
Abstract
Aims:
Parietal epithelial cells (PECs) are potential stem cells within the glomerulus, migrating into site of podocyte loss to differentiate into podocytes. Little is known about the mechanism mediating differentiation of PECs into podocytes.
Results:
In vitro differentiation of PECs into podocytes led to upregulation of podocyte markers such as Wilms’ tumor gene 1 (WT-1), Forkhead box C1 (FOXC1), synaptopodin and podocin, accompanied by increased mitochondrial abundance. Preincubation with a mitochondrial reactive oxygen species (ROS) inhibitor prevented all these events in PECs. In vivo, adriamycin (ADR)-treated mice exhibited albuminuria, decreased WT1 positive cells, and claudin-1 expressed in glomerular capillary tuft, as well as peroxisome proliferator-activated receptor-γ coactivator-1α (PGC1α) overproduction in PECs. Expression of the ROS-related molecule nuclear factor erythroid 2-related factor 2 (Nrf2) and its target protein Brahma-related gene 1 (Brg1) increased during differentiation of PECs into podocytes. Suppressing Nrf2 or Brg1 reduced the differentiation of PECs, whereas overexpression had the opposite effect. Brg1 directly regulated WT-1 transcription in PECs. Activation of Nrf2 with bardoxolone-methyl (CDDO-Me) resulted in less proteinuria and more WT1 positive cells in ADR mice. PECs conditional human Nrf2 knock-in mice showed increased WT1 cell numbers.
Conclusion:
It concluded that mitochondria-derived ROS mediated differentiation of PECs into podocytes via Nrf2 and Brg1 signaling. Antioxid. Redox Signal. 42, 393–407.
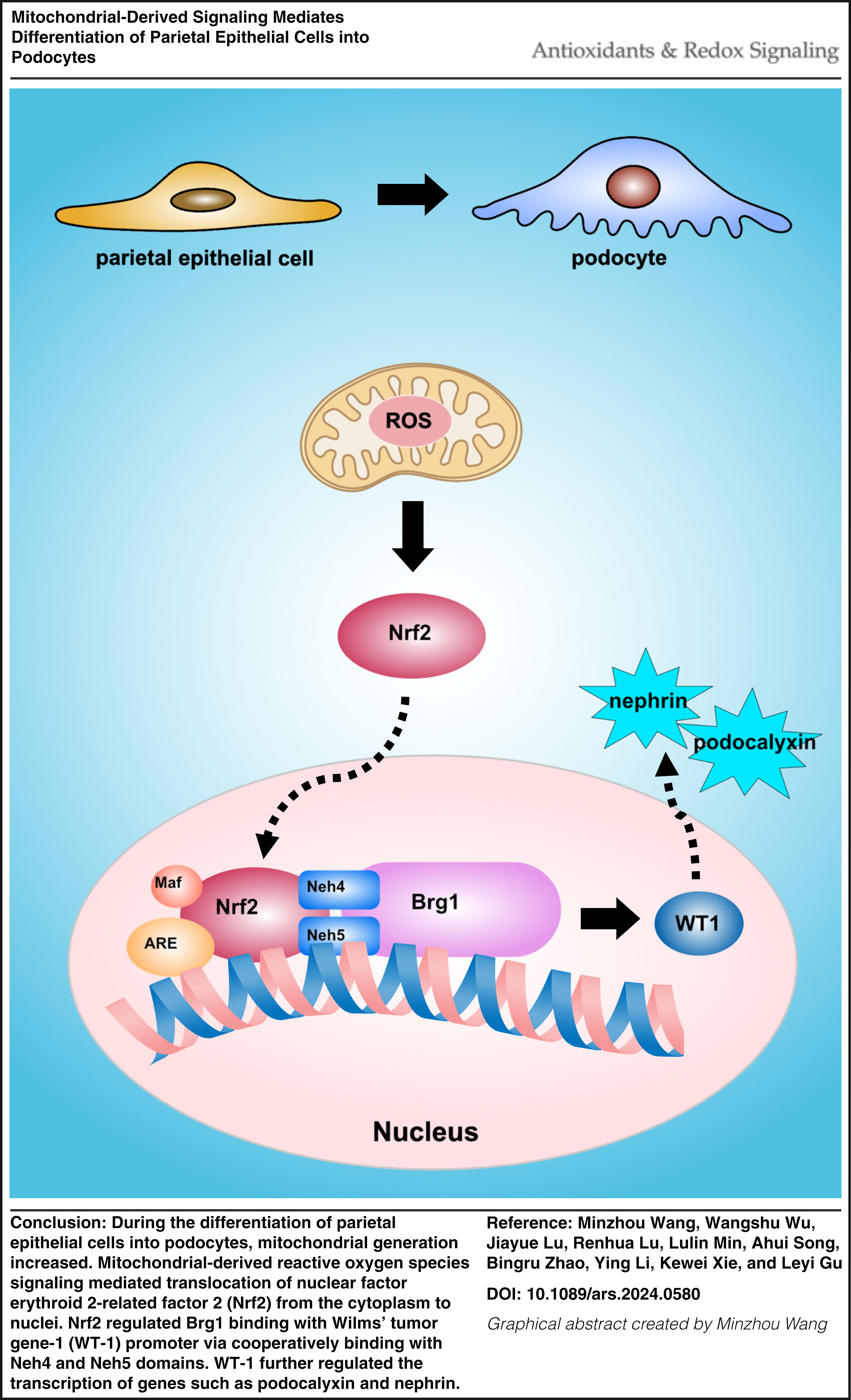
Innovation
Glomerular parietal epithelial cells (PECs) are potential progenitor cells. Previous studies found that PECs differentiate into podocytes compensating for lost podocytes. In this study, we found mitochondria generation increasing combined with mitochondrial-derived reactive oxygen species (mtROS); Nrf2 and Brg1 increased during differentiation of PECs into podocytes in vitro. PECs differentiated into podocytes dependent on mtROS-Nrf2-Brg1. Brg1 regulated WT-1 transcription directly in PECs. In vivo, bardoxolone-methyl, an Nrf2 agonist, and PECs conditional overexpression of human Nrf2 increased WT1-positive cells in glomeruli of adriamycin nephropathy mice.
Introduction
Podocytes are one of the most important cells to maintain the barrier for glomerular filtration (Shankland, 2006). However, podocytes are terminally differentiated cells that usually have only a very limited capacity to regenerate, which hinders repair after podocyte injury.
Glomerular parietal epithelial cells (PECs) are potential progenitor cells within the glomerulus (Eng et al., 2015; Ronconi et al., 2009), capable of migrating into the site of podocytes loss to express podocyte-specific marker protein, as well as differentiating into podocytes (Ohse et al., 2010; Zhang et al., 2012a). Several molecules may be involved in PECs differentiation into podocytes. Meyer-Schwesinger et al. found that microRNA-193a (miR-193a) downregulation switches on the podocyte phenotype in PECs (Kietzmann et al., 2015). Growth inhibitor-specific gene 1 (Gas1) deficiency in PECs caused by early diabetes injury promoted the proliferation and differentiation of PECs into podocytes (Luna-Antonio et al., 2017). Activation of Notch signaling promotes PEC proliferation, and downregulation of the Notch signaling was required for the differentiation of PECs to podocytes in vitro (Lasagni et al., 2010). In addition, glucocorticoids (Zhang et al., 2013), angiotensin-converting enzyme inhibitors (Zhang et al., 2015), insulin growth factor 1 (Tan et al., 2016), and all-trans retinoic acid (ATRA) (Suzuki et al., 2003) can also promote the differentiation of PECs to podocytes. However, the roles of PEC organelles and the detailed mechanisms have not been completely investigated.
Imasawa et al. highlighted that during podocyte differentiation in vitro, mitochondrial size, oxygen consumption, mitochondrial DNA number, as well as mitochondria-related protein peroxisome proliferator-activated receptor-γ coactivator-1α (PGC1α), increased, suggesting a role for mitochondria in this process (Imasawa et al., 2017).
In the present study, we evaluated the roles of mitochondria generation and mitochondria-derived signaling molecule production during PEC differentiation into podocytes. Mice with overexpression human nuclear factor erythroid 2-related factor 2 (Nrf2) were developed to detect the podocyte density. We also estimated the effects by activation of Nrf2 signal in adriamycin (ADR)-induced nephropathy mice.
Results
Mitochondria generation increased during PEC differentiation into podocytes
Primary cultured PECs were incubated with differentiating medium (VRADM: ATRA [10 nM], calcitriol [100 nM], and dexamethasone [50 nM]) for 1 week, the podocyte markers WT-1, synaptopodin, podocin and FOXC1 increased and claudin-1 decreased (Fig. 1A, 1B). In addition, the abundance of mitochondria and the expression of mitochondrial DNA (mtDNA) significantly increased in differentiating medium incubated PECs (Fig. 1C, 1D, 1E, 1F, 1G). Figure 1H showed that PGC1α, cytochrome oxidase subunit IV (COX IV), translocase of the outer mitochondrial membrane member 20 (TOMM20), and cytochrome c (CytC) production increased when PECs differentiated into podocytes. Furthermore, small-interfering RNA (siRNA) was used to silence PGC1α in PECs (Supplementary Figure S3A). Following the induction of differentiation in PGC1α-knockdown PECs, the protein expression of podocyte markers WT-1 and synaptopodin was reduced significantly (Fig. 1J, 1K).
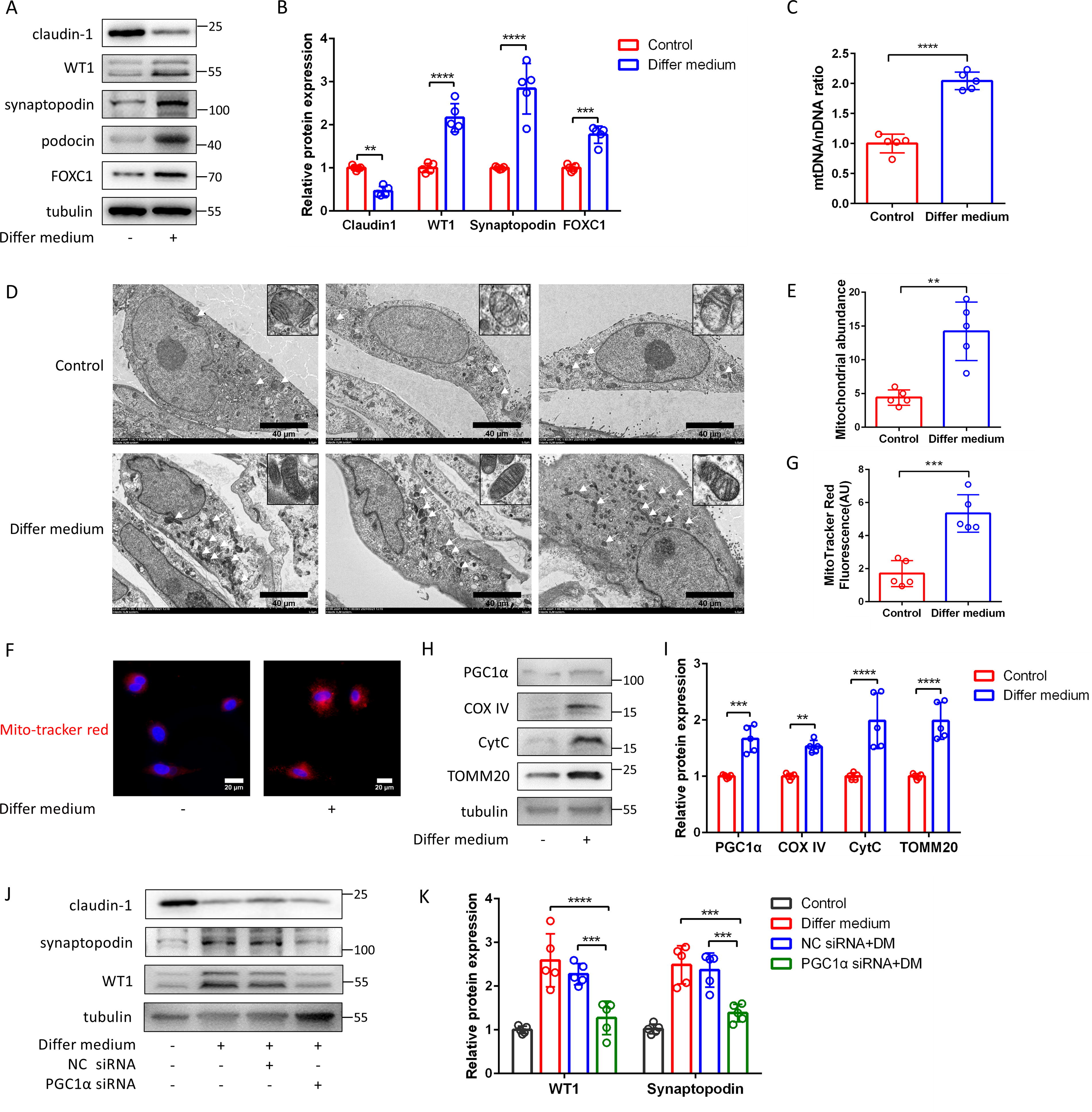
Inducing PECs differentiated into podocytes is accompanied by increased generation in mitochondria.
To verify the differentiation of PECs to podocytes in vivo, we established ADR nephropathy mouse model. 2 and 6 weeks post-ADR administration, mice developed middle-molecular-weight proteinuria, foot process effacement, and focal segmental glomerulosclerosis, accompanied by a reduction in podocyte levels (Supplementary Figure S2A-S2D) and an increased urinary albumin-to-creatinine ratio (UACR) at both time points (Fig. 2A). Podocyte markers (WT-1, nephrin, podocin, and synaptopodin) showed a decline starting from 2 weeks after ADR injection (Fig. 2B, 2C). Increased proliferating cell nuclear antigen (PCNA) production in some PECs and a reduction of WT-1 positive cells were observed in mice with ADR nephropathy (Fig. 2D, 2E). Notably, some claudin-1 positive cells were seen migrating into glomerular capillary tuft (Fig. 2D white arrows), and there was an overproduction of PGC1α (Fig. 2D yellow arrows). Immunoelectron transmission microscopy revealed claudin-1-positive gold particles at the tight junctions of PECs and within the cytosol of some podocytes (Fig. 2F black arrows).
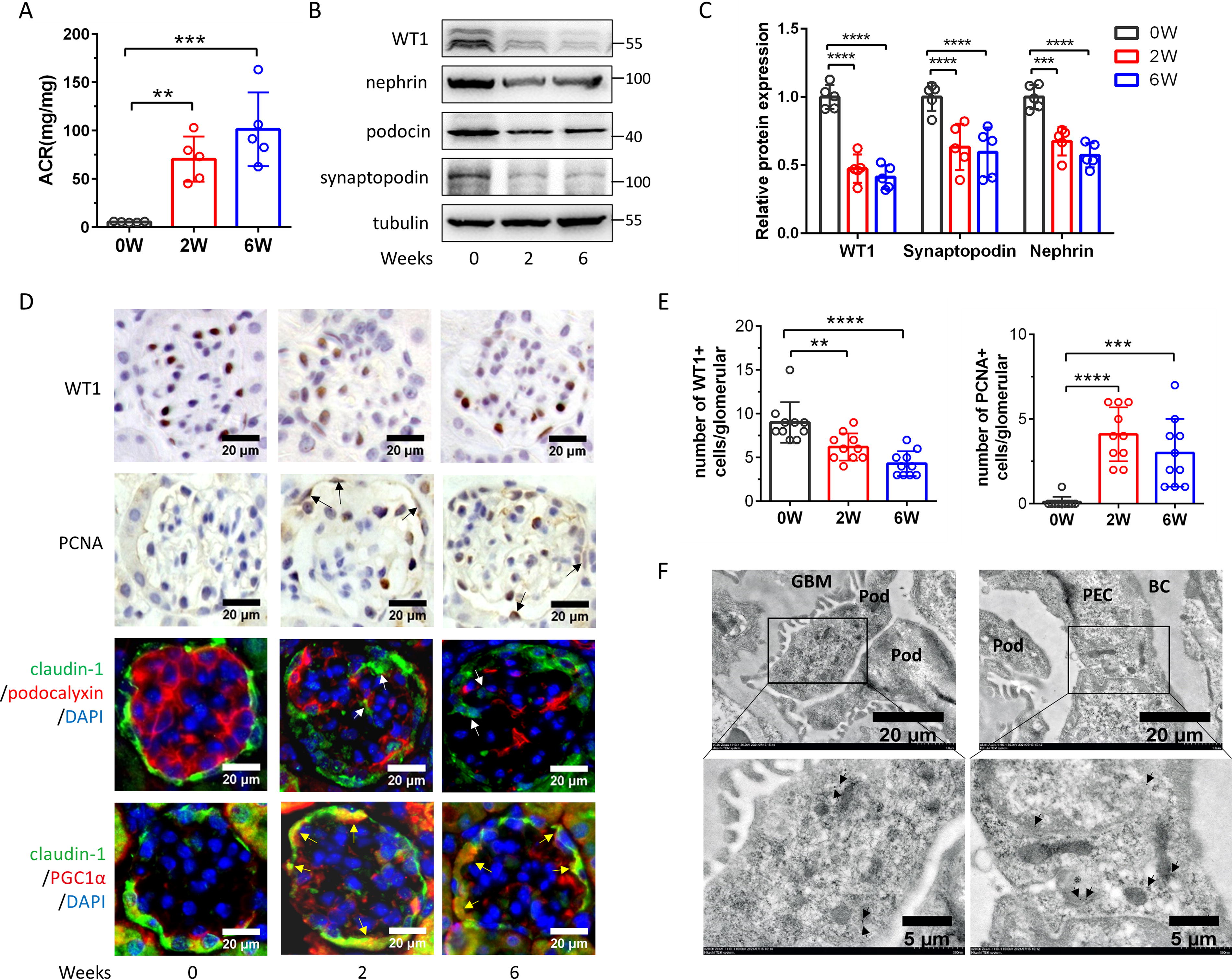
PECs differentiated into podocytes in adriamycin-induced nephropathy.
Mitochondria-derived reactive oxygen species is a key signal for the differentiation of PECs into podocytes
The role of mitochondrial reactive oxygen species (ROS) in the differentiation of PECs into podocytes was investigated by incubating PECs with the VRADM differentiating medium. Relative to the control group, there was a significant increase in both total ROS levels and mitochondrial ROS levels in PECs treated with VRADM. Preincubation with N-acetyl-L-cysteine (NAC, an ROS scavenger) or MitoTempo (a mitochondria-targeted antioxidant) effectively prevented this increase (Fig. 3A, 3B). Following the induction of differentiation of PECs, 3-nitrotyrosine (3-NT) increased and superoxide dismutase (SOD) decreased, suggesting that the differentiation of PECs to podocytes was accompanied by an increase in ROS (Fig. 3C, 3D). The preincubation with MitoTempo and NAC completely inhibited differentiation medium (DM)-induced upregulation of WT1 and synaptopodin (Fig. 3E and 3F). These data implicated mitochondrial ROS as crucial signaling molecules involved in the differentiation of PECs into podocytes.
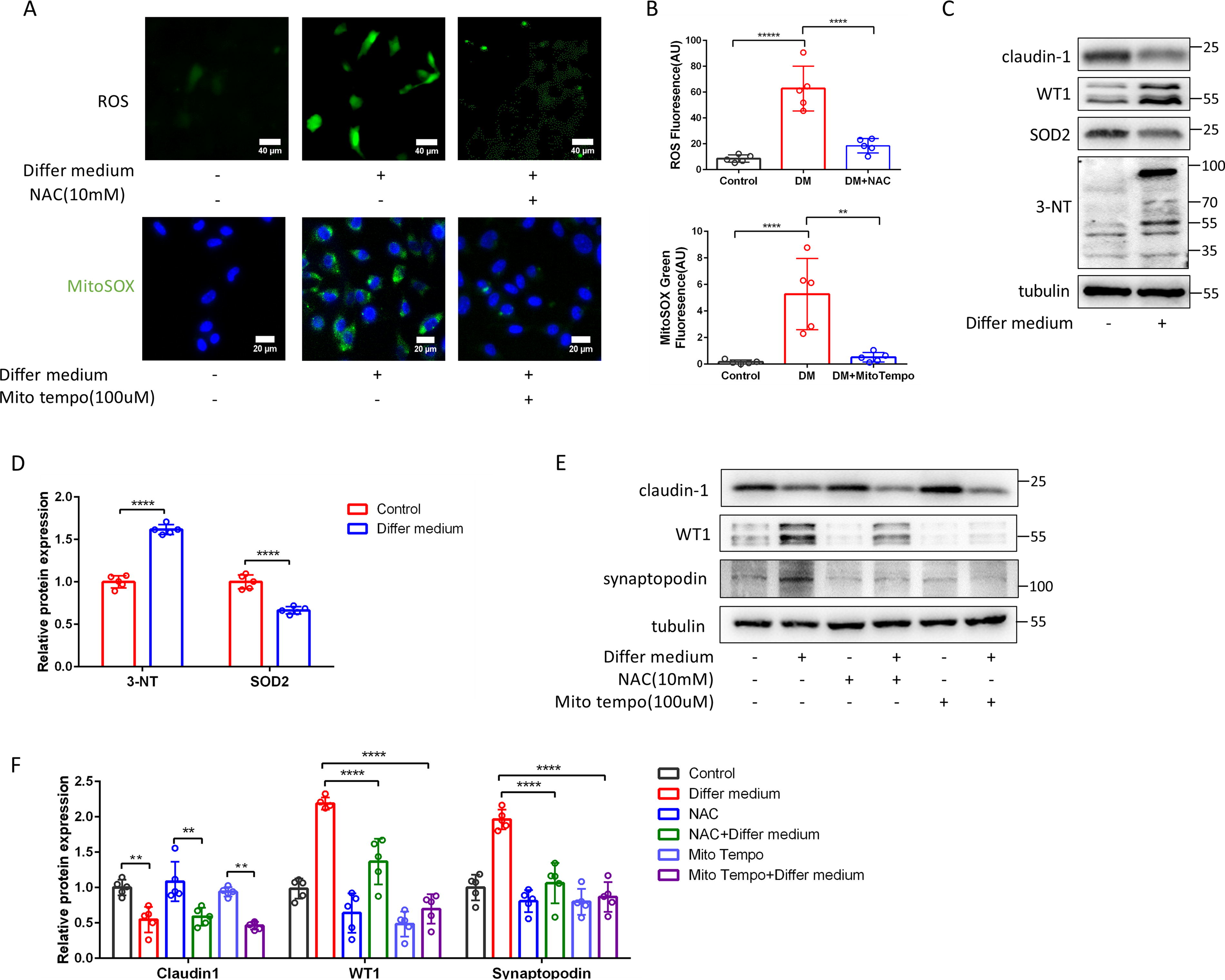
Increased mitochondria-derived ROS generation during the differentiation of PECs to podocytes.
Nrf2 mediated the differentiation of PECs to podocytes
Next, we investigated the ROS-associated molecule Nrf2: a key transcription factor that regulates the transcription of antioxidant proteins and, together with other molecules, regulates various cellular signaling pathways (Mitsuishi et al., 2012; Murakami et al., 2023; Paul et al., 2014; Tsai et al., 2013). Upon incubation of PECs with the VRADM differentiating medium, a significant increase in Nrf2 levels was observed in both nuclei and cytoplasm (Fig. 4A, 4B, 4C), which can be prevented by NAC or MitoTempo pretreatment (Fig. 4D, 4E). Nrf2 siRNA repressed Nrf2 production (Fig. 4K) and eliminated DM-induced WT1 or synaptopodin overexpression (Fig. 4F, 4G). Neh4 and Neh5 are structural domains of Nrf2 involved in regulating the expression of several downstream target genes. PECs infected with a lentivirus overexpressing Neh4 and Neh5 showed increased Nrf2, WT1, synaptopodin, and FOXC1 production (Fig. 4H, 4I; Supplementary Figure S3C). Continued administrating of dimethyl fumarate (DMF, an Nrf2 activator) enhanced the effects of DM on PECs (Fig. 4J). Increased Nrf2 overproduction was detected in ADR mice glomeruli, specifically localized to PECs at 2 and 6 weeks post-ADR injection (Fig. 4L).
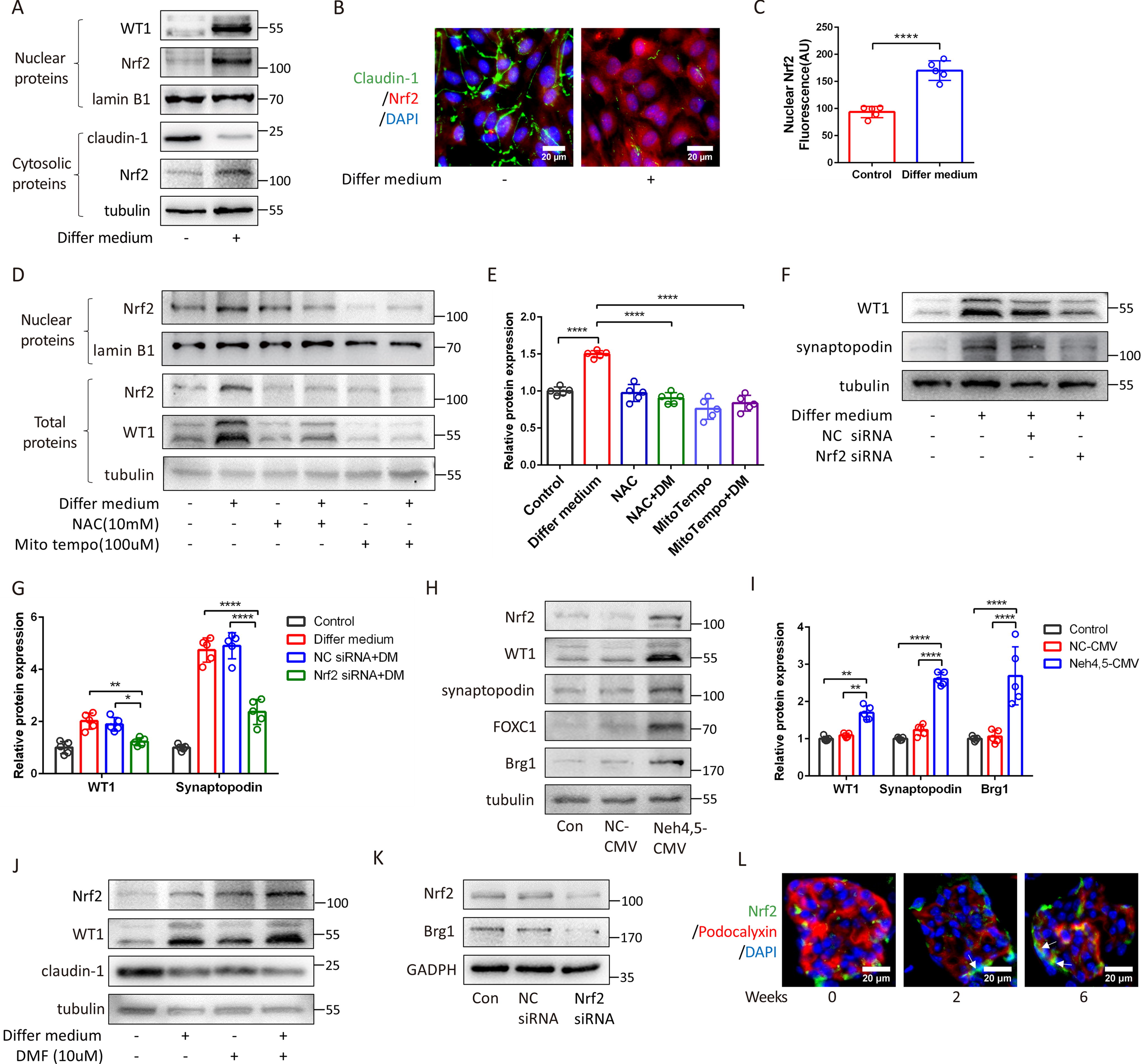
Mitochondrial ROS activates Nrf2 to mediate the differentiation of PECs to podocytes.
Nrf2 regulates Brg1 binding with WT-1 in the differentiation of PECs to podocytes
There is no direct evidence that Nrf2 directly regulates WT-1 or other podocyte marker gene expressions. However, Brahma-related gene 1 (Brg1) has been shown to mediate Nrf2-regulated heme oxidase-1 (HO-1) gene transcription (Sun et al., 2020). Therefore, we explored whether Brg1 is involved in the Nrf2-mediated transformation of PECs into podocytes. Our results demonstrated that incubation with VRADM led to a 2.82-fold upregulation of Brg1 mRNA (Fig. 5A). In addition, Brg1 and WT-1 proteins were overexpressed in differentiated PECs (Fig. 5B). Using siRNA to suppress Brg1 in PECs (Supplementary Figure S3B), we observed that the overexpression of WT-1 and FOXC1 in VRADM-incubated PECs was reduced by Brg1 knockdown (Fig. 5C, 5D). Next, we infected PECs with a Brg1-overexpressing adenovirus (Supplementary Figure S3D). A significant increase in WT-1, synaptopodin, as well as FOXC1, has been found in the infected PECs (Fig. 5E, 5F). To determine if Brg1 directly binds to the WT-1 promoter in PECs, we examined the binding of Brg1 to seven evolutionary conserved regions (ECRs) in PECs using chromatin immunoprecipitation (ChIP) analysis (Supplementary Figure S3E). We confirmed that Brg1 bound to two ECRs (+4.0 kb and +5.8 kb) within the intron of WT-1. Enrichment of Brg1 was more than 20-fold at the +4.0 kb ECR and more than 15-fold at the +5.8 kb ECR compared with the control (IgG) (Fig. 5G).

Nrf2 regulates the differentiation of PECs to podocytes by mediating Brg1.
Furthermore, we investigated the relationship among ROS, Nrf2, and Brg1. After ROS depletion, the overproduction of Brg1 was prevented by MitoTempo or NAC pretreatment (Fig. 5H, 5I). Suppressing Nrf2 with siRNA or increasing Nrf2 with Neh4-Neh5 lentivirus reduced or promoted Brg1 production (Fig. 4K, 4H). The interaction between Nrf2 and Brg1 was confirmed (Fig. 5H).
We also detected Brg1 expression in ADR mice. The number of Brg1-positive PECs increased by 3.19-folds (p < 0.0001) and 2.59-folds (p < 0.001) after ADR injection at 2 weeks and 6 weeks compared with that at 0 week (Fig. 5K, 5L).
Nrf2 activation or overexpression increases WT-1 positive cells in ADR mice
Normal mice and mice with ADR-induced nephropathy were treated with or without the nrf2 agonist bardoxolone-methyl (CDDO-Me). Pretreatment with CDDO-Me reduced UACR levels and glomerulosclerosis in ADR mice (Fig. 6A, 6B). WT-1 positive cells in ADR mice were decreased, which can be partially relieved by CDDO-Me pretreatment (Fig. 6A, 6C). We also found that mice treated with CDDO-Me exhibited Nrf2 overproduction in PECs (Fig. 6A).
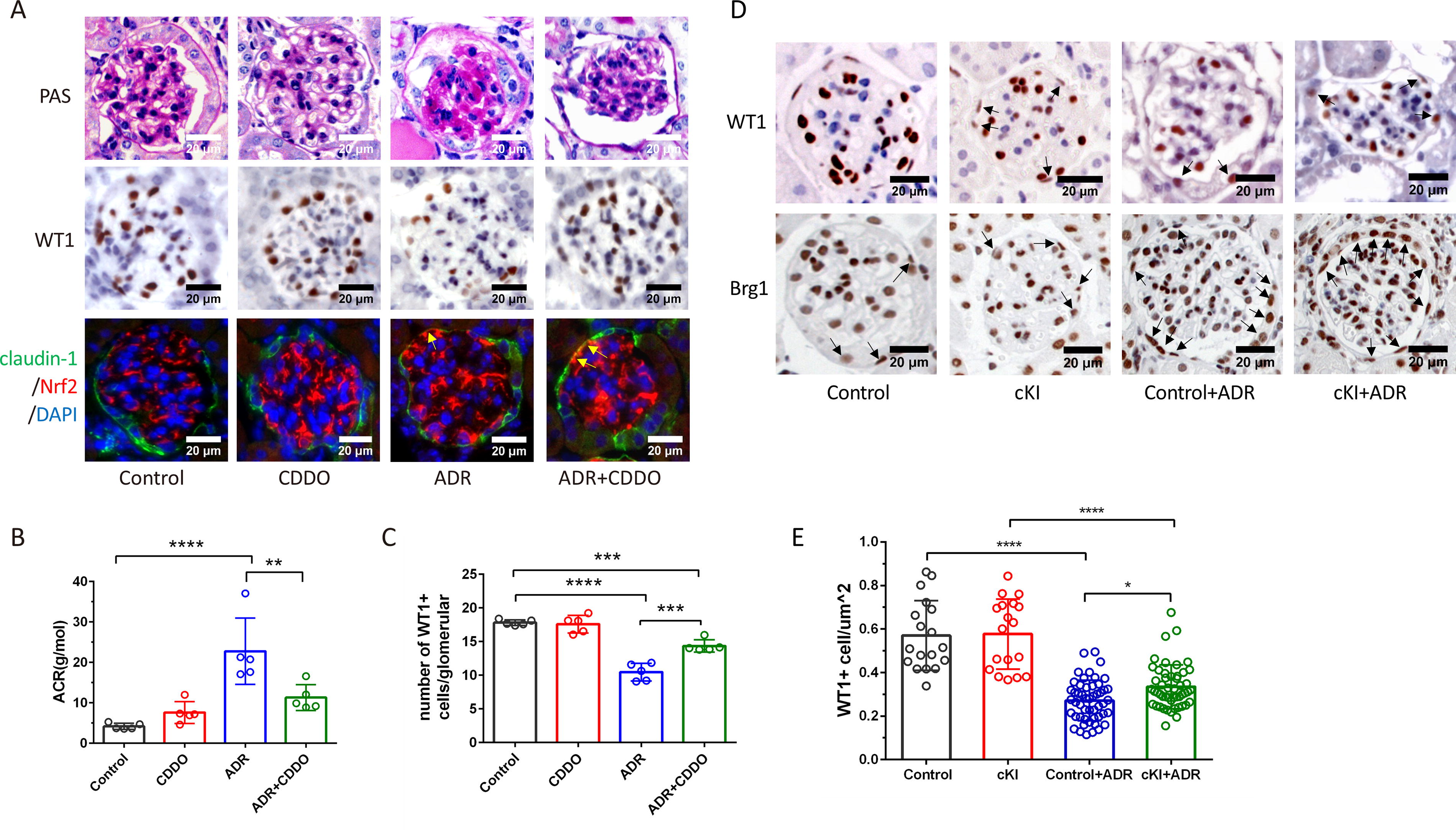
Nrf2 activation promotes PEC differentiation in vivo.
PEC specific human NRF2 knock-in mice were generated by crossing Pax8 Cre mice (PAX8 Cre+) with hNRF2flox/flox mice. Genotype identification results are shown in Supplementary Figure S4A. We found that human Nrf2 gene was expressed in the kidney and liver and Nrf2 protein produced in PECs in Pax8 Cre+: hNRF2flox/+ mice (Supplementary Figure S4B, S4C). However, the area of glomeruli in Pax8 Cre+: hNRF2flox/+ mice was smaller than that in hNRF2flox/+ mice (Supplementary Figure S4D). Thus, Pax8 Cre+: hNRF2flox/+ (conditional knock-in [cKI]) mice and hNRF2flox/+ (Control) mice were used for further study (Supplementary Figure S4E). We found that the number of Brg1 positive PECs in cKI mice was more than that in Control mice (Fig. 6D). Furthermore, WT1 positive cells per μm2 of glomeruli in cKI mice were more than that in Control mice (Fig. 6D, 6E).
Discussion
In the present study, we found an increase in mitochondrial generation during the differentiation of PECs into podocytes. Mitochondrial-derived ROS signaling mediated the translocation of Nrf2 from the cytoplasm to nuclei. Nrf2 regulated Brg1 binding with WT-1 promoter.
Recent studies have identified PECs as potential stem cells within the glomerulus that can differentiate toward podocytes and facilitate repair following podocyte injury. Retinoic acid plays an important role in the differentiation of PECs (Zhang et al., 2012b). However, in mice with ADR nephropathy, retinoic acid may be sequestered by proteinuria (Peired et al., 2013). In the present study, we observed a decrease in the number of WT1 positive cells in mice with ADR nephropathy, suggesting that podocyte regeneration by PECs is low efficiency although there was Nrf2 and Brg1 activation in PECs. We believed that endogenous retinoic acid alone activated ROS-Nrf2-Brg1 signaling insufficiently when retinol is blocked to generate retinoic acid by proteinuria. Direct activation of Nrf2 in vivo and in vitro study significantly increased PEC differentiation into podocyte, implicating that this may be a potential strategy to promote podocyte regeneration in proteinuria-related diseases.
Some studies have proposed that Notch signaling, miR-193a, and Gas1 are involved in regulating the differentiation of renal progenitors or PECs into podocytes (Kietzmann et al., 2015; Lasagni et al., 2010; Luna-Antonio et al., 2017), which implies that different signaling pathways regulate PEC transformation. Several studies have highlighted the significant role of mitochondria in stem cell differentiation (Chakrabarty and Chandel, 2021). Adenosine triphosphate (ATP) generation from glycolysis metabolism shifting to oxidative phosphorylation-dependent metabolism has been demonstrated in the differentiation of progenitor cells or stem cells (Hsu et al., 2016; Jorgensen and Khoury, 2021). Therefore, the generation of mitochondria may be necessary for energy supply in PEC transformation, as PECs had less mitochondria in the stable status, which may be similar with podocyte differentiation in vitro (Imasawa et al., 2017). Besides ATP generation, ROS production, release of cytochrome C as well as calcium regulation from mitochondria also control cell fate (Bigarella et al., 2014; Sun et al., 2016). In induced pluripotent stem cells and embryonic stem cells, a moderate increase in ROS is necessary for differentiation into different lineages (Tan and Suda, 2018). ROS affect multiple intracellular signaling pathways, such as nuclear factor kappa-B (NFκ B), mitogen-activated protein kinases (MAPKs), phosphoinositide 3 kinase-protein kinase B (PI3K-Akt), mitochondrial permeability transition pore (mPTP) and protein kinases, by activating/inactivating receptors, proteins, ions, and signaling molecules (Bae et al., 2011). Inhibition of the mitochondrial respiratory chain or depolarization of the mitochondrial membrane potential prevents the differentiation of pluripotent stem cells but maintains pluripotency (Mandal et al., 2011). Thus, fewer mitochondria maintaining a low basal level of ROS in stable PECs may be necessary to maintain the characteristics of potential stem cells.
ROS accumulation in mitochondria induces the transcription and protein production of antioxidant genes. The Forkhead box protein O (FOXO) family members and Nrf2 are the most important transcription factors that regulate antioxidant genes and protect cells from high levels of ROS. Recent advancements in understanding the Nrf2 pathway have underscored its crucial role in bolstering cellular resilience against oxidative stress and its potential therapeutic applications across a range of degenerative diseases(Calabrese et al., 2005; Catino et al., 2015; Mancuso et al., 2008; Scuto et al., 2020). Meanwhile, Nrf2 is involved in the regulation of the proliferation and differentiation of stem cells and progenitor cells (Jang et al., 2016; Paul et al., 2014; Tsai et al., 2013). Nrf2 belongs to the CNC transcription factor family and has a highly conserved basic leucine zipper structure (Motohashi et al., 2002). The Nrf2 protein consists of seven highly conserved Neh structural domains (1–7) (Baird and Dinkova-Kostova, 2011). Among them, Neh4 and Neh5 are N-terminal structural domains with different trans-activating properties that synergistically bind the coactivator CPEB binding protein and Brg1 to activate transcription of target genes (Katoh et al., 2001; Zhang et al., 2006). Yamamoto et al. found that under oxidative stress, Brg1 interacts with Nrf2 to enhance the transcriptional activity of Nrf2 by promoting the formation of Z-DNA and consequent recruitment of RNA polymerase II, which is critical for HO-1 activation (Zhang et al., 2006). Our findings indicate that PECs in conditional hNRF2 knock-in mice exhibit higher podocyte density, suggesting that Nrf2 plays a key role in podocyte repair. In fact, we screened Nrf2 expression in Nephroseq: Nrf2 expression increased in patients with chronic kidney disease (CKD) compared with those with normal kidney function (Nakagawa et al., 2015). Activation of Nrf2 signaling protects against podocyte injury and reduces 24-h urinary albumin in diabetic nephropathy mice (Lv et al., 2023), while Nrf2 deficiency worsens diabetic kidney disease (Liu et al., 2022), implicating Nrf2 as crucial for podocyte repair. However, Su K et al. found that Nrf2 knockout reduced UACR in Db/Db mice. In their study, Nrf2 deficiency was not conditional in PECs and was associated with reduced fasting blood glucose and glomerular filtration rate (GFR)/body weight (BW) ratio (Su et al., 2023). Thus, the decrease in UACR might be attributable to metabolic control. CDDO-Me has been shown to improve progression of renal dysfunction in patients with CKD and type-2 diabetes mellitus (Pergola et al., 2011). The urinary ACR, however, did not decrease in CDDO-Me groups, potentially due to the advanced stage of CKD in the enrolled patients. In patients with an estimated GFR of 20–45 mL/min/1.73 m2, more than half of the glomerulus might be destroyed. Thus, most PECs would be lost, and the capability of differentiation would decrease. Curcumin (Nrf2 activator) has been found to attenuate urinary excretion of albumin in patients with early-stage kidney disease due to diabetes mellitus (Yang et al., 2015).
The Neh4 and Neh5 structural domains of Nrf2 interact with Brg1 to regulate transcription. Brg1 is the central catalytic ATPase subunit of the switch/sucrose nonfermentable (SWI/SNF) chromatin remodeling complex (Clapier and Cairns, 2009). As a major factor in chromatin remodeling, Brg1 plays a key part in the replication, repair, recombination, and transcriptional regulation of DNA by interacting with various nuclear proteins (e.g., receptors, transcription factors, chromatin-modifying enzymes) (Mohrmann and Verrijzer, 2005; Trotter and Archer, 2007). Brg1 is involved in the proliferation, apoptosis, and differentiation of cells (Marathe et al., 2017; Singh et al., 2016; Wu et al., 2016). It also plays roles in cancer (Pyo et al., 2018), liver fibrosis (Li et al., 2018), and heart disease (Vieira et al., 2017). Riley et al. constructed a mouse model of epicardial conditional knockout of Brg1 expression. They found that targeting Brg1 reduced its binding to the ECRs of WT-1, leading to reduced WT-1 expression in the heart, indicating that BRG1 is required for WT-1 activation (Vieira et al., 2017). We demonstrated that Brg1 could bind to two ECR sites (+4.0 kb and +5.8 kb) in the WT-1 intron. Within the glomerulus, WT-1 (the most complex transcription factor of a podocyte) directly regulates the transcription of genes such as podocalyxin (Palmer et al., 2001) and nephrin (Wagner et al., 2004). Therefore, Brg1 might mediate the differentiation of PECs to podocytes through regulation of WT-1 transcription.
There are some limitations in the present study. We were unable to perform PECs lineage tracing in vivo, which would provide more intuitive evidence. Second, we do not establish a conditional knockout mouse line of Nrf2 or Brg1 mouse line to further confirm their roles in PEC differentiation in vivo. We believe only a small number of PECs differentiated into podocytes can be found in limited glomeruli. Thus, knockout Nrf2 or Brg1 in PECs may be unable to show the effects. Third, Pax8 presents in tubular epithelial cells and may not be so specific for PECs. To our knowledge, there is currently no suitable specific marker for PECs that can be used to develop conditional knockout or knock-in mice.
In summary, the present study demonstrated that mitochondrial-derived ROS generation, Nrf2 translocation, and Brg1 binding to WT-1 promoter are key events that regulate the differentiation of PECs into podocytes.
Materials and Methods
Reagents
ADR was purchased from MedChemExpress (Monmouth Junction, NJ, USA, cat# HY-15142). NAC was obtained from Sigma (Burlington, MA, USA, cat# A7250). MitoTempo was sourced from Abcam (Cambridge, UK, cat# ab144644). DMF was from APExBIO Technology (Houston, TX, USA, cat# B1931). CDDO-Me was purchased from Selleck Chemicals (Houston, TX, USA, cat# S8078).
Cell culture of PECs
To isolate glomeruli, mice were euthanized by overdose of pentobarbital sodium (1 g/kg BW). The kidney cortex was dissected free and cut into small pieces in culture medium. Then, the tissue suspension was poured onto a stainless-steel 80-mesh screen (pore size: 180 μm). The resultant mixture was transferred onto a 150-mesh screen (pore size: 100 μm) and a 200-mesh screen (pore size: 75 μm) successively. Glomeruli-enriched tissue was retained on a 200-mesh screen placed under a 150-mesh screen. Tissue fragments were collected and suspended in RPMI 1640 medium (Gibco, Grand Island, NY, USA, 11875085) supplemented with 5% fetal bovine serum, penicillin, streptomycin (Gibco, cat# 15070063), EGF (Gibco, cat# PMG8045), hydrocortisone (Tocris Bioscience, Bristol, UK, cat# 4093), and ITS (Gibco, cat# 41400045). After culture for one week, cellular outgrowths from glomeruli were detached using a trypsin-EDTA solution and filtered with a 40-μm strainer to remove glomerular fragments. Identification of PECs is shown in Supplementary Figure S1. PECs incubated with differentiated medium (RPMI 1640 with calcitriol 100 nM, ATRA 10 nM, and dexamethasone 50 nM) for 1 week.
Conditional knock-in of Nrf2 in mouse PECs
To generate PEC specific Nrf2 knock-in mice, CAG-LSL-human NRF2-IRES-luciferase-2A-tdTomato mice (C57BL/6Smoc-Gt[ROSA]26Sorem1(CAG-LSL-NRF2-IRES-luciferase-2A-tdTomato)Smoc ) were crossed to Pax8-Cre mice (C57BL/6Smoc-Pax8em1(Cre-WPRE-polyA)Smoc ). Both mice were purchased from Shanghai Model Organisms Center, Inc. (Shanghai, China). Nrf2 transgenic/PAX8-Cre mice were confirmed by PCR genotyping. We found that the livers and kidneys from Pax8 Cre+: hNRF2flox/+ mice expressed human nfe2l2 gene well (Supplementary Figure S4B). Furthermore, we demonstrated that PECs in Pax8 Cre+: hNRF2flox/+ mice can produce Nrf2 protein (Supplementary Figure S4C) and the glomeruli smaller in Pax8 Cre+: hNRF2flox/+ mice than those in hNRF2flox/+ mice (Supplementary Figure S4D).
Animal experiments
Animal experiments were undertaken using a protocol approved by Renji Hospital within Shanghai Jiaotong University School of Medicine (Shanghai, China). Animals were housed under defined conditions (22°C, specified pathogen-free, 12-h day/12-h night cycle) and fed standard laboratory chow ad libitum. Male specific-pathogen-free BALB/c mice (6–8 weeks; Shanghai SLAC Laboratory Animals, Shanghai, China) were divided into two groups as follows: control and ADR treatment. Nephropathy was induced by a single injection of ADR (10 mg/kg BW, i.v.). At 2 weeks and 6 weeks of age, mice were sacrificed using pentobarbital sodium. A urine sample was collected and kidneys harvested for subsequent analyses.
At the age of 8 weeks, 20 male BALB/c mice were weighed and randomized into four groups of five as follows: vehicle control, CDDO-Me treatment, ADR treatment, and ADR+CDDO-Me treatment. Mice in vehicle control group were administered 0.1 mL of vehicle consisting of dimethyl sulfoxide and Sulfobutylether-β-Cyclodextrin (SBE-β-CD) in saline by oral gavage daily. In the CDDO-Me treatment group, mice were administered CDDO-Me at a dose of 10 μmol/kg/day in 0.1 mL of vehicle. At the age of 6 weeks, mice from the four groups were killed 24 h after the final administration of vehicle or CDDO-Me.
Transgenic Pax8 Cre+: hNRF2flox/+ mice (cKI) and hNRF2flox/+ mice (Control) grouping is shown in Supplementary Figure S4E. Mice were subjected to unilateral nephrectomy at 6–8 weeks old. Mice with ADR (10 mg/kg BW, i.v.) or saline were injected into mice in ADR or control groups 1 week after nephrectomy. Six weeks after injection, mice were anesthetized (pentobarbital injection) and perfused with 1% phosphate-buffered saline (PBS). Samples of urine and blood were collected, and the kidneys were harvested for processing.
Immunoblotting
Protein was extracted from fractionated cells. The protein concentration was determined using a Bicinchoninic-Acid Assay Kit (Thermo Scientific, Waltham, MA, USA). We wished to extract protein from cell supernatants. The cell supernatant was collected and centrifuged at 12,000×g for 10 min at 4°C, and 20 μL of the cell supernatant was aspirated from near the bottom of the centrifuge tube (taking care not to touch the cell pellet). Protein samples were resolved by sodium dodecyl sulfate polyacrylamide gel electrophoresis (SDS-PAGE) and transferred to nitrocellulose membranes. Immunoblot bands were visualized using an imaging system from Tanon (Zhejiang, China).
The antibodies we used were as follows: anti-synaptopodin (Proteintech, Wuhan, China, cat# 21064), anti-PGC1α (Proteintech, cat# 66369), anti-SOD2 (Proteintech, cat# 24127), anti-Nrf2 (Proteintech, cat# 80593), anti-tubulin (Proteintech, cat# 10094), anti-WT1 (Cell Signaling Technology, Danvers, MA, USA, cat# 83535), anti-GAPDH (Santa Cruz, CA, USA, cat# sc-47724), anti-CytC (Santa Cruz, cat# sc-13560), anti-TOMM20 (Santa Cruz, cat# sc-17764), anti-COX IV (Abcam, cat# ab16056), anti-claudin-1 (Abcam, cat# ab307692), anti-Brg1 (Abcam, cat# ab150077), anti-FOXC1(Abcam, cat# ab223850), anti-nephrin (Abcam, cat# ab216692), and anti-3-NT (Abcam, cat# ab314438).
RNA extraction and real-time reverse transcription–quantitative polymerase chain reaction
Total RNA was extracted using TRIzol® Reagent (Thermo Scientific) according to manufacturer’s instructions. After reverse transcription, complementary (c)DNA samples were denatured and amplified using a LightCycler™ 480 real-time PCR system (Roche Applied Science, Mannheim, Germany). Amplification conditions were 45 cycles of 95°C for 30 s, 95°C for 10 s, and 60°C for 20 s. The primers we used are shown in Table 1.
Primers Used in Real-Time Reverse Transcription–Quantitative Polymerase Chain Reaction for Selected Genes
RNA interference
PGC1α siRNA, Nrf2 siRNA, Brg1 siRNA, and negative control siRNA were purchased from Jiman (Shanghai, China). siRNAs (100 nmol/L) were transfected transiently using Lipofectamine™ 3000 according to manufacturer’s (Thermo Scientific) instructions.
Immunofluorescence and immunohistochemical staining
Cryosections of thickness of 2 μm were prepared using a cryostat. They were fixed in 4% paraformaldehyde for 15 min. After blocking, the cryosections were incubated with primary antibodies and then with Alexa Fluor® 488 secondary antibody and Alexa Fluor® 555 secondary antibody (1:200; Abcam, cat# ab150077, ab150078). Fluorescence images were recorded using a confocal microscope (TCS SP5II; Leica, Bensheim, Germany). The primary antibodies we used were anti-podocalyxin (1:200; R&D Systems), anti-claudin-1 (1:200; Abcam), and anti-PGC1α (1:100; Proteintech).
For immunohistochemical analyses, after deparaffinization, rehydration, antigen retrieval, and blockade, cryosections were incubated with anti-PCNA (1:100; Proteintech, cat# 60097) and anti-Brg1 (1:200, Abcam). Then, cryosections were incubated with a horseradish peroxidase-labeled secondary antibody (Beyotime Institute of Biotechnology, Shanghai, China).
Transmission electron microscopy and immunoelectron microscopy
Cortical kidney sections were fixed with 2.5% glutaraldehyde in PBS at 4°C overnight, postfixed in 1.0% OsO4, and embedded in LR White Resin (London Resin, Hampshire, England). Ultrathin sections (70 nm) were stained with uranyl acetate and examined under an electron microscope (CM120; Philips, Eindhoven, the Netherlands) at 80 kV.
For immunoelectron microscopy, kidneys were perfused through the heart with 4% paraformaldehyde/2.5% glutaraldehyde in PBS for 20 min at 4°C. Cortical kidney sections were cut and dipped in 4% paraformaldehyde/2.5% glutaraldehyde for 1 h. Sections were embedded in LR White Resin, and ultrathin sections were prepared. Ultrathin sections were incubated with rabbit anticlaudin-1 antibody (1:50) and gold conjugate nanoparticles (10 nm; 1:50) overnight at 4°C. An electron microscope (CM120; Philips) was used for examination of sections.
Construction and transfection of plasmids
A plasmid directing overexpression of the Nrf2 ECH homolog Neh4 and Neh5 domains of Nrf2 was purchased from Jiman. A fragment of the target gene was first amplified using the following primers: forward, 5′-CGAATTCGAAGTATACCTCGAGGAATTCGCCACCATGGGATC-3′ and reverse, 5′-GATCGCAGATCCTTGGATCCTCACGAGGAGATCGATGAGTAAAAATG-3′. Then, the amplified fragment was ligated into a lentiviral vector (PGMLV-CMV-NeH4-P2A-NeH5-EF1-ZsGreen1-T2A-Puro) through restriction sites contained at both ends. The ligation product was transfected into competent bacterial cells. Newly grown monoclonal colonies were sequenced to confirm their identity. We aligned the correct clones to construct the overexpressing adenoviral vector containing the gene of interest. Plasmid transfection was undertaken using a FuGENE™ Transfection Kit (Promega, Madison, WI, USA) according to manufacturer’s instructions.
The brg1 overexpression plasmid was purchased from Shanghai Jikai Gene Chemical Technology (Shanghai, China). Theforward primer was 5′-AGGTCGACTCTAGAGGATCCCGCCACCATGTCTACTCCAGACCCACCCTTGGGTG-3′, and the reverse primer was 5′-TCCTTGTAGTCCATACCGGTGTCTTCCTCACTGCCACTTCCTGAGCG-3′. The amplified fragment was ligated into an adenoviral vector (CMV-MCS-3FLAG-SV40-EGFP).
Detection of albumin and creatinine in urine
Mouse albumin was determined using an Albumin Assay Kit (Jiancheng, Nanjing, China). Absorbance was determined at 510 nm using a microplate reader. The creatinine concentration in urine was determined using a creatinine assay kit (Cayman Chemicals, Ann Arbor, MI, USA) according to manufacturer’s instructions. The creatinine concentration in urine samples was determined using a standard curve for creatinine. The dynamic range of the kit was 0–15 mg/dL of creatinine. The absorbance at 500 nm was determined using a microplate reader.
Coimmunoprecipitation
The cells were lysed using a commercial kit (abs955, Absin Bioscience Inc., Shanghai, China). After 10-min centrifugation at 14,000×g, the supernatants were incubated with anti-Flag (Nrf2) antibody and control IgG, respectively, at 4°C overnight followed by the treatment with protein A/G agarose beads and further incubated at 4°C overnight. Then, lysates were centrifuged, and the precipitate was washed thrice with cold wash buffer. Finally, the immunoprecipitated proteins of Nrf2 and Brg1 were detected by Western blot analysis.
ChIP
PECs were crosslinked with 1% formaldehyde at room temperature for 10 min, incubated first in nuclei extraction buffer, then in lysis buffer, and sonicated. Solubilized chromatin was incubated overnight at 4°C with a rabbit monoclonal anti-Brg1 antibody (Abcam) coupled to antirabbit immunoglobulin (Ig)G Dynabeads. After primary ChIP, specific immunoprecipitates were washed, eluted, and incubated with dithiothreitol (10 mM) for 30 min at 37°C. Then, they were diluted 10-fold in ChIP buffer (EDTA [10 mM], Tris-HCl [50 mM, pH 8.0], 1% SDS, protease inhibitors) and reimmunoprecipitated with secondary antibodies. Isolation of reimmunoprecipitated chromatin was done according to manufacturer’s instructions (Millipore, Bedford, MA, USA). Extracted DNA was used for real-time reverse transcription–quantitative polymerase chain reaction (RT-qPCR) (see above). Fold enrichment was determined as percent fold-change in ChIP signal/IgG signal at the target region.
Statistical analyses
The results are expressed as mean ± standard deviation (SD) for each group. Statistical analyses were undertaken using one-way analysis of variance (ANOVA) followed by a Newman–Keuls multiple-comparisons test using Prism 6.0 (GraphPad, La Jolla, CA, USA) for analyses of data from histology, immunohistochemistry, real-time RT-qPCR, and Western blot studies. Repeat ANOVA was used for analyses of albumin excretion and serum levels of creatinine. p < 0.05 was considered significant.
Footnotes
Acknowledgments
The authors thank the staffs in Renal Pathology Laboratory of Shanghai Jiao Tong University affiliated Renji Hospital for their technical support.
Authors’ Contributions
Conceived and designed the experiments: L.G. and R.L. Performed the experiments: M.W., W.W., J.L., and K.X. Analyzed the data: A.S., B.Z., and W.W. Contributed reagents/materials/analysis tools: L.M. and Y.L. Wrote the first draft of the article: M.W., W.W., and K.X. Revised the article: L.G. L.G. is also the guarantor of this work and, as such, had full access to all the data in the study and takes responsibility for the integrity of the data and the accuracy of the data analysis.
Ethics Approval
The animal study was approved by the Institutional Animal Care and Use Committee (IACUC) of Shanghai Jiaotong University.
Availability of Data and Materials
All of the data of this study are available from the corresponding author.
Author Disclosure Statement
All authors declared no financial or scientific conflicts of interest with regard to the research described in this article.
Funding Information
This work was supported by the
Supplementary Material
Supplementary Figure S1
Supplementary Figure S2
Supplementary Figure S3
Supplementary Figure S4
Abbreviations Used
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.