
Abstract
Aims:
Myocardial fibrosis is an important medium for atrial fibrillation (AF). Exosomes have been demonstrated to affect the development of AF. This study explored the molecular mechanism of exosomes from patients with AF (AF-exo) mediating myocardial fibrosis and thus affecting the development of AF.
Results:
Prolactin-induced protein (PIP) is highly expressed in AF-exo. AF-exo promoted the proliferation and activation of cardiac fibroblasts (CFs) as well as the migration and endothelial-to-mesenchymal transition (EndMT) of human umbilical vein endothelial cells (HUVECs). However, the effect of AF-exo on CFs and HUVECs was mitigated by PIP-specific short hairpin RNA (shPIP). Adeno-associated virus (AAV)-shPIP reduced the incidence and duration of AF in rats, and improved myocardial fibrosis and collagen deposition. ATPase plasma membrane Ca2+ transporting 2 (ATP2B2) overexpression or inhibition reverses the role of PIP or shPIP in CFs, HUVECs, and AF rats. Activation of the cyclic guanosine monophosphate/protein kinase G (cGMP/PKG) pathway is beneficial to alleviate myocardial fibrosis, but this effect is mitigated by shATP2B2.
Innovation:
Our investigation substantiates the pivotal role of the PIP/ATP2B2 axis in both HUVEC myocardial fibrosis and EndMT progression. Our findings suggest that AF-exo can suppress the activation of the cGMP/PKG pathway mediated by ATP2B2 through exosomal PIP, thus promoting myocardial fibrosis, indicating potential targets for novel antifibrotic drug development targeting either PIP or ATP2B2.
Conclusion:
Exosomal PIP can inhibit the activation of cGMP/PKG pathway mediated by ATP2B2, thus promoting the development of AF. Antioxid. Redox Signal. 43, 14–36.

Introduction
Atrial fibrillation (AF) is the most common tachyarrhythmia in clinic at present. Severe adverse complications such as stroke, heart failure, and even sudden death caused by its hemodynamic changes and hypercoagulability have caused a very heavy economic burden to families and society (Sagris et al., 2021). There is extensive evidence that atrial fibrosis is an important pathophysiological factor of AF, because fibroblasts will differentiate into myofibroblasts after stimulation, which can reduce the conduction velocity of myocardium, promote the production of arrhythmogenic substrate, and finally change the cardiac ultrastructure (Li et al., 2021). Although the relationship between atrial fibrosis and AF has been widely reported, the molecular mechanism of this process has not been fully clarified. Therefore, knowing more about the mechanism of atrial fibrosis may provide new strategies for the treatment and prevention of AF.
Recent studies show that exosomes play an important role in the development of AF (Huang et al., 2021). Exosomes are extracellular vesicles with a diameter of 40–160 nm, which can carry various goods to target cells. Studies have shown that there are differences in the expression of circulating exosomes between patients with AF and the control group (Wang et al., 2019), so some differential exosome proteins have the potential to be used as biomarkers of AF. On the contrary, it has been demonstrated that exosomes derived from atrial myocytes can promote atrial fibrosis and lead to AF (Hao et al., 2022). Therefore, we analyzed the plasma-derived exosomes of patients with AF by proteomics, and obtained 12 proteins with abnormally high expression in the exosomes of patients, among which prolactin-induced protein (PIP) attracted our attention because of its role in promoting fibrosis (Liang et al., 2022). PIP is a glycoprotein first found in breast milk and saliva (Haagensen et al., 1980; Haagensen et al., 1979). Subsequent studies have found that PIP can be expressed in cancer and is related to the migration and adhesion of cancer cells (Urbaniak et al., 2018). However, the role of PIP in AF is still unknown and needs further exploration.
Innovation
Our investigation substantiates the pivotal role of the PIP/ATP2B2 axis in both HUVEC myocardial fibrosis and EndMT progression. Our findings suggest that AF-exo can suppress the activation of cGMP/PKG pathway mediated by ATP2B2 through exosomal PIP, thus promoting myocardial fibrosis, indicating potential targets for novel antifibrotic drug development targeting either PIP or ATP2B2.
By analyzing the differential proteins of AF rats with or without PIP-specific short hairpin RNA (shPIP) injection, we found that these proteins were enriched in the cyclic guanosine monophosphate/protein kinase G (cGMP/PKG) pathway. It is reported that the cGMP/PKG pathway is a common medium of cardiac protection and can alleviate cardiac fibrosis (Meng et al., 2023). Among these differential proteins, ATPase plasma membrane Ca2+ transporting 2 (ATP2B2) is the most significant. ATP2B2 belongs to the P-type primary ion transport ATPase family, which has been reported to be related to cardiac conduction and sound sensory processing (Agha et al., 2019; Tao et al., 2023). Previous studies reported that the expression of ATP2B2 decreased when the heart was damaged (Xia et al., 2011). However, the role of ATP2B2 in AF is still unknown. Therefore, we intend to study whether PIP can promote cardiac fibrosis by regulating ATP2B2-mediated cGMP/PKG pathway.
Results
PIP was highly expressed in exosome derived from patients with AF
The clinical characteristics of study population are given in Table1. In total, the majority of demographic characteristics, all concomitant diseases, did not show statistically significant differences between these two groups, except left atrial diameter (LA) (p < 0.001). We extracted exosomes from the sera of patients with AF and healthy volunteers (HC), and identified the exosome markers CD9, CD63, and Tsg101 by Western blot. Interestingly, we found that the levels of CD9 and Tsg101 in the exosomes from patients with AF were higher than those from HC, while the levels of CD63 in the exosomes from patients with AF were lower than those from HC (Supplementary Fig. S1A–D). We also analyzed the exosomes from patients with AF and HC by liquid chromatography-mass spectrometry (LC-MS) (Supplementary Fig. S2A). The principal component analysis (PCA) result showed that the samples between groups were scattered, indicating significant differences between groups (Supplementary Fig. S2B). Among the 323 coexpressed proteins (Supplementary Fig. S2C), there were 29 differentially expressed proteins (Supplementary Fig. S2D, E). We further identified PIP, which has not been studied in AF but can promote fibrosis among these 29 differentially expressed proteins through literature search for subsequent analysis. Consistently, the expression of PIP in exosomes from patients with AF is much higher than that from HC (Fig. 1A, B, p < 0.001).

Baseline Clinical Profile of the Study Population
BMI, body mass index; LA, left atrial diameter; LVEDD, left ventricular end diastolic diameter; LVEF, left ventricular ejection fraction.
Subsequently, we cocultured the exosomes from patients with AF with CFs and HUVECs, respectively, and used PKH26 to trace the exosomes. As shown in Figure 1C, D, it can be seen that the exosomes were internalized by CFs and HUVECs. To study the role of exosomal PIP in AF, we transfected oe-PIP and shPIP into CFs and HUVECs, respectively, and the transfection efficiency is shown in Figure 1E, F (p < 0.01). Because shPIP-3 has the best inhibition efficiency, shPIP-3 is selected for PIP expression interference.
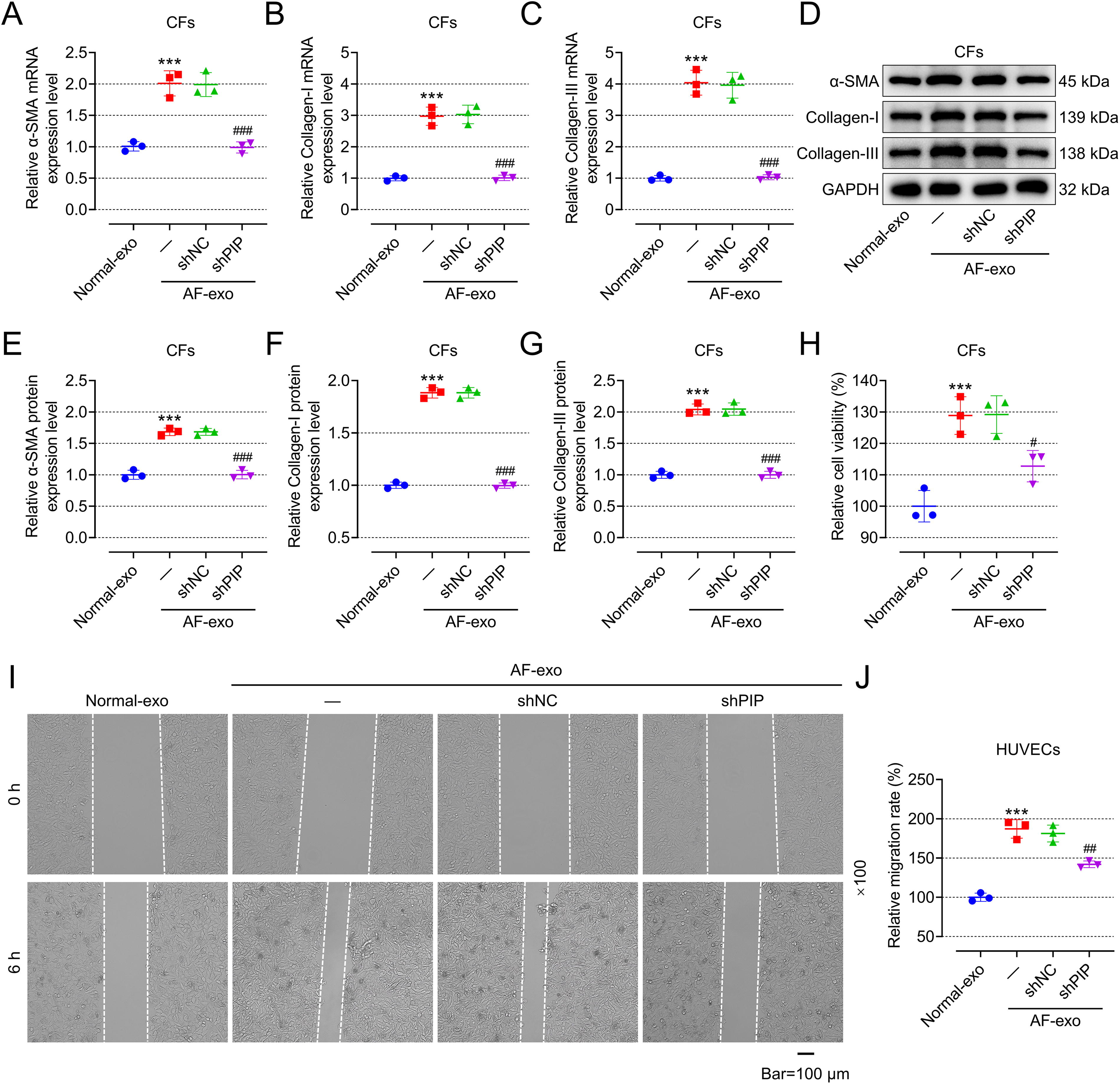
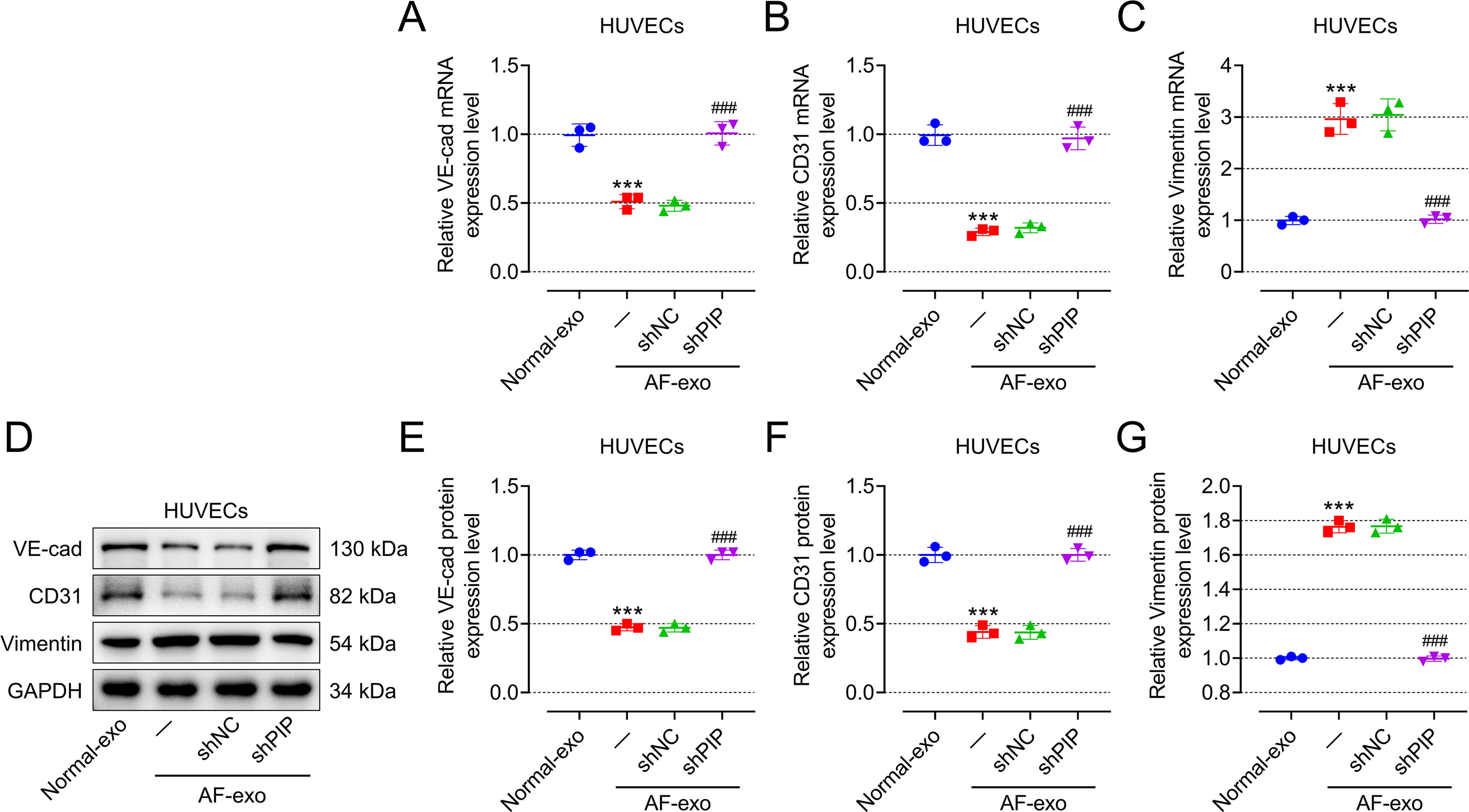
Exosomal PIP derived from patients with AF facilitated fibrosis and viability of CFs as well as promoted migration and EndMT of HUVECs
Compared with those CFs or HUVECs cocultured with exosomes from HC (normal-exo), CFs incubated with exosomes from patients with AF (AF-exo) showed higher fibrosis phenotype (Fig. 2A–G, p < 0.001) and cell viability (Fig. 2H, p < 0.001), while HUVECs incubated with AF-exo showed higher migration ability (Fig. 2I, J, p < 0.001) and EndMT phenotype (Fig. 3A–G, p < 0.001). To study whether AF-exo affects the function of CFs or HUVECs by overexpressing PIP, CFs or HUVECs were transfected with shNC or shPIP, and then were incubated with AF-exo. Interestingly, the promotion of AF-exo on the fibrosis (Fig. 2A–G) and viability (Fig. 2H) of CFs and the promotion of AF-exo on migration (Fig. 2I, J) and EndMT (Fig. 3A–G) of HUVECs were mitigated by shPIP (p < 0.05).
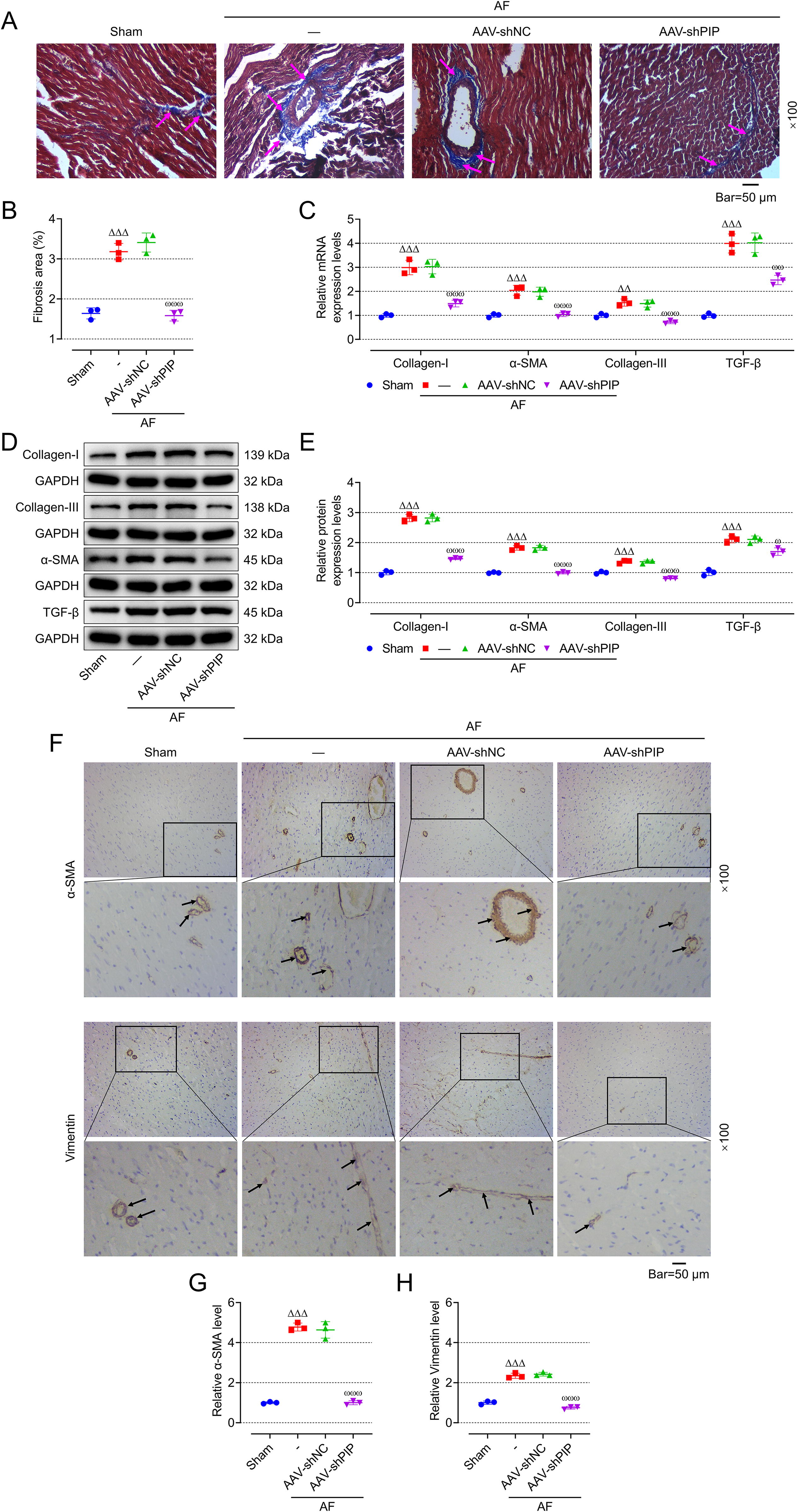
AAV-ShPIP alleviated the incidence and duration of AF as well as inhibited myocardial fibrosis in AF rats
We found that compared with the Control group, p wave disappeared in the electrocardiogram of rats in Model group, and changed into f wave with inconsistent frequency, amplitude, and shape, isopotential line disappeared, and QRS wave shape changed (Fig. 4A), indicating modeling was successful. Moreover, constructing an AF model using ACH-CaCl2 can increase the incidence and duration of AF in rats (Fig. 4B, C, p < 0.001), but has no effect on cardiac function (Fig. 4D–G), Representative ultrasound figures are presented (Fig. 4H). In addition, the myocardial fibrosis of AF rats was enhanced by Masson staining (Fig. 5A, B), which was also confirmed by an increase in the mRNA and protein expression of fibrosis-related proteins (collagen-I, α-smooth muscle actin [α-SMA], collagen-III, transforming growth factor-β [TGF-β]) in left atrial tissue (Fig. 5C–E p < 0.001). At the same time, immunohistochemistry (IHC) also showed that the expression of α-SMA and vimentin (Fig. 5F–H, p < 0.001) in the left atrial tissue of AF rats was enhanced, indicating that cardiac fibrosis and EndMT in AF rats were enhanced. To further verify the effect of PIP on AF, AAV-shPIP was injected into the tail vein of AF rats to inhibit the expression of PIP. The incidence and duration of AF in AAV-ShPIP-treated rats were significantly lower than those in AAV-ShNC-treated rats (Fig. 4B, C, p < 0.001), but there was no significant change in cardiac function (Fig. 4D–G) between the two groups. In addition, AAV-shPIP reduced the cardiac fibrosis (Fig. 5A, B) in AF rats and decreased the expression of fibrosis-related proteins and vimentin in left atrial tissue (Fig. 5C–E and 5F–H, p < 0.05).

Identification of downstream target proteins of PIP
To further explore the molecular mechanism of PIP downstream, we collected the left atrial tissue of rats injected with AAV-shNC and AAV-shPIP and performed proteomic analysis. Supplementary figure S3A shows the differential expression of proteins. Then we analyzed the enrichment of Gene Ontology (GO) and Kyoto Encyclopedia of Genes and Genomes (KEGG) and found that these proteins were enriched in the cGMP/PKG pathway (Supplementary Fig. S3B, S4A, B, S5A, B). Subsequently, we analyzed the differentially expressed proteins related to the cGMP/PKG pathway, the most significant of which was ATP2B2, whose expression decreased significantly when PIP was knocked out. Therefore, we intend to study whether PIP influences the development of AF by regulating ATP2B2.
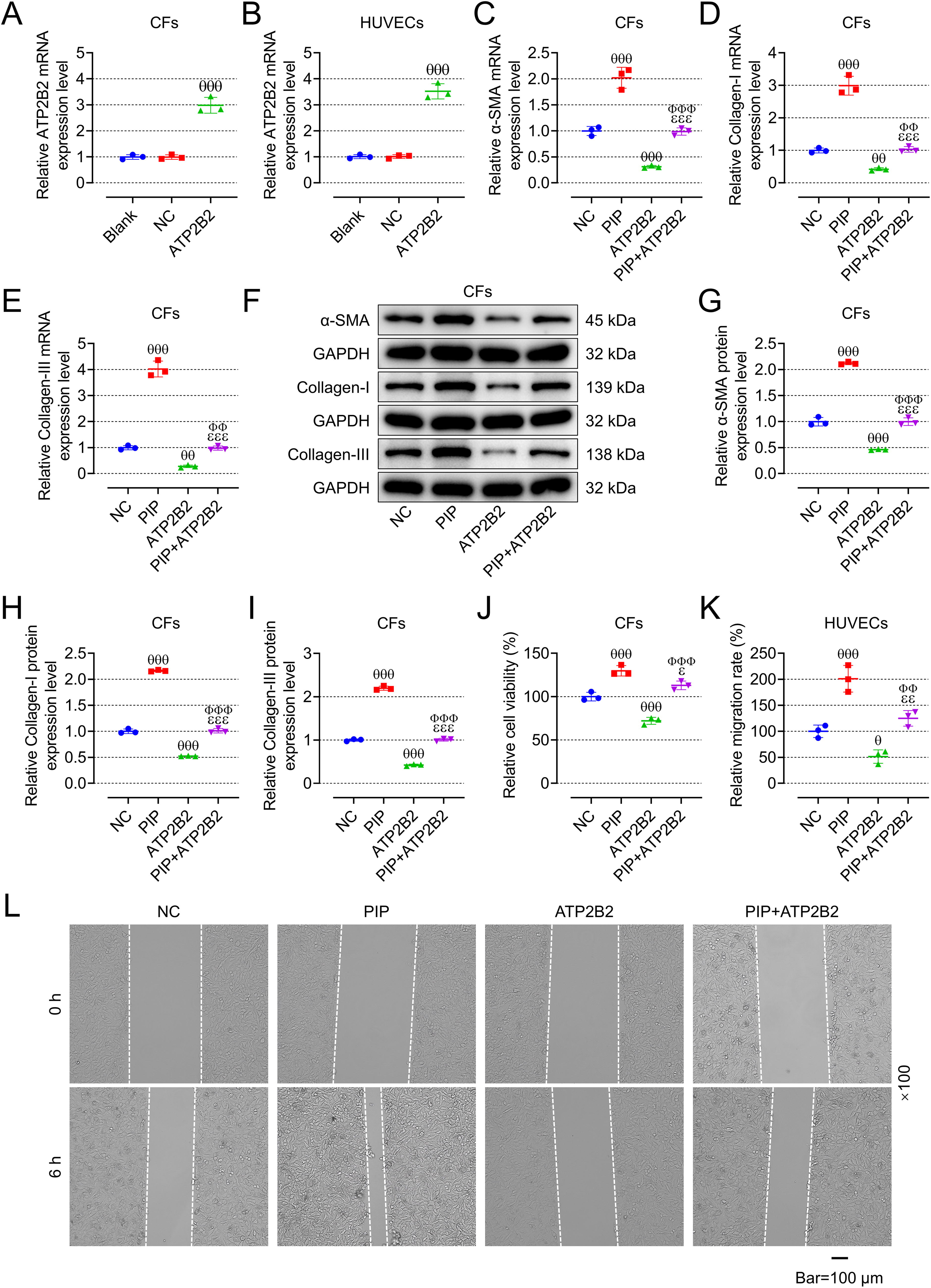
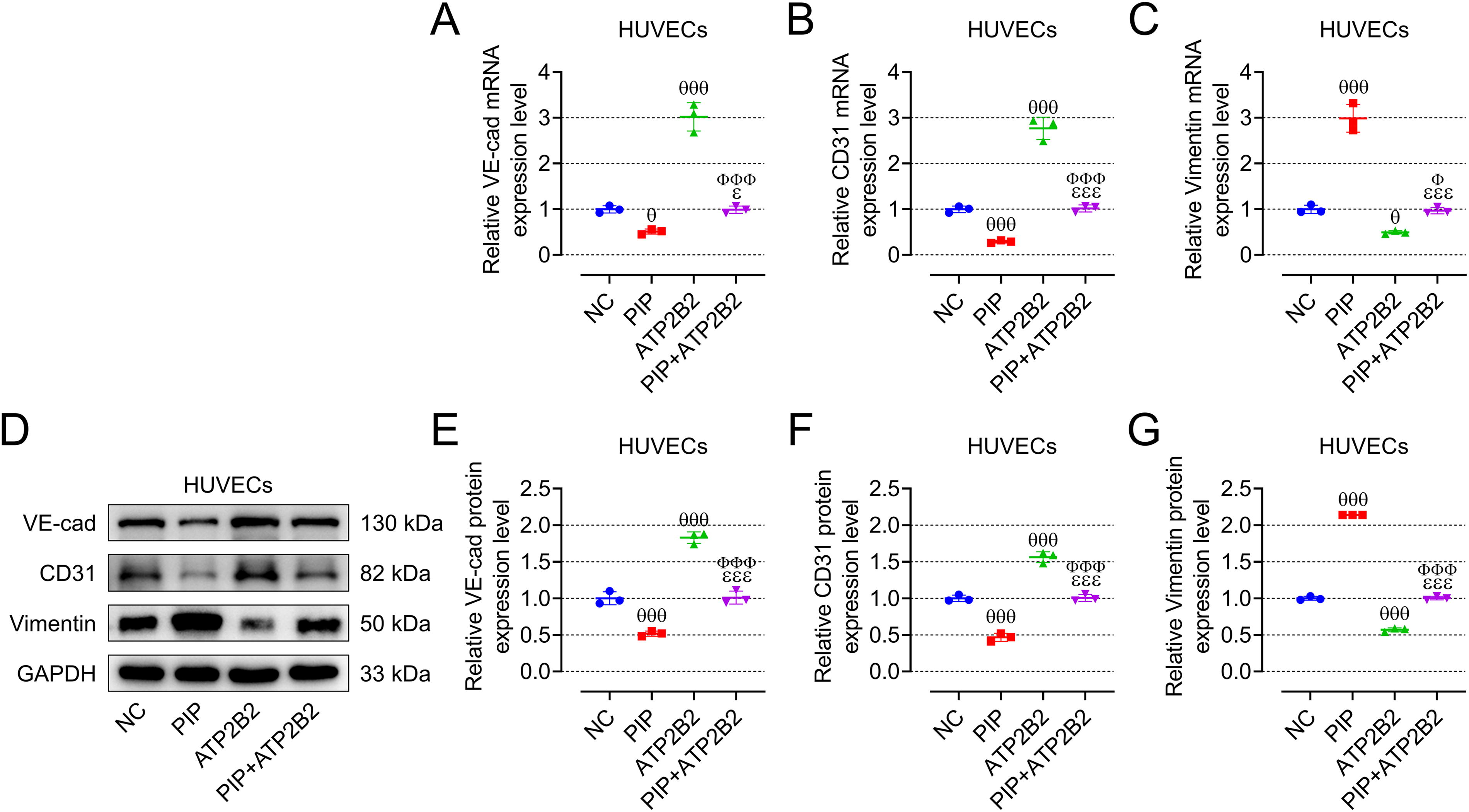
ATP2B2 overexpression mitigated the effects of PIP on fibrosis and viability of CFs as well as migration and EndMT of HUVECs
We first verified the role of PIP/ATP2B2 axis in AF in vitro, and so we constructed oe-ATP2B2 and verified the transfection efficiency in CFs and HUVECs, respectively (Fig. 6A, B, p < 0.001). Contrary to oe-PIP, oe-ATP2B2 inhibited the expression of fibrosis-related genes and proteins in CFs (Fig. 6C–1, p < 0.01), promoted the vitality of CFs (Fig. 6J, p < 0.001), and the migration of HUVECs (Fig. 6K, L, p < 0.05), and at the same time upregulated the expression of VE-cadherin and CD31 in HUVECs and downregulated the expression of vimentin (Fig. 7A–G, p < 0.05). Moreover, oe-ATP2B2 also mitigated the regulation of oe-PIP on the fibrosis and vitality of CFs as well as the migration and EndMT of HUVECs (Fig. 6C–L and 7A–G, p < 0.05).
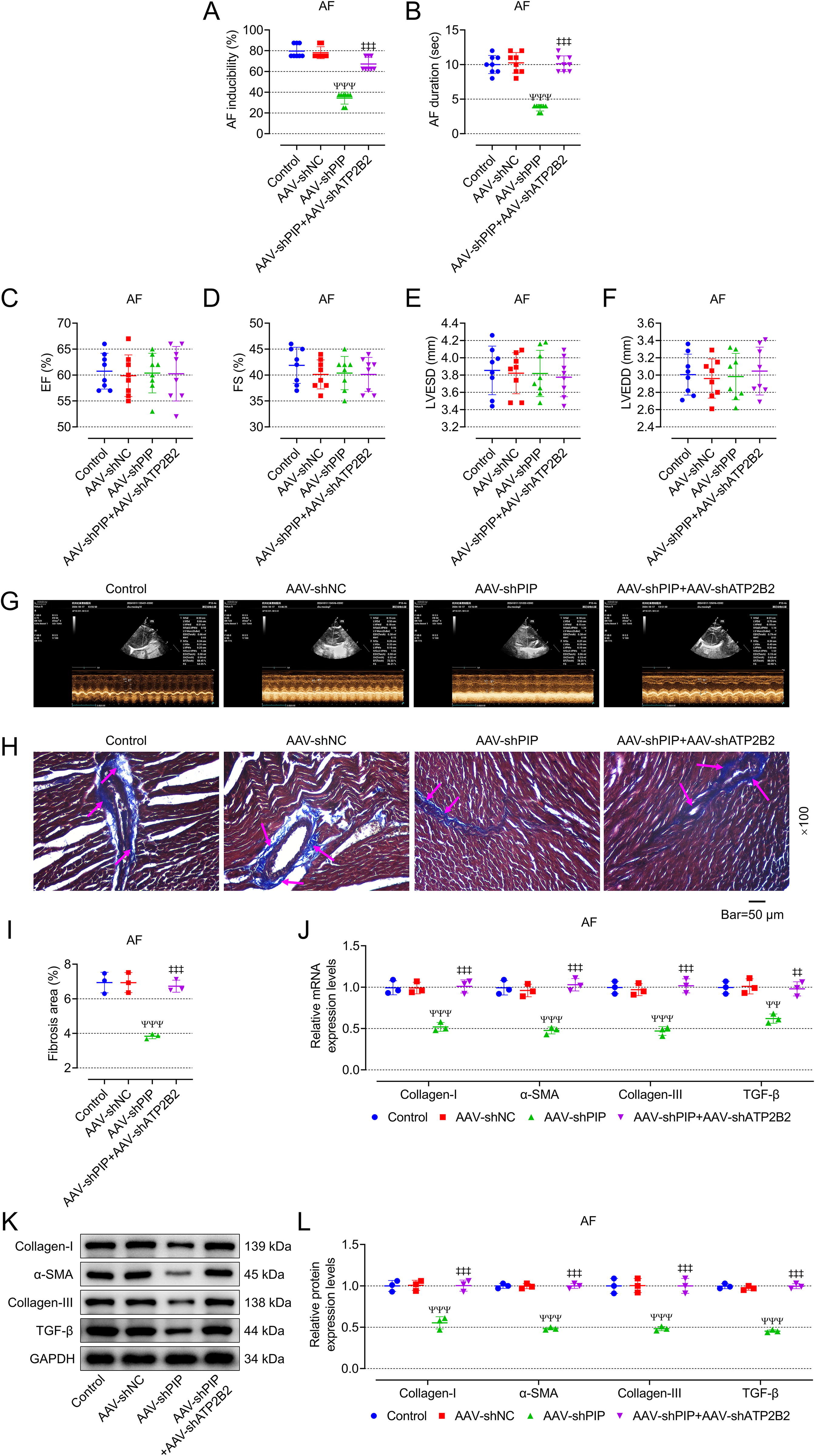
AAV-shATP2B2 mitigated the effects of AAV-shPIP on the incidence and duration of AF as well as myocardial fibrosis in AF rats
To further explore the role of PIP/ATP2B2 axis in AF in vivo, AAV-shPIP and AAV-shATP2B2 were injected into the tail vein of AF rats. Interestingly, AAV-shATP2B2 mitigated the effects of AAV-shPIP on the incidence and duration of AF as well as myocardial fibrosis in AF rats, including increasing incidence and duration of AF (Fig. 8A, B, p < 0.001), promoting myocardial fibrosis (Fig. 8H, I) and increasing the expression of genes and proteins related to myocardial fibrosis (Fig. 8J–L, p < 0.01), but it did not affect the cardiac function of AF rats (Fig. 8C–F). Representative ultrasound figures are presented (Fig. 8G). In addition, IHC showed that the addition of AAV-shATP2B2 blocked the inhibition of AAV-shPIP on the expression of α-SMA and vimentin (Fig. 9A–C, p < 0.001). Also, AAV-shATP2B2 mitigated the promotion effects of AAV-shPIP on the ratio of p-VASP/VASP and intracellular cGMP (Fig. 9D–F, p < 0.001).

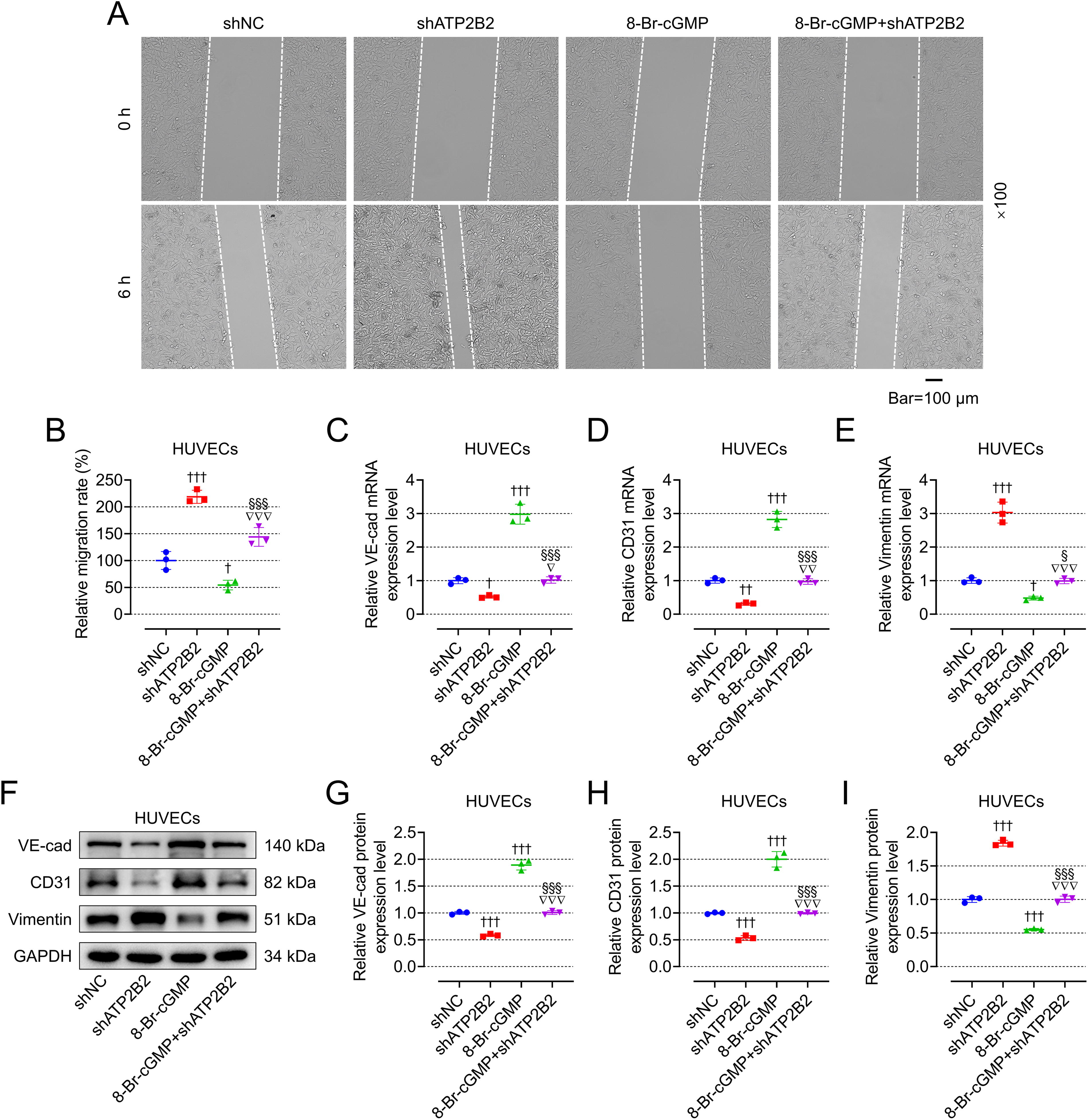
8-Br-cGMP mitigated the effects of shATP2B2 on fibrosis and viability of CFs as well as migration and EndMT of HUVECs
To verify whether ATP2B2 affects AF by regulating the cGMP-PKG signaling pathway, we transfected shATP2B2 into CFs and HUVECs and used PKG activator 8-Br-cGMP. The transfection efficiency of shATP2B2 in CFs and HUVECs is shown in Figure 10A, B (p < 0.001). Contrary to shATP2B2, 8-Br-cGMP promoted the ratio of p-VASP/VASP in CFs and HUVECs (Fig. 10C–E, p < 0.001), inhibited the expression of fibrosis-related genes and proteins in CFs (Fig. 10F–L, p < 0.05), decreased the activity of CFs (Fig. 10M, p < 0.001) and migration of HUVECs (Fig. 11A, B, p < 0.05), increased the expression of VE-cadherin and CD31, as well as decreased the expression of vimentin in HUVECs (Fig. 11C–I, p < 0.05). Moreover, 8-Br-cGMP also mitigated the regulatory effect of shATP2B2 on CFs and HUVECs (Figs. 10C–M and 11A–I, p < 0.05).

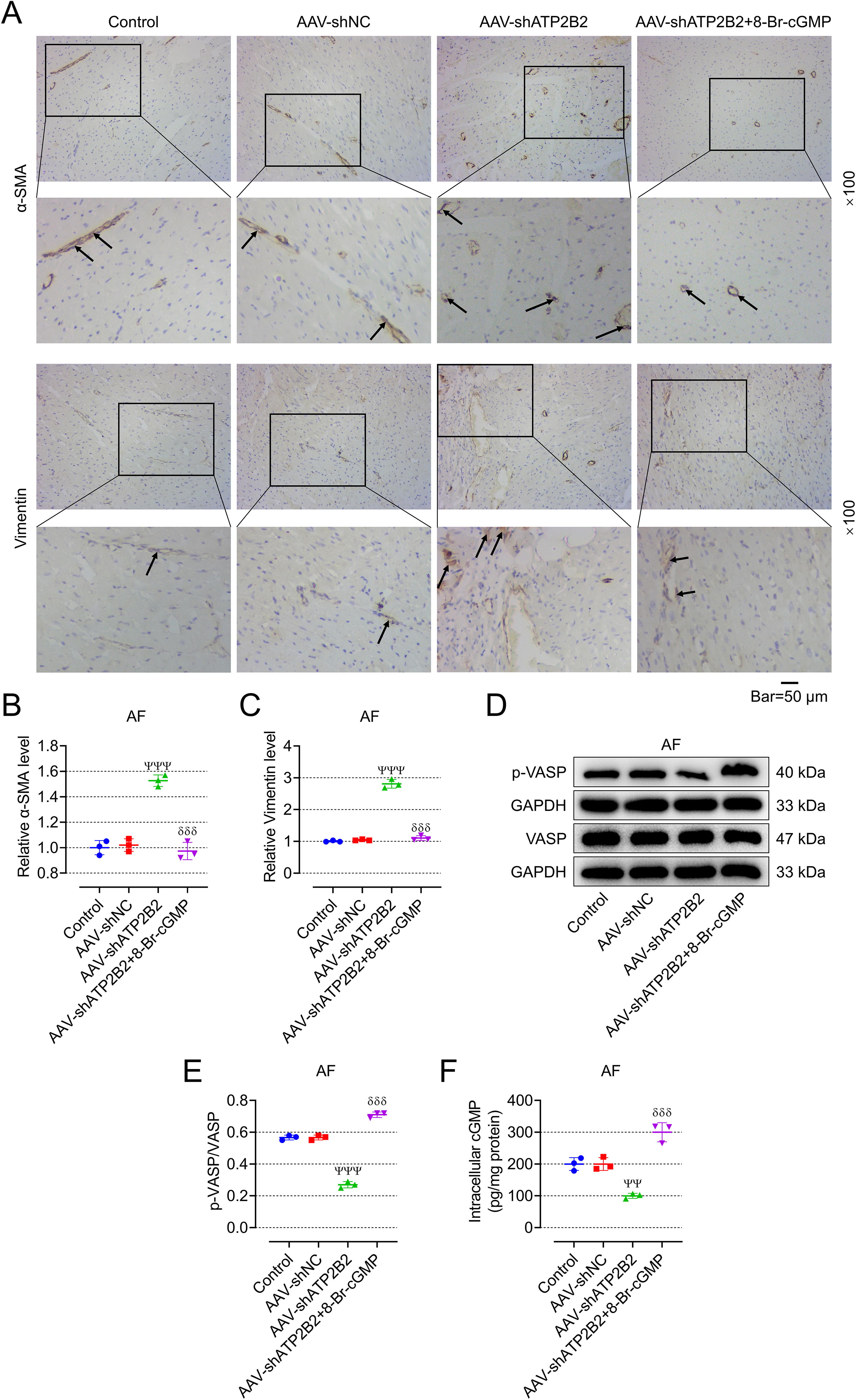
8-Br-cGMP mitigated the effects of AAV-shATP2B2 on the incidence and duration of AF as well as myocardial fibrosis in AF rats
We also used 8-Br-cGMP in AF rats to verify the regulatory effect of ATP2B2 on cGMP-PKG in vivo. As expected, 8-Br-cGMP mitigated the effects of AAV-shATP2B2 on the incidence and duration of AF as well as myocardial fibrosis in AF rats, including decreasing incidence and duration of AF (Fig. 12A, B, p < 0.001), inhibiting myocardial fibrosis (Fig. 12H–I), and decreasing the expression of genes and proteins related to myocardial fibrosis (Fig. 12J–L, p < 0.01), but it did not affect the cardiac function of AF rats (Fig. 12C–F). Representative ultrasound figures are presented (Fig. 12G). In addition, IHC showed that the addition of 8-Br-cGMP blocked the promotion of AAV-shATP2B2 on the expression of α-SMA and vimentin (Fig. 13A–C, p < 0.001). Also, 8-Br-cGMP mitigated the suppressing effects of AAV-shATP2B2 on the ratio of p-VASP/VASP and intracellular cGMP (Fig. 13D–F, p < 0.001).

Discussion
The activation and proliferation of CFs will produce excessive extracellular matrix (ECM) components, such as collagen-1 and collagen-3, and their excessive deposition will lead to the formation of scar tissue in intercellular matrix, leading to myocardial fibrosis, which has been demonstrated to be a sign of AF structural transformation (Li et al., 2022). In addition, CFs can differentiate into myofibroblasts (expressing α-SMA), which further improves the synthesis ability of collagen (Venugopal et al., 2022). Previous studies have reported that AF-exo can improve the vitality and proliferation of CFs (Zhu et al., 2022). Consistently, our study found that AF-exo promoted the vitality and differentiation of CFs, thus increasing the expression of collagen in CFs. Interestingly, transfection of shPIP into CFs can reverse the blocking effect of AF-exo, and in vivo experiments also demonstrated that AAV-shPIP can inhibit myocardial fibrosis in AF rats, which means that AF-exo may promote myocardial fibrosis by upregulating PIP expression in CFs. Previous studies have reported that PIP can bind a variety of proteins, including fibronectin, actin, β-tubulin, and integrin protein, but this interaction of PIP has not been clearly studied at present (Urbaniak et al., 2018). However, based on the important role of fibronectin in ECM synthesis, PIP may participate in ECM synthesis by interacting with fibronectin (Dalton and Lemmon, 2021).
In addition, the differentiation of endothelial cells will also have an impact on cardiac fibrosis, especially endothelial cells that have experienced EndMT are one of the sources of myofibroblasts (Lajiness and Conway, 2012). A preclinical study revealed that about 30% of fibroblasts in the heart fibrosis model were derived from endothelial cells (Zeisberg et al., 2007). EndMT can be induced by TGF-β. During the whole process, endothelial markers (VE-cadherin and CD31) gradually decreased, while the expression of mesenchymal marker vimentin gradually increased (Jordan et al., 2021). Previous studies have shown that the cardiac EndMT of AF rats increases with time (Wang et al., 2024). At the same time, EndMT was also found in the atrium of patients with AF (Kato et al., 2017). Inhibition of EndMT of HUVECs is beneficial to improve cardiac fibrosis in patients with AF (Liu et al., 2021). In our study, AF-exo can promote the migration and EndMT of HUVECs, which is consistent with the previous report. In addition, we also found that transfection of shPIP in HUVECs can block the effect of AF-exo, and in vivo experiments also demonstrated that AAV-shPIP can inhibit the expression of vimentin and TGF-β in the myocardium of AF rats, which means that AF-exo may promote the EndMT process by upregulating PIP expression in HUVECs. Previous studies have reported that the expression of migration-related genes in breast cancer cells with PIP expression increased significantly (Debily et al., 2009), and silencing PIP can inhibit invasion by regulating integrin signaling (Naderi and Meyer, 2012). As a bridge, integrin protein directly connects ECM and intracellular skeleton, which is related to the epithelial–mesenchymal transition (EMT) (Yuan et al., 2022). EndMT belongs to EMT, which means that PIP may participate in the EndMT process of HUVECs through interaction with integrin protein.
To further study the mechanism of PIP in AF, we studied the downstream gene ATP2B2 of PIP. ATP2B2 gene encodes PMCA2, which can transport calcium out of cells and maintain a low level of calcium in resting cells, and so, it plays a key role in maintaining intracellular Ca2+ homeostasis (Lopreiato et al., 2014). It is found that driving cytosolic Ca2+ into cells will promote the activation of fibroblasts and promote fibrosis (Nattel, 2017). At the same time, it is found that inhibiting the release of Ca2+ from endoplasmic reticulum will reduce the EndMT process of endothelial cells (Xu et al., 2022). In our study, oe-ATP2B2 can inhibit myocardial fibrosis and the EndMT process of HUVECs, which means that oe-ATP2B2 may maintain intracellular calcium homeostasis by transporting Ca2+ out of cells, thus preventing fibroblasts from activating and reducing the EndMT process of endothelial cells. At the same time, oe-ATP2B2 and AAV-shATP2B2 mitigated the effects of oe-PIP and AAV-shPIP on myocardial fibrosis and EndMT of HUVECs in vitro and in vivo, respectively, indicating that PIP may mediate the development of AF by inhibiting ATP2B2.
Our study found that the differential proteins in left atrial tissue of AF rats treated with shPIP and shNC were enriched in cGMP/PKG pathway. CGMP plays its physiological role through PKG, which phosphorylates many intracellular proteins. VASP is one of the substrates of PKG, and its phosphorylation at Ser239 site is a key marker to evaluate PKG activation (Yu et al., 2018). Many studies have demonstrated that cGMP/PKG pathway is related to cardiac fibrosis, and activating cGMP/PKG pathway can alleviate cardiac fibrosis (Huang et al., 2023; Qin et al., 2021). Consistently, the use of 8-Br-cGMP in our study alleviated the myocardial fibrosis and the EndMT process of HUVECs in vivo and in vitro. It is found that the cGMP/PKG pathway has complex interaction with Ca2+ signal, which may be because PKG can phosphorylate protein, such as TRPC6, which plays an important role in Ca2+ signal regulation (Rainer and Kass, 2016). Activation of cGMP/PKG pathway can protect myocardial cells from injury by regulating Ca2+ homeostasis during myocardial reperfusion injury (Inserte and Garcia-Dorado, 2015). In addition, it is found that activating cGMP/PKG pathway seems to alleviate irregular collagen metabolism and myocardial fibrosis by regulating Ca2+ influx (Zhang et al., 2021). In our study, shATP2B2 can inhibit cGMP/PKG pathway activation, while 8-Br-cGMP can reverse the inhibitory effect of shATP2B2 on cGMP/PKG pathway activation, which indicates that ATP2B2 may regulate Ca2+ homeostasis by activating cGMP/PKG pathway. In addition, the activation of cGMP/PKG pathway by shPIP was mitigated by shATP2B2, which means that PIP may promote the development of AF by inhibiting the activation of cGMP/PKG pathway mediated by ATP2B2.
Moreover, imbalances in the production of vasoactive substances such as nitric oxide (NO) by endothelial cells may lead to endothelial dysfunction and promote the vasoconstriction response associated with cardiovascular disease (Suzuki et al., 2020). The production of endodermal NO results from the activation of endothelial nitric oxide synthase (eNOS) by calcium-calmodulin through phosphatidylinositol 3-kinase (García et al., 2008). Diffusion of NO into smooth muscle cells increases intracellular cGMP levels and activates protein kinase G1, leading to inositol-triphosphate receptor-associated cGMP kinase substrate and sarcoplasmic reticulum ATPase phosphorylation, which reduces intracellular calcium flux and promotes vascular relaxation (Govers and Rabelink, 2001; Walford and Loscalzo, 2003). Previous study showed that the prolactin family hormones regulate vascular tone through NO and prostacyclin production in isolated rat aortic rings (Gonzalez et al., 2015). This reminds us that inhibition of PIP might reduce oxidative stress in the system and the protective effect involves eNOS-NO signaling, which needs more experiments to verify.
However, there are some limitations in this study. The modeling time of this study was short, and the electrocardiogram showed that AF occurred, but it may still be in the early stage, and the structure and function of the heart were basically normal, and thus, the result of ejection fraction (EF), fractional shortening (FS), left ventricular end-systolic diameter (LVESD), and left ventricular end-diastolic diameter (LVEDD) had little variation. Therefore, based on the results of this study, more comprehensive studies are needed in patients with AF who have already developed clinically impaired heart function. Besides, although no intracellular PIP expression has been reported in CFs and HUVECs, we cannot rule out the possibility that PIP/ATP2B2 axis could be the result of an as-yet unidentified intracellular PIP. Moreover, it is not clear whether PIP directly regulates ATP2B2 directly or indirectly, the details remain to be further elucidated.
In summary, our research demonstrated important role of the PIP/ATP2B2 axis in myocardial fibrosis and EndMT process of HUVECs. Our data show that AF-exo can promote myocardial fibrosis by carrying PIP to inhibit the activation of cGMP/PKG pathway mediated by ATP2B2, which means that PIP or ATP2B2 may be potential targets for the development of AF therapeutic agents.
Materials and Methods
Electronic laboratory notebook was not used.
Human sample collection
Plasma was collected from HC (n = 6) and patients with AF (n = 6) diagnosed in Rujin Hospital, Shanghai JiaoTong University School of Medicine. Fasting plasma, 10 mL, of the patients was collected in an EDTA anticoagulant tube before breakfast, and then the samples were centrifuged at 3000 g at 4°C for 10 min; the upper layer of pale-yellow plasma was collected and stored in an EP tube, labeled, and stored at −80°C. This study was approved by the ethics committee of the hospital (No.139, 2021) following the signing of an informed consent form by the selected patients. In accord with the HRS/EHRA guidelines, all participants are afflicted with paroxysmal or persistent AF (January et al., 2014). Patients suffering from infectious diseases, acute coronary syndrome, chronic/end-stage liver disease, severe hepatic and renal dysfunction, malignancies, valvular heart disease, dilated cardiomyopathy, congenital heart disease, myocarditis, and ischemic cardiomyopathy were excluded from the study.
Cell culture
HUVECs, H0173, were ordered from Anweisci (China) and cultured in DMEM complete medium (SNM-002E, Sunncell, China) supplemented with 10% fetal bovine serum (FBS) and 1% penicillin–streptomycin. Rat CFs were ordered from Sunncell (China) and cultured in CF-specific medium (SNPM-R009, Sunncell, China). Cells were placed in an incubator (51032874, Thermo Fisher, USA) at 37°C with 5% CO2. All cell lines were identified by the short tandem repeat method. Cells from generations 3–7 were selected for the experiment.
Exosome extraction, characterization, and uptake
The exosomes were separated by ultracentrifugation, as described above (Wu et al., 2019). Briefly, plasma from AF patients and HC was placed in an anticoagulant tube and centrifuged at 4°C and 3000 × g for 10 min to collect the supernatant. Subsequently, the supernatant was absorbed into a 15-mL centrifuge tube, and phosphate-buffered solution (PBS) and extraction reagent were added sequentially according to the exosome extraction kit. After mixing, the supernatant was left for 2 h at 4°C. Then, the samples were resuspended with PBS and centrifuged at 4°C and 12,000 × g for 2 min to extract the supernatant rich in exosome. Exosomes were collected and stored at −80°C. To verify the exosomes, the specific markers of the exosomes (CD63, CD9, and Tsg101) were detected by Western blot. The exosomes were labeled by PKH26 (40780ES20, YEASEN, China). In short, PKH26 is diluted with a dilute buffer, and then the diluted PKH26 (100 μM, 50 μL) is added to the exosomes. Excess dyes were bound by 10% FBS without exosomes. After centrifugation at 100,000 g for 70 min, the exosomes were resuspended with PBS (60158ES10, YEASEN, China). To visualize exosome uptake, exosomes were incubated with CFs or HUVECs at 4°C overnight, then the cells were stained with DAPI (T19827, TargetMol, China), and the images were captured by an inverted fluorescence microscope (IXplore, Olympus, Japan).
MS analysis
MS was performed as previously described (Huang et al., 2022). In short, exosomes from HC and AF patients were precipitated by DL-dithiothreitol, iodoacetamide, and 6 volumes of acetone. The precipitate collected by centrifugation was dissolved with enzymolysis diluent and then freeze-dried. After the freeze-dried samples were labeled by tandem mass tag (TMT), they were reversed-phase separated on a 1100 high-performance LC system (Agilent, USA). The separated peptide was detected by Q-Exactive mass spectrometer (IQLAAEGAAPFALGMAZR, Thermo Fisher, USA). The MS data were analyzed by MaxQuant and Proteome Discover 2.1. R software was used to generate heat map of protein differential expression.
Cell transfection
ShPIP- and ATP2B2-specific short hairpin RNA (shATP2B2) was purchased from VectorBuilder (China). The target sequence is shown in Table 2. PIP/ATP2B2 overexpression plasmids (oe-PIP/oe-ATP2B2) were constructed by inserting the full-length sequences of PIP/ATP2B2 into pcDNA3.1+ vector, and the empty vector was used as negative control (NC). Cells at their logarithmic growth phase were seeded into the 96-well plate at 2 × 104 cells/well and left for 24 h at 37°C with 5% CO2 to reach 80% confluency. With the help of transfection reagent (CBMD250, Abace-biology, China), the plasmid was transfected into cells by the liposome transfection technology, and then, the transfection efficiency was evaluated by real-time quantitative polymerase chain reaction (RT-qPCR).
Target Sequences for shRNA Used in This Study
PIP, prolactin induced protein; ATP2B2, ATPase plasma membrane Ca2+ transporting 2.
Real-time quantitative polymerase chain reaction
RNA extraction kit (NP06011, NucleoTech, China) was applied to extract and detect total RNA from cells or left atrial tissue. The cDNA created by 1 μg RNA using the Reverse transcription kit (NN01012, NucleoTech, China), SYBR green kit (NL02021, NucleoTech, China) was used for RT-qPCR. Reaction conditions: predenaturation at 95°C for 10 min and denaturation at 95°C for 15 s; The elongation was extended by annealing at 60°C for 1 min and expanded by 45 cycles. Table 3 lists the sequences of specific primers. The target genes’ expression was normalized to that of glyceraldehyde-3-phosphate dehydrogenase (GAPDH) using the 2-ΔΔCT formula.
Primers Used in This Study
GAPDH, glyceraldehyde-3-phosphate dehydrogenase.
Cell treatments
To verify the effect of exosomal PIP on HUVECs or CFs, HUVECs or CFs were transfected with shNC or shPIP, and then cocultured with exosomes derived from AF patients. To verify the effect of PIP/ATP2B2 axis on HUVECs or CFs, HUVECs or CFs were cotransfected with oe-PIP/NC and oe-ATP2B2/NC. To verify whether ATP2B2 regulates AF by activating PKG pathway, HUVECs or CFs were transfected with shATP2B2 and incubated with PKG activator 8-Br-cGMP (0.3 mM, HY-135110, MedChemExpress, China).
Cell viability
MTT kit (22768, xabiolite, China) was used to determine cell viability. In short, after 96 wells were inoculated with cells (3000 per well), MTT working solution was added to the cells and incubated with them for 2 h. The absorbance at 560 nm was determined by microplate reader (CMax Plus, Molecular devices, China).
Wound healing assay
Scratches were made on the monolayer of HUVECs with a pipette, and the scraped cells were removed with PBS. The migration area of cells was measured immediately after scratch and 6 h later. The ratio of the migration area after 6 h to the initial scratch area is regarded as the mobility of HUVEC.
Animals
Six-week-old male, 200–250 g, Sprague Dawley (SD) rats (n = 96) were bought from Hangzhou Medical College. Rats were maintained in a room with a natural circadian alternation, 50% humidity, and a constant temperature of 21 ± 0.5°C. According to the China Council on Animal Care and Use rules, all experiments in this study were approved by the Institutional Animal Care and Use Committee, ZJCLA (Certificate No. ZJCLA-IACUC-20010435).
The animal experiment was divided into third parts. In the first part, 32 rats were randomly divided into four groups: sham group, AF group, adeno-associated virus (AAV)-shNC group, and AAV-shPIP group. Sham group rats were injected with 0.9% saline (1 mL/kg) through tail vein every day for 7 days. Rats in the other three groups were injected with acetylcholine (ACH)-CaCl2 (60 μg/mL ACH and 10 mg/mL CaCl2) (1 mL/kg) daily through the tail vein for 7 days (Lv et al., 2019). The last two groups of rats were injected with AAV-shNC or AAV-shPIP (2 × 1011 vg/rat) via the tail vein 14 days before the injection of ACH-CaCl2. ACH (PHR1546) and CaCl2 (C4901) were purchased from Merck (Germany). In the second part, 32 rats were randomly divided into four groups: Control group, AAV-shNC group, AAV-shPIP group, and AAV-shPIP+AAV-shATP2B2 group. AAV-shPIP+AAV-shATP2B2 group was injected with AAV-shPIP+AAV-shATP2B2 (2 × 1011 vg/rat) via the tail vein 14 days before the injection of ACH-CaCl2. In the third part, 32 rats were randomly divided into four groups: control group, AF+AAV-shNC group, AF+AAV-shATP2B2 group, and AF+AAV-shATP2B2 + 8-Br-cGMP group. AF+AAV-shATP2B2 + 8-Br-cGMP group was injected with AAV-shATP2B2 (2 × 1011 vg/rat)+8-Br-cGMP (4.8 μg/kg/day) via the tail vein 14 days before the injection of ACH-CaCl2. AAV-shNC, AAV-shPIP (target sequence: 5′-TGGTCAAAGCTTACCTTATAA-3′), and AAV-shATP2B2 (target sequence: 5′-CCAAGAGAGACAUUCACAUTT-3′) were ordered from VectorBuilder (China). The rats were anesthetized by isoflurane (2%) and euthanized by cervical dislocation. The heart tissue was collected for the next experimental analysis.
Proteomics
The left atrial tissue of rat treated by AAV-shNC and AAV-shPIP was collected, and then the protein was extracted from left atrial tissue and quantified by BCA, followed by enzymolysis samples, TMT labeling, and reverse-phase classification. Then LC-MS analysis was carried out according to the MS method mentioned above. GO and KEGG databases were applied to annotate protein.
Electrocardiogram recording
Before the animals were euthanized, rats were anesthetized with isoflurane (2%), and then limb leads of 12-lead ECG were inserted into four limbs of animals, and the inducibility and duration of AF were recorded (Lv et al., 2019). The ECG acquisition time was 5 min. AF inducibility (%) = Number of rats with atrial FA/total number of rats; AF duration (sec) = the interval from initiation to spontaneous AF termination.
Echocardiography
Before the rats were euthanized, the rats in each group were anesthetized with isoflurane (2%) and the hair on the chest wall of the rats was shaved with scissors and surgical blades. Then, the rats were examined by Vevo®3100 microultrasound imaging system (FUJIFILM VisualSonics Inc., Canada). The frequency is 8–12 MHz, the detection depth is 4 cm, and the detection angle is 90° (Lv et al., 2019). The images were obtained in M-mode, and the EF, FS, LVESD, and LVEDD of rats were calculated. The mean value of three cardiac cycles was used for statistical analysis.
Masson staining
The hearts of the rats were removed under anesthesia and the left atrial tissue was isolated and placed in 4% formaldehyde fixation solution (20010, xabiolite, China), and then dehydrated step by step by gradient ethanol. After paraffin embedding and slicing, the left atrial tissue was dewaxed by xylene and hydrated by gradient ethanol. The slices were stained with Masson staining kit (G1340, Solarbio, China) according to the manufacturer’s instructions. The results were observed by microscope (BX53F, Olympus, Japan). Also, three visual fields were tested in each sample. Results were analyzed with Image-Pro 6.0 software (Media Cybernetics, Bethesda, USA). Fibrosis area (%) = fibrosis area/total area × 100%.
Immunohistochemistry
The left atrial tissue slices prepared above were first subjected to microwave antigen repair after dewaxing and hydration, and then BSA (GC35542, GLPBIO, China) was used to block specific antigens. Next, slices were incubated with anti α-SMA antibody (AF1032, Affinity, USA) and anti-vimentin antibody (AF7013, Affinity, USA), and then continued to be incubated with Horseradish peroxidase (HRP)-labeled secondary antibodies (ab205718, Abcam, UK). Coloring was performed using DAB (36201ES03, YEASEN, China), followed by restaining with hematoxylin (C0107, Beyotime, China). The slices were then sealed with neutral resin (36313ES60, YEASEN, China) and visualized under a microscope. In addition, three visual fields were tested randomly in each sample. Results were analyzed with Image-Pro 6.0 software (Media Cybernetics, Bethesda, USA).
Western blot
After the protein was extracted using RIPA buffer (PK10020, Proteintech, China), the total protein was quantified using the Protein Quantitative Kit (SK1070, Coolaber, China). The proteins (15 μg) were loaded onto SDS-PAGE gels (PH1493, Phygene, China), and then transferred to polyvinylidene fluoride membranes (FFP24, Beyotime, China). The membranes were blocked with BSA and then incubated for a whole night at 4°C with the primary antibody (Table 4). After that, the membranes were left to incubate for 2 h at room temperature with the secondary antibody (Table 4). Finally, the membranes were identified using an iBright FL1500 Imaging System (A44115, Invitrogen, USA) following their exposure to the enhanced chemiluminescence (ECL) Western Blotting Substrate (MF078, mei5bio, China). GAPDH was the internal control used in this experiment.
Antibodies Used in This Study
P-VASP, phosphor-VASP.
Measurement of cGMP
The left atrial tissue was washed with PBS, then cut and ground in a homogenizer. The homogenate was centrifuged at 1500 g for 10 min to obtain the supernatant. Then the cGMP level in the supernatant was detected by the ELISA kit (E-EL-0083, Elabscience, China) according to the manufacturer’s instructions, with interassay and intra-assay coefficient of variation being <10%, limit of quantification: 0.47 pmol/mL.
Statistical analysis
Statistical analysis was experimented by GraphPad Prism 8.0. The measurement data were expressed by mean ± standard deviation. One-way or two-way ANOVA was used for multiple group comparisons. Independent-sample t test was used to compare the two groups of data. The Shapiro–Wilk test was used for normality test and the Levene test was used for homogeneity of variance test. All data conformed to normality and homogeneity of variance. Differences with p < 0.05 were considered statistically significant.
Footnotes
Authors’ Contributions
Substantial contributions to conception and design: Y.W. Data acquisition, data analysis, and interpretation: X.L., Z.M.S., J.M.L., G.H.W., T.J.Z., C.J.L., Y.X., Y.Y.B., Q.Z.L., T.Y.L., W.Q.P., Y.C.X., N.Z., Q.J., and L.Q.W. Drafting the article or critically revising it for important intellectual content: All authors. Final approval of the version to be published: All authors. Agreement to be accountable for all aspects of the work in ensuring that questions related to the accuracy or integrity of the work are appropriately investigated and resolved: All authors.
Author Disclosure Statement
The authors declare no conflicts of interest.
Funding Information
This work was supported by the Research Project of Shanghai Municipal Health Commission [20224Y0028] and the National Natural Science Foundation of China [81870250, 81900290, and 82100329].
Supplementary Material
Supplementary Figure S1
Supplementary Figure S2
Supplementary Figure S3
Supplementary Figure S4
Supplementary Figure S5
Abbreviations Used
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.