
Abstract
Physical or chemical stress is commonly known to inhibit protein translation at the cellular level. Since the process of protein translation requires catalysis by a multi-component machinery containing eukaryotic initiation factors (eIFs) and ribosomes in a sequence of reactions, how the process fails to proceed and whether certain genes can escape such blockade have provoked research efforts. Lines of evidence have demonstrated that phosphorylation of eIF4E or dephosphorylation of 4E-binding proteins (4E-BPs) prevents the formation of the eukaryotic translation initiation factor 4F (eIF4F) complex, whereas phosphorylation of eukaryotic translation initiation factor 2 alpha (eIF2α) due to activation of heme-regulated inhibitor (HRI), general control nonderepressible 2 (GCN2), protein kinase RNA-like endoplasmic reticulum kinase (PERK), or protein kinase R (PKR) by a diverse array of stressors prevents eIF2-GTP-tRNAi Met ternary complex assembly. These signal the abandonment of translation initiation via 5′-7-methylguanine (m7G) cap recognition by eIF4E. Stress can promote cleavage of tRNAs, impediment of rRNA processing, changes in the epitranscriptomic landscape, ribosome stalling or collision, activation of ribosomal surveillance systems, and assembly of the stress granules. Although these events contribute to the general inhibition of protein translation, a few proteins can bypass such negativity and become translated selectively. Such selective protein translation is primarily m7G cap independent through the integrated stress response or Internal Ribosomal Entry Site (IRES). The newly synthesized proteins often influence cell fate, facilitate cell survival, and build endogenous defense. Insights into the general inhibition of protein translation and selective translation of specific proteins will advance our understanding of the etiology or progression of human diseases involving cellular stress from viral infection or inflammation to myocardial infarction, stroke, or neurodegenerative disease. Antioxid. Redox Signal. 40, 943–947.
The description of physical or chemical stress originated from experimental evidence of protein synthesis inhibition. Such coupling was first observed in the mid-20th century when cell culture systems and molecular biology tools became available. New protein synthesis occurs continuously in healthy cells to feed proliferation or to maintain metabolism. When mammalian cells encounter a stressful condition, such as nutrient deprivation, temperature extremes, radiation, or exposure to xenobiotics, an immediate response is cessation of protein synthesis. This event can be detected by decreases in amino acid incorporation into the cells. Historically, inhibition of protein synthesis has served as a standard measure for cellular stress. How the process of protein translation ceases its operation has been a recent topic of research.
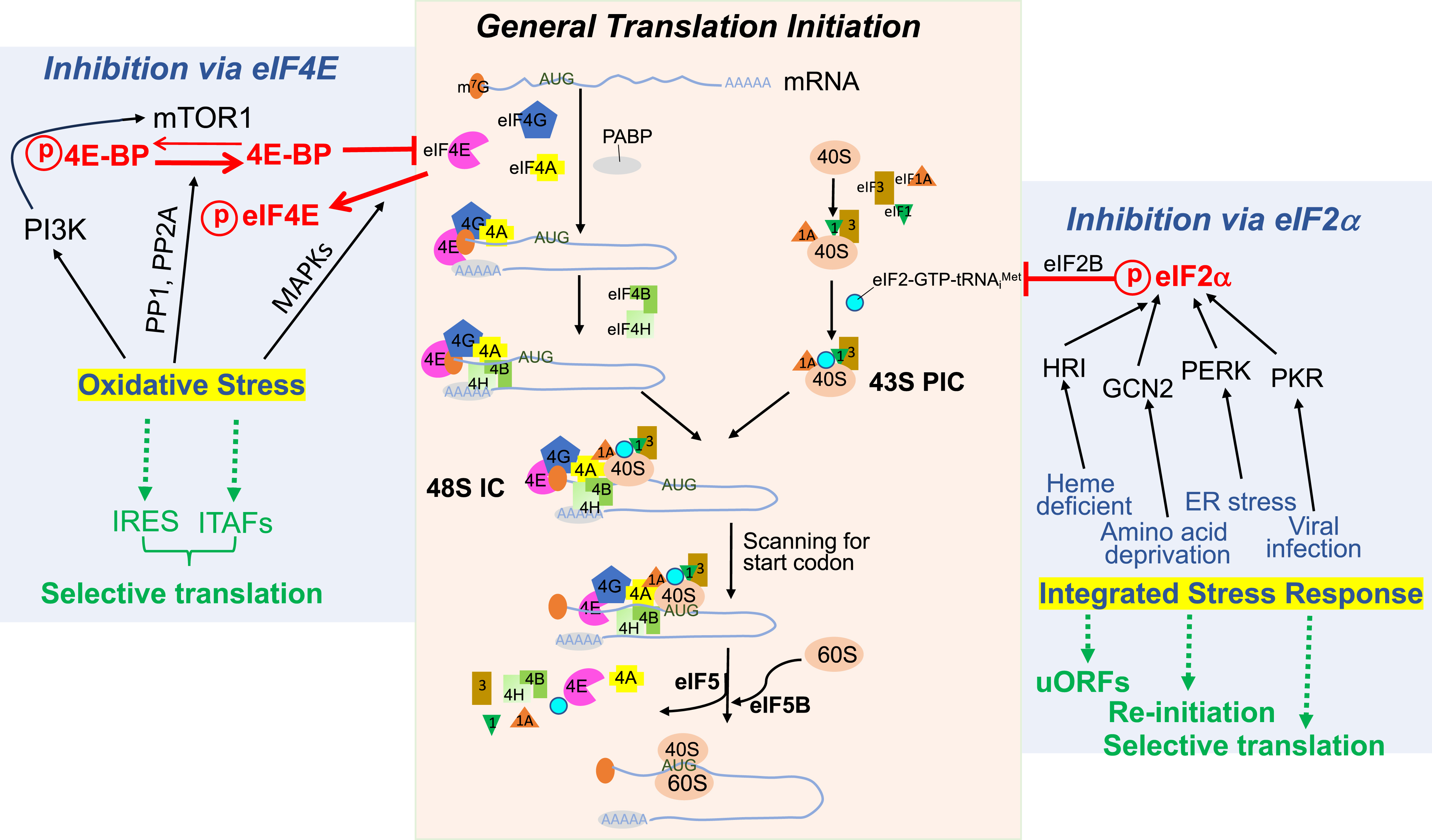
A protein is translated from its encoding mRNA in three sequential steps: initiation, elongation, and termination, with the initiation being a rate-limiting step. Translation initiation requires engagement of the translation machinery to a target mRNA strand (Brito Querido et al., 2024, Holcik and Sonenberg, 2005). The train of the translation machinery contains 12 eukaryotic initiation factors (eIFs) plus poly A binding proteins (PABPs) (Table 1). The process starts with eukaryotic translation initiation factor 4E (eIF4E) recognizing and binding to the 5′-7-methylguanine cap (m7G) cap of an mRNA strand. The eIF4E protein belongs to the eIF4F complex, which contains the eIF4G scaffold protein and the ATP-dependent helicase eIF4A. Binding of the eIF4F complex to the 5′ m7G cap of mRNAs triggers the joining of eIF4B and eIF4H, the enhancers of eIF4A helicase, for the removal of inhibitory RNA structures (Fig. 1, middle panel).
List of eIFs for Protein Translation Initiation
eIF1, eukaryotic initiation factor 1; eIF1A, eukaryotic translation initiation factor 1A; eIF2, eukaryotic initiation factor 2; eIF2B, eukaryotic translation initiation factor 2B; eIF3, eukaryotic initiation factor 3; eIF4A, eukaryotic translation initiation factor 4A; eIF4B, eukaryotic translation initiation factor 4B; eIF4G, eukaryotic translation initiation factor 4G; eIF4E, eukaryotic translation initiation factor 4E; eIF4H, eukaryotic translation initiation factor 4H; eIF5, eukaryotic initiation factor 5; eIF5B, eukaryotic translation initiation factor 5B; eIF2α, eukaryotic translation initiation factor 2 alpha; eIF2β, eukaryotic translation initiation factor 2 beta; eIF2γ, eukaryotic translation initiation factor 2 gamma; m7G, 7-methylguanine cap; PABP1, poly A binding proteins 1; PIC, pre-initiation complex; tRNAi Met, initiator methionine.
In parallel with the recognition of target mRNA and removal of inhibitory structures, the 43S pre-initiation complex (PIC) is assembled from the 40S small subunit of the ribosome by joining of eIF1, eIF1A, eIF3, and the ternary complex of eIF2-GTP-tRNAi Met. Attaching of the 43S PIC to the mRNA strand preoccupied with the eIF4F complex forms the 48S initiation complex (IC), which is capable of scanning for the start codon on the mRNA strand and recruiting the 60S large subunit of the ribosome to form the 80S IC. Addition of Met to the start codon marks the completion of the translation initiation phase (Fig. 1, middle panel).
Growth factors activate the phosphoinositide 3-kinase (PI3K) signaling pathway to trigger an onset of translation initiation. When PI3K is activated, its downstream protein kinase B (PKB), also known as Ak strain transforming gene (Akt), removes tuberous sclerosis complex 1 or 2 (TSC1/2) inhibition on mammalian target of rapamycin (mTOR1) kinase. This allows mTOR1 to phosphorylate 4E-binding proteins (4E-BPs), an inhibitory partner of eIF4E, leading to 4E-BP dissociation from eIF4E. This enables eIF4E to tag the 5′ m7G cap of target mRNA stands for translation initiation.
Stressors, exemplified by oxidants, hypoxia, or arsenite, ignite a network of signaling events, with two main branches being mitogen-activated protein kinases (MAPKs) and PI3K. Downstream of p38 MAPK or extracellular signal-regulated kinases (ERKs) is eIF4E phosphorylation by MAPK-interacting kinases-1/2 (Mnk-1/2). This phosphorylation hinders the formation of the eIF4F complex, therefore blocking its engagement to the target mRNA strand (Fig. 1, left panel).
Although stressors such as oxidants activate PI3K, differing from growth factors, oxidants can also activate phosphatases, i.e., phosphatase 1 (PP1) and phosphatase 2A (PP2A). PP1 or PP2A, in turn dephosphorylates 4E-BPs, resulting in the sequestration of eIF4E. As a result, 4E-BP binding to eIF4E prohibits eIF4E recognition of the 5′ m7G cap and prevents the eIF4F complex from landing on an mRNA strand for translation initiation.
The integrated stress response senses a myriad of stressors for altering protein translation (Wek et al., 2023). The concept of the integrated stress response, developed after 1992, has provided insights into how cellular stress is communicated to inhibit general protein translation (Dever et al., 1992). Immediate activation of heme-regulated inhibitor (HRI), general control nonderepressible 2 (GCN2), protein kinase RNA-like endoplasmic reticulum kinase (PERK), or protein kinase R (PKR) kinase by a specific type of stress causes eukaryotic translation initiation factor 2 alpha (eIF2α) phosphorylation (Fig. 1, right panel). As an engine for the eIF2-GTP-tRNAi met ternary complex assembly and landing of the 43S PIC cargo onto the target mRNA track, eIF2α phosphorylation results in an increased affinity for the eIF2B GDP/GTP exchange factor. The multimeric eIF2B protein normally replenishes eIF2-GTP from eIF2-GDP, preparing for the eIF2-GTP-tRNAi Met ternary complex joining the 40S ribosome to form the 43S PIC. As a result of eIF2α phosphorylation, eIF2B prohibits the formation of the eIF2/GTP/tRNAi Met complex.
Whereas phosphorylation of eIF4E, dephosphorylation of 4E-BPs, or phosphorylation of eIF2α provides a satisfactory answer to the pathways leading to translation cessation, stress can also cause ribosomes to stall or collide on the mRNA track (Fig. 2). The ribosomal surveillance systems deal with such ribosomal deformities by ribosomal subunit splitting, ribosomal quality control, and ribosomal integrated stress response (Vind et al., 2023). A rapid evolvement in the range of ribosomal surveillance systems leads to a growing list of unique proteins with designated functions for ribosomal subunit splitting, repressing ribosomal participation in bulk protein translation, ubiquitination and degradation of peptidyl-tRNA-ribosome complex, or catalyzing a ribotoxic response into inflammation or cell death.
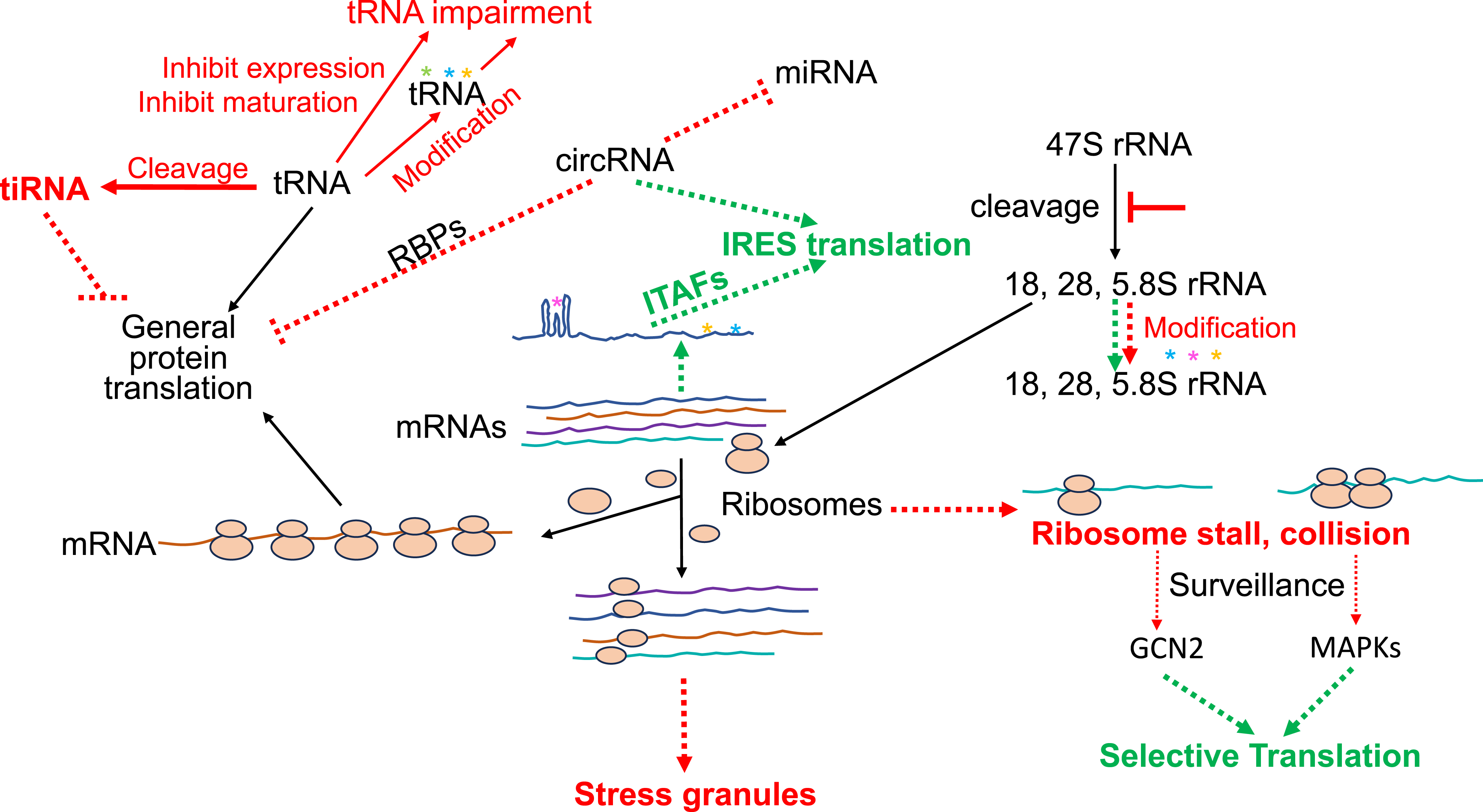
The discovery of stress granules has evoked our appreciation for biological evolution. After all, giving up protein translation or cell survival has expensive consequences, requiring extensive resources and energy for replacement. Apparently, the stalled 43S PICs on the track of target mRNAs are sheltered into membraneless dense aggregates in 0.1-2.0 µm diameters (Baymiller and Moon, 2023). The fascinating world of stress granules has its own conductors, operators, and mediators, emerging from invisible seeds to microscopically visible granules, and then resolving the state of hibernation by transforming into the P-bodies for re-entry into translation-active stature.
How many transcripts are made into their encoded proteins under stress conditions from the 25,000 genes of the human genome remains a question unanswered. Do all transcripts cease to be translated into proteins during cellular response to stress? Common sense evolution argues against a total shutdown of protein translation. Indeed, selective protein translation occurs under stress conditions, serving to avoid aberrant protein synthesis yet conserve energy, since over 30% cellular ATP is consumed for protein translation (Buttgereit and Brand, 1995).
One mechanism for a functional protein to be made involves the Internal Ribosomal Entry Site (IRES), bypassing 5′ m7G cap-dependent translation. The IRES-mediated protein translation was first reported in viral proteins during infection when cellular protein synthesis was inhibited (Pelletier and Sonenberg, 1988). As it stands now, about 60 cellular genes have been reported to undergo IRES-associated protein translation (www.iresite.org). While the nature of IRES remains to be fully defined, it is clear that an IRES contains “stems”-and-“loops” or complex tertiary structures, and attracts the binding of its trans-acting factors, i.e., IRES Trans-acting factor (ITAFs) (Fig. 2). It is presumed that a unique structural fold of IRES attracts the binding of one or a set of ITAFs (Yamamoto et al., 2017). These ITAFs presumably recruit eIFs and ribosomes for translation initiation. Whether IRES is the main mechanism for protein translation under stress is an issue of controversy. Nevertheless, most proteins translated via the IRES mechanism participate in cellular defense, repair, or resistance against apoptosis. These outside-the-box findings further illustrate the beauty of evolution: when there is a stress, there are multiple mechanisms fighting against such stress.
Traditionally, the primary focus in understanding protein translation control under stress relies on eIFs, RNA binding proteins, and IRES in the 5′untranslated region (5′UTR) of mRNAs. tRNA as a component for regulating protein translation had been a neglected corner of the field, until recent revelations of noncanonical functions of tRNAs. Advances in nucleic acid sequencing technologies and mass spectrometry allow the definition of tRNA species, expression, modifications, cleavage, localization, and metabolism under oxidative stress (Akiyama and Ivanov, 2023). For example, tRNA can be cleaved by endoribonucleases to form stress-induced tRNA (tiRNA), which in turn suppresses translation initiation (Fig. 2). Additionally, tRNA may be inhibited for maturation or undergo covalent modifications. The extensive intricacies of these new entities of tRNAs undoubtedly call for comprehensive efforts to decipher the details of ins and outs.
Equally intriguing are the roles of rRNA, circular RNA (circRNA), microRNA (miRNA), and epitranscriptomic elements in protein translation control (Fig. 2). Unlike tRNAs, these non-protein-coding RNA species exhibit a broad spectrum of complexity, commanding in-depth investigations on each category of its own alike. For instance, oxidative stress inhibiting rRNA processing thereby decreasing the number of functional ribosomes is a narrative distinct from that of circRNAs, which lack both 5′UTR and 3′UTR but can sequester miRNAs or RNA binding proteins (Payea et al., 2023). These elements contribute to the artistry of inhibiting bulk protein translation without sacrificing selective synthesis of the proteins essential for cellular defense, repair, or adaptation.
Stress-associated protein translation has gained prominence owing to its linkage to a long list of human diseases, from viral infections, inflammation, and neurodegenerative diseases, to stroke, cardiovascular disease, or cancer. In theory, any disease that involves cellular stress warrants considerations of general protein synthesis inhibition versus selection of specific proteins for translation. Our understanding of protein translation occurring under stress conditions and the significance of such event will expand along with the advancement of bioscience research tools.
Footnotes
Author Disclosure Statement
No competing financial interests exist.
Funding Information
The research works at the author's laboratory is supported by NIH R01 GM125212, NIH R01 GM126165, NIH R56 HL166330, University of Arizona College of Pharmacy Start-up fund, and Holsclaw Endowment.