural, and economic factors, how much the biological difference contributes to gender-related CVD outcome remains unclear and future studies to elucidate this important issue are warranted. However, female mice have a better survival rate and lower rupture incidence post-MI compared with males (Lindsey et al., 2021), which makes the male mice a preferred selection for studies expecting a beneficial effect in MI. A strategy to balance this conflict is to perform the initial target validation in male mice but evaluate therapeutic efficacy in both genders. Intervention or treatment time window The healing process of MI in the mouse heart has been comprehensively reviewed by Prabhu and Frangogiannis (2016). Briefly, the first 24 h post-MI is the peak of cell death followed by about 3 days' inflammation storm to clear out the dead cells and debris. Starting from day 3 post-MI in mice, reparative and proliferative phase emerges to heal the damaged heart with early granulation tissue formation composed of edema, capillaries, fibroblasts, macrophages, polymorphonuclear leukocytes, and lymphocytes. Around day 7 post-MI, the granulation tissue becomes dense and continues maturation in the next 2 weeks to form a stiff fibrotic scar. Based on dynamic changes of the endogenous healing process, pro-regeneration intervention starting around day 3–7 post-MI would be optimal to avoid the initial inflammation storm and prime the endogenous reparative signaling (Fig. 6A). Plenty of inducible genetic manipulation mouse lines and adenovirus-based gene therapy have been developed and used to successfully trigger the target expression during the reparative phase post-MI (Table 1). At 3–4 weeks post-MI in mice, the infarct area gradually become a paper-thin stiff fibrotic scar and interstitial fibrosis can be observed in the non-infarct myocardium. One would assume that repair or regeneration in such a fibrotic microenvironment could be much less effective. However, Leach et al. (2017) have tested a YAP-based therapeutic window from 3 to 6 weeks post-MI and observed a significant recovery of cardiac function, suggesting that pro-CM-regeneration therapy might also be effective for chronic MI. More related research is warranted to define the optimal treatment window for pro-CM-regeneration therapy. Quality of the MI surgery Although quality of the MI model is essential for scientific studies in the MI field, it is not emphasized enough in the published studies. The procedure of MI surgery has been introduced in detail (Kolk et al., 2009). It is a procedure that has been used for over 60 years (Johns and Olson, 1954) and requires a high level of surgical skills. Quick exposure of the location of LAD root and ligating the LAD while the heart is beating at 450–550 bpm is challenging. The major factor that affects the quality of a MI surgery is how experienced the microsurgeons are. Normally it takes about 1 year for a technician with previous microsurgery experience to generate a reproducible consistent level of infarction in mice. Besides the consistency of infarct size, there are other critical factors related to the surgery quality, including short surgery (about 5–8 min) and chest-open time (about 3 min), no or minimum bleeding, leakage free after rib cage closure, and minimum wound infection post-MI. These factors also directly affect the survival and outcome of the MI. But one must keep in mind that even the best surgeon would have about 10%–15% failure in MI induction (unpublished evaluation based on three surgeons with more than 10-year MI modeling experience) and could generate infarct size range from 20% to 60% of left ventricle due to the heterogeneity of the coronary anatomy (Chen et al., 2017; Kumar et al., 2016). The potential MI surgery quality issue needs to be aware of and sample size should be calculated based on the variability in infarct size from the surgeon before conducting an in vivo MI study. Exclusive criteria, animal grouping, and data presentation According to the guideline presented by Lindsay et al. (2021; 2018), including MI mice with infarct size ≥30% at day 1–3 post-MI and excluding MI mice with EF >40% or FS >20% at day 1 post-MI is recommended. However, most laboratories are using two-dimensional echo (2D echo), which might not be able to accurately detect failed MI (Thielen et al., 2021) and evaluate infarct size. The main limitations of traditional 2D echo are its geometric assumptions, the acoustic window characteristics, probe positioning errors, and a highly subjective approach (Gao et al., 2011; Russo et al., 2019; Scherrer-Crosbie and Thibault, 2008). On the other hand, 4D echo can generate a video covering several systolic and diastolic cycles with the whole 3D view of the heart without subjective area selection. It is like a mini version of MRI but less costly. The parameters generated are volume based not calculation from distance change and reflect the real-time heart pumping capacity. The limitations of applying this relatively new technology include longer data collecting time (10–15 min per mouse), requirement for minimum noise from the front chest skin, time-consuming data analysis (0.5–1 h per mouse vs. 5 min per mouse for 2D analysis), and big data storage space. We are using the 4D echo to evaluate infarct size and EF on day 2 or 3 post-MI to exclude the failed MIs. We also assign infarct size into three score levels (small: 30%–40%, large: 40%–50% or x-large: >50%). Infarct size less than 30% will be excluded, as this level of infarct normally would not affect cardiac function in mice. With the infarct size score, mice will be divided into three cohorts and mice from each cohort will be randomly assigned to control or experiment groups to make sure there is a similar proportion of infarct levels in each group. We hope this kind of grouping would help to grade the MI and answer the question about which MI grade would benefit most from CM regeneration therapy. In most CM regeneration studies, the function data were presented in a combined group, whereas in clinic studies, the information of each patient at different timepoints is dynamically traced to monitor the progression of MI. Some might argue that the lab mice are in a well-controlled similar genetic background, so it is not necessary to monitor each mouse. However, for the MI model the heterogeneity of the coronary anatomy could cause an unavoidable wide range of infarct size, which turns the clean background mice into a group with a heterogenic injury background. Anyhow, it is not difficult to change the format of data presentation. When the EF data are presented in individual MI mice, a clear time-dependent declining of cardiac function can be visualized in mice without any therapy (our unpublished data, Fig. 6B). If the treatment group is also presented in this fashion, the information of when the beneficial effect starts, how long the benefit lasts, and which subgroup is benefited the most can be clearly visualized. More importantly, using the change of cardiac function in each animal could unmask the beneficial effects that might be buried in the noise caused by heterogenic injury levels at the start point. Efficacy readouts The readouts for CM regeneration studies normally include cardiac function measurement, infarct size measurement, proof for CM proliferation, and cardiac repair index. These methodologies have been intensively described in numerous papers. In clinical trials, cardiac MRI is used to evaluate cardiac performance, myocardial perfusion, and scar size (Saraste et al., 2008). Similarly, cardiac function can be monitored in MI mice at multiple time points by volume-based echo or MRI to trace the dynamic progress of cardiac healing post-MI. Applying similar parameters for cardiac function would help translate the readout from preclinic studies. Unfortunately, the resolution of echo in mice to date is not high enough to effectively trace the change of infarct area. Another piece of key readout is the evidence of CM proliferation. In a recent review by Auchampach et al. (2022), the potential problems and limitations of current methodology for CM proliferation measurement were discussed. It is recommended to use multiple strategies to confirm the cell cycle activity or proliferation in adult CM. We prefer to use a combination of at least two cell cycle markers (EdU, Ki67, or pHH3), sarcomere marker, and WGA via Z-stack scanning to identify proliferating CM. We do not recommend using Aurora B kinase staining in cardiac tissue to show midbody, because it is largely dependent on orientation and has a high chance to generate false positive results (Auchampach et al., 2022). For confirmation of CM division, several transgenic reporter mouse lines have been used to study clonal expansion of daughter cells after cell division (Auchampach et al., 2022), such as MADM, FUCCI, and Confetti. Showing the adult CM division is convincing evidence for proliferation, but the obvious limitation is the complicated and time intensive breeding, and efficiency of Cre-mediated recombination. It would not be practical to use these in vivo systems for drug candidate selection, but it would be an important readout for target validation.
Conclusion and Future Perspectives
New therapy that could promote CM regeneration to enhance myocardial repair and preserve cardiac function in IHDs has long been considered to be paradigm-shifting breakthrough in the field. A considerable amount of time and resources have been invested in identifying target genes that drive adult CM proliferation. Meanwhile, there was an increased awareness and recognition of difficulties in data interpretation and comparative examination across published studies in the CM proliferation field, mostly due to the lack of a common standard.
We reviewed the published targets for mammalian adult CM proliferation, and 17 potentially druggable targets that have been validated in an in vivo rodent MI model in a therapeutic setting were selected for efficacy ranking. We found that the proliferation rate and cardiac function data from most targets could only be used for confirmation of the target's involvement, but not sufficient for meaningful ranking. We managed to score the current targets by several criteria, and the Hippo-Yap pathway has met all criteria and might be the most promising target for CM regeneration based on current knowledge.
However, for new drug development centering around a given target, a consistent, robust, and reliable experiment design is critical for compound selection and lead optimization because readout from each round of in vivo study needs to be consolidated and interpreted to lead to a clear meaningful conclusion. We summarized some suggestions and recommendations based on our experience, as well as some published guidelines for in vivo MI models, to help establish the common guidelines for translational studies in the CM regeneration field.
Moreover, drug development-related characterizations of current targets need to be further investigated. For example, whether and how aging or base CVDs, including diabetes, hypertension, and hyperlipidemia, would affect target's capacity to drive CM proliferation remains mostly unknown. The long-term safety and efficacy of current targets are still missing. Further, critical translational information such as patient subpopulation selection, biomarker discovery, as well as the optimal treatment time window and delivery routes for pro-CM-regeneration therapy are waiting to be completed to help clinical trial design.
It will be a long tough way before we reach the point to announce that the field is ready to turn CM regeneration into reality, but with the huge effort the field has investigated we do see potential promising targets that could one day help extend the lifespan and improve the life quality of CVD patients.
Footnotes
Authors' Contribution
X.Z.: writing – original draft (equal); review and editing (equal). D.L.: writing–original draft (equal); writing–review and editing (equal). Y.S.: writing–original draft (equal); review and editing (equal). J.X.: Conceptualization (supporting); Writing–original draft (equal); Writing–review and editing (equal); writing–review and editing (equal). F.-L.X.: Conceptualization (leading); Writing–original draft (equal); Writing–review and editing (equal).
Author Disclosure Statement
The authors declare no competing financial interest.
Funding Information
This work was supported by the National Natural Science Foundation of China (82070311, 82250610229, 82270274), and the Fundamental Research Funds of the Central Universities, Sun Yat-sen University (2022).