
Abstract
Significance:
Extracellular traps (ETs) represent structured frameworks that comprised DNA embellished with histones and granular proteins extruded by immune cells in response to various stimuli. Immune cells contribute to adverse effects of chronic inflammation via ET generation, promoting the release of nuclear chromatin, reactive oxygen species (ROS), and bioactive proteins into the extracellular matrix.
Recent Advances:
The occurrence of ET formation has been documented across diverse immune cell types. The excessive production of ROS during the activation of these cells has the potential to initiate substantial DNA damage, culminating in chromosome decondensation. The inflammatory microenvironment fosters ROS and ET generation, impacting tissue microenvironment remodeling. Recent studies reveal ET involvement in sustaining persistent inflammation, promoting angiogenesis, and initiating thrombotic processes.
Critical Issues:
This review elucidates ET participation in chronic inflammatory disease etiology, detailing ROS-dependent and ROS-independent ET formation mechanisms and their contextual manifestations. It discusses diverse immune cell-derived ETs in the inflammatory milieu and their responses to therapies. Furthermore, the review emphasizes the significance of ETs as potential biomarkers and envisions prophylactic strategies against ET-associated chronic inflammation.
Future Directions:
Subsequent investigations are warranted to uncover the intricate mechanisms governing the resolution of inflammation through ETs in normal physiological processes. Moreover, a comprehensive understanding of the aberrant pathways driving ET formation in persistent inflammation is imperative. Prospective research endeavors should focus on executing expansive clinical studies to discern the involvement of ETs in both the diagnostic and prognostic facets of inflammatory diseases, thereby shedding light on their prospective utility as biomarkers.
Introduction
Chronic inflammatory diseases, including autoimmune disorders, cancer, cardiovascular diseases (CVDs), chronic kidney diseases, diabetes mellitus, neurological diseases, nonalcoholic fatty liver disease, and pulmonary diseases, represent predominant contributors to disability and mortality worldwide (GBD 2017 Causes of Death Collaborators, 2018; GBD 2017 Disease and Injury Incidence and Prevalence Collaborators, 2018). The phenomenon of systemic low-grade persistent inflammation exhibits an upward trajectory in correlation with advancing age. Additional etiological factors responsible for this escalating incidence include chronic and recurrent infections, microbial dysbiosis, environmental factors, sedentary lifestyle, dietary patterns, obesity, psychological stress, and exposure to toxins (Furman et al., 2019).
A diverse array of intracellular signaling pathways are implicated in the context of chronic inflammation.
These pathways, including cell surface receptors (e.g., cytokine and chemokine receptors, epidermal growth factor receptor [EGFR], G-protein-coupled receptors [GPCRs], insulin-like growth factor receptor-1 [IGF-1R], nucleotide-binding oligomerization domain-like receptors [NOD-like receptors/NLRs], retinoic acid-inducible gene I [RIG-I]-like receptors [RLRs], toll-like receptors [TLRs], vascular endothelial growth factor receptors [VEGFRs]), growth factors (e.g., colony stimulating factor [CSF], epidermal growth factor [EGF], fibroblast growth factor [FGF], hepatocyte growth factor [HGF], insulin-like growth factor [IGF], transforming growth factor [TGF], vascular endothelial growth factor [VEGF]), inflammatory cytokines (e.g., interleukins [IL], tumor necrosis factor [TNF]), chemokines (e.g., chemokine [C-X-C motif] ligand 12 [CXCL12], C-C motif chemokine ligand 2 [CCL2]), interferons (IFN), enzymes (cyclooxygenase-2 [COX-2], kinases, lipoxygenase, phospholipase A2), transcription factors (activator protein 1 [AP-1], hypoxia inducible factor-1 [HIF-1], nuclear factor kappa B [NF-κB], nuclear receptors [NRs], mammalian repressor activator protein 1 [RAP1], and signal transducer and activator of transcription 3 [STAT3]), tumor suppressors (e.g., p53, retinoblastoma protein [pRb], and phosphatase and tensin homolog [PTEN]), apoptosis regulatory proteins (B cell lymphoma-2 [Bcl-2], B cell lymphoma-extralarge [Bcl-xL], inhibitor of apoptosis [IAP], and survivin), collectively contribute to the exacerbation of the inflammatory state through the modulation of proinflammatory genes (Alvarez-Garcia et al., 2019; Anderton et al., 2020; Arienti et al., 2019; Barnes, 2006; Bordoloi et al., 2020; Bordoloi et al., 2019; Futosi et al., 2013; Goldman, 2004; Guzman et al., 2020; Hao et al., 1999; Hegde et al., 2020; Jayaprakash et al., 2022; Kanwar et al., 2013; Keshavarz et al., 2021; Khatoon et al., 2022; Khattar et al., 2019; Koelink et al., 2012; Kunnumakkara et al., 2020; Lacroix et al., 2020; Rahman and MacNee, 1998; Riemekasten et al., 2020; Roy et al., 2019; Roy et al., 2017; Singh et al., 2020; Wong et al., 2016; Yuan et al., 2022; Zhao et al., 2021).
In accordance with a growing body of recent research findings, it is becoming increasingly evident that the interactions occurring between immune cells and nonimmune cellular constituents within host tissues exert a pivotal influence on both the onset and evolution of chronic inflammatory processes (Tsai et al., 2021). Research over the last decade has illustrated that tissue-infiltrated immune cells trigger the positive feedback loop of NF-κB through the STAT3 axis by releasing inflammatory mediators such as IL-6 leading to chronic low-grade inflammation (Grivennikov and Karin, 2010; Hirano, 2021). Immune cells also play a critical role in pathogen clearance and host tissue inflammation by orchestrating extracellular traps (ETs) (Daniel et al., 2019; Papayannopoulos and Zychlinsky, 2009; Tsai et al., 2021).
The mechanism of ETosis, cell death that occurred by forming ETs, was first discovered in neutrophils and was suggested to play a vital role in the host defense mechanism against bacterial infection by exposing them to certain bactericidal proteins and peptides (Brinkmann et al., 2004; Daniel et al., 2019). In addition, ETs immobilize pathogens, constraining the spread of bacteria. Under physiological circumstances, ETs monitor ducts and arteries, immobilize and sequester pathogens during host defense, and protect viable tissues from necrosis and the associated massive tissue injury (Daniel et al., 2019). In recent years, the phenomenon of ETosis has been illustrated in several immune cells, including basophils, eosinophils, macrophages, mast cells, and lymphocytes, under various circumstances (Conceicao-Silva et al., 2021; Daniel et al., 2019). ETosis is instigated by a myriad of stimuli, the nature of which varies depending on the specific cell type engaged (Daniel et al., 2019).
In addition, it is noteworthy that ETs participate in both the onset and resolution phases of inflammatory processes (Hahn et al., 2016). At the site of inflammation, ETs have the propensity to group to form larger functional units with the machinery of a variety of enzymes and proteins that can substantially modify biomolecules and their activities (Jimenez-Alcazar et al., 2015; Leppkes et al., 2016). ETs that evade routine clearance from the body after their regular functions have the potential to impair immunological tolerance and act as autoantigens, causing autoimmune malfunctions and exacerbating chronic inflammation (Leffler et al., 2012; Mahajan et al., 2016).
The participation of ETs has been demonstrated in the initiation, resolution, and pathology of a spectrum of noninfectious inflammatory diseases such as asthma, autoimmune disorders, CVDs, thrombotic diseases, metabolic syndromes, neurological manifestations, and neoplasms (Berthelot et al., 2017; Castano et al., 2023; Cristinziano et al., 2022; Farhan et al., 2023; Jimenez-Alcazar et al., 2017; Lee and Cho, 2022; Manda-Handzlik and Demkow, 2019; Manda-Handzlik and Demkow, 2015; Martins-Cardoso et al., 2020; Poto et al., 2022; Wu et al., 2023). Aberrant ETs have been detected in solid tissues, blood, saliva, sputum, and cerebrospinal fluid in these diseases (Garley et al., 2022; Grabcanovic-Musija et al., 2015; Hwang et al., 2019; Joshi et al., 2020; Manda-Handzlik and Demkow, 2019; Michiba et al., 2022; Obermayer et al., 2014; Thakur et al., 2023; Tillack et al., 2013; Yang et al., 2020; Yazdani et al., 2019).
Hence, targeting the defective production of ETs or promoting their efficient systemic and tissue-level clearance are appealing objectives for future therapeutics, but diverse stimuli and actions of ETs pose major challenges to these approaches.
In this contemporary review, we have discussed the underlying mechanisms governing ET formation in the context of chronic inflammation, originating from diverse immune cell types, including basophils, eosinophils, lymphocytes, mast cells, macrophages, and neutrophils. The multifaceted functions of these ETs at the inflammation site have been described. A particular focus was directed toward their deleterious role in the liberation of ROS and proteolytic enzymes. In addition, we have provided a comprehensive elucidation of the potential utility of ETs as diagnostic indicators for a spectrum of chronic ailments. Furthermore, a detailed exposition has been presented on how modulation of ET formation and clearance pathways could offer avenues for therapeutic strategies targeting inflammation, thrombosis, and concomitant tissue damage with due consideration of ongoing clinical trial endeavors.
ETs: What Are They?
ETs are structures that comprised a DNA scaffold entangled with granular proteins that are extruded by immune cells in a process known as ETosis, although cell death is not always necessary for ET formation (Uddin et al., 2019). ETs were first described in neutrophils and were hypothesized to play a role in host defense due to their potential to destroy bacteria by exposing them to bactericidal proteins and peptides (Brinkmann et al., 2004). ETs also immobilize the pathogens within their web-like structures, which subsequently prevent the infection from spreading. In addition, studies have also revealed that ETs can be formed either through nuclear DNA extrusion (suicidal) or via the expulsion of mitochondrial DNA (mtDNA; vital) (Brinkmann and Zychlinsky, 2007; Yipp and Kubes, 2013).
Succeeding studies have shown ETs in systemic lupus erythematosus (SLE), which was considered to be the cause of improper elimination of dead cells and debris (Pisetsky and Jiang, 2006). Extensive research in the field identified the phenomenon in other granulocytes and mononuclear phagocytes, and ETs are later named according to the cell types that generated them, for instance, neutrophil ETs as NETs, eosinophil ETs as EETs, and macrophage ETs as METs (Conceicao-Silva et al., 2021; Daniel et al., 2019). ETs were also found to be instrumental in the modulation of inflammation. Importantly, ETs are heterogenous, and diverse scaffolds of macromolecules and DNA-based framework may alter the characteristics of ETs depending on the surrounding tissue microenvironment and stimulating agents (de Andrea et al., 2021; Hilscher and Shah, 2020; Papayannopoulos, 2018; Petretto et al., 2019).
Nonetheless, the generation of ETs may turn into pathogenesis under specific conditions, resulting in immunological activation and inflammation, followed by organ damage (Czaikoski et al., 2016; Huang et al., 2015; Knight et al., 2012; Yousefi et al., 2012).
Molecular Mechanisms of ET Formation
The release of ETs is a conserved defensive strategy by numerous organisms from various kingdoms (Neumann et al., 2020). As stated earlier, ETosis is a signature of different immune cells against infection and acute-phase inflammation (Conceicao-Silva et al., 2021; Daniel et al., 2019). Nevertheless, the current knowledge about ETs and their underlying mechanism is enormously based on the evidence from neutrophils (Conceicao-Silva et al., 2021; Daniel et al., 2019; Mukherjee et al., 2018). In the course of NETosis, activated neutrophils go through significant morphological changes that cause the nucleus to lose its distinctive multilobular nature (Fuchs et al., 2007). In addition, the inner and outer nuclear membranes gradually separate, chromatin decondenses, and the membranes concurrently rupture. An enzyme peptidylarginine deaminase 4 (PAD4) has been demonstrated to be involved in the citrullination of histones and the subsequent decondensation of chromatin (Anzilotti et al., 2010).
The nuclear envelope then disintegrates entirely into vesicles that merge with the cytoplasm, resulting in a homogenous mass consisting of cytoplasmic, nuclear, and granular contents (Fuchs et al., 2007). Following that, cell contraction occurs, culminating in plasma membrane rupture and the subsequent expulsion of cellular material, resulting in NETs (Fuchs et al., 2007). NETs comprised mainly of decondensed chromatin and proteins, which include cathepsin G, defensins 1–3, histones, high-mobility group box 1 protein (HMGB1), myeloperoxidase (MPO), and neutrophil elastase (NE) (Brinkmann et al., 2004; Pinegin et al., 2015; Zou et al., 2018). However, the complete composition of these ETs is unclear. Urban et al. revealed 24 proteins involved in NETs, including cytoplasmic, granular, and nuclear proteins. Histones represent 70% of all NET-associated proteins, although, among nonhistones, NE was the most prevalent, and catalase (CAT) was the least prominent (Urban et al., 2009).
Of note, there are no basic hallmarks of apoptosis seen in ETosis, such as DNA fragmentation or the initial exhibition and recognition of phosphatidylserine on the cell membrane. NETs are also found to be independent of caspases (Thompson-Souza et al., 2022; Ueki et al., 2013). This process of ET formation was also found to differ from necrosis, in which the nuclear membrane disintegrates in ETosis, while it remains intact during necrosis (Fuchs et al., 2007).
Numerous factors can trigger ETosis, including lipopolysaccharides (LPS), bacteria, fungus, viruses, cytokines, IFN, ROS, and chemicals such as phorbol 12-myristate 13-acetate (PMA) (Aulik et al., 2012; Chen et al., 2022; Jenne et al., 2013; Jin et al., 2016; Schultz et al., 2022; Silva et al., 2021; Souza et al., 2018; Twaddell et al., 2019; Zawrotniak and Rapala-Kozik, 2013). The NET formation is either nicotinamide adenine dinucleotide phosphate (NADPH) oxidase (NOX)-dependent, where after releasing the content the neutrophils will eventually die (suicidal NETosis), or independent, where after releasing the content the neutrophils will be alive (vital NETosis) (Chen et al., 2022; Clark et al., 2007; Fuchs et al., 2007; Pilsczek et al., 2010).
Various signaling mechanisms have been implicated in NET generation so far, including PI3K/Akt/mTOR, TLR/receptor interacting protein kinase (RIPK), extracellular regulated protein kinase 1/2 (ERK1/2), protein kinase C (PKC), c-Jun N-terminal kinase (JNK), and Src (Castano et al., 2023; Desai et al., 2016; Douda et al., 2014; Gray et al., 2013; Hakkim et al., 2011; Keshari et al., 2013; Khan et al., 2017; Lv et al., 2021; Nani et al., 2015; Papayannopoulos et al., 2018). Furthermore, NETs have been demonstrated to interact with various processes in ailments, including those that participate in inflammasomes and autophagy (Chargui and El May, 2014; Huang et al., 2020; Kahlenberg et al., 2013; Remijsen et al., 2011).
Mitochondria also play a key role in ET formation by serving three important functions. They facilitate the production of mitochondrial ROS (mtROS), expel mtDNA out of the cell, and offer support by providing energy for the process via glycolytic ATP synthesis (Amini et al., 2018; Cristinziano et al., 2020; Lood et al., 2016; van Dam et al., 2019). It has also been observed that mtROS might release short mtDNA fragments through pores generated in the mitochondrial outer membrane by voltage-dependent anion channel (VDAC) oligomers (Kim et al., 2019). The putative involvement of ROS, mtROS, and mtDNA has gained significant attention since these ROS play an immunomodulatory function in humans (Caielli et al., 2016). The molecular mechanism of NETosis has been illustrated in Figure 1.
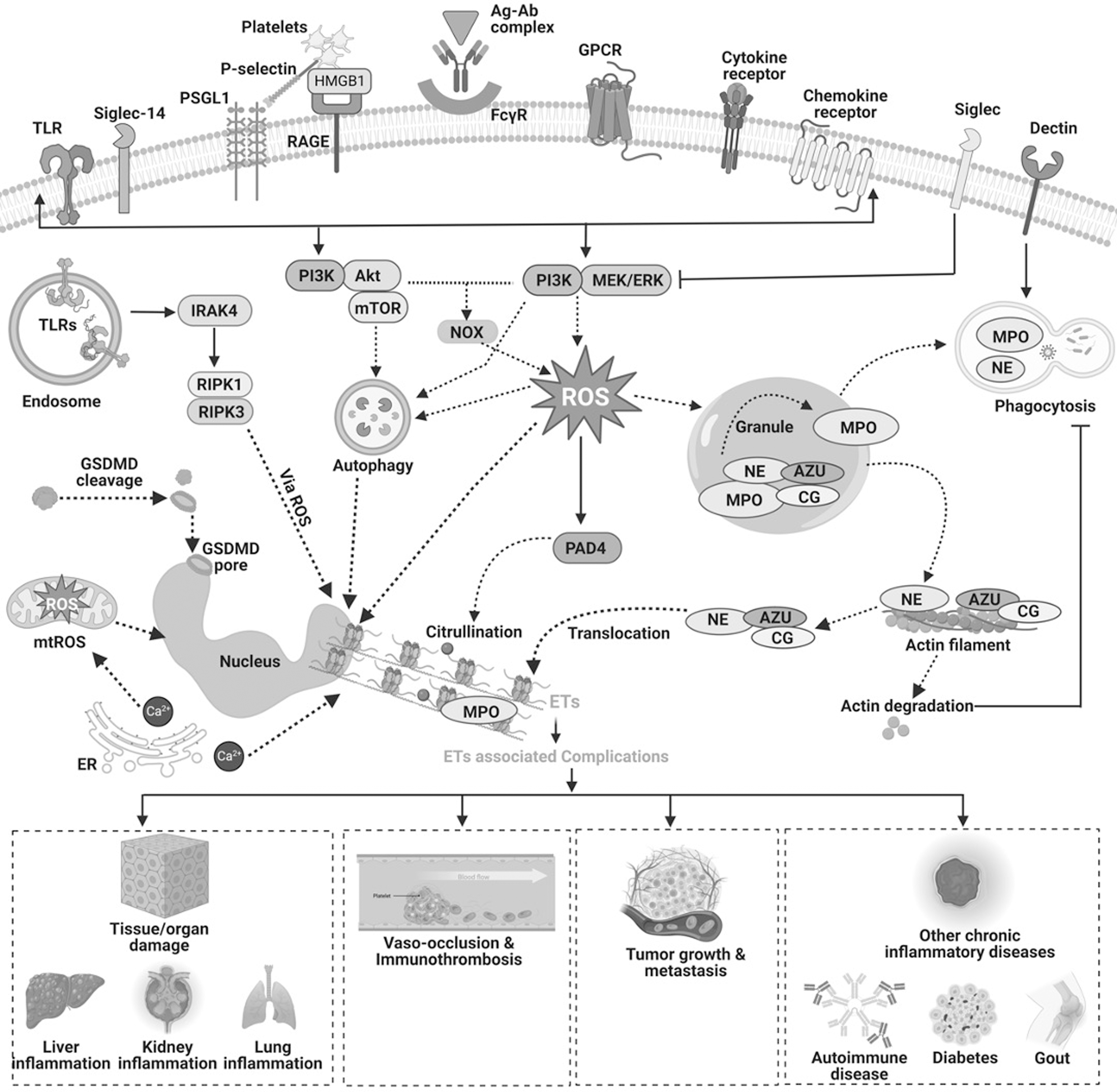
Role of ROS in the Modulation of ETs
Recent evidences suggest that ROS are an integral aspect of most of the reaction cascades that entail the extrusion of ETs (Arazna et al., 2013; Guimaraes-Costa et al., 2012; Kirchner et al., 2012). The coordination between ROS production and NETosis has been well-known (Azzouz and Palaniyar, 2022; Azzouz et al., 2021; Bjornsdottir et al., 2015). Figure 2 illustrates the involvement of ROS from various sources in NET generation. Three enzymes, NOX, MPO, and NE, play a critical role in ROS-mediated NETosis. NOX-mediated ROS stimulate MPO to trigger the activation and subsequent translocation of NE from azurophilic granules to the nucleus (Branzk and Papayannopoulos, 2013; Douda et al., 2015; Papayannopoulos et al., 2010). NE promotes histone processing leading to the disruption of chromatin packaging.
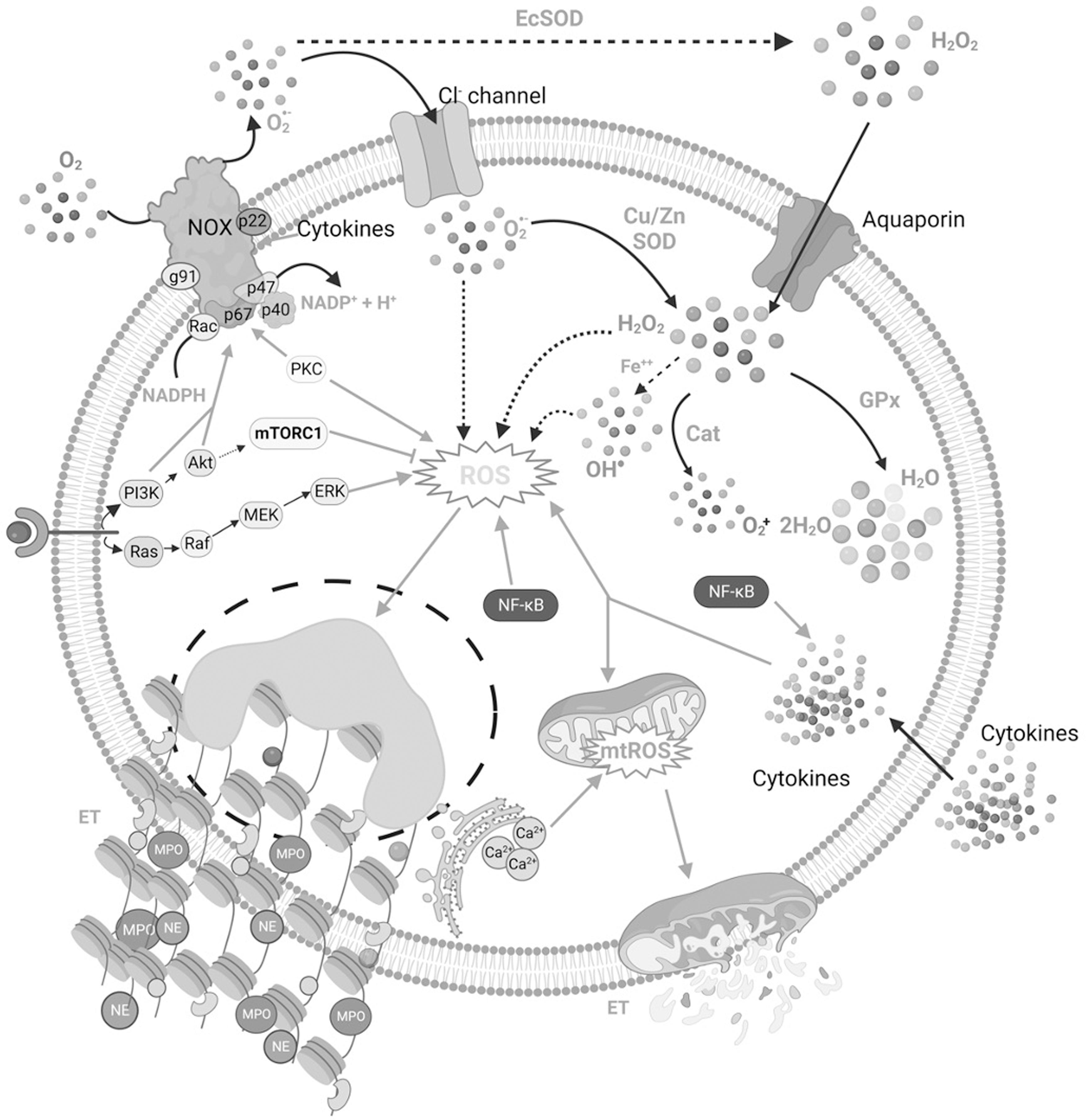
In addition, the binding of MPO to chromatin synergistically with NE decondenses the chromatin succeeding in the extrusion of ETs (Papayannopoulos et al., 2010; Stoiber et al., 2015). Redundancy in NOX activities has been reported in response to specific stimuli, including immune complex formation, in which mitochondrial-originated ROS are sufficient to trigger ETosis (Douda et al., 2015; Naffah de Souza et al., 2017). NOX comprised cytoplasmic subunits-p40phox, p47phox, and p67phox, membrane-anchored subunits-p22phox and gp91phox, and a low-molecular-weight G protein- Rac 2 or Rac1 (Babior, 2004; Bedard and Krause, 2007; Vignais, 2002). Accumulating research demonstrated that enhanced production of superoxide anions by NOX participates in signaling pathways associated with cytokines, hormones, and growth factors (Cho et al., 2017; Ewald, 2018; Fu et al., 2014).
Several studies have documented the role of NOX in the pathogenesis of CVDs, cancers, neurodegenerative disorders, etc. (Ameziane-El-Hassani et al., 2016; Cave et al., 2006; Gao et al., 2012; Lassegue et al., 2012; Sun et al., 2007; Ushio-Fukai and Nakamura, 2008). NOX transfers an electron from cytosolic NADPH to oxygen (O2) in the extracellular matrix, generating superoxide anions (Babior, 2004). These superoxide radicals enter the cells through chloride channels and are dismuted into hydrogen peroxide (H2O2) by extracellular superoxide dismutase (EcSOD) (den Hartog et al., 2014; Fattman et al., 2003; Hawkins et al., 2007). Superoxide radicals can also be converted to H2O2 by cytoplasmic superoxide dismutase (CuZnSOD) and mitochondrial superoxide dismutase (mtSOD) (Bortoluzzi et al., 2021).
H2O2 can either diffuse freely through the membrane or enter via aquaporins and is further metabolized by enzymes, including CAT and glutathione peroxidase (GPx) (Bienert and Chaumont, 2014; Hara-Chikuma et al., 2015; Miller et al., 2010). H2O2 can also generate hydroxyl radicals in the presence of ferrous ions through the Haber–Weiss and Fenton reactions (Kehrer, 2000). Deregulation in ROS production and clearance along with enzymes involved including superoxide dismutase (SOD), CAT, and GPx have been reported in various chronic diseases, including cancers, CVDs, diabetes, liver diseases, kidney diseases, and neurological manifestations (Collin, 2019; Irazabal and Torres, 2020; Jaeschke, 2011; Kehrer, 2000; Schumacker, 2006; Sugamura and Keaney, 2011; Volpe et al., 2018; Waris and Ahsan, 2006). The dynamics involving the manner in which NOX-derived ROS impact NET formation within the cell are seemingly complicated and are yet not entirely conclusive.
Azzouz et al. (2021) stated that NOX-derived ROS functioning at the cellular level can and, in some situations, are required to trigger the generation of NETs. Singlet O2 participation in NOX-dependent NET generation in human neutrophils stimulated with PMA has been validated using O2 scavengers (Nishinaka et al., 2011). The NET formation has been demonstrated to be prevented by experimental inhibition and mutation-caused failure of functioning of NOX. NOX-deficient neutrophils from mutant mice and chronic granulomatous disease (CGD) patients were unable to produce NETs (Bianchi et al., 2011; Fuchs et al., 2007; Rohm et al., 2014). In addition, recent research showed that accumulating intracellular ROS favor autophagy-mediated NETosis (Chargui and El May, 2014; Huang et al., 2020; Kahlenberg et al., 2013; Remijsen et al., 2011). However, it remains unclear how exactly ROS influence the sequential steps of NETosis, that is, neutrophil activation, chromatin decondensation, histone citrullination, granule interaction with the nucleus, binding of MPO and NE to DNA, membrane rupture, and DNA expulsion.
ETs in Chronic Inflammation
It is important to note that recent research suggests that the type and severity of the inflammatory response induced by DNA traps depend on the cell type that produces the traps, the source of the DNA, and whether or not other DNA-related products such as enzymes, histones, and plasma proteins are present. This section summarizes the role of ETs from various immune cells in chronic inflammatory diseases. Table 1 provides insights into in vitro and in vivo studies that demonstrated the role of ETs from various sources in the pathogenesis of chronic diseases. Figure 3 illustrates the general concept of various ETs in health and diseases.

Role of Extracellular Traps in Various Inflammatory Diseases
BET, basophil extracellular traps; CK, cytokeratin; COX-2, cyclooxygenase-2; CLL, chronic lymphocytic leukemia; CLS, crown-like structures; DVT, deep vein thrombus; EET, eosinophil extracellular trap; EMT, epithelial–mesenchymal transition; ET, extracellular trap; G6PD, glucose 6 phosphate dehydrogenase; G-CSF, granulocyte colony stimulating factor; GI, gastrointestinal; IFN, interferon; IICs, immobilized immune complexes; IL, interleukin; LAMP2, lysosomal associated membrane protein 2; LLC, Lewis lung cancer; LPS, lipopolysaccharides; MMP, matrix metalloproteinase; MPO, myeloperoxidase; MSU, monosodium urate; mtDNA, mitochondrial DNA; NADPH, nicotinamide adenine dinucleotide phosphate; NAPA, NSF attachment protein alpha; NETs, neutrophil extracellular traps; NF-κB, nuclear factor kappa B; NRF1, nuclear respiratory factor 1; NSF, N-ethylmaleimide sensitive factor; PAD2, peptidylarginine deaminase 2; PAF, platelet activating factor; PINK1, PTEN induced kinase 1; PS-MPs, polystyrene-microparticles; ROS, reactive oxygen species; SLE, systemic lupus erythematosus; SNAP, synaptosome associated protein; SOD, superoxide dismutase; STX2, syntaxin 2; TFAM, transcription factor A, mitochondrial; TGF, transforming growth factor; TIMP-1, tissue inhibitor of metalloproteinase 1; TLR, toll-like receptor; TNF, tumor necrosis factor; TOM1, target of Myb1 membrane trafficking protein; ULK2, Unc-51 like autophagy activating kinase 2; ZEB1, zinc finger E-box binding homeobox 1.
Basophil Extracellular Traps
Basophils, although a minority among circulating leukocytes constituting merely about 2%, possess a multifaceted role in host defense against parasites, pathogens, and substantial contributions to the pathogenesis of a spectrum of inflammatory ailments, autoimmune disorders, renal fibrosis, and neoplastic conditions (Chauvin and Bayry, 2022; Karasuyama et al., 2018). Basophils participate in T helper (Th) cell-mediated immunity, sense signals from cytokines, TLR agonists, and allergens resulting in the rapid activation of basophils and the release of inflammatory mediators such as IL-4, IL-6, IL-13, leukotrienes, chemokines, and histamine. These inflammatory mediators have been reported to be the potential regulators of basophil-associated pathogenesis (Karasuyama et al., 2018; Morshed et al., 2014; Motomura et al., 2014; Schorn et al., 2012). Besides, basophils expel the DNA known as BETosis within a few minutes of exposure to IgE, cytokines, chemokines, TLRs, and proinflammatory lipids. Basophil extracellular traps (BETs) are orchestrated generally by mtROS-dependent and NOX-independent mechanisms (Morshed et al., 2014; Schorn et al., 2012).
Apart from infectious diseases, the role of BETs has been explained in sterile inflammatory diseases. A salient example that features the BETs in inflammation involves the molecular mechanism of ET formation after the treatment of monosodium urate (MSU) crystals, which mimic the biological inflammation in arthritis in vitro (Schorn et al., 2012). In addition, Morshed et al. (2014) demonstrated that the release of BETs from basophils was largely influenced by IgE and can also be stimulated by other factors, including LPS, platelet activating factor (PAF), lipoteichoic acid (LTA), and f-methionine-leucine-phenylalanine (fMLF). This study also showed that BETs comprised mtDNA and are produced in a mitochondrial-dependent manner. The BETs were also reported to be NOX-independent (Morshed et al., 2014).
Together, these findings suggested that the basophils exert immune effector functions through ET formation as well. However, in-depth examinations are required to acknowledge their participation in the pathogenesis of inflammatory diseases. Indeed, an urgent imperative arises with regard to the establishment of standardized models conducive to the comprehensive investigation of BETs within both in vitro and in vivo contexts.
Eosinophil Extracellular Traps
Eosinophils, a subset of multifunctional granulocytes constituting <5% of circulating leukocytes, with repercussions for allergies, immune response to helminths, viral infections, immunomodulation, homeostasis, and tissue remodeling (Thompson-Souza et al., 2022). Unlike the majority of leukocytes, eosinophils do not perform phagocytosis, instead, they carry out defensive functions by selectively releasing granular content into the surroundings (Thompson-Souza et al., 2022). They feature a bilobed nucleus and cytoplasmic granules containing primary basic granules—galectin 10 and eosinophil lysophospholipase, and secondary cationic granules—major basic protein (MJB), eosinophil cationic protein (ECP), eosinophil-derived neurotoxin (EDN), and eosinophil peroxidase (EPO) (Rosenberg et al., 2013; Thompson-Souza et al., 2022). Typically, EPO interacts with H2O2 during degranulation, releasing catastrophic O2 radicals that kill neoplastic cells, viruses, and Schistosoma mansoni. EDN possesses ribonuclease activity against RNA viruses, whereas ECP possesses antibacterial and antiparasitic properties. MBP-1 is lethal to bacteria and can cause eosinophilic infiltration in host tissues. MBP-1 also has an immunoregulatory function, as evidenced by an elevation in the proinflammatory cytokine IL-8 (Gigon et al., 2021; Yousefi et al., 2012).
A significant advancement in eosinophil biology was introduced by Yousefi et al. who unveiled a novel mechanism termed “EETosis,” through which eosinophils foster host–pathogen interactions and instigate inflammation. They demonstrated the presence of eosinophil extracellular traps (EETs) in colon biopsy samples from patients with Crohn's disease, spirochetosis, and schistosomiasis (Yousefi et al., 2008). These researchers also reported that the DNA content released from eosinophils was of mitochondrial origin along with granular proteins in vitro. This mechanism was found to be NOX- and ROS-dependent. Strikingly, eosinophils persisted in a viable state throughout the EETosis process (Yousefi et al., 2008). Increased entrapment and reduced burden of pathogens due to catapult-like expulsion of mtDNA from eosinophils led to the increased survival rate in in vivo models of Crohn's disease (Yousefi et al., 2008). These revelations marked a transformative paradigm shift in the realm of eosinophil functions.
In another study, eosinophils were shown to secrete IL-8 and expel the ETs in response to MSU indicating its participation in gouty arthritis (Schorn et al., 2012). Proinflammatory cytokine, thymic stromal lymphopoietin, was shown to induce the formation of EETs consisting of mtDNA from the purified blood eosinophils in both time- and dose-dependent manner (Morshed et al., 2012). EETs were also reported in both in vitro and in vivo models of severe asthma in association with increased cytokeratin (CK)18 and CK19 (Lee et al., 2022). Subsequent mechanistic studies revealed that the EET formation from eosinophils derived from asthma patients is ROS-dependent and also found to enhance the secretion of IL-6 and IL-8 (Cao et al., 2023; Choi et al., 2018).
The simultaneous increase in tissue inhibitor of metalloproteinase 1 (TIMP-1) was also noticed in the ROS-dependent activation of eosinophils and generation of EETs (Cao et al., 2023). The formation of EETs was implicated in the increased entrapment of pathogens, including Staphylococcus epidermidis, Staphylococcus aureus, Escherichia coli, Aspergillus, spp., and Candida albicans (Gevaert et al., 2017; Ueki et al., 2016). The catapult of EETs in chronic diseases, including chronic rhinosinusitis and eosinophilic otitis, was found to be NOX- and ROS-dependent (Gevaert et al., 2017; Ueki et al., 2016). The treatment of eosinophils with the NOX inhibitor abrogated the formation of EETs indicating the prerequisite of NOX in the generation of EETs (Ueki et al., 2016).
The aforementioned investigations have delineated that eosinophils extrude mtDNA in reaction to diverse inflammatory provocations. The implication of EETs in a spectrum of maladies, encompassing but not confined to Crohn's disease, gouty arthritis, and asthma, has been ascertained through diverse in vitro and in vivo settings. Consequently, these revelations substantiate the indispensable involvement of EETs in the repercussions stemming from ROS generation and inflammatory processes. Nonetheless, there exists a compelling imperative for more refined models to comprehensively scrutinize the attributes and functions of EETs in pathological contexts.
Lymphocyte Extracellular Traps
T and B lymphocytes are the key orchestrators of immune responses that originate from lymphoid progenitor cells in the bone marrow (BM). The later selection and clonal maturation of these cells take place both in BM and thymus (Conceicao-Silva et al., 2021). These cells are the crucial components of adaptive immunity and induce tailored responses to pathogens and reinforce immunopathological memory (Gray and Gibbs, 2022). In recent years, pathogenic mechanisms of lymphocytes have also been related to DNA release referred to as LETosis, however, the functions and underlying mechanisms of lymphocyte extracellular traps (LETs) are still elusive (Conceicao-Silva et al., 2021; Koh et al., 2020). LETs have been reported to be mtROS-dependent and are associated with a substantial increase in proinflammatory molecules, including TNF-α, IFN-γ, granulocyte-macrophage colony stimulating factor (GM-CSF), and IL-2 in in vitro and in vivo models of autoimmune encephalomyelitis (Costanza et al., 2019).
LETs were also reported from B and T cells derived from healthy individuals treated with serum from SLE patients (Rocha Arrieta et al., 2017). This study suggested that the factors in the serum of SLE patients activated B and T lymphocytes and induced LET formation. Besides, another study reported the formation of LETs by B lymphocytes in response to a cultured medium of chronic lymphocytic leukemia cells, type I IFN, and CpG oligonucleotides of class C (Ingelsson et al., 2018). Research endeavors pertaining to LETs remain notably restricted in scope. Consequently, it is imperative to embark upon further investigations aimed at elucidating the intricate physiological functions of LETs and delineating alterations in their functionality within pathological contexts. Such elucidation is indispensable for the prospective utilization of LETs as viable therapeutic targets.
Macrophage Extracellular Traps
Macrophages are produced from myeloid progenitor cells in the BM as monocytes and differentiate into macrophages in tissues (Han et al., 2021). The heterogeneity of tissue-resident macrophages in human health and diseases, including angiogenesis, neurogenesis, osteogenesis, and tumorigenesis, has been increasingly evident from the decades of research (Park et al., 2022). It was Chow et al. (2010) who reported that murine macrophages, Raw264.7 cells, exposed to S. aureus or PMA expel ETs. Subsequently, preclinical and clinical studies have also confirmed the presence of METs in various inflammatory diseases. One prominent example is inflammation, where METs were observed upon stimulation with TNF-α in RAW264.7 cells (Mohanan et al., 2013). This study also reported that the METs promote the increase in peptidylarginine deaminase 2 (PAD2) in THP-1 cells (Mohanan et al., 2013).
Another study limiting the formation of METs by injecting DNases through the tail vein of mice showed suppression of inflammation and enhanced recovery of motor function in mice models of spinal cord injury (Zhang et al., 2022a). A plethora of research has evidenced that obesity is linked with persistent inflammation and is a well-known causative factor of inflammatory diseases, including breast diseases and breast cancer (Khan et al., 2013; Larson et al., 2023; Naaman et al., 2022). Increased METs have also been noticed in the adipose tissues of mice mammary gland lesions (Mohanan et al., 2013). Mechanistic studies showed that obesity-associated adipose inflammation enhances METs within the lesions of mammary gland (Mohanan et al., 2013). However, studies need to be conducted to validate the role of METs in mammary gland development, remodeling, and pathogenesis.
Recently, another study illustrated enhanced macrophage infiltration and MET formation in microplastic-induced inflammatory models (Wang et al., 2023). This study also elucidated that polystyrene microplastic-induced MET formation leads to hepatic injury and epithelial–mesenchymal transition (EMT) changes in hepatocytes indicating the role of METs in environmental-induced pathologic changes (Wang et al., 2023). Collectively, the findings from these investigations delineated that the METs are present within the inflamed tissues. Nevertheless, it is imperative to undertake comprehensive studies aimed at elucidating the specific contextual factors and the extent of MET formation, as well as their intricate participation in distinct stages of these pathological conditions. Such an in-depth understanding holds the promise of opening novel avenues for the prophylactic management of these diseases.
Mast Cell Extracellular Traps
Mast cells are immune cells that reside within tissues that function as barriers, such as the skin, cardiovascular system, respiratory system, and intestinal mucosa (Cardamone et al., 2016; Varricchi et al., 2020). During microbial infection, mast cells are crucial for the identification and removal of microbial pathogens, onset of adaptive immune responses, and activation of cell–cell interactions (Jimenez et al., 2021; Voehringer, 2013). Notably, mast cells perform immunopathological functions in a variety of inflammatory diseases since they generate chemokines that attract leukocyte subtypes, which are toxic to host tissues (Jimenez et al., 2021; Sobiepanek et al., 2022). The well-known destructive role of mast cells has been described in an autoimmune disease, rheumatoid arthritis, where mast cell-released chemokines, including CCL2, CCL3, CCL4, CCL5, and CXCL12, are found to be associated with tissue deterioration (De Filippo et al., 2013; Sobiepanek et al., 2022; Walker et al., 2012).
Mast cell extracellular traps (MCETs) have also been reported to be effective in limiting Streptococcus infections through NOX-dependent pathways (von Kockritz-Blickwede et al., 2008). Recent evidence has provided a more sophisticated understanding of MCETs in chronic diseases. Lin et al. (2011) elucidated the expulsion of ETs from mast cells in skin explants treated with IL-1β or IL-23. Another study demonstrated the enhanced MCETs in response to hemin and TNF-α in mice models of sickle-cell disease (Tran et al., 2015). Hence, it is evident that ROS-dependent MCETs are the key regulators of immunomodulation; however, further studies are crucial to elucidate the underlying mechanisms, their effects, and consequences in the prophylaxis of inflammatory diseases.
Neutrophil Extracellular Traps
Neutrophils are the most abundant circulating leukocytes in humans and are known for their essential role in host defense against pathogens and are implicated in the various processes of inflammation, including initiation, resolution, and progression (Cristinziano et al., 2022; Erpenbeck and Schon, 2017). The immunomodulatory functions of neutrophils include phagocytosis, secreting ROS, cytokines, and chemokines, degranulation, and entrapment of pathogens through the formation of NETs (Cristinziano et al., 2022; Yang et al., 2016). Among the ETs from various immune cells, NETs have been well-studied and are implicated in various inflammatory diseases both in in vivo and clinical settings (Fig. 4). For instance, Zenaro et al. (2015) demonstrated that neutrophil infiltration, NETs, and IL-17 have been increased in 5XFAD mice models of Alzheimer's disease. NETs have also been implicated in thrombosis, and platelet aggregation in atherosclerotic models (Fuchs et al., 2010; Warnatsch et al., 2015).
NETs were observed to be increased especially in atherosclerotic lesions in ApoE−/− mice (Warnatsch et al., 2015). Moreover, in atherosclerotic baboons, increased amounts of NETs in deep vein thrombus and circulating DNA levels in blood were observed (Fuchs et al., 2010). These studies depicted the key role of NETs in CVDs. NETs were also found to play a crucial role in autoimmune diseases, including psoriasis, rheumatoid arthritis, and SLE (Behnen et al., 2014; Caielli et al., 2016; Cheng et al., 2023; Hollstein et al., 2023; Lande et al., 2011; Lood et al., 2016). The study conducted by Cheng et al. showed the involvement of inflammatory molecules SHIP1, TNF-α, and p38 in NET formation in rheumatoid arthritis (Caielli et al., 2016). In addition, NETs formed in SLE models were found to be mtROS-dependent, which is an unusual mechanism compared with regularly reported NOX-dependent cytosolic ROS (Caielli et al., 2016; Lood et al., 2016). The levels of NETs and oxidized mtDNA were found to be in association with modulated levels of Unc-51-like autophagy activating kinase 2 (ULK2), Ras-related protein Rab27a (Rab27a), Rab4a, N-ethylmaleimide sensitive factor (NSF), intracellular transcription factor A, mitochondrial (TFAM), NSF attachment protein alpha (NAPA), synaptosome associated protein 23 (SNAP23), SNAP29, syntaxin 2 (STX2), lysosomal associated membrane protein 2 (LAMP2), presenilin-1, and target of Myb1 membrane trafficking protein (TOM1) in SLE (Caielli et al., 2016).
Another study showed that NETs substantially influence the secretion of IL-6, IL-8, and glucose 6 phosphate dehydrogenase (G6PD) in airway epithelial cells of severe asthma models (Lachowicz-Scroggins et al., 2019). Defective NETs were detected upon stimulation with high glucose concentration (mimicking diabetic conditions) in the neutrophils in vitro (Joshi et al., 2016). The generation of NETs was also found to be associated with profound metabolic changes in neutrophils (Joshi et al., 2020). These NETs were also reported to be ROS- and NOX-dependent, where treatment with DPI inhibited the formation of NETs (Joshi et al., 2020; Joshi et al., 2016).
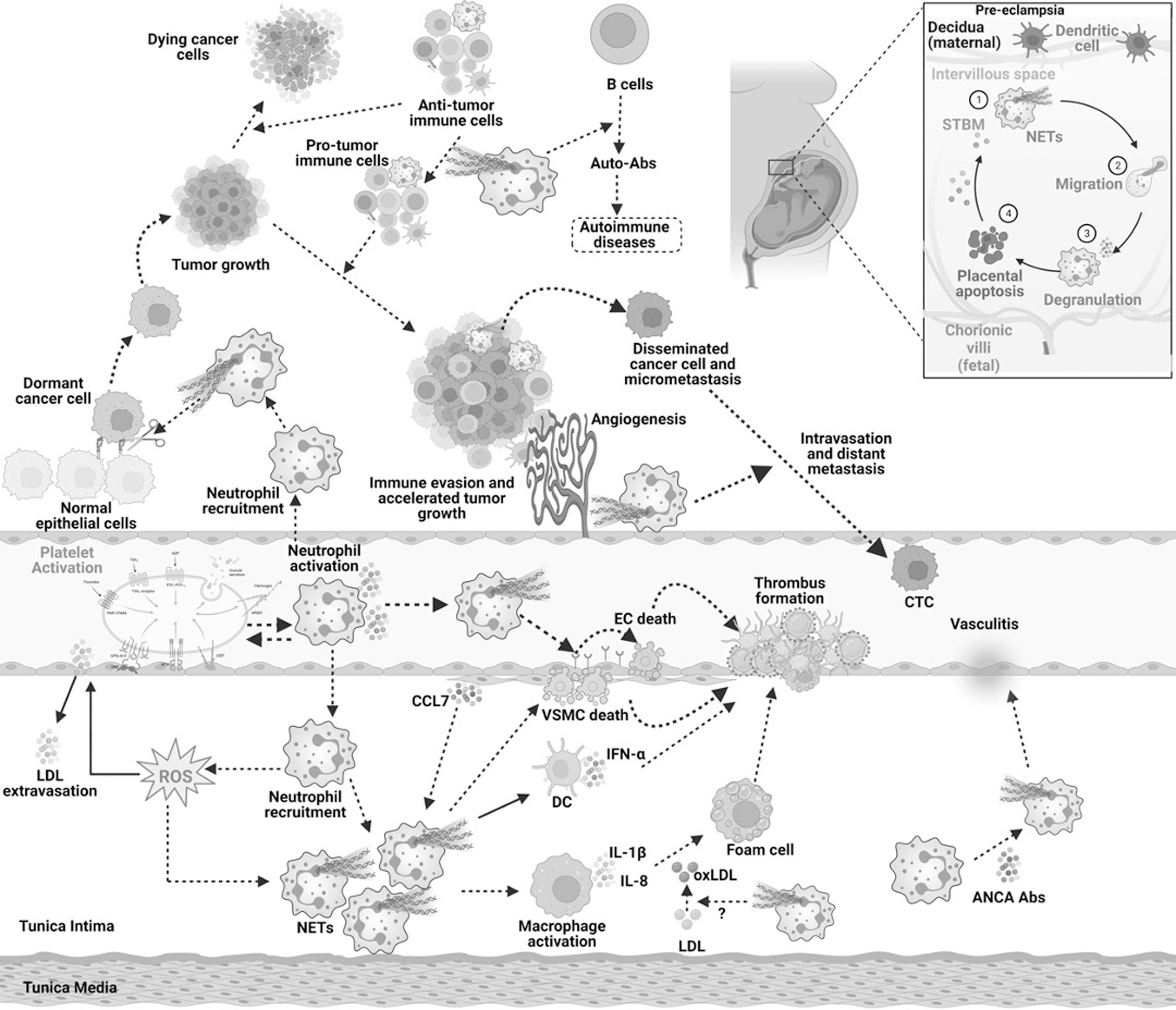
A considerable body of evidence has suggested the association of many forms of cancer, metastatic lesions, and cancer-induced platelet aggregation and thrombosis with enhanced NETs and their components. A persuasive example of NETs in cancer involves the upregulated neutrophil infiltration, NETs in response to LPS, DNA in peripheral blood, tumor growth, and lung thrombosis in 4T1 breast cancer cell-bearing nude mice (Demers et al., 2012). In continuation with this, Gomes et al. (2019) identified the simultaneous increase in NETs and prothrombotic state in nude mice bearing 4T1 cells. In addition, Park et al. (2016) demonstrated that NETs promoted invasion and migration of cancer cells in vitro and lung metastasis in vivo.
The release of NETs in the neutrophil and 4T1 coculture conditions was also found to be dependent on NOX (Park et al., 2016). NETs were also shown to promote liver metastasis in 4T1 cell-bearing nude mice models (Yang et al., 2020). In another study, NET-induced migration and EMT were evident in both luminal breast cancer cells, MCF-7 and triple-negative MDA-MB-231 cells (Martins-Cardoso et al., 2020). The gene expression of those that are positively associated with EMT, including zinc finger E-box binding homeobox 1 (ZEB1), SNAI1, N-cadherin, matrix metalloproteinase (MMP)-2, and MMP-9, was reported to be upregulated, and E-cadherin, an anti-EMT gene, was found to be downregulated in response to NETs (Martins-Cardoso et al., 2020).
This study also demonstrated that NETs stimulated the release of proinflammatory molecules from breast cancer cells, including IL-6, IL-8, IL-1β, and COX-2. The augmentation of breast cancer cell stemness by NETs was also evident by the significant downregulation of CD24 (Martins-Cardoso et al., 2020). Another novel study delineated the enhanced breast cancer liver metastasis association with NETs, in which treatment with DNAse inhibited the metastatic spread to the liver in in vivo models (Yang et al., 2020). This study also demonstrated the crucial role of cell division cycle 25 (CCDC25) in NET formation. It was found that CCDC25 overexpressing MCF-7 cells when injected into mice induced NET formation resulting in increased liver and lung metastasis of these breast cancer cells (Yang et al., 2020). Contrarily, implantation of CCDC25 knockout patient-derived breast cancer cells was demonstrated to inhibit NET formation and the subsequent liver metastasis (Yang et al., 2020).
Similar results were observed in CDCC25−/− mice, where reduced NETs were correlated with resistance to liver metastasis of breast cancer cells (Yang et al., 2020). These results elucidated the novel mechanism in which CCDC25 might play a crucial role in regulating NETs in breast cancer cells and targeting CCDC25 and/or NETs may aid in the effective treatment of breast malignancy (Yang et al., 2020). Interestingly, this study also reported the possible association of NETs and CCDC25 in mice models of colon cancer metastasis to the liver and lung (Yang et al., 2020). Another study reported the enhanced NETs formation and its subsequent role in surgical stress-induced gross and liver metastases in mice with colorectal cancer (Tohme et al., 2016). It was found that surgery-induced TLR9 upregulation positively regulated NET formation and liver micrometastases in C57BL/6J mice bearing colorectal tumors (Tohme et al., 2016).
Yazdani et al. (2019) achieved enhanced mitochondrial biogenesis and reduced metastasis through the inhibition of NETs in animal models of colorectal cancer. Furthermore, in vitro mechanistic studies revealed the association of inflammation and metastatic inducible genes, including PTEN induced kinase 1 (PINK1), Parkin, peroxisome proliferator-activated receptor gamma coactivator-1 alpha (PGC-1α), nuclear respiratory factor 1 (NRF1), and TFAM, with NETs (Yazdani et al., 2019). Furthermore, ROS-independent NETs were found to be increasingly involved in platelet activation and thrombosis formation in pancreatic cancer models (Abdol Razak et al., 2017). Indeed, the release of NETs was also found to be stimulated by anaphylactic thyroid cancer cells in an mtROS-dependent manner and the process upregulated inflammatory mediators, including CXCL8 and IL-8 (Cristinziano et al., 2020).
Studies have also reported the indispensable involvement of NETs in lung cancer growth, invasion, migration, and metastasis (Cools-Lartigue et al., 2013; Najmeh et al., 2017; Zhang et al., 2022b). Recruitment of neutrophils to lung cancer tissues was also found to be associated with upregulated proinflammatory cytokines, including TNF-α, IL-1β, IL-6, and IL-18 (Li et al., 2019; Zhang et al., 2022b). Collectively, these exemplary instances encourage the proposition of diverse functions of ROS-dependent or ROS-independent NET formation in the pathogenesis of neoplastic diseases and associated malignancies. Moreover, it is noteworthy that NETs exhibit notable diversity in their formation, with distinctions observed between those triggered by encounters with bacteria and those generated during the pathological occurrences associated with chronic inflammatory disorders, such as diabetes. These discernible variations underscore the potential efficacy of targeting NETosis as a therapeutic strategy for the treatment of these diseases.
ETs as Biomarkers and Therapeutic Targets: A Clinical Perspective
A plethora of research has shown the ET involvement in various chronic ailments in humans and these results suggest that ETs might be possible biomarkers and novel therapeutic targets (Tables 2 and 3 and Fig. 5). Studies have reported the general occurrence of ETs in thrombectomy aspirates from patients with myocardial infarction and mitochondrial extracellular traps (mtETs) have been detected in synovial fluid and plasma from rheumatoid arthritis patients indicating their role in the development and progression of these diseases (Hajizadeh et al., 2003; Pertiwi et al., 2020). Apart from these, studies have also revealed the presence of specific ETs in various human diseases. For instance, Morshed et al. (2014) reported the presence of BETs in the skin biopsies obtained from patients with inflammatory skin diseases indicating the role of BETs in the inflammation and pathogenesis of the disease.
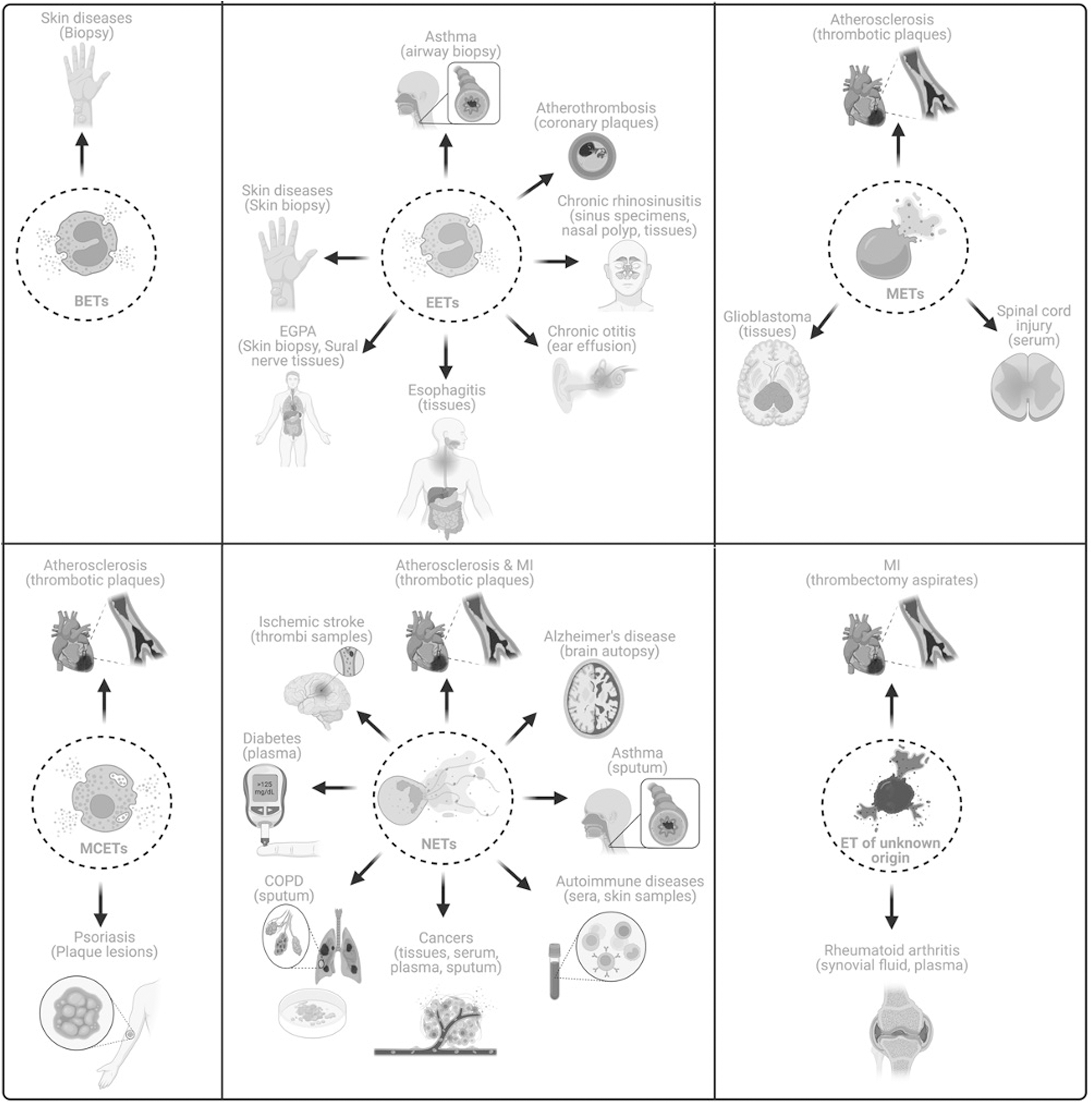
Clinical Applications of Extracellular Traps
ACT, asthma control test; ASIA, American Spinal Injury Association; BAL, bronchoalveolar lavage; cfDNA, cell-free DNA; CitH3, citrullinated histone 3; COPD, chronic obstructive pulmonary disease; DPU, delayed-pressure urticaria; EGPA, eosinophilic granulomatosis with polyangiitis; MCETs, mast cell extracellular traps; METs, macrophage extracellular traps; mtETs, mitochondrial extracellular traps; NE, neutrophil elastase; PAD4 peptidylarginine deaminase 4; SJS, Sjogren's syndrome; SLE, systemic lupus erythematosus.
Summary of Clinical Trials Evaluating the Role of Extracellular Traps in Prophylaxis of Chronic Inflammatory Diseases
The table shows clinical trials regestered on https://beta.clinicaltrials.gov as of October 6, 2023.
In another study, Simon et al. (2011) reported the occurrence of EETs in the skin biopsies derived from patients with eosinophilic skin diseases. Besides, various studies illustrated the occurrence of EETs in respiratory tract diseases, including asthma—bronchoalveolar lavage and airway biopsies and chronic rhinosinusitis—nasal polyps, sinus specimen, and tissues (Dworski et al., 2011; Gevaert et al., 2017; Hwang et al., 2019; Ueki et al., 2016). These studies suggest the possible role of eosinophils as biomarkers and might be novel targets to design therapeutic agents against chronic respiratory diseases. In addition, EETs have been detected in the inflammation of the middle ear (Ohta et al., 2018; Ueki et al., 2016). The presence of EETs were also confirmed in other chronic diseases, including atherothrombosis, bullous delayed-pressure urticaria, eosinophilic granulomatosis with polyangiitis, and eosinophilic esophagitis (Hashimoto et al., 2021; Kawakami et al., 2023; Kerstan et al., 2012; Pertiwi et al., 2019; Simon et al., 2015). Hence, further large-scale clinical studies need to be conducted to establish the possible role of EETs as potential biomarkers in these diseases.
The essential functions of macrophages and mast cells in atherosclerosis and vascular occlusion have been increasingly appreciated. Subsequently, manifestations of both METs and MCETs have also been identified in thrombotic plaques in patients with atherosclerosis (Pertiwi et al., 2019). Another study showed the presence of METs in the tissues derived from glioblastoma patients (Michiba et al., 2022). In addition, Zhang et al. (2022a) and the team revealed that cell-free DNA (cfDNA), CD68, and citrullinated histone 3 (CitH3) in the serum of patients are associated with spinal cord injury. This study also showed that enhanced cfDNA was correlated with exacerbated spinal cord injury and associated inflammation (Zhang et al., 2022a). Moreover, another study detected MCETs in psoriatic plaque lesions (Lin et al., 2011). However, more tailored clinical studies are necessary to conclude MET's and MCET's role as biomarkers in inflammatory diseases.
Over the last two decades, research has shed light on the association of NETs with several inflammatory diseases. Salient examples involve the detection of NETs in the thrombotic samples collected from patients with acute myocardial infarction (AMI) and atherosclerosis (Novotny et al., 2020; Pertiwi et al., 2019). Studies have confirmed the occurrence of NETs in thrombi samples derived from ischemic stroke patients (Laridan et al., 2017; Novotny et al., 2020). An interventional study has been registered and currently recruiting patients suffering from venous thromboses and thromboembolic disease (NCT05711173). The main objective of this study is to identify whether NETs could be the biomarkers for venous thromboses and thromboembolic diseases (NCT05711173). Interestingly, NETs have also been reported in brain autopsy samples from Alzheimer's disease (Zenaro et al., 2015).
Another study aimed at the influence of NETs on immune complexes in multiple sclerosis (MS) patients (Paryzhak et al., 2018). Serum was collected from 23 healthy donors and MS patients (n = 56). These MS patients were divided into five groups as follows: untreated (n = 17), patients undergoing corticosteroid therapy (n = 12), patients undergoing IFN β-1b treatment (n = 15), patients undergoing anti-B cell antibody treatment—ocrelizumab (n = 6) and alemtuzumab (n = 6) (Paryzhak et al., 2018). This study showed that NE activity was increased in MS patients undergoing corticosteroid treatment, which was accompanied by enhanced immune complexes and lectin binding to capture IgG molecules. On the contrary, NE activity and immune complexes were found to be lower in IFN-treated MS patients (Paryzhak et al., 2018). NETs were also shown to be increased in males with relapsing remitting MS (RRMS) compared with females with the same diseases, and prognosis was also reported to be poor in males with RRMS (Tillack et al., 2013). These results indicate the possible prognostic role of NETs in patients with MS. NETs have also been found in both serum and tissues derived from breast and colon cancer patients (Tohme et al., 2016; Yang et al., 2020; Yazdani et al., 2019). In addition, enhanced neutrophil infiltration and NETs have also been reported to be negatively associated with disease-free and overall survival rates of colon cancer patients (Yazdani et al., 2019).
Of note, NETs were detected in the plasma, tissues, and sputum of lung cancer patients (Li et al., 2019; Zhang et al., 2022b). Increased plasma levels of MPO-DNA complexes were related to increased inflammatory cytokines, including TNF-α, IL-1β, IL-6, and IL-18, and a reduced survival rate among lung cancer patients (Zhang et al., 2022b). These studies indicated that NETs might be a possible indicator of lung cancer progression and might also be therapeutic targets. Several registered clinical trials have also been found, which have aimed to establish NETs as novel biomarkers in primary tumors or in cancer-associated thrombosis and also in pediatric malignancies (NCT05056857, NCT03781531, NCT01491230, NCT05040347, NCT04177576, NCT04294589, NCT01533779). These studies indicate that NETs are not only present in tissues but also in liquid biopsies in these cancer patients.
Moreover, NETs were found in the sputum of patients with chronic pulmonary diseases, including chronic obstructive pulmonary disease (COPD) and severe asthma (Grabcanovic-Musija et al., 2015; Lachowicz-Scroggins et al., 2019; Obermayer et al., 2014). In the sputum collected from severe asthma patients NETs co-occurred with increased MPO, mucous secretion, caspase-1, and proinflammatory cytokine IL-1β (Lachowicz-Scroggins et al., 2019). Increased NETs were also found to be associated with an increased intake of oral corticosteroids, and reduced asthma control test scores in these patients (Lachowicz-Scroggins et al., 2019). The possible involvement of NETs in chronic respiratory diseases is also being examined in a large-scale clinical trial (n = 300; NCT05575726).
Another clinical trial conducted in Paris aimed to understand the role of NETs in bronchioalveolar aspirates collected 30 min after the anaphylactic shock to neuromuscular blockers (n = 200; NCT01637220). In an interventional study, the effect of danirixin (selective CXCR2 antagonist) on NETs in COPD was evaluated and reported ∼30% reduction in NETs in the sputum of these patients treated with this drug (n = 19) compared with placebo controls (NCT03250689). However, this study was terminated due to unexpected changes in the benefit–risk profile for danirixin in these patients (NCT03250689). A clinical trial has also been registered to identify NETs in patients with inflammatory lung cystic fibrosis (NCT02564393).
Moreover, Joshi et al. (2020, 2016) described the constitutively increased NETs in the plasma of diabetes patients, and the same group subsequently reported the association of these NETs with altered glucose metabolism and antioxidant pathways. They also demonstrated that isolated neutrophils from diabetic patients form defective NETs, which might be responsible for increased microbial invasion and infections in diabetic patients (Joshi et al., 2016; Joshi et al., 2013). An interventional study has also been conducted to reveal the likelihood of contemporary functions of NETs in diabetic foot ulcers (NCT05577104). Furthermore, NETs were also detected in the sera of patients from autoimmune diseases, including rheumatoid arthritis, Sjogren's syndrome, scleroderma, and SLE (de Bont et al., 2020). Another study detected NETs in psoriatic skin and pustule smears of pustular palmoplantar psoriasis (Hollstein et al., 2023).
Studies have also been registered to understand the feasibility of NETs in preeclampsia and antiphospholipid antibody-induced pregnancy loss (NCT03735108, NCT05470712). Nevertheless, the molecular mechanisms of NETs in preeclampsia remain elusive. Various clinical trials have also been registered to reveal the occurrence of ETs and their components in several chronic diseases, including liver cirrhosis, necrotizing enterocolitis, sickle cell anemia, and systemic sclerosis (NCT05012501, NCT01735552, NCT01106209, NCT02004808, NCT03374618). Another study aimed to determine the role of NETs in ulcerative colitis. Interestingly, NETs were detected only in the colonic biopsies from ulcerative colitis patients (n = 9) but not in biopsies obtained from Crohn's disease (n = 9) and normal controls. This showed that ETs play a crucial role selectively in ulcerative colitis (Dinallo et al., 2019). However, more studies are required to elucidate the role of ETs in differential diagnosis.
Collectively, these clinical observations suggest that irrespective of their origin, ETs are found in various chronic diseases and at different stages of diseases, raising a concern of being a “specific biomarker.” However, studies are also not available to assess them as a suitable biomarker for chronic inflammatory diseases till date. Therefore, it can be envisaged that careful monitoring and pharmacologic interventions might serve to elucidate the possible role of ETs in the diagnosis and prognosis of chronic inflammatory diseases (Fig. 6). Hence, extensive research needs to be conducted to evaluate the potential role of ETs as diagnostic and/or prognostic biomarkers in these diseases in comparison with existing biomarkers such as erythrocyte sedimentation rate and C-reactive protein.
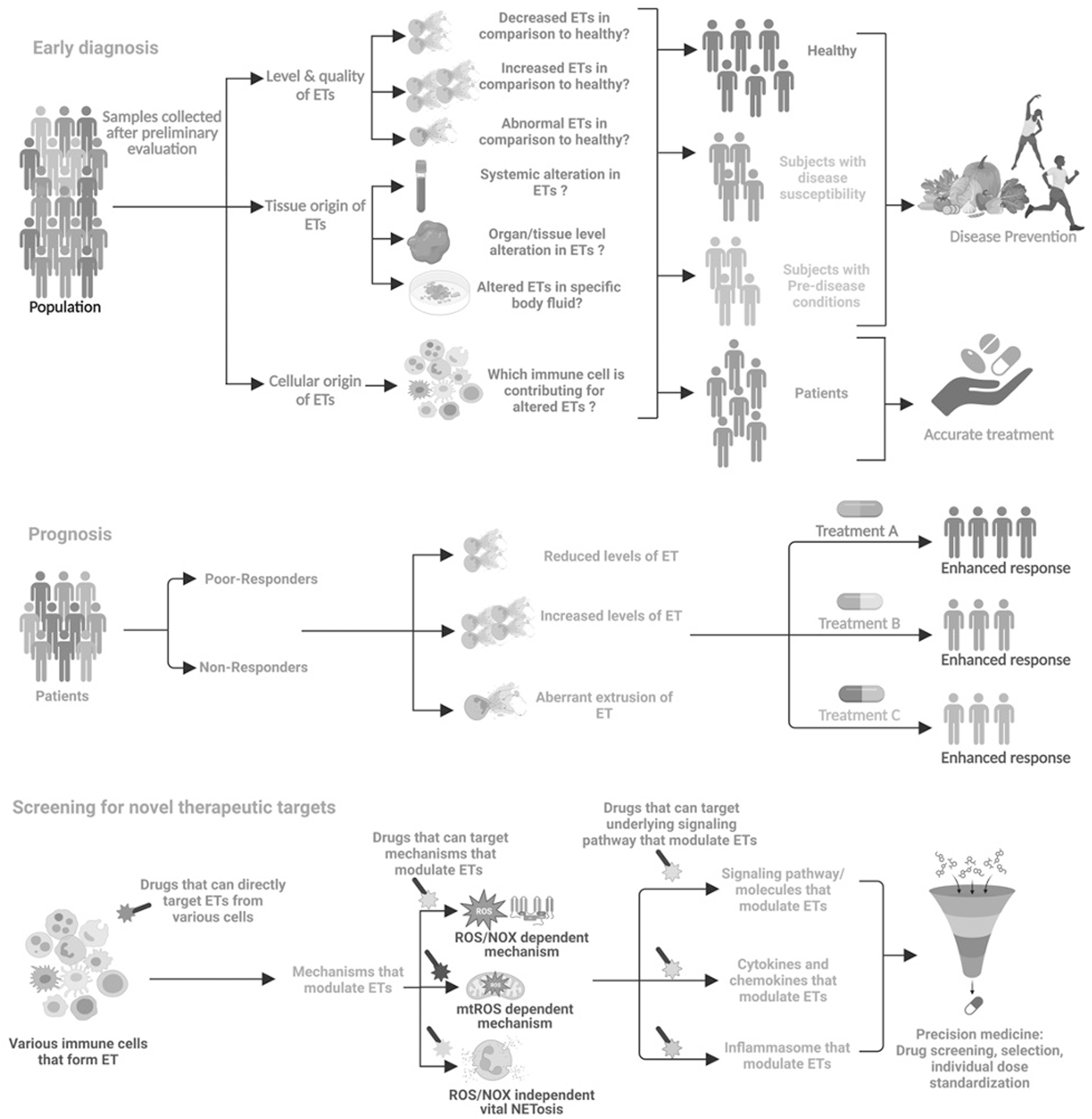
The main features of ETs, including surface receptor-mediated stimulation, gasdermin D (GSDMD) pore formation, NOX-dependent and NOS-independent ROS production, calcium influx, the release of MPO and NE from granules, actin filament degradation, PAD4-mediated histone citrullination, and binding of MPO and NE to DNA, can be utilized for the development of novel therapeutic agents (Fig. 7). However, some of these targets are not specific to ETs alone and targeting these might result in adverse side effects due to their involvement in various signaling pathways. Numerous inhibitors of NETs have exhibited promising effect in the management of diverse medical conditions, encompassing compounds such as Cl-amidine, HDQ, DPI, NAC, recombinant human DNase, vitamin D, and antibiotics.
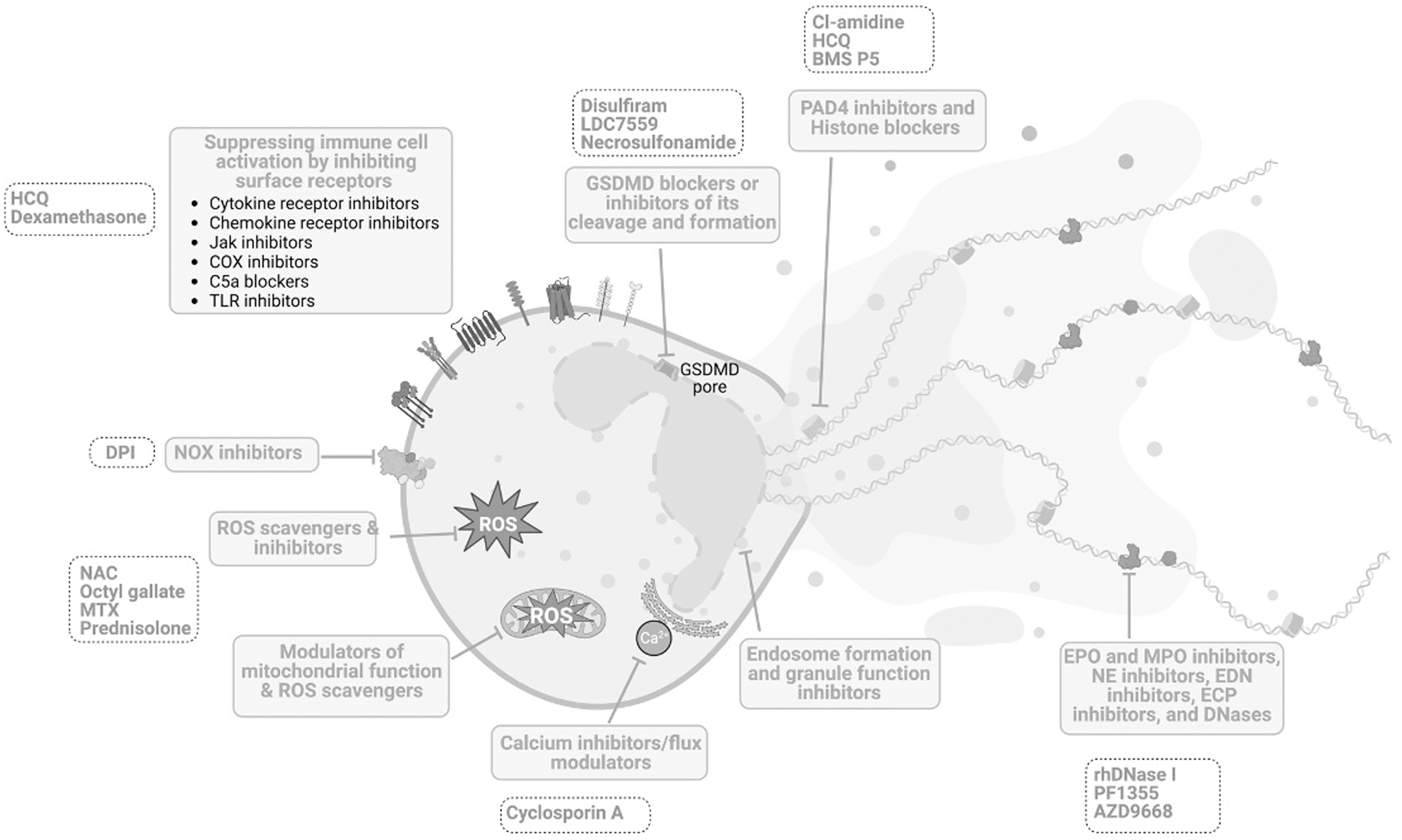
Nevertheless, it is pertinent to acknowledge that these pharmacological agents are associated with adverse impacts on the host's immune milieu, manifesting as enhanced vulnerability to infections and compromised immune function (Bonilha et al., 2023; Chamardani and Amiritavassoli, 2022; Huang et al., 2022). As a potential mitigation strategy, the adoption of combinatorial therapies may prove efficacious in ameliorating these deleterious consequences while concurrently enhancing their therapeutic efficacy. It is also imperative to clinically establish the underlying molecular mechanisms to utilize ETosis and its components as therapeutic targets to develop novel drugs against these inflammatory diseases.
Conclusions and Future Perspectives
Chronic inflammation is the driving force for the initiation and progression of a variety of ailments and provides an excellent route to evade immunosurveillance. Tissue deterioration, modulation of host cell plasticity, neoplastic changes, and response to immunotherapy are substantially influenced by infiltrated and resident immune cells in the tissue microenvironment. Immune cells modulate the tissue homeostasis through various mechanisms. Over the last decade, mounting evidence has suggested that immune cell-derived ETs play a key role in a variety of inflammatory conditions. They are the key regulators in the initiation and perpetuation of chronic inflammation by fostering the inflammatory milieu.
It is now widely acknowledged that these ETs are more diversified and functionally distinctive than previously thought, and currently, understanding specific ETs in health and diseases is an essential part of the research. ETosis has been reported to be a major defense mechanism not only in neutrophils but also in various immune cells, including basophils, eosinophils, lymphocytes, mast cells, and macrophages. The molecular cues that govern the migration of these immune cells to various tissues with distinct topologies and molecular dynamics of their respective ET formation are major open questions in ETosis biology. These characteristics of ETs apart from their friendly microbicidal activities during infection make them ‘a trap in real sense,’ which traps the whole tissue system under a never-ending inflammatory milieu.
Therefore, further research is required to determine whether the mechanisms that control these immune cells' recruitment associated with tissue homeostasis are distinct from those that occur during infection and inflammation. It is yet uncertain whether tissue-specific recruitment patterns and diversified molecular mechanisms of ET formation could aid in the development of treatment strategies. In addition, an imbalance in the on-time clearance of ETs from the host tissue initiates chronic inflammation, making it one of the well-recognized defense mechanisms to a master initiator of inflammation. Hence, future research should concentrate on elucidating the extended function of ETs in physiological tissue homeostasis, disease onset, and secondary perturbations due to various other stimuli in inflammatory diseases. Using an appropriate cell line model is imperative to study ETs, their associated mechanisms, and their role in pathology.
However, the use of leukocyte- or lymphocyte-like cell lines, for instance, RAW264.7 cells, as models for studying METs raises a cautious concern of their nonresemblances to circulating macrophages. Nonetheless, the ease of obtaining and the growing conditions of these cell lines over primary macrophages made them a model to study macrophage-associated mechanisms. Also, in vivo studies are lacking to appreciate the essential functions of ETs derived from these immune cells. Therefore, future studies should be focused to develop suitable models to study these novel mechanisms in physiology and pathology. To fully comprehend their potential use as biomarkers, extensive clinical research encompassing large patient cohorts is imperative to decipher the involvement of ETs in the diagnosis and prognosis of diverse inflammatory diseases.
Footnotes
Acknowledgment
Authors' Contributions
M.H. contributed to the conceptualization, initial drafting of the article, table preparation, visualization, and overall editing; S.G. and T.N.D. contributed to critical article revision; G.S. and A.B.K. contributed to conceptualization, funding, and overall supervision, and supported review development and overall editing.
Author Disclosure Statement
The authors declare that they have no known competing financial interests or personal relationships that could have appeared to influence the work reported in this article.
Funding Information
This project was supported by the BT/556/NE/U-Excel/2016 grant awarded to A.B.K. by the Department of Biotechnology (DBT), Government of India. The authors extend their appreciation to the Deanship of Scientific Research at King Khalid University (KKU) for funding this research through the Research Group Program Under the Grant Number: (R.G.P.2/516/44).