
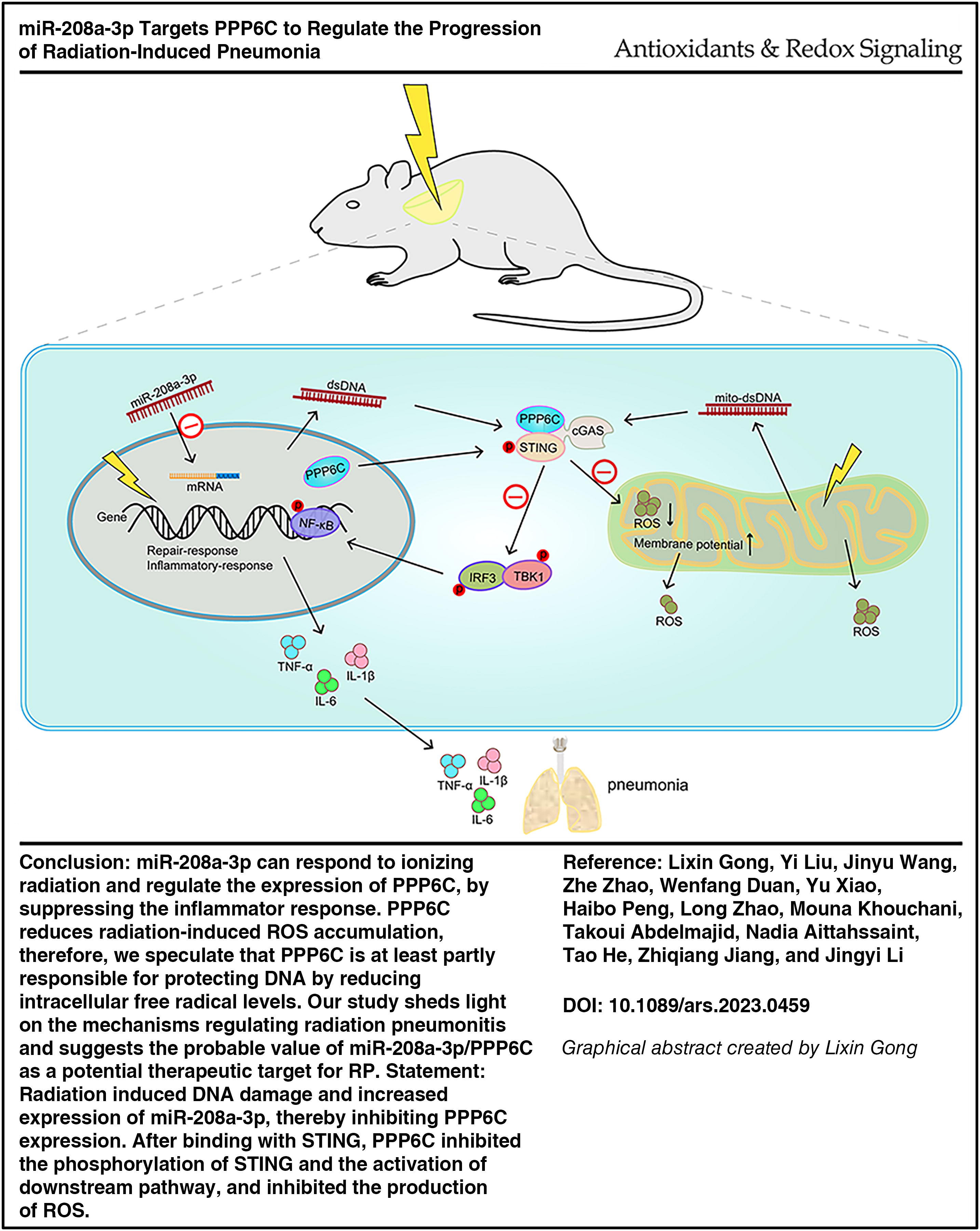
Abstract
Aims:
Radiation-induced pneumonia (RP) is a common complication after radiotherapy for clinical thoracic tumors, and increasing evidence suggests that miRNAs have potential value in regulating radiation-induced lung injury. However, the potential mechanism is still obscure. Here, we evaluated the miRNAs-dependent mechanism involved in the progression of RP.
Results:
Our data showed that mmu-miR-208a-3p was consistently highly expressed in the lung tissue of irradiated mice. In vitro studies demonstrated that the expression of miR-208a-3p in cells was significantly increased after X-ray irradiation. Further mechanism studies indicated that radiation-induced upregulation of miR-208a-3p promoted inflammatory responses by suppressing the expression of protein phosphatase 6C (PPP6C) and activating the cyclic GMP-AMP synthase/stimulator of interferon genes protein pathway. Overexpression of PPP6C can alleviate radiation-induced DNA damage and excessive accumulation of ROS. It was also observed that PPP6C inhibited ionizing RP in vivo.
Innovation and Conclusion:
miR-208a-3p/PPP6C represents a potential therapeutic target for RP which needs to be verified by future clinical studies. Antioxid. Redox Signal. 43, 239–253.
Introduction
The lung is one of the most radiation-sensitive organs (Hanania et al., 2019). Radiation-induced pneumonia (RP) is a general complication of clinical radiotherapy for thoracic tumors (lung cancer, esophageal cancer, breast cancer, etc.) and a dose limiting factor for radiation therapy in patients with thoracic tumors. RP occurs within 6 months of treatment (usually within 12 weeks), cellular injury leads to cytokine release, and cytokine recruitment of the inflammatory infiltrate causes acute pneumonitis (Hanania et al., 2019). Typically, RP is relatively mild for patients with no obvious symptoms other than imaging abnormalities. However, RP may progress into a serious event, and the continued progression of symptomatic RP can cause respiratory failure and even death (Wang et al., 2020).
At present, there is a lack of recognized clinical treatment guidelines for RP, and most of them are handled based on clinical experience. Therefore, an in-depth study of the mechanism of RP progression, the search for early warning markers of RP, and the development of targeted intervention preparations are practical problems that need to be solved urgently in the field of RP treatment. Recent research has shown that miRNAs are tightly connected to the occurrence and development of pulmonary diseases, and the differential expression profiles of miRNAs were observed in response to external stimuli (Chen et al., 2022; Gao et al., 2022; Zhang et al., 2022). Therefore, in various diseases such as malignant tumors, miRNAs are considered disease early warning and diagnostic markers of good value (d’Adda di Fagagna, 2014; Liu et al., 2020; Panir et al., 2018). However, the expression profiles of miRNAs and their roles in the progression of RP are still obscure.
Cyclic GMP-AMP synthase (cGAS)/stimulator of interferon genes protein (STING)/TANK binding kinase 1 (TBK1) is a classical innate immune signaling pathway. When pathogens such as viruses invade cells, cGAS can acquaint the viral DNA in the cytoplasm and induce a large amount of cyclic GMP-AMP (cGAMP). As a second messenger, cGAMP can bind to STING on the endoplasmic reticulum, making it undergo conformational changes, and recruit protein kinase TBK1 and transcription factor IFN regulatory factor 3 (IRF3). STING first stimulates the activation of TBK1 and then phosphorylates IRF3, which promotes the translocation of IRF3 into the nucleus, activates the expression of a large number of IFN-related genes, and promotes the activation of downstream related immune cells and inflammatory factors, thereby exerting antiviral functions (Li et al., 2020; Ma and Damania, 2016; Wan et al., 2020). Recent studies have shown that cGAS can also recognize DNA fragments originating from the nucleus caused by ionizing radiation (Liu et al., 2021).
Innovation
miR-208a-3p can respond to ionizing radiation and regulate the expression of PPP6C, overexpression of PPP6C or inhibition of miR-208a-3p inhibits STING phosphorylation at Ser366 and alleviates radiation pneumonitis by suppressing the inflammatory response. Our study demonstrates that PPP6C reduces radiation-induced ROS accumulation, therefore, we speculate that PPP6C is at least partly responsible for protecting DNA by reducing intracellular free radical levels. Our study sheds light on the mechanisms regulating radiation pneumonitis and suggests the probable value of miR-208a-3p/PPP6C as a potential therapeutic target for RP.
Studies have reported that PPP6C can inhibit STING activity by interacting with and dephosphorylating STING (Ni et al., 2020a; Tan et al., 2017). PPP6C is the catalytic subunit of PP6 and belongs to the Ser/Thr protein phosphatase family (PSP), which is highly conserved in eukaryotes (Ohama, 2019). PP6 displays broad roles in adjusting to a variety of cellular procedures, including cell cycle, autophagy, DNA damage repair, and lymphocyte development. Many studies have reported that PSP family members are closely related to inflammation processes (Geraghty et al., 2013; Wallace et al., 2012; Ziembik et al., 2017).
Herein, we demonstrate that under the condition of ionizing radiation, forced expression of miR-208a-3p decreases PPP6C expression and activates the inflammatory response by activating the cGAS/STING pathway, ultimately leading to the occurrence of RP. Our research will conduce to a deeper comprehension of the outburst of RP and optimize the current treatment strategies.
Results
Pneumonitis and pulmonary fibrosis caused by ionizing radiation
A mouse model of radiation-induced lung injury was established by irradiating the whole chest of C57BL/6 mice with a single 15 Gy X-ray. After irradiation, the mice were sacrificed by cervical dislocation at 14, 30, 90, and 180 days, respectively. The lung tissue structure was observed by hematoxylin and eosin (H&E) staining of paraffin-embedded sections. The results showed that radiation led to inflammatory infiltration of alveoli and damaged alveolar structure, and the damage degree of alveolar structure showed a time-dependence after irradiation (Fig. 1A). Masson staining revealed that after 90 days since irradiation, more obvious collagen fibers and muscle fibers were deposited in the lung tissue (Fig. 1B). The expression of type I collagen in the lung tissue of irradiated mice was significantly higher than that of the control group after 180 days of irradiation (Fig. 1D). As one of the main components of collagen tissue and the “raw material” for later collagen synthesis and deposition of collagen fiber (Srivastava et al., 2016), hydroxyproline (HYP) in mouse lung tissue was significantly increased 30 and 90 days after irradiation (Fig. 1C). To investigate the effect of radiation on inflammation, the content of TNF-α, interleukin (IL)-1β, IL-6, and IL-8 in serum was measured by enzyme-linked immunosorbent assay (ELISA), and it was found that the content of TNF-α, IL-1β, and IL-6 in serum was significantly higher than that of the control group from 14 days after irradiation (Fig. 1G). At the same time, we irradiated Beas-2B cells with different doses of X-rays, and the clone formation experiments proved that the clone formation rate reduced with the increase of the radiation dose (Fig. 1E), and the ROS content in the cells accumulated with the increase of the radiation dose (Fig. 1F). Our results are in good agreement with existing reports. In conclusion, the mice in the early stage of irradiation (about 30 days) began to develop symptoms such as radiation pneumonia, and the mice in the late stage of irradiation (after 90 days) gradually developed symptoms of radiation pulmonary fibrosis.

miR-208a-3p expression is associated with RP
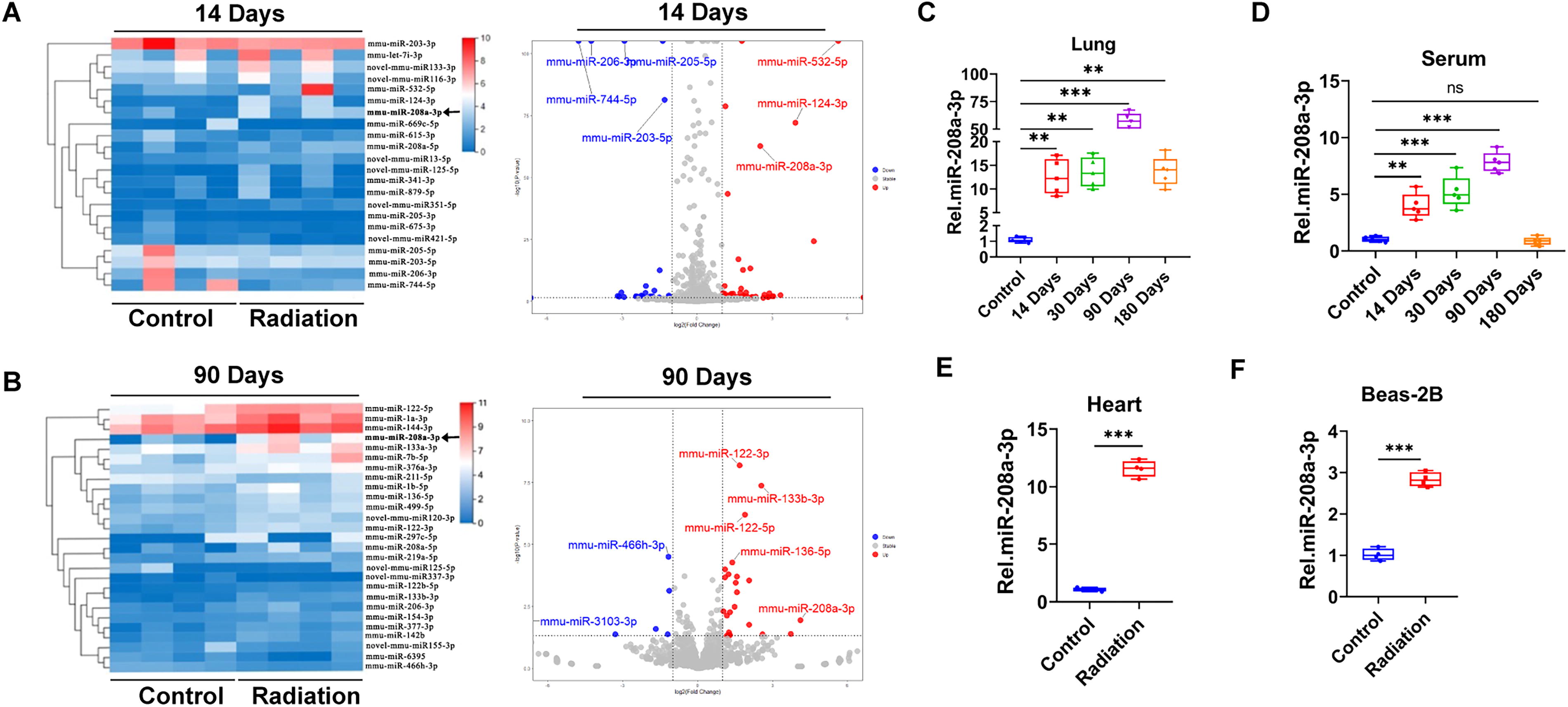
To identify miRNAs that may play a role in radiation-induced lung injury, C57BL/6 mice were irradiated with 15 Gy chest cavity and sacrificed after 14 and 90 days, and the mouse lung tissues were sent to BGI for high-throughput RNA-seq. The results showed that mmu-miR-208a-3p and mmu-miR-124-3p were significantly highly expressed in lung tissues from the irradiation group (after 14 days of irradiation) (Fig. 2A). mmu-miR-208a-3p, mmu-miR-122-5p, mmu-miR-133a-3p in lung tissues were significantly overexpressed after 90 days of irradiation (Fig. 2B). Given that mmu-miR-208a-3p is consistently overexpressed in irradiated mouse lung tissues (after 14 and 90 days of irradiation), we speculated that miR-208a-3p plays a major character in the progression of radiation-induced pneumonia. Meanwhile, we verified the expression of miR-208a-3p by extracting RNA from mouse lung tissue. The results showed that the expression level of miR-208a-3p was higher in irradiated mouse lung tissue. It peaked at 90 days and then gradually decreased (Fig. 2C). We further isolated RNA in serum, and the expression of miR-208a-3p in serum showed the same results (Fig. 2D). Similar results were observed in the heart tissue after X-ray irradiation (Fig. 2E). Furthermore, the expression detection of miR-208a-3p by real-time polymerase chain reaction (PCR) in Beas-2B cells revealed that miR-208a-3p was overexpressed after 10 Gy X-ray irradiation (Fig. 2F). These results suggest that ionizing radiation induces forced expression of miR-208a-3p in the stage of RP.
miR-208a-3p directly targets the PPP6C gene to regulate its expression
In order to study the biological function and mechanism of miR-208a-3p in RP, we first investigated the potential target genes of miR-208a-3p through four databases TargetScan, miRanda, PITA, and PicTar, and 14 potential genes were obtained by the intersection of the four databases (Fig. 3A, Supplementary Table S1). Among them, PPP6C is the catalytic subunit of PP6 and is involved in various cellular activities including inflammation. PPP6C inhibits NF-κB signaling by stabilizing IκBε or by suppressing sustained TAK1 activation. PPP6C helps prevent the continued production of inflammatory factors that would otherwise lead to severe autoimmune disease (Ni et al., 2020b). Gene sequence analysis revealed that there is a miR-208a-3p target site in the 3′-UTR of PPP6C mRNA (Fig. 3B).
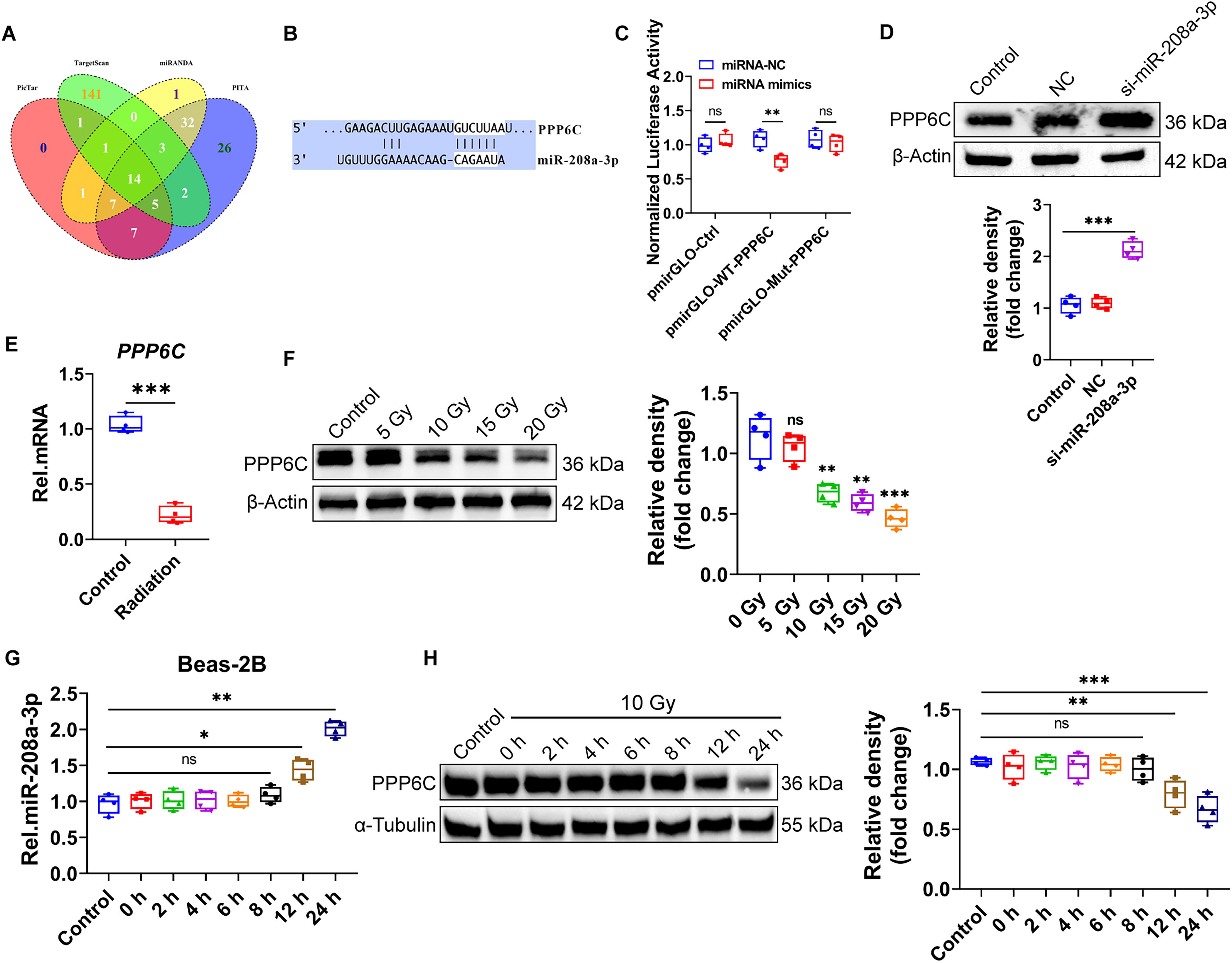
To determine whether miR-208a-3p directly binding to PPP6C 3′ UTR, we designed PPP6C wild type (WT) and PPP6C mutation (Mut) plasmids and utilized a dual-luciferase reporter assay. 293T cells were co-transfected with the miR-208a-3p mimics or miRNA-NC and plasmids. A direct interaction between miR-208a-3p and WT-PPP6C mRNA was confirmed by reduced luciferase activity after 24 h (Fig. 3C). Furthermore, miR-208a-3p knocked down in Beas-2B cells promoted the expression of PPP6C (Fig. 3D). These results indicate that miR-208a-3p directly binds the 3′ UTR of PPP6C mRNA to regulate the expression of PPP6C.
We next investigated whether PPP6C, as a direct downstream target of miR-208a-3p, could respond to ionizing radiation. As expected, the expression of PPP6C mRNA and protein levels were both significantly depressed after X-ray irradiation (Fig. 3E–H).
miR-208a-3p/PPP6C is partially responsible for ionizing RP in vivo
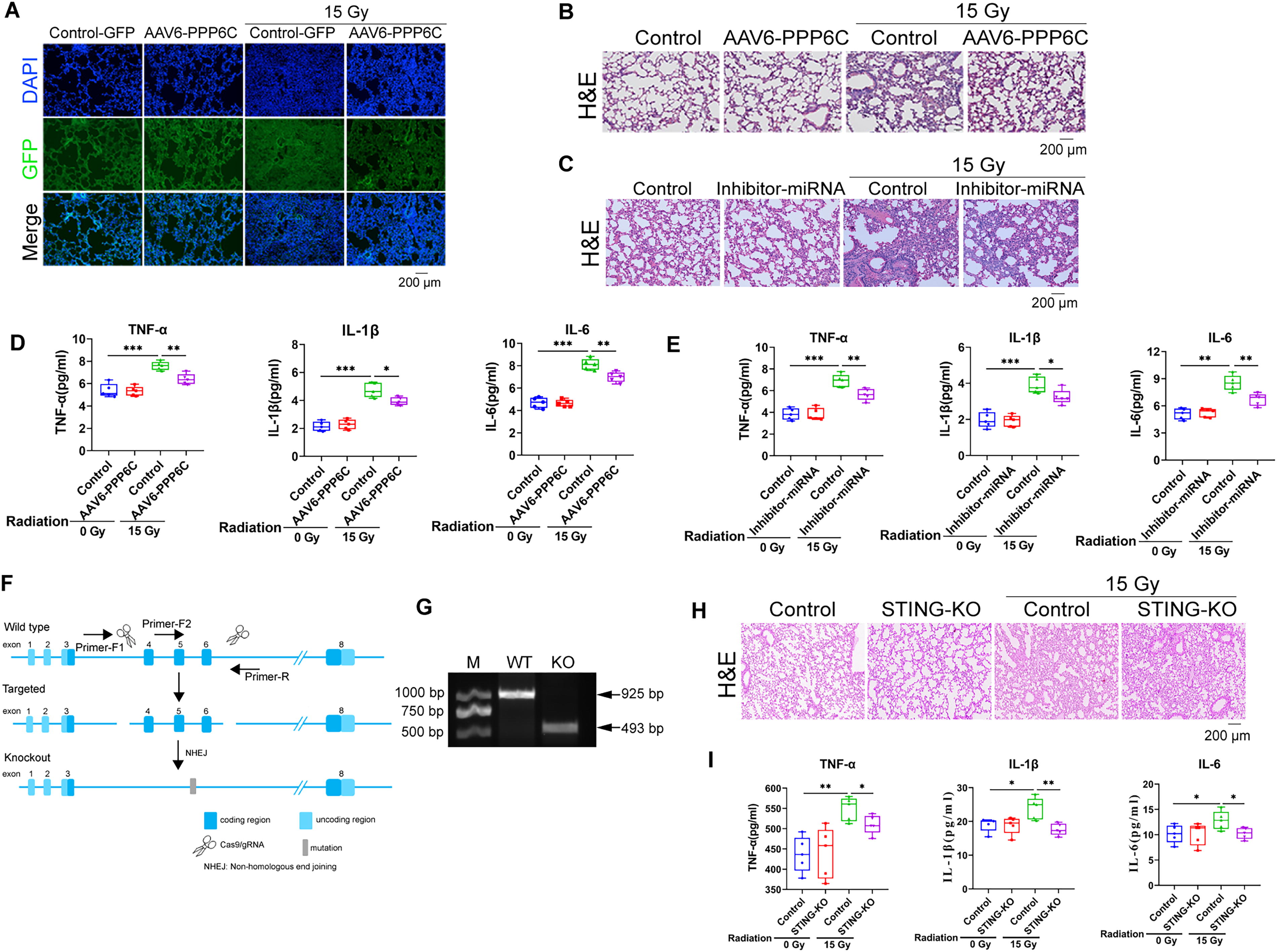
Our in vitro data indicated that miR-208a-3p mediates RP by targeting PPP6C. To further investigate the biological function and mechanism of miR-208a-3p/PPP6C pathway in RP in vivo, mice were exposed to radiation of 15 Gy and after radiation exposure, 100 μL of virus AAV6-PPP6C was inhaled into the lungs of mice by noninvasive tracheal drip or miR-208a-3p inhibitor was injected into the tail vein. The mice were sacrificed 30 days later for further experiments.
As shown in Figure 4A, PPP6C-GFP protein was normally expressed in lung tissues as expected. Compared to the irradiation control group, overexpression of PPP6C or inhibition of miR-208a-3p significantly alleviated RP, mainly reflected in less infiltration of cells observed in the wall and alveolar interstitium (Fig. 4B and C). In addition, ELISA results also showed that overexpression of PPP6C or inhibition of miR-208a-3p can inhibit the increase in the expression levels of radiation-induced inflammatory cytokines such as TNF-α, IL-1β, and IL-6 (Fig. 4D and E). At the same time, STING gene knockout transgenic mouse models under ionizing radiation conditions (Fig. 4F and G) showed alleviated lung tissue structural changes in the radiation-induced lung injury model, accompanied by decreased levels of inflammatory factors such as TNF-α (Fig. 4H and I). These findings align with previous reports indicating that STING activation may promote pathological progression through enhanced inflammation (Frémond and Crow, 2021). Taken together, these data indicate that miR-208a-3p/PPP6C is at least partially responsible for ionizing RP in vivo.
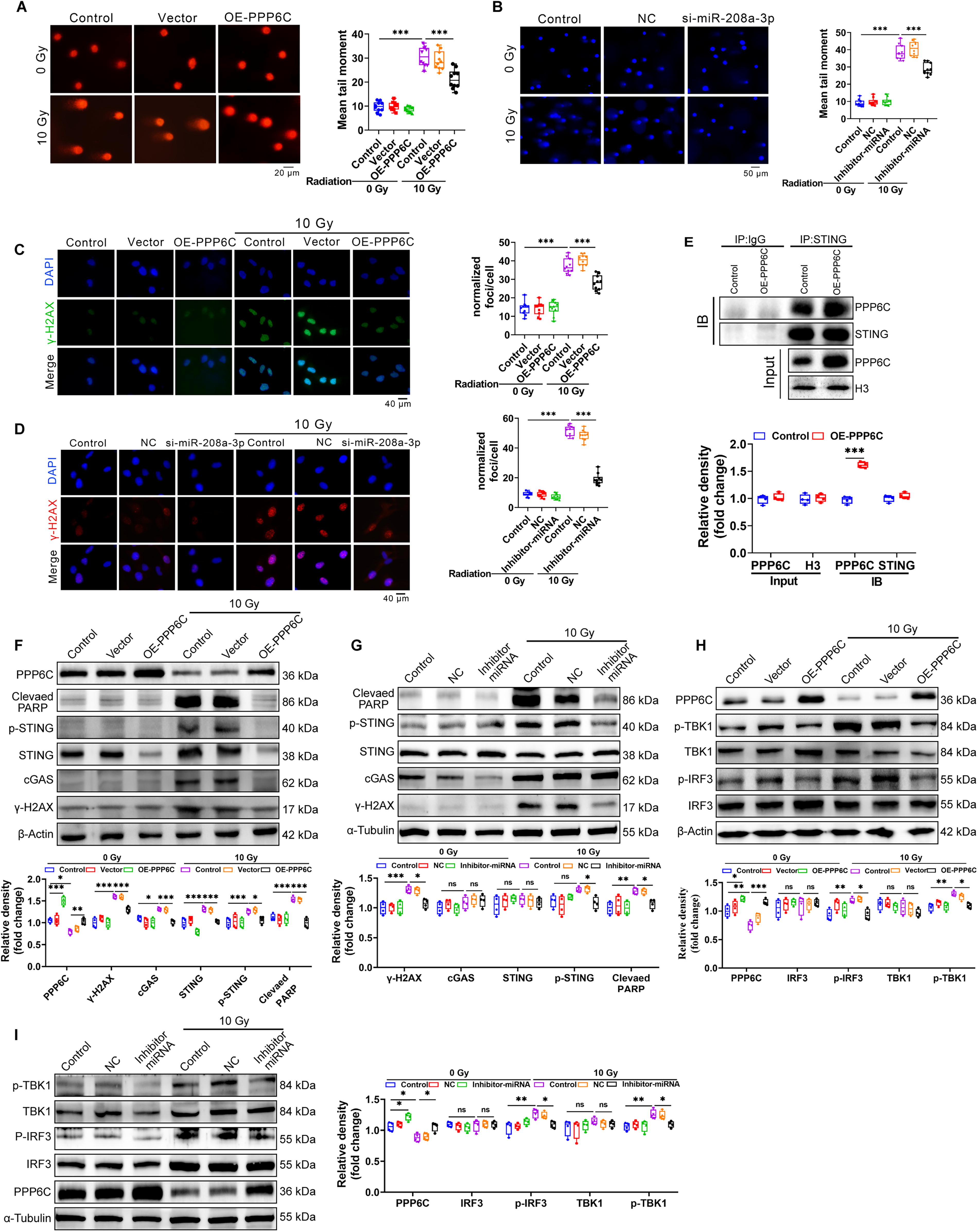
miR-208a-3p/PPP6C regulates DNA damage and inhibits cGAS/STING/TBK1 axis
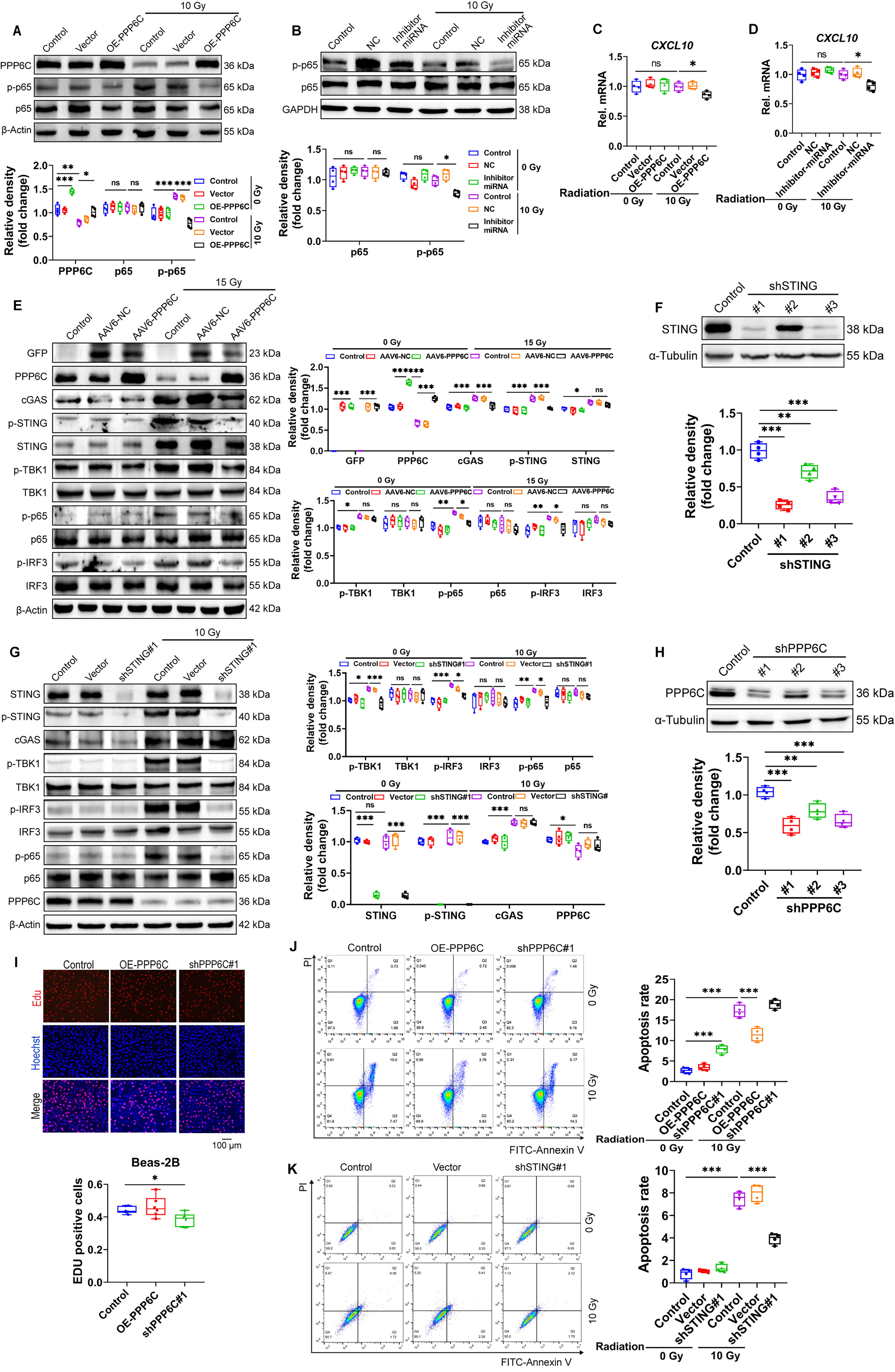
Ionizing radiation can cause DNA fragmentation, and the fragmented DNA fragments enter the cytoplasm through nuclear pores, where they are recognized by cGAS and activate the STING pathway to remove DNA fragments. We found that intervention of the miR-208a-3p/PPP6C pathway can inhibit DNA damage and maintain cell activity under ionizing radiation conditions (Fig. 5A–D). It has been reported that PPP6C negatively regulates STING-dependent innate immune responses (Ni et al., 2020b). To assess the function of PPP6C in regulating the STING pathway during ionizing radiation, we first confirmed the interaction of PPP6C with STING by IP experiments (Fig. 5E). Using the stable cell line expressing PPP6C or miR-208a-3p inhibitor, it was found that radiation-induced accumulation of DNA damage with the detection of DNA damage marker γ-H2AX and PARP as well as activation of cGAS and phosphorylated STING were obviously extenuated (Fig. 5F and G). STING is a key regulator of activation of downstream TBK1 and IFN regulator 3 (Chen et al., 2016; Kwon et al., 2017). Therefore, the effect of PPP6C on ionizing radiation-induced phosphorylation of TBK1 and IRF3 was investigated. Overexpression of PPP6C or miR-208a-3p inhibitor effectively inhibited the phosphorylation of TBK1 and IRF3 in irradiated cells (Fig. 5H and I). We further assessed whether PPP6C also regulates STING-dependent NF-κB signaling activation, with NF-κB subunit p65 Ser536 representing the most effective possible inducible phosphoric acid site in response to inflammatory stimuli (Ghosh and Karin, 2002). The results showed that p65 phosphorylation was significantly inhibited in cells overexpressing PPP6C or miR-208a-3p knockdown after irradiation (Fig. 6A and B). The transcription of some type I IFN genes and proinflammatory cytokines (e.g., CXCL10) is co-regulated by IRF3, NF-κB (Konno et al., 2013; Singhirunnusorn et al., 2005). To further confirm whether miR-208a-3p/PPP6C can regulate the activation of IRF3 and NF-κB, we examined the mRNA expression levels of CXCL10 in PPP6C-overexpressing or miR-208a-3p knockdown cells after irradiation. CXCL10 mRNA expression was obviously decreased in PPP6C-overexpressing or miR-208a-3p knockdown cells after irradiation (Fig. 6C and D). These results propose that miR-208a-3p/PPP6C regulates radiation-induced activation of IRF3 and NF-κB through STING-TBK1. Meanwhile, overexpression of PPP6C inhibited the activation of STING-TBK1-IRF3-p65 (Fig. 6E), and knockdown of STING also suppressed the activation of STING-TBK1-IRF3-p65 (Fig. 6F and G). In addition, we also found that cell proliferation levels decreased and the proportion of apoptosis increased after knocking down PPP6C (Fig. 6H and K).
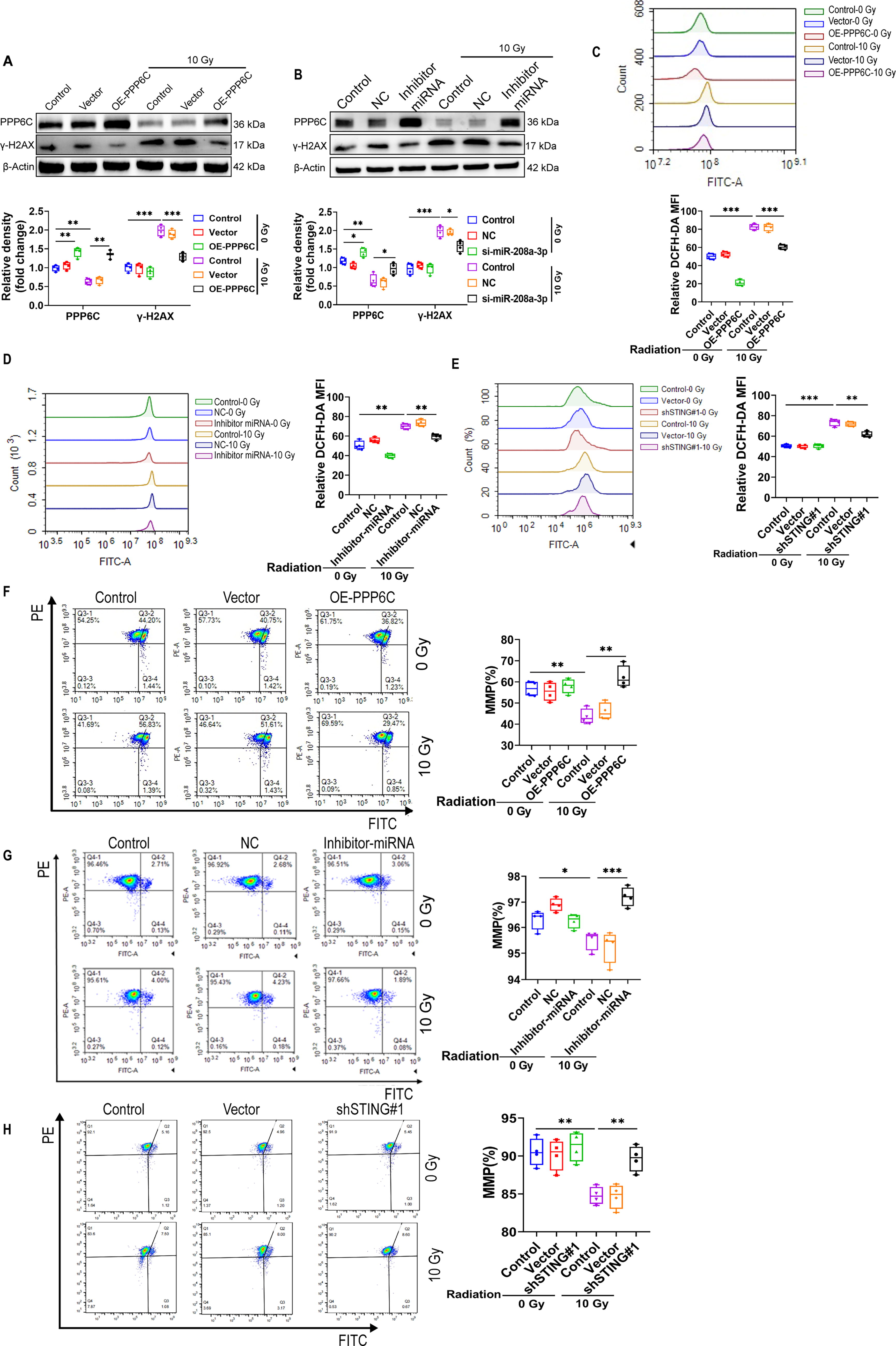
Intervention of miR-208a-3p/PPP6C inhibits ionizing radiation-induced ROS elevation and protects mitochondria from damage
Mitochondria are the energy factories of cells, and studies have reported that ionizing radiation can cause mitochondrial damage, thereby reducing intracellular ROS removal. Our experimental results showed that ROS levels decreased significantly in cells that overexpressed PPP6C or inhibited miR-208a-3p and STING, suggesting that overexpression of PPP6C could reduce ionizing radiation-induced ROS accumulation (Fig. 7A–E). We further evaluated mitochondrial status by measuring mitochondrial membrane potential and found that inhibition of miR-208a-3p and STING or overexpression of PPP6C could effectively protect mitochondria from ionizing radiation damage (Fig. 7F and G).
Discussion
Radiation-induced lung injury can generally be divided into early radiation pneumonitis and late radiation pulmonary fibrosis. Although corticosteroids can be used clinically to treat radiation pneumonitis, they are not suitable for long-term use due to their side effects. In addition, how to accurately diagnose and identify patients who may progress from radiation pneumonitis to fibrosis has been a challenging problem. Therefore, the search for novel radiation-induced lung injury diagnosis and treatment markers has always been a research hot-spot in this field. miRNAs are considered to have good value as a disease early warning and diagnostic markers (Beermann et al., 2016; Liu et al., 2020). Recent studies have shown that miRNAs are closely related to the occurrence and development of lung diseases such as fibrosis. For example, miR-424 activates TGF-β through Smurf2, thereby regulating myofibroblast differentiation during EMT (Xiao et al., 2015). TGF-β can inhibit the expression of let-7d, thereby activating EMT to participate in pulmonary fibrosis (Liang et al., 2016). miR-199a-5p facilitates the progression of pulmonary fibrosis by restraining the expression of CAV1 (Lino Cardenas et al., 2013). Here, we report that radiation promotes immune responses by suppressing PPP6C expression through miR-208a-3p upregulation. Mechanistically, this influence is through the activation of cGAS/STING/TBK1 pathway and persistent inflammation which further induces fibrosis. A growing number of reports have confirmed miRNAs may have clinical relevance as biomarkers (Biswas, 2018; Hu et al., 2018; Scott et al., 2015). In our study, we demonstrate that miR-208a-3p can respond to ionizing radiation and is involved in radiation pneumonitis. Consistent with previous studies, our data have connected the cGAS/STING axis to ionizing radiation-elicited DNA breakage. Ionizing radiation-induced DNA fragments are dispersed into the cytoplasm through nuclear pores, and activation of the cGAS-STING pathway by cytoplasmic DNA rapidly induces hundreds of type I IFN genes and proinflammatory cytokines within hours (Ni et al., 2020a). During host cell response to viral infection, loss of PPP6C promotes STING phosphorylation and activation (Ni et al., 2020a). PPP6C scarcity broadly improves the expression of many other IFN and cytokine genes (e.g., CXCL10), members of the IFN-α family, and several IL genes in response to DNA stimulation(Ni et al., 2020a). It has also been previously reported that PPP6C inhibits NF-κB signaling by stabilizing IκBε or by suppressing persistent TAK1 activation (Kajino et al., 2006; Stefansson et al., 2008). Consistently, our study shows that overexpression of PPP6C or inhibition of miR-208a-3p inhibits STING phosphorylation at Ser366 and alleviates radiation pneumonitis by suppressing the inflammatory response. It is possible that the cytoplasmic DNA fragmentations induced by ionizing radiation can produce downstream biological effects similar to viral nucleic acid invasion.
Using cells overexpressing PPP6C or miR-208a-3p knockdown, we found that radiation-induced accumulation of DNA damage was obviously extenuated. Considering that ionizing radiation can induce DNA damage through direct and indirect effects (via free radicals formed) (Huang and Zhou, 2020; van de Kamp et al., 2021; Wallace, 2022), our study demonstrates that PPP6C reduces radiation-induced ROS accumulation, therefore, we speculate that PPP6C is at least partly responsible for protecting DNA by reducing intracellular free radical levels. However, we do not rule out that PPP6C can protect DNA from radiation-induced direct damage or be involved in DNA repair through some unknown mechanism.
In summary, our data propose that miR-208a-3p regulates the cGAS-STING pathway by inhibiting the expression of PPP6C. Thus, PPP6C gives assistance to avoid the continued production of STING-dependent cytokines that will otherwise arouse severe immune reactions. Our study sheds light on the mechanisms regulating radiation pneumonitis and suggests the probable value of miR-208a-3p/PPP6C as a potential therapeutic target for RP.
Materials and Methods
Cell lines and animal studies
HEK293T cells and Beas-2B cells were purchased from the American Type Culture Collection. HEK293T cells and Beas-2B cells were maintained in Dulbecco’s minimal essential medium (VivaCell, China, C3113-500) with 10% fetal bovine serum (VivaCell, China, C04001-500) and 1% penicillin-streptomycin (VivaCell, China, C3421-0100). All cells were held in a 37°C laboratory brooder in 5% CO2.
This study was carried out according to the principles established by the Declaration of Helsinki, and all animal experiments were approved by the Medical Ethics Committee of the Second Affiliated Hospital of Chengdu Medical College Nuclear Industry 416 Hospital (2020-13). In total, 6- to 8-week-old C57BL/6 mice were divided into control group and experimental group. Mice were held under circumstances (24°C ± 2°C, 40%–70% relative humidity, 12 h light/12 h dark cycle) and were given a regular diet and water.
Generation of STING−/− mice
STING knockout C57BL/6 mice were purchased from Shanghai Model Organisms. Genomic DNA was extracted using the mouse tail genotype rapid identification kit (Beyotime, China, D7283M). The primers for PCR analysis: F1 5′-ACCTGATGGGAGGTATCTACCGG-3′, F2 5′-CCTCCCACCTTTACCTGTGTTACG-3′, and R 5′-CCAGCAACTAGCATCAGAA-CCTCC-3′.
X-ray exposure
Mice were irradiated using an X-RAD 225 irradiator (X-ray: 225 kV and 12.5 mA; SSD: 30 cm; beam filter: 2.0 mm AI). The mice were settled in the center of the radiance field at a dose rate of 200 cGy/min at a distance of 30 cm from the irradiation source and the mice were immobilized in a specially designed fixture designed to irradiate the entire thoracic cavity of the mouse, A total dose of 15 Gy while avoiding exposure to other parts of the body.
Second-generation high-throughput sequencing
Mice were sacrificed by cervical dislocation, 100 mg of fresh lung tissue was quickly put into liquid nitrogen for preservation, and BGI was commissioned to perform high-throughput sequencing analysis. Altogether 16 samples were sequenced on the BGISEQ-500 platform, with an average yield of 28.8 M reads per sample. The average alignment ratio of the sample was 95.23%. A total of 3364 miRNAs were detected. Using negative binomial regression models in DESeq2. |log2FoldChange|≥ 1 and adj. p Value ≤ 0.05, these two statistical thresholds were considered significant.
Immunohistochemistry
The tissues were fixed in 4% buffered formalin and embedded in paraffin. Overall, 3 μm thick sections were stained with H&E (Servicebio, China, G1004), and elastica-Masson trichrome stain (Servicebio, China, G1006). Antigen retrieval was performed by microwave heating in citrate buffer, pH6, for 20 min. After peroxidase blocking, sections were incubated for 1 h with a COL1 (1:50; Santa, USA, sc-59772). DAB Chromogenic Kit (ZSGB-Bio, China, ZLI-9018) was used for signal amplification. Use “imageJ” for quantitative analysis.
Immunofluorescence
Cells were fixed with 4% paraformaldehyde, 0.25% Triton X-100 permeable, 2% BSA, incubated overnight at 4°C with primary antibody (γ-H2AX, 1:200; ZEOBIO, China, R381558), incubated at room temperature with secondary antibody (Fluorescein secondary antibody, 1:1000; ThermoFisher, USA, A-11001/A-11005), and stained by 4′−6-diamidino-2-phenylindole (DAPI). Use “ZEISS” software for quantitative analysis.
Quantitative PCR quantification
RNA was extracted using the EasyPure miRNA Kit (TransGen, China, ER601-01-V2) to conform to the manufacturer’s protocol. Gain the purity of the RNA by checking the optical density 260/280 ratio using a Nanodrop spectrophotometer. For miRNA, cDNA synthesis was performed using the HiScript III 1st Strand cDNA Synthesis Kit (+gDNAwiper) (Vazyme, China, R312-01) with reverse transcription primer 5′-GCGAGCACAGAATTAATA-CGACTCACTATAGG(T)12VN-3′. For mRNA, 1.0 μg of total RNA was reverse transcribed into cDNA in a 20 μL reaction system using the HiScript III 1st Strand cDNA Synthesis Kit (+gDNAwiper) (Vazyme, China, R312-01) in the light of the manufacturer’s protocol. Real-time PCR of miRNA was applied on BioRad CFX96 sequence detection system in 2 × T5 Fast qPCR Mix (Probe) (TSINGKE, China, TSE301), and real-time PCR of mRNA was performed on BioRad CFX96 sequence detection system in SYBR Green I Real-Time PCR Kit (Roche, Germany, 04913850001). The primers for quantitative qRT-PCR analysis: miR-208a-3p 5′-CGGATAAGACGAGCAAAAAGCTTG-3′, Universal reverse 5′-GCGAGCA-CAGAATTAATACGAC-3′, TaqMan probe 5′-FAM-CACCTCGATGCAGCC-ATCTGC-MGB-3′, U6 5′-CTCGCTTCGGCAGCACA-3′, PPP6C-F 5′-CGTT-TTTCTAAAGGAGCGGCTA-3′, PPP6C-R 5′-GGAACCTGACCTCCAGTTC-TGA-3′, GAPDH-F 5′-GTCTTCACCATGGAGAAGGC-3′, GAPDH-R 5′-TTG-TTG-TCATGGATGACCTTGGCC-3′.
The qRT-PCR assays were performed in triplicate and the change in expression level was calculated using the 2−ΔΔCt method.
Plasmid construction
The pCDH-CMV-MCS-EF1-Puro vector and pLVX-shRNA1 vector were purchased from Addgene, and the pmirGLO plasmid was purchased from Promega (USA). The CDS sequence of the PPP6C gene was cloned into pCDH-CMV-MCS-EF1-Puro using a seamless molecular cloning protocol to generate pCDH-CMV-PPP6C. The WT-PPP6C-3′UTR sequence (5′-CTGTCTAGAACTGTTGGAAGA
Lentiviral infection and stable cell line generation
The pCDH-CMV-MCS-EF1-Puro plasmid and the pCDH-CMV-PPP6C plasmid use the helper plasmids psPAX2 and PMD2.G to generate lentiviral particles in HEK293T cells according to the lentiviral expression system. For lentiviral transduction, cells were grown to 70% and inoculated with virus in the presence of 6 μg/mL polybrene. The virus-containing medium was removed after 24 h and cells were grown in fresh medium for 24 h. Forty-eight hours after lentiviral transduction, puromycin medium with a final concentration of 0.25 μg/mL was added for selection. We needed to change the medium every 3 days, and when the cells were no longer dying in the medium containing puromycin, stable cell lines were maintained in a medium containing the appropriate concentration of puromycin to maintain protein expression.
Adeno-associated virus production and infection
Construction of PPP6C overexpressing adeno-associated virus-containing SP-C lung-specific promoter and GFP. AAV6 was produced by transfection of 293T cells, virus enrichment was performed. Mice were exposed to radiation of 15 Gy (for details, see experimental method “X-ray exposure” section). After radiation exposure, 100 μL of the virus was inhaled into the lungs, about 5 × 107 VG of the virus. The mice were sacrificed 30 days later for further experiments.
miRNA mimic transfection and RNA interference
The miR-208a-3p mimics, mimics negative control (mimics NC), miR-208a-3p inhibitor, and inhibitor NC were bought from Ribobio (Guangzhou, China). Mimics were transfected into HEK293T and Beas-2B cells with FuGENE6 (Promega, USA, E2692). Inhibitors were injected 200 nM/kg/3 days via mouse tail vein.
Western blotting and immunoprecipitation
Cell lysates boiled in buffer were separated by SDS-polyacrylamide gel electrophoresis (PAGE) and transferred to polyvinylidene fluoride membranes (Millipore, USA, 03010040001) and immunoblotted with primary antibodies. For immunoprecipitation, lysates were incubated with antibodies overnight at 4°C, followed by the addition of 20 μL of protein A/G beads (Byeotime, China, P2108) for 2 h. Beads were washed four times with buffer and boiled in the appropriate amount of sample buffer before analysis by SDS-PAGE.
Enzyme-linked immunosorbent assay
Mouse serum was collected using an ELISA kit (RUIXIN Biotech, China) and determined according to the manufacturer’s protocol (TNF-α, RX202412M; IL-6, RX203049M; IL-1β, RX203063M).
HYP content assay
Tissue samples were measured using the HYP Content Assay Kit (Solarbio, China, BC0255) according to the manufacturer’s protocol.
Determination of mitochondrial membrane potential and ROS
The 1 × 105 cells were cultured in 6-well plates, cultured for 24 h, subjected to X-ray irradiation, and continued to be cultured for 24 h. The serum-free medium was prepared with a DCFH probe working solution, and the cells were incubated at 37°C without light for 30 min. The ROS levels were collected and measured by flow cytometry. After X-ray irradiation, 24 h cultured cells were collected in 1.5 EP tubes, and the cells were suspended with the prepared JC-10 working solution, incubated at 37°C without light for 15 min, cleaned, and the mitochondrial membrane potential was detected by flow cytometry.
Comet assay
The cells were collected, wrapped in agarose gel with a low melting point, placed on a slide, lyzed at 4°C for 2 h, and neutralized for 30 min after cleaning, the slide with gel attached was placed in a low-temperature unwinding solution for 1 h, electrophoresis was carried out for 30 min, PI/DAPI staining was performed after cleaning, and data were collected by fluorescence microscope. Use the CASP software for analysis.
Apoptosis assay
The Annexin V/FITC kit was used according to the manufacturer’s instructions (C1062M; Beyotime, China). Cells with a density of 1 × 105 cells/well were plated in 6-well plates. After overnight culture, exposure in X-ray. A flow cytometer (Agilent) was used to detect apoptotic cells. Data were analyzed by FlowJo software (Version 10).
Dual-luciferase reporter gene assay
HEK293T cells were co-transfected with 100 ng of reporter plasmid and control empty vector or various expression plasmids. Forty-eight hours after transfection, luciferase activity was measured using the Dual-Lucy Assay Kit (Solarbio, China, D0011) according to the manufacturer’s instructions.
Reagents and antibodies
Antibodies and sources used in this study are as follows: anti-PPP6C (ab70634; Abcam, USA), anti-IRF3 (4302; Cell Signaling, USA), anti-phospho-IRF3 (29047; Cell Signaling, USA), anti-TBK1 (3504; Cell Signaling, USA), anti-phospho-TBK1 (5483; Cell Signaling, USA), anti-p65 (8242; Cell Signaling, USA), anti-phospho-p65 (3033; Cell Signaling, USA), anti-STING (13647; Cell Signaling, USA), anti-phospho-STING (50907; Cell Signaling, USA), γ-H2AX (R381558, ZEOBIO, China), cGAS(29958-1-AP, proteintech, China), PARP (13371-1-AP, proteintech, China), Tubulin (11224–1-AP, proteintech, China), β-Actin (R23613, ZEOBIO, China), GAPDH (R24404, ZEOBIO, China), and HRP-secondary antibodies (550036/511203 ZEOBIO, China).
High-throughput sequencing data
The original data has been uploaded to the GEO database, GEO data ID: GSE273437.
Quantitative and statistical analysis
Statistical analysis was performed using the GraphPad Prism version 8 (GraphPad Software, Inc.). For all numerical values, the mean and error bars show SD. p < 0.05 was considered statistically significant using the t-test (two-tailed).
Footnotes
Acknowledgment
The authors thank Prof. Rui Liu, State Key Laboratory of Oral Diseases Research, for his valuable discussions on this work.
Availability of Data and Materials
The datasets used and analyzed during the current study are included in the article and the supplementary materials.
Authors’ Contributions
L.G. and Y.L.: Conducted experiments and data analysis. L.G. and J.L.: Wrote the article. Z.Z., W.D., J.W., Y.L., Y.X., L.Z., H.P., M.K., T.A., and N.A.: Participated in the project management and contributed to the formal analysis. T.H., J.L., and Z.J.: Conceived and designed the study. All authors have read and approved the final article.
Author Disclosure Statement
The authors declare no conflicts of interest.
Funding Information
This work was supported by Sichuan Provincial Department of Science and Technology (2025YFHZ0091, 2025YFHZ0252); the Scientific Research Program for Young Talents of China National Nuclear Corporation (CNNC202482); the Nuclear Medical Technology Innovation Project (ZHYLYB2021011); the Chengdu Health Commission’s Urban Medical Research Project (2024347); the Sichuan Medical Research Youth Innovation Initiative (Q2024029); and the Foundation of Chengdu Medical College (2022LHFSZYB-02, 23LHHGYMP05).
Supplementary Material
Supplementary Table S1
Abbreviations Used
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.