
Abstract
Renal transplantation is an effective treatment for severe chronic kidney diseases. However, young patients often face a scarcity of kidneys from donors of similar age, resulting in the transplantation of older organs, which increase the risk of graft rejection and several complications compared with older individuals who receive kidneys from donors of similar age or younger. This article focuses on studying different senescence biomarkers in donors and patients who received kidneys from various age ranges complying with the STROBE requirements. We studied 61 patients subjected to renal transplant isolating blood samples 24 h before, and 24 h, 3 days, 7 days, 3 months, and 6 months after transplant. The patients were divided into three groups: older donor than the patient (Old Donor), younger donor than the patient (Young Donor), and similar age (Matched). We studied different senescence markers such as p16, p21, interleukin 6 (IL-6), and senescence-associated secretory phenotype (SASP) release. Young patients who receive older organs showed increased mRNA and protein expression of the senescence makers. Hence, increased SASP release was also observed in patients from older donor. In contrast, older patients who receive younger organs showed a slow but consistent improvement in their initial senescent phenotype. In addition, macrophage cell model treated with blood-derived serum from patients 6 months after the transplant showed a pro-senescence environment in macrophages proposed by the SASP from the patients. These results lead the hypothesis that senolytics could reduce the presence of senescent cells and mitigate the complications associated with the transplantation of older organs in young patients.
Introduction
Kidney transplant offers significant advantages in terms of survival for patients with end-stage renal disease compared with dialysis (Augustine, 2018). Although the immediate risk of death is higher after transplantation, it decreases significantly within a few months. Among those who benefit the most from transplants in terms of extended lifespan are younger individuals, particularly those with diabetes. For instance, individuals aged 20–39 live ∼8 years on dialysis compared with 25 years after receiving a transplant (Kaboré et al., 2017).
Recent advancements have led to notable improvements in post-transplant survival and quality of life. In response to the limited availability of living and deceased organ donors, elderly individuals are increasingly participating as donors (Sergeyenko et al., 2018). Age alone is no longer considered a contraindication for renal transplant. However, studies have indicated that patients receiving a kidney transplant during adolescence or young adulthood face a higher risk of graft loss compared with those transplanted at other ages (Augustine, 2018; Sergeyenko et al., 2018). Specifically, young patients with a graft from a deceased donor are at a higher risk of graft failure than those with a graft from a living donor.
Innovation
Kidney transplantation is the preferred treatment for severe kidney failure, improving patient quality of life and reducing mortality compared with hemodialysis; however, it is not exempt from numerous complications during the process and even long after the transplant, which worsens the quality of life of the patients, inflammation being a large part of the problem. In this study, we show that senescence are mediating the senescence-associated secretory phenotype (SASP)-mediated inflammation during the transplant being associated to the donor age on which old donors induce significant activation of senescence cells and SASP release. The outcomes described in this article may serve as a new way for designing treatments to improve the quality of life of transplanted patients.
Similarly, patients with a donor older than 40 years have a higher risk of graft failure compared with those with a donor younger than 20 years (Sergeyenko et al., 2018). Several factors may contribute to this phenomenon, including the immunosuppressive regimen (Xu et al., 2018) and physiological differences between the donor and the recipient, such as the aging process, which also affects the pharmacokinetics and pharmacodynamics of immunosuppressive drugs and other medications (Xu et al., 2018).
Cellular senescence refers to a permanent state of cell cycle arrest that occurs in proliferating cells under various stresses. The senescent state is characterized by a failure to re-enter the cell cycle in response to mitogenic stimuli, an enhanced secretory phenotype, and resistance to cell death (Uyar et al., 2020). Senescence occurs in various tissues during physiological and pathological processes such as tissue remodeling, injury, cancer, and aging. It is considered one of the causal processes of aging and is responsible for aging-related disorders (Uyar et al., 2020). Studies using mouse models have demonstrated that transplanting small numbers of senescent cells into young mice is sufficient to cause prolonged physical dysfunction (Le Meur, 2015).
Results
This study focuses on analyzing various senescence biomarkers after kidney transplantation. The senescence biomarkers examined include gene and protein expression of P16 and P21, the presence of γH2AX histone, the senescence-associated secretory phenotype (SASP) profile, and the release of interleukin 6 (IL-6). In addition, we measured the concentration of IL-6 in the media of a macrophage cell model after supplementing it with patient serum.
We selected individuals from different age groups and divided them into three categories: older donor than the patient (Old Donor), younger donor than the patient (Young Donor), and similar age (Matched). The selection was based on sample availability, absence of severe complications, and higher age difference. Clinical data for all patients are provided in Supplementary Table S1. Three patients were selected in for each group.
The mean age of older donors was ∼60 years, whereas the mean age of younger donors was 45 years. The similar age group was defined as having a difference of <5 years in age, with a mean age of 60 years. According to the human transplant protocol, immunosuppressant treatments commence before organ transplantation to prevent immune responses and enhance success rates. Owing to ethical considerations, it is not feasible to conduct studies of this nature without administering immunosuppressant treatments to patients during sample isolation. Therefore, all our patients received these treatments at the time of every sample collection.
First, we conducted an immunofluorescence assay on pretransplant donor kidney biopsies, focusing on the γH2AX histone, which is associated with DNA damage and senescence (Le Meur, 2015; Uyar et al., 2020). Biopsies from donors spanning various age groups were chosen, including individuals aged 23, 43, 57, and 80 years. Our findings demonstrated a gradual elevation in γH2AX expression corresponding to advancing age (Fig. 1A and Supplementary Fig. S1A).

Then, we studied the expression levels of two main genes and proteins associated with senescence and aging in the patients: p16Ink4a and p21 (Iske et al., 2020). Blood samples were collected from patients at various time points: pretransplant, 24 h, 72 h, 7 days, 3 months, and 6 months post-transplant. RNA and proteins were extracted from their peripheral blood mononuclear cells (PBMCs). Our findings revealed a correlation between donor age and p16Ink4a expression levels. Young patients who received an older kidney showed an increase in p16 expression over time, whereas those who received a younger kidney experienced the opposite effect (Fig. 1B).
However, in the final time point, the old donor group exhibited a reduction in p16 gene and protein expression, possibly due to a compensatory mechanism. The matched group demonstrated consistent p16Ink4a expression over time, likely because the average age between the donor and patient was similar. Moreover, p21 gene expression remained high across all groups and time points, although protein expression increased over time in the Old Donor and Matched groups (Fig. 1C). Loading controls can be found in Supplementary Figure S2.
Because senescence promotes the production of cytokines, chemokines, proteases, and growth factors known as SASP, we determine the inflammatory grade of the patients assessing a panel of SASP components in serum by cytokine array. Middle (<60 years old) and old (>60 years old) age patients showed increased pretransplant SASP profile (Supplementary Fig. S3). These results suggest that the release of SASP factors is associated to the age of the patients. Furthermore, a comparative analysis of the patients according to the donor age showed a more significant SASP factors levels in both groups with old donor such as matched and old donor groups, which was more significant in the previous and final time points 24 h, 72 h, and 6 months post-transplant (Fig. 2A).
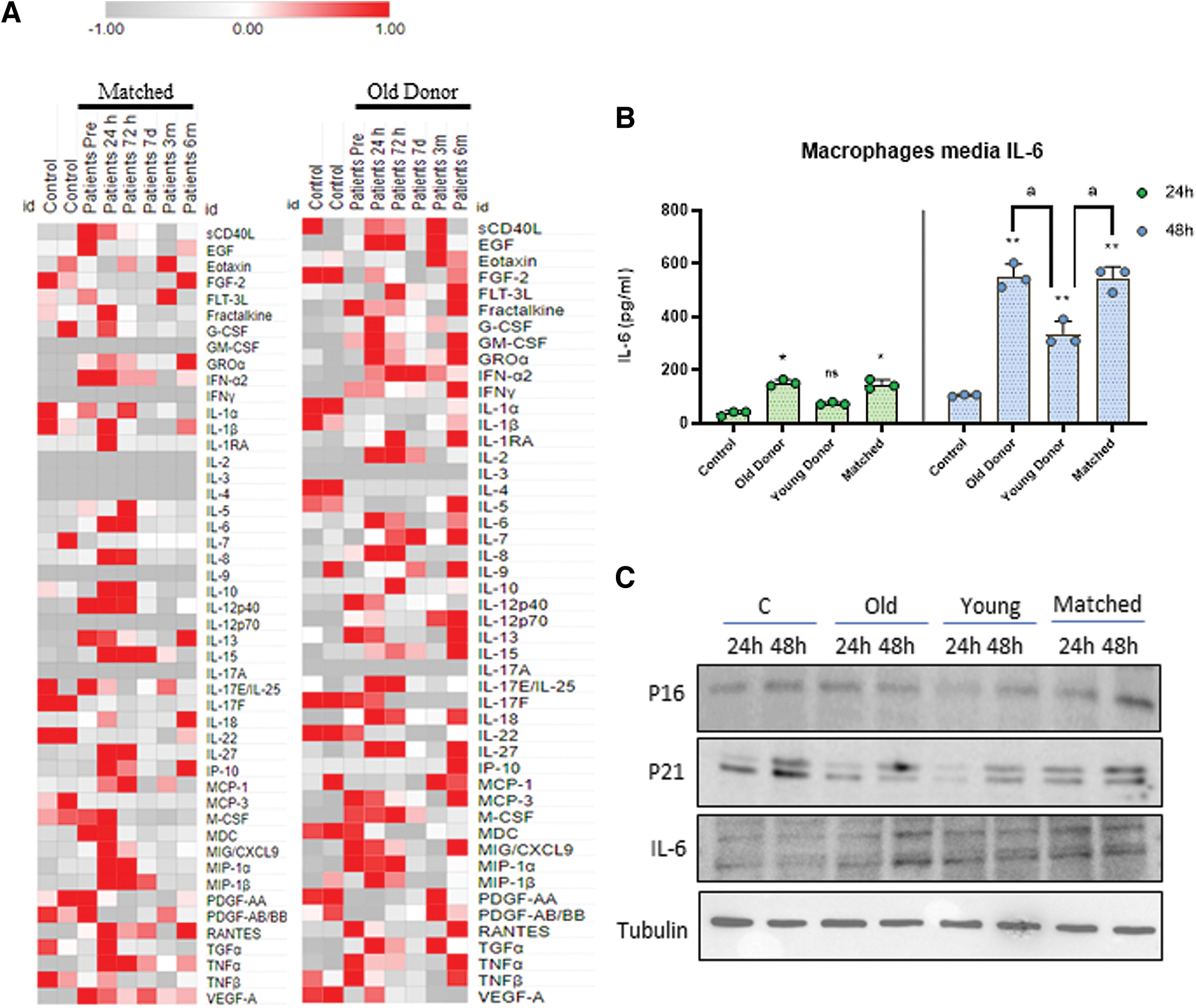
Next, we attempted to mimic each patient's inflammatory environment in a cellular model using THP-1-derived macrophages. After growing and differentiating the THP1 cells into macrophages, we replaced the fetal bovine serum (FBS) medium with patient serum at 6 months post-transplant and measured IL-6 release after 24 and 48 h (Fig. 2B). Both the 24- and 48-h time points exhibited a similar pattern, with a slight increase in IL-6 levels observed in the young donor group compared with the control, and a higher increase observed in the old donor and matched groups.
Furthermore, we conducted an analysis of protein expression under identical conditions for P16, P21, and IL-6 (Fig. 2C). Overall, these proteins exhibited a consistent pattern, with the Young Donor group showing lower expression compared with the other groups. These findings support previous studies suggesting that the emergence of a group of senescent cells contributes to a pro-senescence environment within the entire organism (Iske et al., 2020; Le Meur, 2015; Uyar et al., 2020).
Discussion
The concept of transferring the senescence phenotype is not new (Uyar et al., 2020). With aging, senescent cells accumulate and have been identified as crucial in promoting the immunogenicity of older organs, which accelerates alloimmune responses (Kaboré et al., 2017). Senescent cells can induce a senescent phenotype in neighboring cells, which could potentially explain how the engraftment of older organs may contribute to the spread of senescence. Studies in mouse models have demonstrated that depleting senescent cells using senolytics can significantly improve a wide range of age-associated disabilities and diseases (Le Meur, 2015; Uyar et al., 2020).
In contrast to the potential transfer of senescence to recipients, it is also possible that a young donor organ could have rejuvenating effects when transplanted into an older recipient. Although there are limited studies on this topic, research involving young parabiotic animals has shown the ability to rejuvenate brain, heart, and muscle function in older mice (Villeda et al., 2014). In addition, the transfer of plasma derived from young mice to older mice has been found to enhance neuronal plasticity in the hippocampus and improve cognitive functions, supporting the concept of rejuvenation (Villeda et al., 2014).
Overall, our findings support these hypotheses and may help explain why older donor organs are associated with higher rates of acute rejection. At the same time, we suggest that transplanting an organ from an elderly donor may pose risks for young recipients (Fig. 3). This effect underscores the need to explore age-specific treatment approaches, such as senolytics, which could improve the quality of older organs while reducing immunogenicity.

Despite potential age-related changes in immunology and pharmacology, immunosuppression protocols are generally similar regardless of age, lacking specific age-based guidelines (Sergeyenko et al., 2018). However, recent research suggests that senescent cells create a basal immunosuppressive microenvironment that disrupts tissue homeostasis (Tchkonia et al., 2013). This process is associated with “inflammaging,” a chronic low-grade inflammation that occurs with aging and leads to a state known as “immunosenescence” (Tchkonia et al., 2013; Uyar et al., 2020).
Age-related immunosuppression involves the activation of immunosuppressive phenotypes, impairing the surveillance and clearance of senescent cells. Consequently, the accumulation of pro-inflammatory senescent cells further amplifies inflammation and immunosuppression. This self-reinforcing process contributes to inflammaging and progressively worsens age-related tissue degeneration.
Although immunosuppression is necessary after organ transplantation, combining immunosuppressive treatment with the presence of an older allogeneic organ may exacerbate aging-like conditions at an organism level in younger patients. Optimizing the utilization of available organs for transplantation, including those from older donors, remains crucial for bridging the gap between organ demand and supply. Employing senolytic drugs that selectively deplete senescent cells or targeted anti-inflammatory drugs that alleviate inflammaging may represent potential approaches to improve outcomes for older organs while limiting the spread of senescence and its associated side effects.
Limitations of the study
Despite the interest of our findings, we recognize some limitations of our study. First, many of these results are derived from the blood of the transplanted patients. Thus, most likely, more notable results would be obtained in case of having the possibility to study the senescence cells and SASP in other specific tissues. Second, the confirmation of our results in larger cohorts is also desirable. In this sense, we are trying to increase the repository of this precious samples. Thus, further research on this topic is warranted.
Notes
Biological samples
Ethical statements
Approval of the ethical committee of the Hospital Puerta del Mar was obtained, according to the principles of the Declaration of Helsinki and all the International Conferences on Harmonization and Good Clinical Practice Guidelines. All the participants in the study gave their written informed consent before initiating it. The samples obtained during this study was part of the samples isolated during the routine procedure with the patients complying with the STROBE requirements.
Kidney transplanted patients were recollected from Puerta del Mar Hospital between January 1, 2021 and January 9, 2022 who received a kidney transplantation from young (mean age 45 years) and old donor (mean age 60 years) (Supplementary Table S2). In total, we recruited 61 patients. We perform a preimplantation biopsy in all kidney grafts. All patients had a minimum follow-up of 1-year post-kidney transplant.
An immunosuppression regimen was administered to kidney transplant recipients to prevent rejection episodes and loss of the renal allograft. The components were tacrolimus, mycophenolate mofetil, steroids, and basiliximab/thymoglobulin.
Kidney biopsy samples of the upper pole of the kidney graft are taken by expert nephrologists using the 16-gauge biopsy under ultrasound guide. All kidney biopsies were processed by standard techniques for light microscopy and immunofluorescence. Renal tissues were fixed in 10% neutral buffered formalin and embedded in paraffin. The processed sections were 5 μm each.
Blood samples were collected at various time points: before transplantation, as well as at 24 h, 72 hurs, 7 days, 3 months, and 6 months post-transplantation. PBMCs were obtained from patients (Supplementary Table S2) blood using Histopaque 1077 (Sigma-Aldrich, San Lois, MO). Protein and RNA were obtained from patient PBMCs using the AllPrep RNA/Protein Kit (Qiagen, Hilden, Germany).
Commercial THP-1 monocyte cells line was purchased from ATCC (Manassas, VI; TIB202).
Reagents
Bovine serum albumin (BSA; A7030), Triton X-100 (X100-100ML), and phorbol-12-myristate-13-acetate (PMA; P8139) were purchased from MERCK (Darmstadt, Germany). The following reagents were obtained from Thermo Fisher Scientific (Waltham, MA): Secondary Alexa Fluor 488 Goat Anti-Rabbit Antibody (A-21424), QuantumRNA™ Classic 18S Internal Standard (AM1716) and RPMI 1640 medium (31870074).
Primary antibodies: anti-p16 INK4A human (18769S; Cell Signaling, Danvers, MA), anti-p21 NBP2-29463 (NBP2-29463; Novus Biologicals, Centennial, CO), anti-gamma H2AX (ab81299; Abcam, Cambridge, United Kingdom), anti-IL-6 (sc-32296; Santa Cruz Biotechnology, Santa Cruz, CA).
Cell culture
THP-1 cells were seeded in six-well plates in RPMI medium supplemented with 10% FBS and 1% antibiotics (11548876; Thermo Fisher) until 90% confluence. Then, THP1 cells were differentiated into macrophages using PMA at 50 nM for 24 h. For patient's serum assay, THP1-derived macrophages medium was refreshed with RPMI without FBS and supplemented with patient 6 months post-transplant serum. Finally, medium was aliquoted for enzyme-linked immunosorbent assay (ELISA) analysis.
Immunofluorescence
Immunofluorescent staining of paraffin-embedded sections was performed following standard methods. Paraffin sections were attached to superfrost plus slides (Menzel-Glaser, Braunschweig, Germany) at 60°C for 1 h. Deparaffinization was performed by pure xylol washes three times for 10′ each. Slides were rehydrated by ethanol solutions immersion (from 100% to 70%) for 5′ each and rinsed with deionized water.
For the heat antigen retrieval, slides were immersed in sodium citrate 10 mM (unmasking solution) and microwaved at 800 W for 15′, then samples were kept at room temperature until cool down. Slides were rinsed with phosphate-buffered saline (PBS) 1 × three times, and then blocking solution (2% BSA, 0.05% triton X-100 in PBS 1 × ) was applied for 1 h. The samples were surrounded with a hydrophobic barrier using a barrier pen and primary antibody was applied at 1:100 concentration diluted in blocking solution overnight.
The next day, slides were rinsed three times with PBS 1 × and secondary antibody was applied at 1:400 concentration diluted in blocking solution for 2 h. Again, slides were rinsed three times with PBS 1 × and DAPI (sc-3598; Santa Cruz Biotechnology) staining (1 μg/mL) was applied for 10′. Finally, samples were mounted with coverslips in using Vectashield Mounting Medium (H1000; Vector Laboratories, Burlingame, CA).
IL-6 measurement
IL-6 ELISA kit was purchased from Abcam (ab178013). Assays were performed using patient blood serum or culture medium according to manufacturer's protocol.
SASP array
Blood serum were analyzed for expression of several mouse cytokine and chemokines (MD44) or human cytokine and chemokines (HD48), respectively, using a Multiplexing LASER Bead Assay (Eve Technologies).
Gene expression
Expression of P16 and P21 SYBR Green (RR420W; Takara, Kusatsu, Japan) quantitative polymerase chain reaction (qPCR). cDNA was obtained from extracted mRNA using the iScript cDNA KIT (Biorad, Hercules, CA). The qPCR was performed in a CFX96 Connect Real-Time PCR Detection System (Biorad). Ribosomal 18S expression was used as reference gene (QuantumRNA Classic 18S Internal Standard, AM1716; Thermo Fisher).
Western blotting
PMBCs extracted proteins were used for standard protein electrophoresis and Western blot assays. Gel electrophoresis were performed using 4%–20% Mini-PROTEAN® TGX Stain-Free™ Protein Gels, Biorad at 200 V in Tris-Glycine-SDS buffer for 40′. Protein transfer was made using a TurboTransfer (Biorad) at 25 V for 7′. After transferring the proteins to 0.45 μM nitrocellulose membranes (Biorad), these were incubated for 1 h in BSA 5% in PBS-Tween20 0.05% and then overnight at 4°C with primary antibodies at 1:1000. Then washed twice with PBS-Tween20 and incubated with the corresponding secondary 1:5000 antibody for 1 h at room temperature. Protein loading was checked using stain-free gel activation. Stripping was not used.
Statistics
Nonparametric Student's t-test (Mann–Whitney) was used to compare data between two groups. The Mann–Whitney U test does not rely on the assumption of normal distribution in the data. It is particularly useful when dealing with ordinal or interval data that may not meet the normality assumption required by parametric tests. It can be applied to a wide range of data types, including ordinal, interval, and some ratio data. The test is robust against outliers and skewed data, making it a good choice when dealing with data sets that may have extreme values. The Mann–Whitney U test is sensitive to differences in both central tendency and shape of the distribution, making it versatile for various types of comparisons.
The test assumes that the observations in one group are independent of the observations in the other group. If this assumption is violated, the results may be invalid. It assumes that the shapes of the distributions in the two groups are similar. If the shapes are substantially different, the Mann–Whitney U test may not provide an accurate assessment of differences. Finally, the test may be less accurate when there are ties in the data (i.e., multiple observations with the same value). Some adjustments or corrections may be necessary in such cases.
All results are expressed as mean ± standard deviation of three independent experiments and a p value <0.05 was considered as statistically significant. Level of significance is denoted by asterisks *p < 0.05, **p < 0.01.
Footnotes
Acknowledgments
We thank to all patients for their participation in this study.
Authors' Contributions
M.D.C. and A.M. designed the study. J.M.S.-R., J.L.P., and I.M.-Z. performed the experiments. R.V.M., A.A., T.G., A.N.-D., and A.M. performed the patient selection and according to the protocol of transplant from the hospital. J.M.S.-R. J.L.P., A.M., A.N.-D., and M.D.C. discussed and analyzed the data. M.D.C. and J.M.S.-R. drafted the article. All the authors revised the article and approved the final version.
Funding Information
This study was supported by PI21/01656 grant, Instituto de Salud Carlos III, Spain and SATOT fellowship, Spain. Juan Miguel Suarez-Rivero has the benefit of a postdoctoral fellowship (FPU 13/03173) from Consejería de Sanidad, Junta de Andalucía, Spain.
Supplementary Material
Supplementary Table S1
Supplementary Table S2
Supplementary Figure S1
Supplementary Figure S2
Supplementary Figure S3
Abbreviations Used
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.