
Abstract
Significance:
Mitochondria are recognized as a central metabolic hub with bioenergetic, biosynthetic, and signaling functions that tightly control key cellular processes. As a crucial component of mitochondrial biogenesis, peroxisome proliferator-activated receptor gamma coactivator 1α (PGC-1α) is involved in regulating various metabolic pathways, including energy metabolism and reactive oxygen species homeostasis.
Recent Advances:
Recent studies have highlighted the significant role of PGC-1α in tumorigenesis, cancer progression, and treatment resistance. However, PGC-1α exhibits pleiotropic effects in different cancer types, necessitating a more comprehensive and thorough understanding.
Critical Issues:
In this review, we discuss the structure and regulatory mechanisms of PGC-1α, analyze its cellular and metabolic functions, explore its impact on tumorigenesis, and propose potential strategies for targeting PGC-1α.
Future Directions
: The targeted adjustment of PGC-1α based on the metabolic preferences of different cancer types could offer a hopeful therapeutic approach for both preventing and treating tumors. Antioxid. Redox Signal. 41, 557–572.
Introduction
Although numerous new strategies for cancer prevention and treatment have been implemented in clinical settings, cancer continues to be a leading cause of death globally, accounting for approximately one sixth of all deaths (Sung et al., 2021). The dysregulation of cellular metabolism, widely acknowledged as a hallmark of cancer, has emerged as a prominent area of study in cancer research (Martínez-Reyes and Chandel, 2021). To meet the energy requirements for proliferation and survival in nutrient-poor environments, tumor cells must adjust their energy supply and expenditure pathways through metabolic reprogramming, such as glycolysis or autophagy. Glycolysis has traditionally been recognized as a key metabolic process in tumors for an extended period (Paul et al., 2022). Nowadays, the role of dysfunctional mitochondria in tumors is garnering increasing attention as the central hub of cellular metabolism.
Mitochondria, as bilayer organelles with their own genomes, play a crucial role in tumor anabolism by providing essential building blocks, generating energy, maintaining redox and calcium homeostasis, participating in transcriptional regulation, and influencing cell fate (Dache and Thierry, 2023). Dysfunction of mitochondria can contribute to malignant transformation through increasing production of reactive oxygen species (ROS), overproducting of oncometabolites such as succinate, fumarate, or 2-hydroxyglutarate, and enhancing resistance to oncogenes by preventing cell death or senescence (Bhatti et al., 2017; Klemmensen et al., 2023). In addition, the plasticity and dynamics of mitochondria are crucial for conferring cellular phenotypic and metabolic plasticity, which can lead to resistance against anticancer therapies (Bhatti et al., 2017). Therefore, targeting mitochondria holds promise for cancer prevention and treatment.
Mitochondrial biogenesis involves the growth and division of existing mitochondria, including the synthesis of new mtDNA, proteins, and membranes, and is crucial for mitochondrial function (Liu et al., 2023b). Peroxisome proliferator-activated receptor gamma coactivator 1α (PGC-1α) is a key regulator of mitochondrial biogenesis and other functions like antioxidant defense and oxidative phosphorylation (OXPHOS) (Miller et al., 2019). Research indicates that PGC-1α influences tumor development, metastasis, and treatment resistance by modulating transcriptional programs and metabolic characteristics (Mastropasqua et al., 2018). However, the role of PGC-1α varies across different cancer types, with promoting effects in breast cancer but inhibitory effects in ovarian cancer. Moreover, PGC-1α can have opposing roles in different subtypes or stages of the same tumor, such as prostate cancer. Given the diverse functions of PGC-1α that are tumor specific, a comprehensive understanding of its role in tumors is essential.
Herein, we describe the structure and modulatory mechanisms of PGC-1α, analyze its function in cells and metabolism, highlight its role in tumors, and offer a new target and perspective for antitumor strategies.
A Glance of PGC-1α
Discovery of the PGC-1 family
The PGC-1 family comprises PGC-1α, PGC-1β (also known as PERC), and PGC-1 related coactivator (PRC) (Fig. 1). PGC-1α is located in the nucleus and has been identified as a transcriptional coactivator, serving as the master regulator for adaptive thermogenesis and mitochondrial biogenesis (Puigserver and Spiegelman, 2003). Puigserver et al. discovered this protein, originally named PGC-1, by screening proteins that interact with the transcription factor PPARγ from mouse brown adipocyte cDNA libraries (Puigserver et al., 1998). PRC and PGC1-β were subsequently identified as factors with high sequence similarity to PGC-1, leading to the renaming of PGC-1 as PGC-1α. Ruas et al. identified other isoforms of PGC-1α based on their transcriptional promoters. PGC-1α1, which corresponds to the previously described PGC-1α, was derived entirely from proximal promoters. The other variants, such as PGC-1α2, PGC-1α3, and PGC-1α4, were derived from other promoters (Ruas et al., 2012).
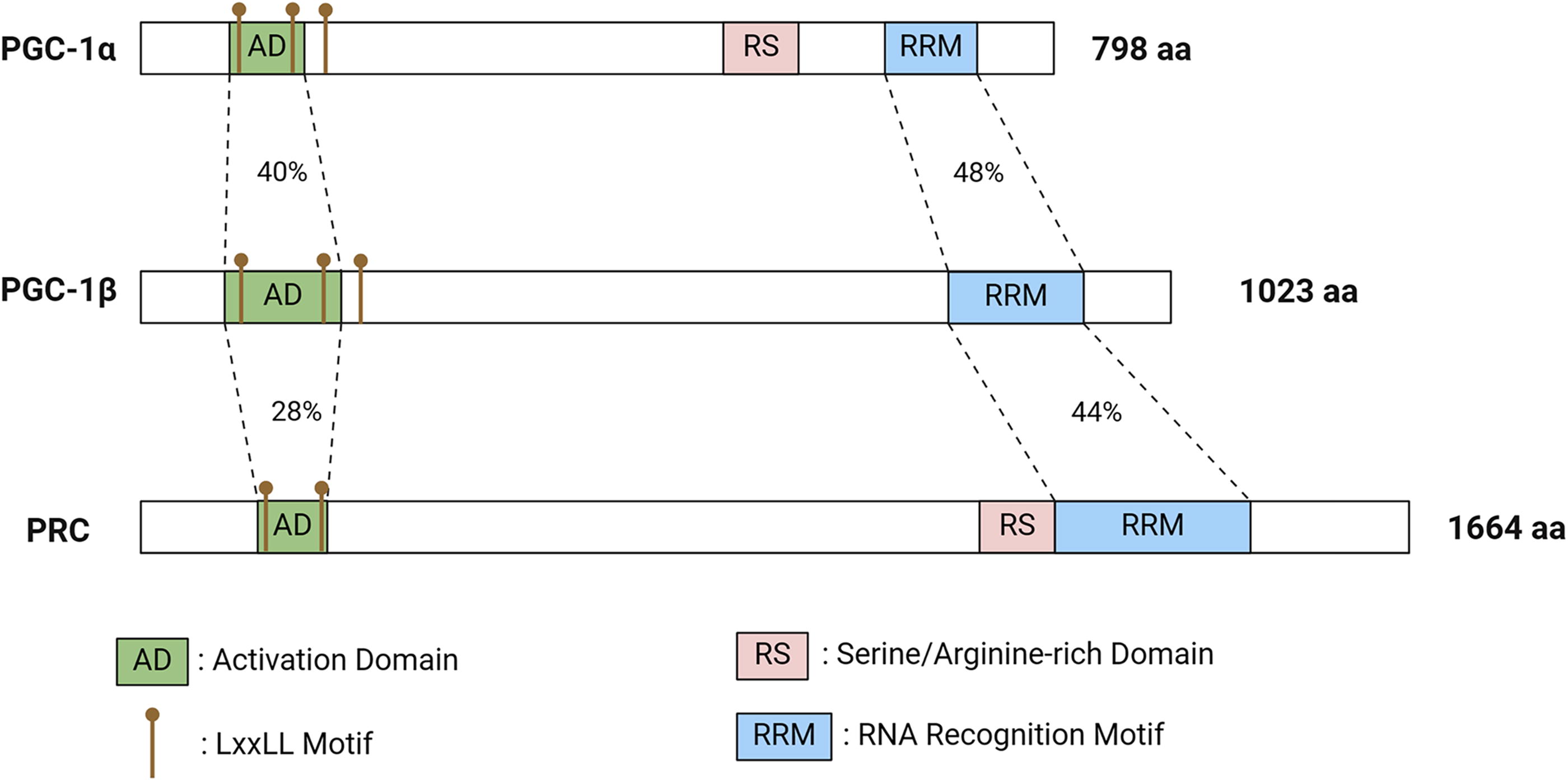
From a structural point of view, the PGC-1 family members exhibit significant variation in their overall sequences, whereas some crucial sequences exhibit high levels of homology (Fig. 1). For instance, all members possess a leucine-rich transcriptional activation domain LXXLL sequence at the N-terminal, which interacts with nuclear receptors (Luo et al., 2019). From a functional perspective, PGC-1β is primarily involved in regulating adipocyte differentiation and lipid metabolism, whereas PRC is linked to mitochondrial proliferation. PGC-1α plays a crucial role in all these metabolic processes and is recognized as a key regulator of mitochondrial biogenesis and antioxidant defense. In addition, PGC-1α can boost the transcriptional activity of nuclear receptors such as PPARγ and various steroid receptors, controlling important genes related to adaptive thermogenesis, glucose, and lipid metabolism. Overall, PGC-1α is essential for maintaining mitochondrial biogenesis and energy metabolism, which are closely associated with tumor initiation and progression.
Structure of PGC-1α
PGC-1α is a coactivator that, similar to others, has a powerful activation domain at the N-terminal that binds to various transcription factors and nuclear receptors, rather than to DNA. However, unlike other coactivators, it does not have histone acetyl transferase (HAT) activity, which promotes chromatin remodeling and gene transcription, nor a recognizable HAT domain. Instead, it serves as a platform to recruit other HAT-containing coactivator proteins, such as SRC and CBP/p300 (Bost and Kaminski, 2019) (Fig. 2a).
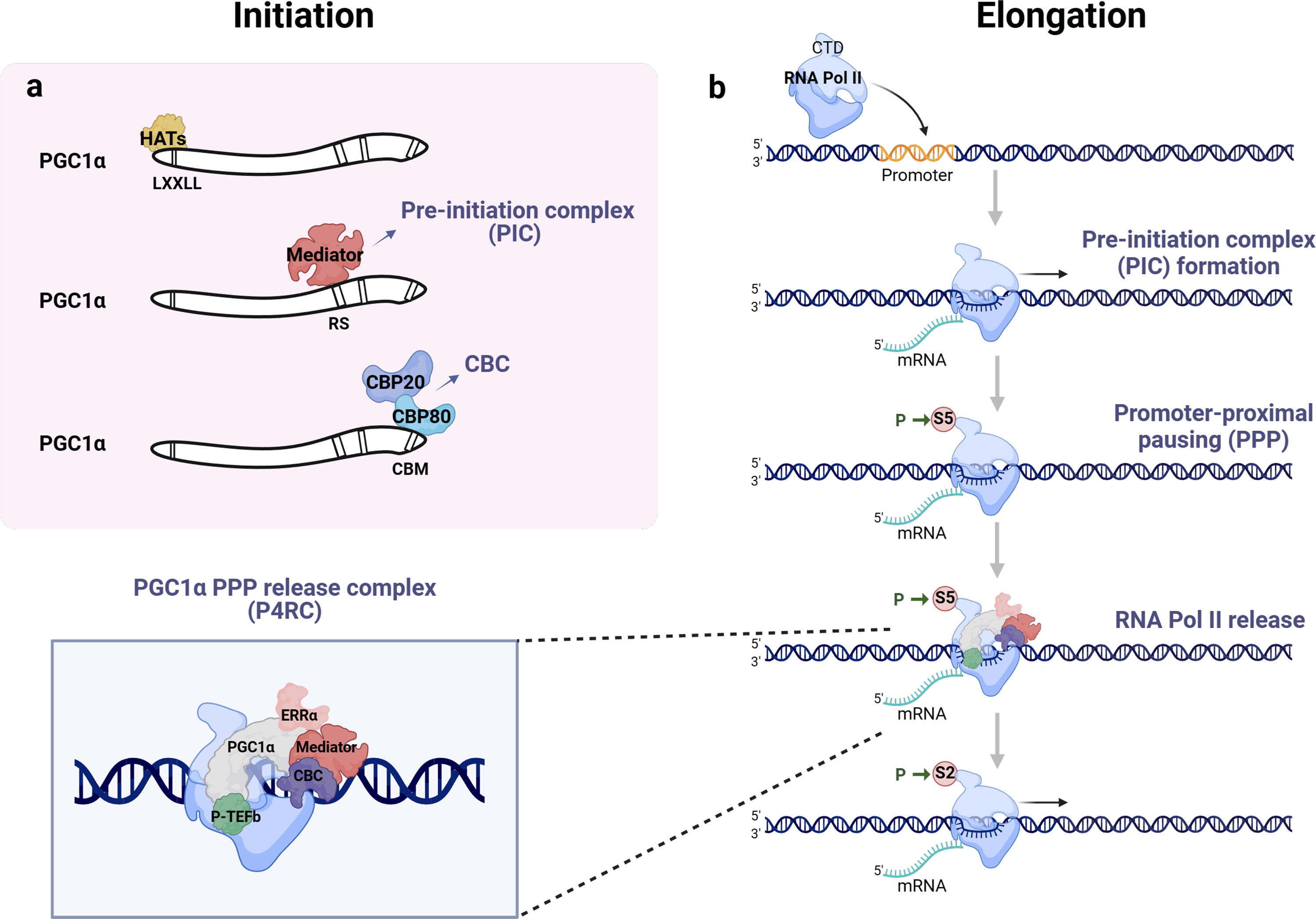
A folded structural domain known as the RNA recognition motif (RRM) is located at the C-terminus of PGC-1α. In addition, the C-terminal domain (CTD) of PGC-1α also includes arginine- and serine-rich domains referred to as RS domains (Rambout et al., 2023). Although this arrangement is similar to the SR splicing factor, their structures are different. The N-terminal is the key domain for PGC-1α transcriptional activation. However, some studies have found that the highly conserved CTDs serve as platforms for protein interactions and protein-RNA interactions. They also control the dynamic recruitment of chromatin to form drop-like aggregates and regulate RNA transcription processing in RNA-dependent or independent ways (Monsalve et al., 2000; Pérez-Schindler et al., 2021). However, RRM deletion did not entirely eliminate PGC-1α interaction with RNA (Miller et al., 2019). The C-terminal of PGC-1α contains a conserved region that includes an α helix known as CBP80-binding motif (CBM). This region binds to CBP80 and promotes RNA synthesis (Cho et al., 2018). Recent studies have shown that PGC-1α forms a complex called P4RC with CBP80 through its CBM, which releases RNAPII and enhances transcriptional elongation (Rambout et al., 2023) (Fig. 2b).
Upstream signals that modulate PGC-1α
As a key regulator of energy metabolism, PGC-1α responds to various environmental and physiological stimuli (Fig. 3). Exercise triggers the activation of the Ca2+/calmodulin-dependent protein kinase IV (CaMKIV) and calcineurin A (CnA) pathways at the mRNA level (Fernandez-Marcos and Auwerx, 2011). CnA facilitates PGC-1α transcription by interacting with myocyte enhancer factor 2 (MEF2), whereas CaMKIV activates PGC-1α through cAMP response element (CRE)-binding protein (CREB). Moreover, postexercise activation of p38 mitogen-activated protein kinase (p38 MAPK) further upregulates PGC-1α via MEF2 and activating transcription factor 2 (Fernandez-Marcos and Auwerx, 2011; Qian, 2024). In cold conditions, the β3 adrenergic receptor (β3-AR) detects low temperatures and triggers cAMP and protein kinase A (PKA) activation, leading to enhanced PGC-1α transcription through CREB or p38 MAPK (Fernandez-Marcos and Auwerx, 2011; Weng et al., 2023). In addition, DNA methyltransferase 3B suppresses PGC-1α promoter activity. The methyltransferase like 3 (METTL3), the core catalytic subunit of the m6A methyltransferase complex, binds to PGC-1α mRNA, promoting its degradation and inhibiting translation and protein expression of PGC-1α (Fernandez-Marcos and Auwerx, 2011; Zhang et al., 2021).
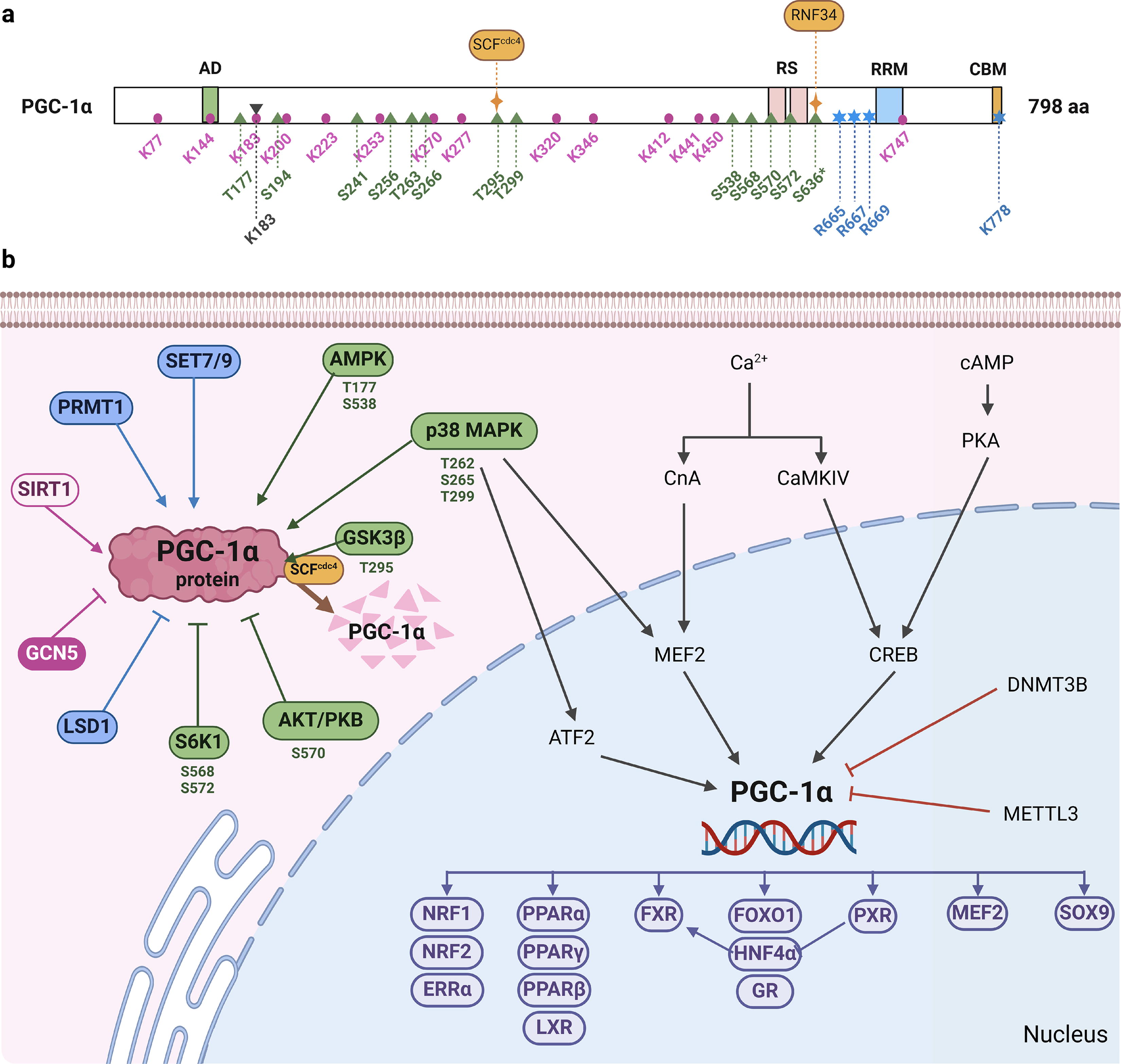
In contrast, PGC-1α activity is primarily regulated by various protein post-translational modifications (PTMs) (Fig. 3). These modifications include phosphorylation, acetylation, methylation, and SUMOylation, among others. Phosphorylation is a well-studied PTM on PGC-1α, with AMP-activated protein kinase (AMPK) phosphorylating T177 and S538 to activate PGC-1α. In contrast, p38 MAPK activates PGC-1α through phosphorylation at T262, S265, and T299 (Fang et al., 2019; Salagre et al., 2024; Suntar et al., 2020). Several natural products such as resveratrol (Yang et al., 2021), fucoxanthin (Winarto et al., 2023), and curcumin (Hann et al., 2013) have been shown to promote mitochondrial biogenesis by enhancing phosphorylation at these sites on PGC-1α. In addition, protein kinase B (PKB/Akt) inhibits S570 phosphorylation of PGC-1α, leading to the activation of forkhead box O (FOXO) 1 and regulation of gluconeogenesis. However, different isoforms of PKB/Akt may not have the same effect on PGC-1α (Qian, 2024). S6 kinase phosphorylates S568 and S572 of PGC-1α, affecting its binding to hepatic nuclear factor-4α (HNF-4α) and negatively impacting gluconeogenesis (Lustig et al., 2011). Moreover, glycogen synthase kinase-3β (GSK3β) phosphorylation of T295 on PGC-1α can trigger Skp1/Cullin/F-box (SCF)CDC4-mediated ubiquitination degradation (Qian, 2024).
As previously mentioned, PGC-1α interacts with transcription factors by recruiting other HAT-containing coactivating proteins. Lerin et al. purified a protein with HAT activity and discovered that GCN5 is an endogenous acetyltransferase of PGC-1α. This acetyltransferase can inhibit PGC-1α’s intrinsic transcriptional activity, leading to the inhibition of gluconeogenesis (Qian, 2024). However, low glucose levels in vivo increase NAD+ levels, which activate sirtuin 1 (SIRT1) and result in PGC-1α deacetylation. Research has demonstrated that the absence or reduction of SIRT1 impedes the stimulatory impact of PGC-1α on various mitochondrial genes or regulatory genes, and also diminishes PGC-1α-induced fatty acid oxidation (FAO) (Li et al., 2011; Wu et al., 2022).
The nuclear receptor coactivator arginine methyltransferase 1 methylates PGC-1α, which interacts with and regulates thermogenic fat activation (Qiao et al., 2019) (Fig. 3). The lysine methylation and demethylation of PGC-1α at the C-terminal can be mediated by the methyltransferase SET7/9 and the lysine-specific demethylase 1A, respectively (Aguilo et al., 2016). In addition, reversible SUMOylation has been shown to modulate the activity of PGC-1α (Rytinki and Palvimo, 2009).
In addition to post-translational modifications, PGC-1α can also receive signals by interacting with signal molecules. For example, the transcriptional repressor protein yin-yang 1-PGC-1α interaction is regulated by the mammalian target of rapamycin, which plays a crucial role in energy metabolism and nutrition (Qian, 2024). Overall, PGC-1α is influenced by various external signals, which in turn affect its function.
The Role of PGC-1α in Cellular Metabolism and Systemic Diseases
Upon receiving various signals, PGC-1α can activate numerous metabolic pathways, coordinate the metabolism of molecular compounds, enhance mitochondrial biogenesis, and regulate cellular energy and ROS homeostasis. Its involvement is evident in the pathogenesis of various diseases, including neurological disorders, cardiovascular diseases, and kidney-related conditions.
PGC-1α is a hub of multiple metabolism pathways
Glucose serves as a vital source of energy and material acquisition for organisms. PGC-1α, a key factor in maintaining energy balance, plays a crucial role in glucose metabolism. Research indicates that fasting triggers the release of glucagon, along with cAMP analogs, which in turn stimulate the elevation of CREB through the PKA pathway, resulting in a notable increase in PGC-1α expression (Tavares et al., 2020). Moreover, PGC-1α forms a complex with HNF-4α or FOXO1 to increase the expression of phosphoenolpyruvate carboxykinase (PEPCK), glucose-6-phosphatase, and other important enzymes in the gluconeogenic pathway, which ultimately leads to the promotion of gluconeogenesis (Qian, 2024). In contrast, insulin-mediated Akt phosphorylation hinders PGC-1α transcription, leading to the opposite effect. Several studies have shown that low levels of PGC-1α in diabetic patients are linked to insulin resistance (Oropeza et al., 2015). Furthermore, PGC-1α aids in the transcription of the Glucose Transporter 4 (GLUT4) gene by interacting with MEF2, thereby facilitating glucose uptake (Qian, 2024). In short, PGC-1α is crucial for maintaining normal glucose homeostasis.
PGC-1α plays a crucial role in lipid metabolism, providing essential energy and substances for the organism. It functions in multiple ways, such as enhancing PPARα-mediated transcriptional activation and interacting with estrogen-related receptor (ERR)α to promote the transcription of medium-chain acyl coenzyme A dehydrogenase (MCAD), which facilitates mitochondrial FAO (Huss et al., 2004; Huss and Kelly, 2004). Furthermore, PGC-1α’s interaction with thyroid hormones stimulates the expression of carnitine palmitoyltransferase1, a key enzyme in FAO (Espinoza et al., 2010). In addition, PGC-1α supports lipid synthesis by promoting the expression of genes like acetyl CoA carboxylase, fatty acid synthase, and diacylglycerol-O-acyltransferase1. These transcriptional changes enable lipid storage to meet future energy demands when needed (Espinoza et al., 2010; Girnun, 2012). Moreover, PGC-1α can also contribute to lipid metabolism through the synergistic activation of liver X receptors (LXR) (Calkin and Tontonoz, 2010). Coactivation of PGC-1α with PPARγ and HNF-4α leads to enhanced transcription of the nuclear receptor farnesoid X receptor (FXR) gene or interaction with the structural DNA structural domain of FXR, resulting in decreased plasma triglyceride levels and hepatic triglyceride synthesis and secretion (Bhalla et al., 2004; Zhang et al., 2004). Therefore, PGC-1α plays a dual regulatory role in lipid metabolism (Espinoza et al., 2010).
PGC-1α interacts with various factors to form a complex metabolic network, including binding to the nuclear hormone receptor family. It binds to PPARβ to prevent ubiquitination and to PPARγ for essential biological functions. The interaction with ERRα influences multiple pathways, regulating PPAR transcriptional activity and FAO enzyme gene activation. Activation of nuclear receptors, such as PPARγ, glucocorticoid receptor, thyroid hormone receptors, and retinoid X receptor, can impact PGC-1α by displacing inhibitors or inducing conformational changes (Knutti et al., 2001).
Overall, PGC-1α serves as a central player in multiple metabolic processes, crucial for maintaining cellular energy balance.
Mitochondrial biogenesis
Mitochondria are widely recognized as the primary sites for cellular energy metabolism, housing numerous biochemical processes and metabolic reactions. Dysfunction in mitochondria has been linked to various diseases, including neurodegenerative diseases, cardiomyopathy, aging, and cancer (Mishra and Chan, 2016). PGC-1α, a key regulator of cellular energy metabolism homeostasis, plays a central role in mitochondrial biogenesis and is involved in physiological processes such as adaptive thermogenesis triggered by cold exposure (Puigserver and Spiegelman, 2003) and muscle fiber transition regulation (Mortensen et al., 2006). In addition, PGC-1α has been implicated in pathological processes. For instance, studies by Lehman et al. and Russell et al. have demonstrated that increased PGC-1α expression in the hearts of newborn mice leads to mitochondrial proliferation and subsequent development of cardiomyopathy (Lehman et al., 2000; Russell et al., 2004). PGC-1α has also been linked to conditions like obesity (Martínez-Jiménez et al., 2019), neurodegenerative diseases (Calabrese et al., 2010; Wang et al., 2013), and Duchenne muscular dystrophy (Suntar et al., 2020).
Furthermore, PGC-1α stimulates the expression of nuclear transcription factors such as nuclear respiratory factor (NRF)−1, NRF-2α, and ERRα and interacts with specific nuclear transcription factors like NRF-1. NRF-1 binds to the promoter regions of various mitochondrial genes in the nucleus, including β-ATP synthase and cytochrome-c, and mediates the activation of transcription factors that promote mitochondrial DNA replication and transcription, such as mitochondrial transcription factor (mtTF)-A and mtTFB. This stimulation leads to the activation of the respiratory chain and mitochondrial transcription (Vernier and Giguère, 2021; Zhao et al., 2021).
Mitochondria, which possess their own genetic system (mtDNA) and are the only organelles outside the nucleus to contain DNA, are crucial for cellular function. PGC-1α acts as a coactivator and interacts with various transcription factors related to mitochondrial oxidation. It integrates the effects of multiple transcription factors to regulate mitochondrial function. The upregulation of PGC-1α expression, which promotes mitochondrial biogenesis and enhances the oxidative capacity of skeletal muscle cells, can be achieved through a CREB-dependent pathway by either the transducer of regulated CREB-binding proteins (TORCs) or CaMK (Wu et al., 2006). This process makes the cells more adaptable to endurance exercise (Wu et al., 2002). There’s no doubt that the impact of PGC-1α on mitochondria is extensive and significant.
According to previous studies, mitochondrial biogenesis varies depending on the tissue. In addition, the interaction between PGC-1α and transcription factors can lead to coupled or uncoupled respiration in a tissue- and cell-type specific manner. For example, exposure to cold temperatures can cause the upregulation of PGC-1α and uncoupled mitochondrial respiration through activation of uncoupling protein (UCP) expression in brown adipose and skeletal muscle tissues. In contrast, forced expression of PGC-1α in cultured cardiomyocytes leads to coupled respiration (Lehman et al., 2000; Leone et al., 2005). This difference may explain why cardiac tissue, which has high levels of PGC-1, is not as important in adaptive thermogenesis.
PGC-1α and redox homeostasis
Mitochondria are indeed the energy powerhouses of cells, but as the proverb goes, “No rose without a thorn.” When ATP is produced through the electron transport chain, a fraction of oxygen is incompletely reduced, generating ROS (Bhatti et al., 2017). ROS primarily originate from mitochondria in cells, serving as signaling molecules to regulate functions like inflammatory responses. However, excessive ROS production can cause oxidative damage to mitochondrial components, leading to dysfunction and disease development (Calabrese et al., 2016). Cells combat oxidative stress induced by ROS by generating various antioxidant enzymes and low-molecular weight scavengers (Liu et al., 2023a).
While promoting mitochondrial biogenesis, PGC-1α also plays a crucial role in regulating the expression of ROS-detoxifying enzymes such as superoxide dismutase, glutathione peroxidase, and UCP. PGC-1α triggers mitochondrial remodeling, which helps maintain cellular redox homeostasis at a lower level (Fig. 4a). During times of stress, there is an excess production of ROS leading to an imbalance in cellular redox levels. Simultaneously, the upregulation of PGC-1α in most cells acts as a protective mechanism by enhancing the expression of ROS detoxification proteins (Suntar et al., 2020). In addition, the overexpression of PGC-1α can mitigate ROS levels by inhibiting dynamin-related protein 1–mediated mitochondrial dynamics (Guo et al., 2015).

Oxidative stress, triggered by an overproduction of ROS, is closely linked to various diseases, such as acute kidney injury, aging, and neurodegenerative diseases (Fig. 4b). PGC-1α has become a key target for improving the clinical outcomes of diseases associated with oxidative stress (Jamwal et al., 2021; Zhao et al., 2021). For instance, resveratrol demonstrates its antioxidant properties through the activation of PGC-1α (Yang et al., 2021).
Systemic disease
PGC-1α is known to have implications in various systemic diseases. In the nervous system, it plays a role in neuronal survival and synaptic transmission. Increased levels of PGC-1α have been shown to combat neuronal damage progression, offering potential therapeutic avenues for neurodegenerative conditions like Alzheimer’s, Huntington’s, and Parkinson’s diseases (Nierenberg et al., 2018; Sack, 2010; Yang et al., 2021). In addition, PGC-1α has a protective impact on the cardiovascular system, with dysregulated expression potentially leading to cardiovascular issues such as postnatal cardiomyopathy, cardiac insufficiency, myocardial hypertrophy, and atherosclerosis (Haemmerle et al., 2011; Persad and Lopaschuk, 2022; Schilling and Kelly, 2011). Furthermore, PGC-1α has been observed to aid in recovery from ischemia-reperfusion injury in acute kidney injury and may also play a role in preventing diabetic nephropathy (Chambers and Wingert, 2020).
PGC-1α is a Double-Edged Sword for Tumors
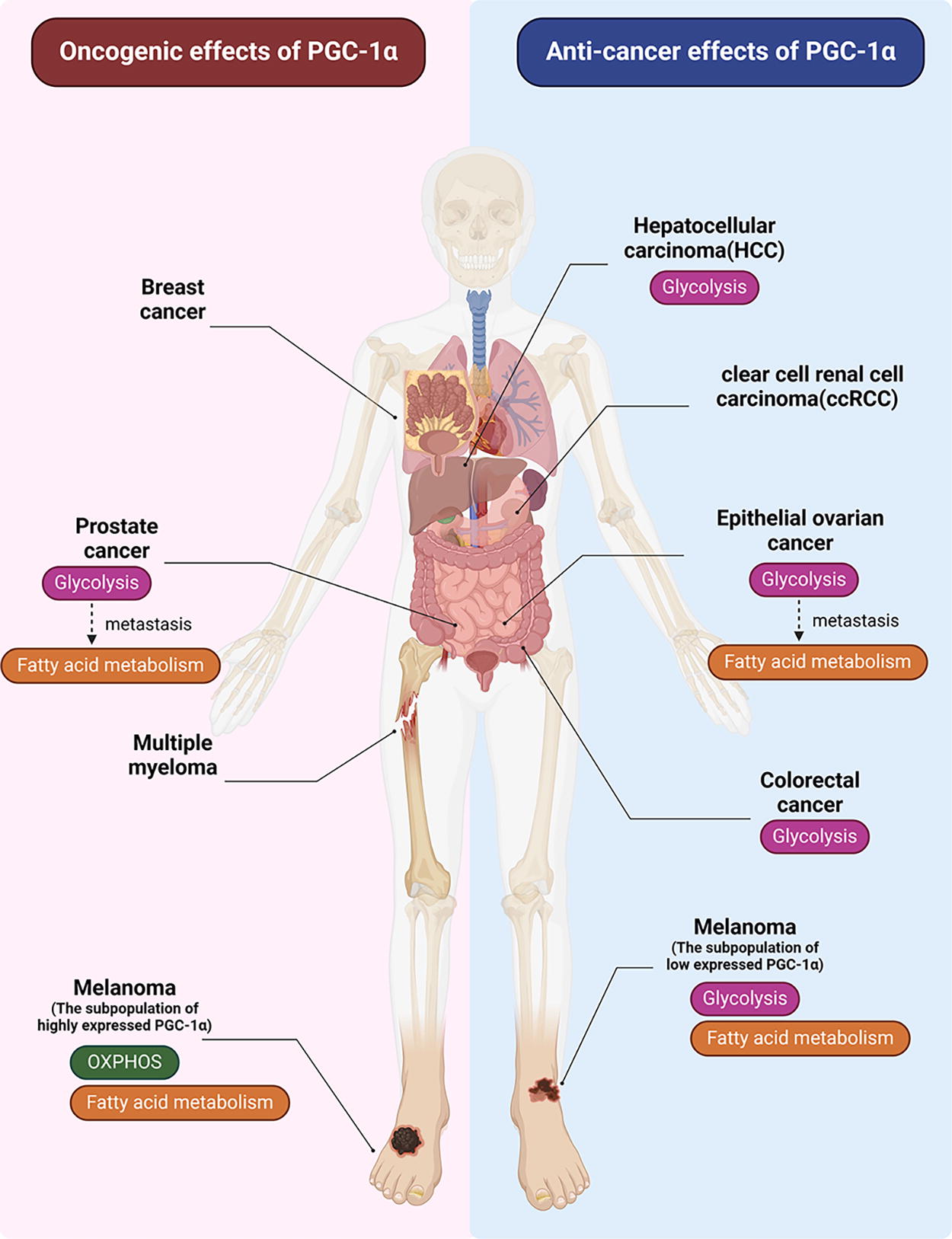
Current evidence suggests that PGC-1α plays a significant role in tumorigenesis, development, metastasis, and treatment resistance by modulating transcriptional programs and metabolic pathways (Mastropasqua et al., 2018). However, the effects of PGC-1α vary across different types of cancer (Fig. 5). For example, PGC-1α can promote cancer in breast cancer, but has an inhibitory effect in ovarian cancer. In addition, PGC-1α may have opposing roles in different subtypes or stages of the same tumor, such as in prostate cancer. Given the diverse functions of PGC-1α that are highly dependent on specific tumor types, a comprehensive understanding of its role in tumors is essential.
Oncogenic effects of PGC-1α
PGC-1α has been found to be upregulated in gastric cancer (Wang et al., 2019), endometrial cancer (Cormio et al., 2009), and certain renal tumors (Klomp et al., 2010) (Table 1), indicating its potential role as a cancer-promoting molecule. Notably, the expression level of PGC-1α has been inversely associated with the prognosis of breast cancer (Audet-Walsh et al., 2016).
Expression of PGC-1α in Different Cancer Species and Its Roles in Tumor Initiation and Progression
PGC-1α acts as a transcription coactivator, interacting with the androgen receptor (AR) to promote tumor cell proliferation via the AMPK pathway in prostate cancer. Studies have shown that knockout of PGC-1α effectively inhibits tumor cell growth (Shiota et al., 2011; Tennakoon et al., 2014). Furthermore, PGC-1α has been implicated in the growth of breast tumors (Klimcakova et al., 2012). The orphan nuclear receptor ERRα is known to synergize with PGC-1α in regulating cellular energy homeostasis and oxidative metabolism (Deblois et al., 2009; Charest-Marcotte et al., 2010).
The tricarboxylic acid cycle is essential for providing intermediates necessary for the synthesis of amino acids and fatty acids, crucial for the survival and growth of cancer cells (DeBerardinis et al., 2008; Luo et al., 2016; Menendez and Lupu, 2007). The ERRα/PGC-1α pathway enhances glutamine pathway–mediated fatty acid synthesis in low glucose and hypoxia conditions, vital for lipid synthesis in cancer (DeBerardinis and Cheng, 2010; Luo et al., 2016; McGuirk et al., 2013). PGC-1α increases the expression of enzymes related to lipid synthesis and regulates genes involved in converting glucose to fatty acids when energy is sufficient (Bhalla et al., 2011; Espinoza et al., 2010). In addition, PGC-1α promotes FAO when coactivated with other transcription factors, contributing to a favorable tumor environment for cancer growth.
Tumors depend on developing a strong blood supply to maintain nutrient levels and facilitate the spread of cancer cells. PGC-1α has been shown to stimulate the growth of new blood vessels in different types of tumors, including breast cancer and multiple myeloma, without needing hypoxia-inducible factor-1 alpha ( Chinsomboon et al., 2009 ; Arany et al., 2008; Cao et al., 2014; Klimcakova et al., 2012), possibly related to ERRα (Girnun, 2012).
Solid cancer cells have the ability to easily spread to other organs through the bloodstream, although only a small percentage of these cells are actually able to successfully metastasize. Stephen Paget, a British surgeon, introduced the “seed and soil” hypothesis in 1889 based on autopsy studies of patients with primary tumors. This hypothesis, which explains the nonrandom nature of tumor metastasis, posits that the outcome of metastasis depends on the affinity of certain tumor cells for specific organ microenvironments (Fidler, 2003). Recent research on tumor metastasis indicates the presence of distinct subpopulations of cells within primary tumors, which may originate from the multicellular nature of the tumor or from changes during its progression (Fidler, 2003; Hoek et al., 2008). LeBleu et al. conducted studies isolating circulating cancer cells (CCCs), primary cancer cells (PCCs), and metastatic cancer cells (MCCs) and found significant differences in gene expression related to the oxidative phosphorylation pathway, particularly in CCCs compared to PCCs. They observed upregulation of genes associated with mitochondrial biosynthesis in CCCs, with some genes showing reversible expression patterns in MCCs that partially reverted to levels seen in PCCs. This suggests a potential role for PGC-1α as an upstream factor in mediating the transition from PCCs to CCCs and then to MCCs (LeBleu et al., 2014). Studies have shown a positive correlation between PGC-1α and tumor metastasis in cholangiocarcinoma and invasive breast cancer (Andrzejewski et al., 2017; Dan Li et al., 2018).
PGC-1α has been shown to be upregulated in drug-resistant cells, including 5-fluorouracil (5-FU)-resistant colorectal cancer cells (Yun et al., 2019), cisplatin-resistant ovarian cancer cells (Deng et al., 2020), and adriamycin–resistant diffuse large B cell lymphoma (Zhou et al., 2020). Silencing PGC-1α in nonsmall cell lung cancer has been associated with a significant reduction in resistance to cisplatin (Cruz-Bermúdez et al., 2019). A recent study revealed that inhibiting PGC-1α in melanoma stem cell subsets can impede the growth and propagation of melanosphere, decrease tumorigenicity, and reduce invasiveness in melanoma, offering new potential targets for combating treatment resistance and cancer recurrence (Fontana et al., 2024).
Tumor suppressive effects of PGC-1α
On the contrary, PGC-1α also functions as a tumor suppressor in various types of cancer. It is typically downregulated in solid tumors such as lung cancer (Bellance et al., 2009), hepatocellular carcinoma (HCC) (Liu et al., 2017; Piccinin et al., 2019), bile duct cancer (Li et al., 2018), colorectal cancer (Feilchenfeldt et al., 2004; Triki et al., 2017), ovarian cancer (Zhang et al., 2007), and thyroid cancer (Liu et al., 2017) (Table 1). In addition, PGC-1α has been found to have a positive correlation with the clinical prognosis of HCC (Zuo et al., 2021).
PGC-1α is known to be highly expressed in normal intestinal epithelial cells. In the context of colorectal cancer, studies have shown that PGC-1α inhibits colorectal carcinogenesis by inducing mitochondria-related metabolic changes in intestinal epithelial cells (D’Errico et al., 2011; Triki et al., 2017). In addition, PGC-1α has been found to inhibit metastasis and invasion of HCC both in vivo and in vitro by suppressing aerobic glycolysis (Zuo et al., 2021).
The production of ROS plays a dual role in tumor cells, contributing to tumorigenic alterations while also exerting toxic effects. Overexpression of PGC-1α has been shown to increase ROS levels, leading to apoptosis in proliferating colorectal cancer cells and ultimately protecting against tumorigenesis (D’Errico et al., 2011; Triki et al., 2017; De Souza-Teixeira et al., 2018). Furthermore, PGC-1α induces apoptosis in human epithelial ovarian cancer (Zhang et al., 2007) and suppresses tumorigenesis in clear cell renal cell carcinoma (ccRCC), with low PGC-1α expression correlating with poor overall survival in ccRCC patients (LaGory et al., 2015).
Pancreatic cancer stem cells with reduced PGC-1α expression display enhanced glycolysis and develop resistance to metformin treatment (Bost and Kaminski, 2019; Sancho et al., 2015). Metformin, widely used in cancer therapy in addition to its antidiabetic properties, is speculated to be more effective against pancreatic cancer when PGC-1α levels are increased (Huang et al., 2023; Triggle et al., 2022).
Pleiotropic regulation of tumors by PGC-1α
The expression of PGC-1α varies among different subtypes of the same cancer and has different effects on tumor cells. For example, high PGC-1α expression in a subpopulation of melanoma was linked to poor survival outcomes compared with a subpopulation with low PGC-1α expression. The subpopulation with low PGC-1α levels showed increased glycolysis and lactic acid production, whereas the subpopulation with high PGC-1α levels had higher rates of mitochondrial oxidative metabolism (Haq et al., 2013; Vazquez et al., 2013). In addition, PGC-1α acts as an inhibitor of tumor growth and metastasis in cancers that rely on glycolysis, such as HCC, ovarian cancer, and colorectal cancer (Chakraborty et al., 2017; Wang et al., 2023; Zhao et al., 2022; Zhao and Li, 2021). The role of PGC-1α also varies at different stages of tumor development, with tumors gradually shifting from glycolysis to fatty acid metabolism (Yoon and Lee, 2022; Zhang et al., 2023). PGC-1α inhibits tumor metastasis by suppressing fatty acid metabolism. Melanoma cells exhibit high heterogeneity, leading to variations in their metastatic potential (Zhang et al., 2017). Some cell subsets with high PGC-1α levels depend on it for survival and tumor progression (Luo et al., 2016). These findings suggest that PGC-1α plays distinct or even opposing roles in different types and subgroups of tumors or at different tumor stages, possibly related to the metabolic pathways utilized by the tumors at that time (Fig. 5).
Overall, PGC-1α acts as a key regulator of multiple metabolic pathways, enabling tumor cells to achieve energy balance. The diversity of PGC-1α expression in different types and subtypes of tumors, along with its dynamic changes during tumor development, progression, and metastasis, highlights the complexity and heterogeneity of cancer.
Application of Targeting PGC-1α in Tumors
Recent studies highlight the pivotal and intricate role of PGC-1α in cancer pathogenesis. Various therapeutic approaches aimed at targeting PGC-1α show promise in both cancer prevention and treatment.
Natural compounds target PGC-1α against tumors
Natural compounds sourced from diverse natural reservoirs have garnered attention for their therapeutic potential in treating a range of diseases with minimal toxicity. For instance, curcumin (Calabrese et al., 2007; Concetta Scuto et al., 2019), resveratrol (Yang et al., 2021), and saponins (Zhang et al., 2023) have demonstrated beneficial effects. Curcumin hinders the proliferation of head and neck cancer cells by downregulating the PGE2 receptor EP4 gene through the activation of AMPKα and p38 MAPK pathways mediated by PGC-1α (Hann et al., 2013). Resveratrol induces mitochondrial dysfunction and reduces PGC-1α levels, leading to increased ROS, elevated Deleted in liver cancer-1 (DLC1) expression, and subsequent cancer cell senescence (Ji et al., 2018). Similarly, momordin Ic, a natural triterpenoid saponin, triggers apoptosis in HCC cells by enhancing PGC-1α and FOXO4 expression via the MAPK signaling pathway (Wang et al., 2014). In addition, ginsenoside Rh2 promotes mitophagy by reducing PGC-1α levels, thereby influencing mammary epithelial cell aging and breast cancer growth (Hou et al., 2020; Jeon et al., 2021). Collectively, targeting PGC-1α with natural compounds emerges as a promising avenue in antitumor therapy.
Target modulation of PGC-1α
Metformin, an FDA-approved antidiabetic agent used to treat type 2 diabetes mellitus, has been shown to significantly increase PGC-1a expression (Aatsinki et al., 2014). There is compelling evidence supporting the potential use of metformin in cancer prevention and treatment. Metformin has been demonstrated to reduce cancer incidence, mortality, enhance response to radiotherapy and chemotherapy, and decrease the risk of cancer recurrence (Saraei et al., 2019). Moreover, studies have revealed that downregulation of PGC-1α in metformin-resistant pancreatic cancer stem cells could provide insight into mechanisms of tumor resistance, although further research is needed to fully understand this mechanism (Sancho et al., 2015).
Radiotherapy is a key component in the treatment of glioma, but its effectiveness is often limited by the development of radiotherapy resistance. Glioma cells with decreased PGC-1α expression have been found to display increased resistance to radiotherapy. In both in vitro and in vivo studies, ZLN005, a PGC-1α activator, has shown the ability to enhance the radiosensitivity of resistant glioma cells by reactivating mitochondria-related ROS production and inducing apoptotic effects (Zhao et al., 2023).
Gene therapy targeting PGC-1α
There is a growing body of evidence supporting the potential of gene therapy as a therapeutic approach in cancer treatment. Numerous in vivo and in vitro studies have been conducted to investigate the impact of editing (knocking out or overexpressing) PGC-1α on tumor growth, invasion, or drug resistance in cells or animals. For instance, research has shown that overexpression of PGC-1α in breast cancer cells can enhance apoptosis and inhibit cell proliferation (Zu et al., 2021). In addition, experiments have demonstrated that overexpressing PGC-1α can restrict the migration and invasion of HCC cells in both in vitro and in vivo settings (Zuo et al., 2021).
Anticancer immunotherapy by orchestrating PGC-1α
Investigating the mechanisms of tumor immunosuppression and developing new approaches for sensitizing immunotherapy are crucial for enhancing the prognosis and survival rates of cancer patients, representing a cutting-edge focus in global oncology research. CD8+ T cells are pivotal in antitumor immunity, with mitochondrial dysfunction directly impairing their antitumor effects (Gebhardt et al., 2023). Abnormal mitochondrial dynamics in tumor-infiltrating CD8+ T cells are often linked to the downregulation or inhibition of PGC-1α (Zhang et al., 2022). Boosting PGC-1α expression enhances T cell activation, thereby bolstering CD8+ T cell–mediated antitumor immune responses (Dumauthioz et al., 2021; Zhong et al., 2023). In addition, activating PGC-1α promotes FAO in tumor-infiltrating lymphocytes (TILs), augmenting the efficacy of programmed cell death protein-1 (PD-1) blockers in lung cancer (Wan et al., 2020). Moreover, PD-1 impedes mitochondrial biogenesis by intensifying the inhibition of AKT-mediated PGC-1α (Ogando et al., 2019). Adoptive cell therapy has shown that pharmacological AKT inhibition enhances antitumor immunity in CD8+ T cells (Crompton et al., 2015), potentially through mechanisms involving PGC-1α. Therefore, PGC-1α emerges as a promising therapeutic target in antitumor immunity.
Conclusion and Prospective
Compelling evidence has revealed that PGC-1α plays a unique role in directing cancer metabolism, although the specific regulatory mechanism of PGC-1α in cancer remains incompletely understood. This review summarizes the structure and upstream and downstream signals of PGC-1α, highlighting its crucial role in metabolism and redox homeostasis, particularly in tumor tumorigenesis and cancer progression. The potential of PGC-1α as a promising target for cancer prevention and treatment is discussed, emphasizing the importance of considering the metabolic profile of the cancer when deciding on activation or inhibition of PGC-1α.
PGC-1α influences cellular metabolism by interacting with various transcription factors, such as nuclear factor erythroid 2-related factor 2 (Nrf2), a key regulator of cellular defense against oxidative stress (Aquilano et al., 2013; Deng et al., 2020). When faced with increased ROS levels, PGC-1α and Nrf2 work together to enhance the expression of antioxidant genes, although the exact mechanism is not fully understood. Given the dual roles of ROS in cancer prevention and therapy (Wang et al., 2021), the complex interplay between PGC-1α, Nrf2, and ROS may contribute to the diverse functions of PGC-1α in different stages of tumor development. Further research is needed to elucidate the mechanisms underlying the cross talk between PGC-1α and Nrf2.
In the tumor microenvironment (TME), TILs often experience severe metabolic defects, leading to functional deficits. Studies have shown that overexpressing PGC-1α in CD8+ T cells can enhance mitochondrial activity, fitness, and memory formation and boost antitumor immunity in a melanoma model (Dumauthioz et al., 2021; Zhong et al., 2023). These findings suggest that targeting PGC-1α could potentially correct TIL metabolism and enhance antitumor immunity.
PGC-1α can have varying and even opposing effects in different types and subgroups of tumors or at different stages of tumor development, potentially due to the metabolic pathways utilized by the tumors at that particular time. For example, PGC-1α often acts as a tumor suppressor in certain cancers that primarily rely on glycolysis, while it can promote tumor growth in OXPHOS-dependent tumors. The interplay between PGC-1α and various transcription factors in the TME is complex and warrants further investigation. The metabolic preferences of different tumors may be influenced by interactions in diverse environments, resulting in varying outcomes. Conducting in-depth studies on tumor metabolism to explore the diverse effects of PGC-1α on tumors could be a promising approach.
The increasing evidence suggests that targeting PGC-1α could be a promising but challenging approach for anticancer therapy. Therefore, developing personalized treatments based on the specific metabolic characteristics of each cancer type may be a more practical strategy.
Footnotes
Authors’ Contribution
L.Y. conceived the review, Z.Y. and Z.H. wrote the article and prepared the figures and tables. All authors contributed to the discussion and revision and the final version.
Author Disclosure Statement
The authors have no relevant financial or nonfinancial interests to disclose.
Funding Information
This work was supported by the Major International (Regional) Joint Research Program of the NSFC of China (No. 81920108027), NSFC (No. 82273212), Funding for National Clinical Key Specialty (Oncology), and Funding for Chongqing Young and Middle-Aged Medical Excellence Team.