
Abstract
Aims:
In Friedreich ataxia (FRDA), early motor discoordination stems from dysfunctional sensory neurons in the spinal cord driven by epigenetic dysregulation, frataxin (FXN) deficiency, oxidative stress, and inflammation. Omaveloxolone, a nuclear factor erythroid 2-related factor-2 (NRF2) inducer, is the only treatment available. In various chronic disease models, sulforaphane (SF) can target NRF2 and the above processes. This study compared the effects of SF with omaveloxolone and dimethyl fumarate (DMF) in sensory neurons generated from FRDA patient-induced pluripotent stem cells and their isogenic control.
Results:
The successful generation of the FRDA and isogenic control sensory neurons was confirmed by the positive expression of β-III TUBULIN, BRN3A, ISLET1, PERIPHERIN, and tropomyosin receptor kinase C. In comparison with the isogenic control, FRDA sensory neurons displayed an aberrant gene expression profile alike to that reported in patients. None of the drugs affected the viability of the isogenic control sensory neurons. SF treatment improved the viability of FRDA sensory neurons by up to 61% versus the untreated control. DMF treatment showed a modest 35% increase, while omaveloxolone lacked an effect. SF-treated FRDA sensory neurons demonstrated increased reduced glutathione/oxidized glutathione ratio and expression of FXN and redox markers, and a reduced expression of selected epigenetic enzymes and inflammatory cytokines, at the respective gene and protein levels. DMF and omaveloxolone treatments only modulated some of these biomarkers.
Innovation:
We revealed the therapeutic potential of SF and how it performs in comparison with omaveloxolone and DMF, in a physiologically and genetically relevant in vitro FRDA model.
Conclusion:
SF offers a multipronged approach to alleviating the different cellular events underlying FRDA. Antioxid. Redox Signal. 43, 308–327.
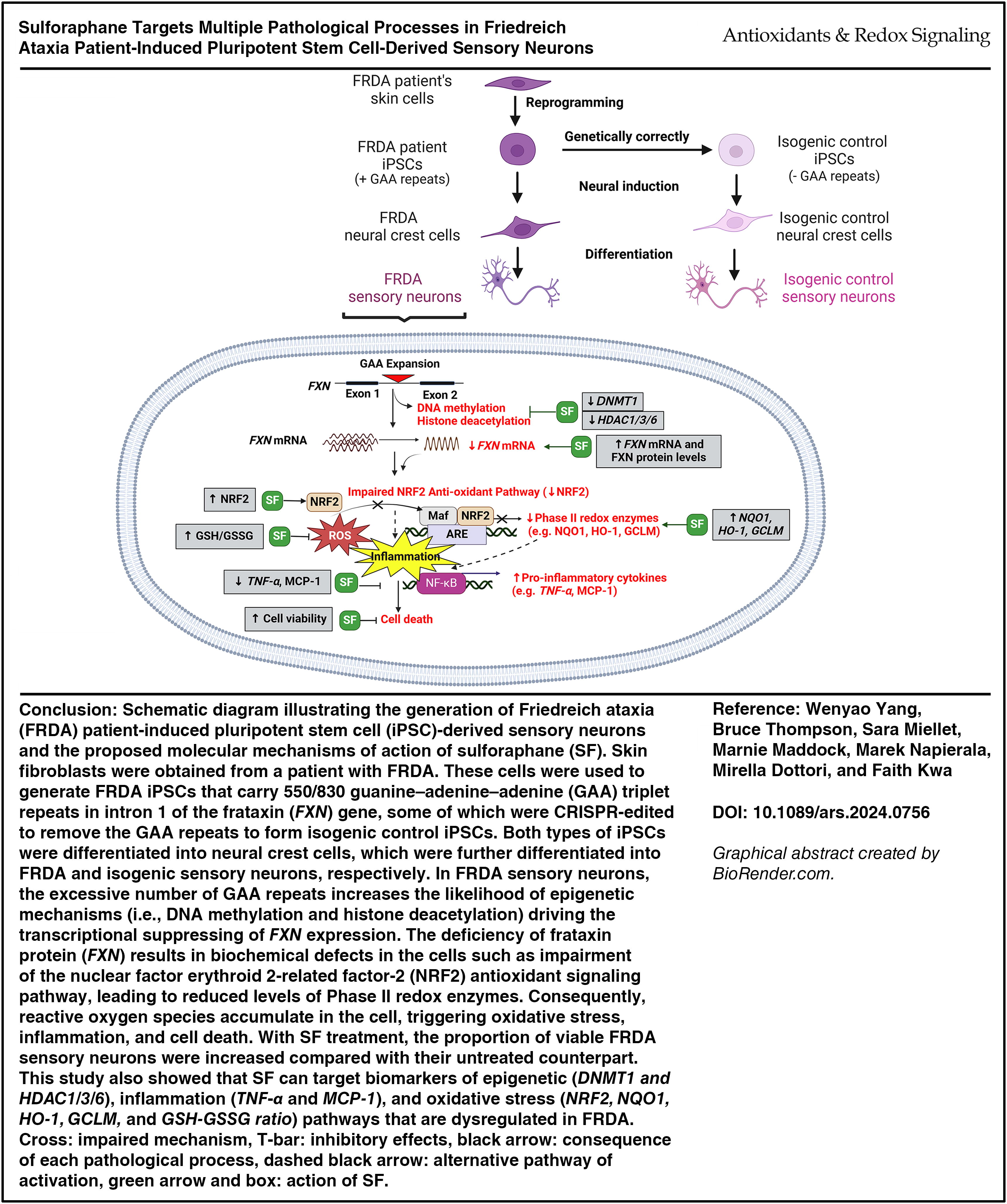
Introduction
Friedreich ataxia (FRDA) is the most prevalent inherited form of ataxia (Campuzano et al., 1996), arising from a guanine–adenine–adenine (GAA) (Mayer et al., 2024) expansion in intron 1 of the frataxin gene (FXN) on chromosome 9 (Al-Mahdawi et al., 2008). The FXN gene encodes a highly conserved frataxin protein (FXN) essential for cell metabolism. The GAA expansion leads to transcriptional suppression of this gene in various cell types, with peripheral sensory neurons originating from the dorsal root ganglia (DRG) being a key cell type affected and the subsequent neuronal loss can lead to progressive motor symptoms seen in patients (Tiberi et al., 2023). Generally, affected individuals are found to carry 70 to 1700 GAA repeats, whereas unaffected individuals have up to 36 repeats (Silva et al., 2015) The length of the GAA repeats is positively correlated with the severity of FRDA symptoms and disease progression but inversely correlated with the age of disease onset (Al-Mahdawi et al., 2006). The transcriptional suppression of the FXN triggered by the GAA expansion involves the epigenetic processes of DNA methylation and histone deacetylation, which are regulated by histone deacetylases (HDACs) and DNA methyltransferases (DNMTs), respectively (Soragni and Gottesfeld, 2016). HDAC1 and HDAC3 have been suggested as targets for reversing silenced FXN gene expression, while HDAC6 is a common target in neurogenerative diseases and increased DNMT1 levels have been observed in cells from patients with FRDA (Quesada et al., 2015; Simões-Pires et al., 2013; Soragni and Gottesfeld, 2016). Moreover, studies have demonstrated heterochromatin formation and hypermethylation of specific CpG islands upstream of the GAA repeat sequence in the FXN gene in FRDA primary cells and mouse models but absent in the respective control groups (Al-Mahdawi et al., 2008). The subsequent pathological consequences of FXN deficiency in patient cells include mitochondrial dysfunction, impaired nuclear factor erythroid 2-related factor 2 (NRF2) antioxidant signaling pathway, and inflammation, thereby ultimately compromising cell viability, which gives rise to various clinical symptoms and shortened life expectancy if left untreated (Chiang et al., 2020; Nachun et al., 2018).
Currently, omaveloxolone (Omav), a potent NRF2 activator, is the only available treatment approved by the U.S. Food and Drug Administration for patients with FRDA aged 16 or older in the United States and countries in the European Union (Lee, 2023). However, this drug was not designed to correct FXN deficiency and for most patients with FRDA who reside in countries where Omav is unavailable, comprehensive disease management strategies remain the mainstay treatment for improving quality of life (Yang et al., 2022). Consequently, there is a need to identify alternative compounds that can address the pathological processes both upstream and downstream of FXN deficiency, which drives the multisystemic symptoms at different stages of FRDA. One such therapeutic agent that can target the same NRF2 antioxidant pathway as Omav but with the potential to exert both epigenetic and anti-inflammatory effects is sulforaphane (SF) (Santin-Marquez et al., 2019).
SF, derived from glucoraphanin, is an isothiocyanate compound abundant in cruciferous vegetables (Santin-Marquez et al., 2019). Its small molecular size and lipophilic nature enable it to exhibit high bioavailability and penetrate DRG sensory neurons and traverse the blood–brain barrier (BBB) to reach the central nervous system (CNS), thereby effectively exerting its neuroprotective effects (Houghton, 2019; Jazwa et al., 2011; Sun et al., 2017; Yang et al., 2018b). As a dietary phytochemical, SF exhibits good tolerability in clinical trials (Mangla et al., 2021; Mazarakis et al., 2020). Daily consumption of broccoli sprout extract containing SF for up to 12 weeks demonstrated antioxidant and detoxification benefits in healthy individuals with no adverse events recorded (Mazarakis et al., 2020). More importantly, SF has gained considerable attention as a therapeutic agent as it can target various molecular pathways regulating epigenetic mechanisms, oxidative stress and inflammation, all of which play significant roles in the pathogenesis of FRDA (Santin-Marquez et al., 2019; Yang et al., 2022).
Studies have demonstrated that SF can inhibit HDAC and DNMT activities in various human cancer cell types (Kaufman-Szymczyk et al., 2015). SF at micromolar doses was found to exhibit an inhibitory effect on the total HDAC activities by upregulating the expression of genes involved in cell cycle and apoptosis (i.e., P53, P21, and BAX) in human lung cancer cells (Jiang et al., 2016). Other studies showed that SF treatment led to reductions in DNMT activity (DNMT1/3a/3b) and a subsequent increase in the expression of tumor suppressor genes, thereby rendering cancer cells more susceptible to apoptosis (Ali Khan et al., 2015; Meeran et al., 2010). Thus, SF may hold promise in reversing the transcriptional silencing of the FXN gene by downregulating chromatin-modifying enzymes that are affected in patients with FRDA. In neurodegenerative conditions such as Alzheimer’s disease, SF has been found to mitigate DNMT-mediated DNA hypermethylation at the promoter region of the NRF2 gene, thereby increasing gene expression and subsequently reducing neurological inflammation (Saleh et al., 2021; Zhao et al., 2018). This suggests that the epigenetic modulatory capability of SF may contribute to its antioxidant and anti-inflammatory properties, which may drive its neuroprotective effects.
As a potent NRF2 inducer, SF has been investigated in a variety of chronic diseases (Du et al., 2022; Folbergrova et al., 2023; Kwa et al., 2023). It can interact with Kelch-like ECH-associated protein 1 to promote NRF2 nucleus translocation and subsequent binding to the cis-acting enhancer sequence antioxidant response element in the upstream promoter region of the target genes encoding redox enzymes and activating their expression. These enzymes include NADPH quinone oxidoreductase-1 (NQO1), heme oxygenase-1 (HO-1), and glutamate–cysteine ligase modifier subunit (GCLM). In addition, SF can suppress the expression genes encoding nuclear factor kappa B proinflammatory mediators such as tumor necrosis factor-alpha (TNF-α) and monocyte chemoattractant protein-1 (MCP-1) (Saleh et al., 2021; Yang et al., 2022). In FRDA patients' cells, the NRF2 signaling pathway is impaired, which is reflected by decreased expression levels of NRF2, NQO1, HO-1, GCL, and reduced ratios of reduced form of glutathione (GSH) and oxidized form of glutathione (GSSG) (Carletti and Piemonte, 2014; Chiang et al., 2020). A proinflammatory transcriptomic signature has previously been established in peripheral blood cells from patients with FRDA, along with increased expression levels of TNF-α (Bolotta et al., 2020; Nachun et al., 2018). This emphasizes the need for drugs to ameliorate oxidative stress and inflammatory pathways in affected cells.
To date, SF has not yet been tested in clinical trials for FRDA. Moreover, the epigenetic and anti-inflammatory effects of SF have not been reported in preclinical models of FRDA. Although some studies have shown the ability of SF to increase FXN expression levels, these studies were limited by the use of cellular models that are not physiologically relevant or do not fully represent the aberrant genetic profile of patients with FRDA (Abeti et al., 2015; La Rosa et al., 2019; Petrillo et al., 2019; Petrillo et al., 2017). In vitro models involving FRDA patient fibroblasts and lymphoblastoid cells are not the key cell types affected by progressive loss of viability despite harboring the inherited GAA abnormality (La Rosa et al., 2021; Schreiber et al., 2019). Animal models used to screen the effects of SF include those generated by knocking in a mouse FXN gene with 230 GAA repeats into KIKO mice or by injecting YG8R mice with a humanized transgene containing 90/190 GAA repeats. These models display only mild disease symptoms attributed to the inherent instability of the introduced GAA repeats (Abeti et al., 2015; La Rosa et al., 2019; Schreiber et al., 2019). Therefore, it is imperative to screen the potential therapeutic impact of SF in reliable FRDA cellular models that exhibit both of the following features: (1) Characteristic of the predominant cell type affected by neurodegeneration (i.e., DRG sensory neurons) and (2) fully recapitulate the genetic hallmarks of FRDA (i.e., pathological number of GAA repeats) (Alshawaf et al., 2018; Hulme et al., 2020; Schreiber et al., 2019). Here, our novel study is the first to compare the therapeutic efficacy and molecular mechanism-of-action of SF with two other NRF2 activators, Omav and DMF, using sensory neurons derived from FRDA patient-induced pluripotent stem cell (iPSC) carrying 550/830 GAA repeats on each of the alleles and its isogenic control line that have been CRISPR-edited to eliminate the repeats (Ku et al., 2010; Miellet et al., 2024a; Miellet et al., 2024b) (see Graphical Abstract). This study has shown that SF can improve the viability of patient sensory neurons by promoting the gene and protein expression levels of FXN and redox enzymes of the NRF2 pathway while inhibiting the expression of genes targeting histone deacetylation, DNA methylation, and markers of inflammation.
Results
Generation of FRDA and isogenic control sensory neurons
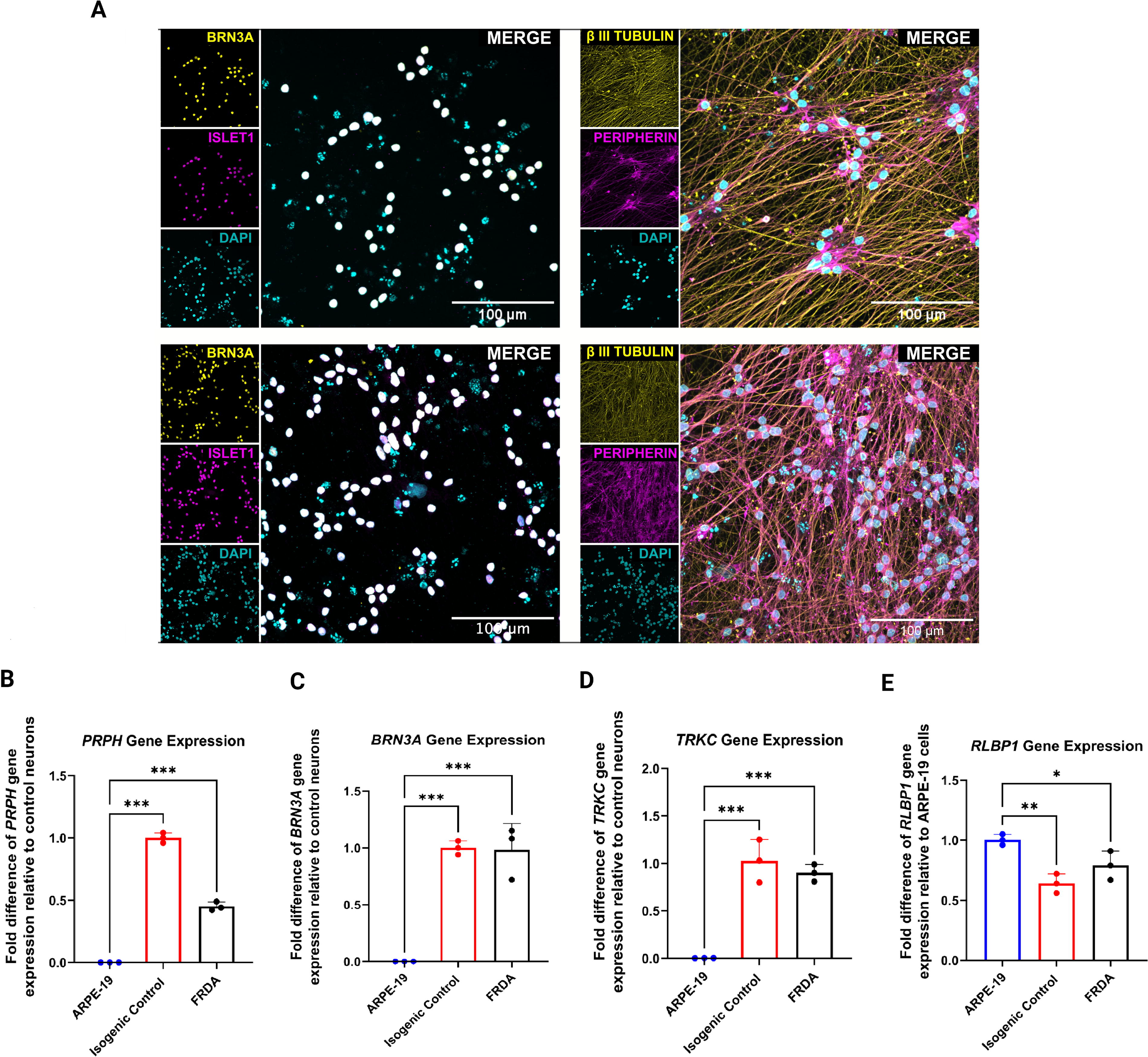
Genetically modified FRDA iPSC lines that carry an inducible neurogenin 2 (NGN2 expression cassette with an mApple gene) were used to generate FRDA and isogenic control sensory neurons (Miellet et al., 2024b). Positive mApple expression was observed throughout the culture of iPSCs (Supplementary Fig. S1), indicating the stability of the NGN2 gene construction that is required for the differentiation of neural crest cells (NCCs) into sensory neurons upon doxycycline induction. Following neural induction of the iPSCs, the NCCs formed distinctive two-dimensional morphologies characteristic of neural crest features (Supplementary Fig. S1) (Kobayashi et al., 2020). These cells were further differentiated into mature sensory neurons through a 3-day doxycycline treatment to activate NGN2 expression (Miellet et al., 2024b). Approximately 2 weeks from the first day of doxycycline treatment, both the isogenic control and patient sensory neurons were observed to develop distinguishable neuronal processes (Supplementary Fig. S1). These sensory neurons also expressed the pan-neuronal marker β-III TUBULIN, as well as sensory neuronal markers BRN3A, ISLET1, and PERIPHERIN (Fig. 1A). Quantitative real-time polymerase chain reaction (qRT-PCR) analyses further confirmed the gene expression profiles of DRG sensory neuronal markers on Day 14 postdoxycycline induction, evidenced by positive gene expression of PRPH, BRN3A, and tropomyosin receptor kinase C (TRKC) (all p-values <0.001; Fig. 1B–D). Human retinal ARPE-19 cells, which served as the negative control, exhibited positive gene expression of the retinal cell marker retinaldehyde binding protein 1 (RLBP1) but showed null expression for the said sensory neuronal markers (Fig. 1).
FRDA sensory neurons exhibit aberrant expression of frataxin and markers of pathological processes driving the disease
To further validate whether the FRDA sensory neurons exhibit disease hallmarks such as aberrant expression levels of FXN and markers of pathways regulating epigenetic mechanisms, oxidative stress, and inflammation, gene expression profiles of the FRDA sensory neurons were compared with that of the isogenic control sensory neurons. FRDA sensory neurons showed a 5.8-fold and 2.5-fold reduction in FXN gene and FXN protein expression levels, respectively, compared with isogenic control sensory neurons (both p-values <0.001; Fig. 2A, B). The FRDA sensory neurons had decreased levels of the antioxidant markers when compared with the isogenic control (Fig. 2C–F). Specifically, a two-fold and 2.1-fold reduction were found in NRF2 (p < 0.001) and HO-1 (p < 0.05) mRNA levels, respectively. Similarly, the gene expression levels of NQO1 (p < 0.001) and GCLM (p < 0.05) were 1.28-fold and 1.47-fold lower in the FRDA sensory neurons compared with the isogenic control, respectively. TNF-α gene expression was elevated by 1.5-fold in the FRDA sensory neurons compared with the isogenic control (p < 0.001; Fig. 2G). The MCP-1 protein expression level was also found to be 13-fold higher in the FRDA sensory neurons compared with the isogenic control (p < 0.001; Fig. 2H). The gene expression levels of HDAC1, HDAC3, HDAC6, and DNMT1 were all upregulated in FRDA sensory neurons by approximately 1.5-fold compared with the isogenic control (all p-values <0.05; Fig. 2I–L). These findings, summarized in Table 1, provide evidence that the FRDA sensory neurons used in this study exhibit abnormal gene expression of the various molecular markers of FRDA, validating it as a reliable disease model.

Effects of Sulforaphane, Dimethyl Fumarate, and Omaveloxolone on Friedreich Ataxia Patient-Induced Pluripotent Stem Cell-Derived Sensory Neurons
Data are presented as an average change in gene expression from three independent experiments, each with three technical replicates.
p < 0.05, **p < 0.01, ***p < 0.001.
DMF, dimethyl fumarate; DNMT, DNA methyltransferase; FRDA, Friedreich ataxia; FXN, frataxin; GCLM, glutamate–cysteine ligase modifier subunit; HDAC, histone deacetylase, HO-1, heme oxygenase 1; iPSC, induced pluripotent stem cell; NQO1, NADPH quinone oxidoreductase 1; NRF2, nuclear factor erythroid 2-related factor-2; Omav, omaveloxolone; SF, sulforaphane; TNF-α, tumor necrosis factor-alpha.
SF treatment increases the proportion of viable FRDA sensory neurons
Following the validation of the cell model, the effects of SF, DMF, and Omav on cell viability were investigated using the MTS assay. None of the drugs, including 0.1% dimethyl sulfoxide (DMSO), significantly reduced the cell viability of the isogenic control and FRDA sensory neurons after 24 hour of exposure (all p-values >0.05; Fig. 3A, B). However, FRDA sensory neurons treated with 20 μM SF significantly increased the proportion of viable cells by 61% compared with the untreated control (p = 0.039). At lower concentrations such as 100 nM (p = 0.049) and 500 nM SF (p = 0.024), a notable 10% and 18% increase in the proportion of viable cells was observed, respectively. The positive drug control DMF also demonstrated an improvement in cell viability by 35% (p = 0.041). However, Omav at the tested nanomolar concentrations did not have any effects on cell viability.

SF treatment increases frataxin gene and protein expression levels in FRDA sensory neurons
To investigate whether the improvement in the viability of FRDA sensory neurons by SF is consistent with an upregulation of frataxin expression, FXN gene and protein expression levels were measured. It was found that treatment with SF at either 100 nM (p = 0.009) or 10 μM (p = 0.013) significantly increased FXN mRNA levels by 1.4-fold (Fig. 4A). Although 500 nM SF appeared to increase the FXN mRNA levels, this was not statistically significant (p = 0.052). At the protein expression level, a 1.58-fold increase in FXN protein expression levels with 100 nM SF treatment (p = 0.003) was observed (Fig. 4B). Meanwhile, SF concentrations at 5 μM (p = 0.005) and 10 μM (p < 0.001) significantly increased FXN protein expression levels by 1.55-fold and 1.7-fold, respectively. None of the positive control drugs was found to increase FXN expression levels (all p-values >0.05).

SF treatment inhibits the gene expression of epigenetic enzymes in FRDA sensory neurons
To determine the epigenetic modulatory effects of SF, changes in the gene expression levels of HDAC1, HDAC3, and HDAC6 and DNMT1 were examined. A consistent 1.28-fold reduction in HDAC1 mRNA levels was observed following treatment with SF at concentrations of 500 nM SF, 10 µM SF, or 20 µM (all p-values <0.05; Fig. 5A). Treatment with DMF at 30 µM (p = 0.04) exerted the same effect. In contrast, despite various concentrations of Omav appearing to decrease HDAC1 mRNA levels, none of the observations was statistically significant (all p-values >0.05). SF at 500 nM, 5 µM, and 20 µM was shown to downregulate the HDAC3 gene expression by 1.27-fold, 1.43-fold, and 1.28-fold, respectively (all p-values <0.05; Fig. 5B). Similarly, treatments with 50 nM Omav (p = 0.002) and 100 nM Omav (p < 0.001) suppressed HDAC3 gene expression levels by 1.5-fold and 1.59-fold. A total of 20 µM of SF (p < 0.001), 30 µM of DMF (p = 0.008), and 50 nM of Omav (p = 0.04) downregulated HDAC6 mRNA levels (Fig. 5C). However, SF having triggered a 2.5-fold reduction in HDAC6 expression seemed to have a more potent effect compared with the other two drugs (DMF: 1.4-fold; Omav: 1.3-fold). A dose-dependent effect was observed when the FRDA sensory neurons were treated with 5 µM SF (p = 0.04), 10 µM SF (p = 0.03), or 20 µM SF (p < 0.001), resulting in a 1.5-fold to 2.3-fold reduction in DNMT1 gene expression levels compared with untreated cells (Fig. 5D). In addition, treatment with 10 nM or 50 nM Omav decreased DNMT1 gene expression (both p-values <0.05). However, DMF had no effect on DNMT1 mRNA levels.

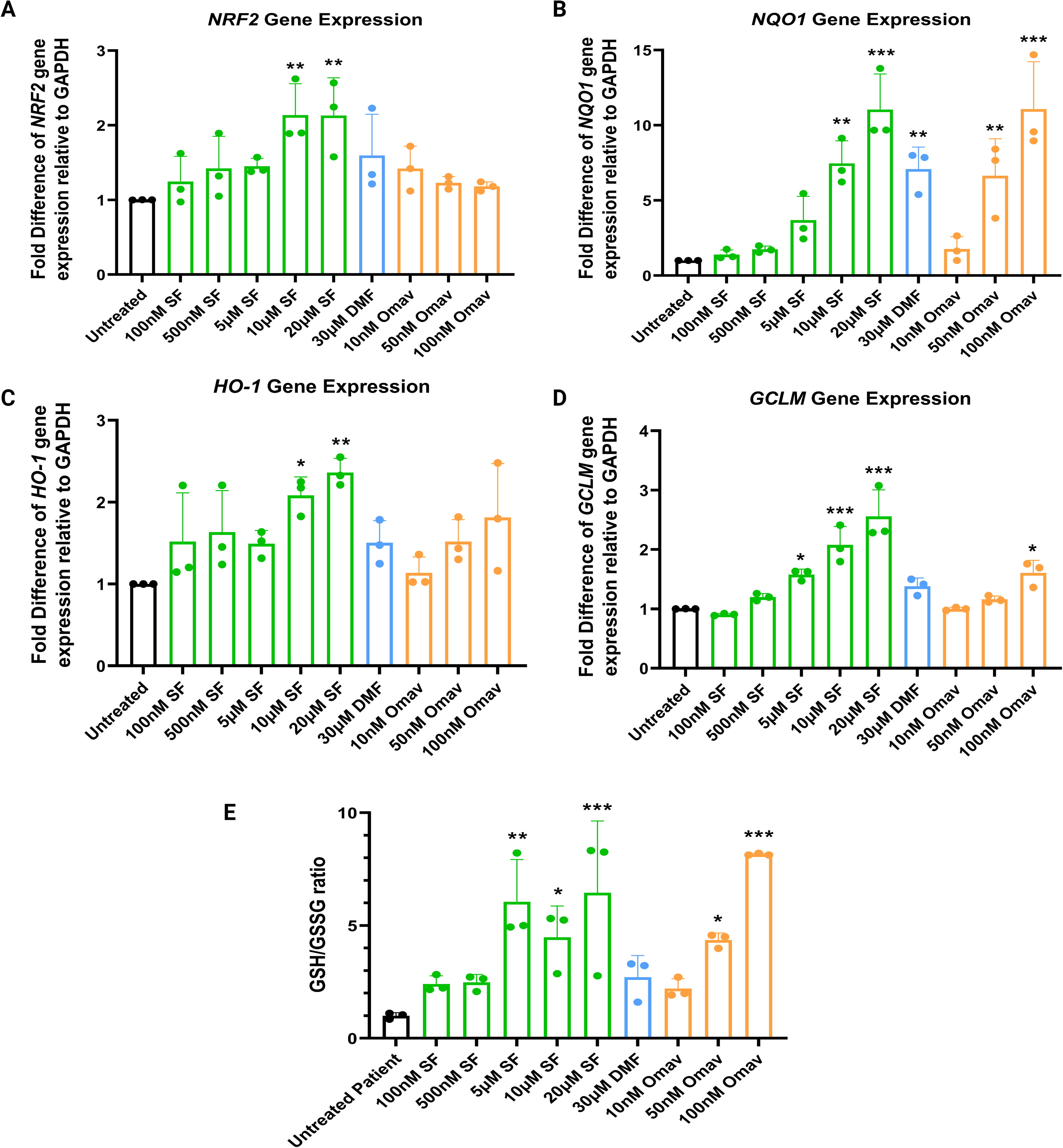
SF enhances the gene expression of redox factors and increases the GSH/GSSG ratio in FRDA sensory neurons
The renowned antioxidant effects of SF have been reaffirmed in this novel FRDA model. SF at 10 µM or 20 µM significantly boosted NRF2 mRNA levels by 2.1-fold (both p-values = 0.005; Fig. 6A). However, NRF2 gene expression levels remained unaffected when treated with either DMF or Omav. The NQO1 mRNA levels were significantly increased by 7.5-fold and 11-fold following treatment with 10 µM (p = 0.002) and 20 µM SF (p < 0.001), respectively (Fig. 6B). The increase in NQO1 mRNA levels has also been observed in cells treated with DMF and Omav. Specifically, Omav at 50 nM and 100 nM significantly boosted NQO1 mRNA levels by 6.6-fold (p = 0.006) and 11.1-fold (p < 0.001), respectively. DMF at 30 µM upregulated the NQO1 mRNA levels by sevenfold (p = 0.003). A similar trend as seen in the NQO1 gene expression profile has been found in HO-1 mRNA levels, where 10 µM SF (p = 0.02) and 20 µM SF (p = 0.002) increased HO-1 gene expression levels by two- and 2.4-fold, respectively (Fig. 6C). However, although DMF and Omav appeared to increase HO-1 mRNA levels, the differences were statistically insignificant (all p-values >0.05). A dose-dependent increase in GCLM mRNA levels was observed with micromolar concentrations of SF (i.e., 5 µM [p = 0.02], 10 µM [p < 0.001], and 20 µM SF [p < 0.001]; Fig. 6D). A significant but modest increase (1.6-fold) in GCLM gene expression levels was identified with the treatment with 100 nM Omav (p = 0.01) but not with DMF. The antioxidant effects of SF and Omav, but not DMF, were reflected in the data from the glutathione assays. FRDA sensory neurons treated with 5, 10, or 20 µM SF and 50 nM or 100 nM Omav increased the GSH-GSSG ratio compared with untreated cells (all p-values <0.05; Fig. 6E).
SF treatment downregulates the expression of inflammatory markers in FRDA sensory neurons
The anti-inflammatory effects of SF treatment have been investigated in a preclinical FRDA model for the first time. The qRT-PCR results demonstrated a dose-dependent 1.6-fold to 2.8-fold decrease in TNF-α mRNA levels at the micromolar concentrations of SF [5 µM SF (p = 0.003), 10 µM SF (p < 0.001), and 20 µM SF (p < 0.001)] (Fig. 7A). At 100 nM, both SF and Omav were found to significantly reduce the gene expression levels of TNF-α by 1.5-fold (p = 0.009) and 1.6-fold (p = 0.002), respectively. The concentration of MCP-1 protein in the FRDA sensory neurons was dose dependently decreased by 1.35-fold to 1.7-fold following treatment with 5 µM SF (p = 0.004), 10 µM SF (p < 0.001), or 20 µM SF (p < 0.001) compared with untreated cells (Fig. 7B). SF at 100 nM concentration was also found to decrease MCP-1 protein concentration by 1.29-fold (p = 0.02). Similarly, 10 nM to 100 nM Omav treatment also resulted in a dose-dependent decrease of 1.39-fold to 2.3-fold in MCP-1 protein levels (all p-values <0.001). Although 30 µM DMF had no effect on TNF-α gene expression levels, it reduced MCP-1 protein expression levels by 1.28-fold (p = 0.02).

Discussion
In FRDA, most symptoms begin to manifest when the DRG sensory neurons succumb to pathological processes such as oxidative stress, inflammation, and ultimately death (Tiberi et al., 2023). To date, there is only one clinically approved therapy for this devastating disease for adults living in the United States and within the European Union, primarily targeting the NRF2 antioxidant signaling pathway (Lee, 2023). Therefore, alternative compounds addressing FXN deficiency and other downstream processes in the pathogenesis of FRDA are needed. Previous interventions targeting various stages of the disease did not have clinical success despite promising in vitro observations (Yang et al., 2022). This is mostly likely due to the limitations of prior FRDA models that lacked physiological relevance or the failure to fully recapitulate the genetic abnormalities of the condition (Schreiber et al., 2019). Therefore, FRDA iPSC-derived sensory neurons that carry the pathological number of GAA repeats from the patient serve as a leading model to screen novel therapies and offer a sustainable source of human sensory neurons that are physiologically and genetically relevant (Hulme et al., 2020). This study investigated the potential therapeutic effects of SF on sensory neurons derived from the FRDA iPSCs.
Upon the generation of isogenic control and FRDA sensory neurons, the phenotypic and gene expression profiles of these cells were established. Immunostaining verified the presence of the DRG neuronal markers PRPH, BRN3A, β-III TUBULIN, and ISLET1 (Fig. 1), in both isogenic control and FRDA sensory neurons, which is consistent with previous studies (Hulme et al., 2020; Viventi et al., 2021). In addition, positive gene expression of the peripheral neuronal marker PRPH and pan-sensory neuronal markers BRN3A and proprioceptive marker TRKC was detected in these cells (Fig. 1). These findings support the DRG sensory neuronal phenotype of the cells used in this drug-screening study (Alshawaf et al., 2018; Hulme et al., 2020). FXN protein expression levels in the FRDA sensory neurons were found to be lower than that of the isogenic controls; a finding that aligns with an observation made in another study where patients with FRDA experience a 70%–80% reduction of FXN levels compared with their healthy counterparts (Nachun et al., 2018). Elevated gene expression levels of gene-encoded chromatin-modifying enzymes such as HDAC1/3/6 and DNMT1 were found in the FRDA sensory neurons in this study (Fig. 2), indicating that the reduced expression of FXN may be associated with dysregulated epigenetic mechanisms, which is consistent with other studies using FRDA human primary cells (Quesada et al., 2015; Soragni and Gottesfeld, 2016; Xu et al., 2009). Compromised antioxidant mechanisms have also been observed in the FRDA sensory neurons showing reduced mRNA levels of NRF2, NQO1, HO-1, and GCLM (Fig. 2). These findings reflect the observations seen in blood samples of patients with FRDA, as well as in previous FRDA models (Chiang et al., 2020; La Rosa et al., 2021; Petrillo et al., 2019). The FRDA sensory neurons also exhibited higher expression levels of inflammatory markers (i.e., TNF-α and MCP-1), which have been reported by other researchers in blood samples from patients with FRDA (Bolotta et al., 2020). Collectively, these findings support the use of FRDA patient iPSC-derived sensory neurons as a robust cell model of the condition that provides measurable parameters suitable for drug screening.
As a neurodegenerative disease, neuronal atrophy observed in the DRG and cerebellum of patients with FRDA underscores the importance of evaluating therapeutic candidates for preventing neuronal cell death (Tiberi et al., 2023). Hence, the cell viability of isogenic control and FRDA sensory neurons was examined following a 24-hour exposure to SF and the positive control drugs, DMF and Omav, at selected concentrations as validated by other FRDA studies (Hayashi and Cortopassi, 2016; Jasoliya et al., 2019; Petrillo et al., 2019). While these drugs are not the main drugs of interest in this study, DMF was included as a well-established antioxidant with epigenetic properties, and Omav was included as the only clinically approved orally administered treatment for FRDA (Lee, 2023; Yang et al., 2022). We found that DMF treatment increased the proportion of viable FRDA sensory neurons, whereas Omav did not have any significant impact on cell viability. SF was shown to be the most effective drug, increasing the proportion of viable FRDA sensory neurons by up to 61% (Fig. 3B). The protective effects of SF were observed even at concentrations as low as 100 nM, which is supported by a study that investigated SF in an in vitro model of Alzheimer’s disease (Zhang et al., 2017). Moreover, the observed biphasic dose–response effect of SF on FRDA sensory neuronal viability aligns with findings reported by others and this effect is associated with the endogenous cellular defense mechanisms of SF activating vitagenes (protective genes), enhancing cellular resilience against neurobiological oxidative stress and promoting cell survival (Calabrese and Kozumbo, 2021). Most importantly, none of the drug treatments reduced the viability of the isogenic control and FRDA sensory neurons in the present study. This seems to be contrary to findings in cancer studies where SF at similar micromolar concentrations induced apoptosis in human breast cancer cell lines (Yang et al., 2018a). Indeed, SF at these concentrations was also found to reduce cellular death of injured cortical neurons and human retinal cells under oxidative stress (Kwa et al., 2019; Wu et al., 2012). These findings suggest the tissue-specific activity of SF, making it an ideal treatment option with potentially minimum off-targeting effects. Furthermore, improvement in cell viability could also be mediated by different mechanisms and there could be dose-dependent regulation of various disease pathways that are not revealed in this study as the potential complex networks involved make it difficult to conclude direct cause and consequence relationships (Ben-Hamo et al., 2020; Hu et al., 2021; Michoel and Zhang, 2023).
As a hallmark of FRDA, restoring FXN expression is one of the key targets for treating FRDA (Tiberi et al., 2023). Here, we found that SF was the only drug capable of increasing FXN mRNA and protein levels in FRDA sensory neurons, whereas DMF and Omav did not modulate FXN expression at either the gene or protein levels. The absent effect of Omav on FXN expression levels in this study corroborates with the earlier findings in FRDA patient fibroblasts (Petrillo et al., 2019). However, our results pertaining to DMF treatment are contrary to previous studies using FRDA patient lymphoblasts, fibroblasts, and mouse models, which exhibit dose-dependent effects on FXN mRNA and protein expression (Jasoliya et al., 2019; Petrillo et al., 2019). This inconsistency may be due to cell-type-specific responses to drug treatments (Perl et al., 2017; Wheeler et al., 2023). The limited number of available articles suggests that SF can increase FXN expression in fibroblast cells and frataxin-silenced motor neurons, but dose-dependent effects of SF are not revealed because these studies only tested one single dose (Petrillo et al., 2019; Petrillo et al., 2017). For the first time, we found that SF at nanomolar and micromolar doses effectively promoted the gene and protein expression of FXN. Although we did not observe a dose-dependent effect, previous studies testing 5 µM or 10 µM SF in FRDA patient fibroblasts and mouse motor neurons also showed comparable increases in FXN gene and protein expression levels (Petrillo et al., 2019; Petrillo et al., 2017). In this study, SF exhibited a multiphasic regulation of FXN expression, likely due to the presence of the complex cellular regulatory mechanisms, differential cellular stress responses, role of SF in cross talk between different signaling pathways, and the heterogenous feature of the iPSC-derived cells (Frohlich et al., 2019; Hulme et al., 2020; Martin and Dimmitt, 2019; Pascual-Ahuir et al., 2020; Ramesh and Krishnan, 2023; Uddin et al., 2020). Furthermore, SF may not always exert dose-dependent effects on all target genes and proteins at different concentrations, it can preferentially act on selective pathways and trigger a significant impact on distinct genes and proteins during a fixed period of time (i.e., 24 hour), as shown by the inconsistent dose-dependent effects observed in other investigations (Kubo et al., 2017; Liu et al., 2014; Vauzour et al., 2010). Numerous researchers have identified nonlinear pharmacological behaviors of other drugs in various studies. For instance, lymphocyte cell lines exhibit a nonlinear dose response to microtubule inhibitors, while resveratrol exhibits nonlinear dose-dependent protective effects in both humans and mice (Cai et al., 2015; Potashnikova et al., 2019). Enrichment analysis in yeast experiments also revealed that 77 genes displaying nonlinear responses in single-drug treatments were associated with diverse biological pathways and functions (Taguchi and Turki, 2021).The upregulation of FXN gene expression levels was not directly correlated to the increase in cell viability in this study. Certain concentrations of SF were unable to improve FXN levels but yet increased cell viability. This observation aligns with a study which reported that FXN depletion does not appear to correlate with the apoptosis of the DRG neurons (Lu et al., 2009). To our knowledge, there is no literature or reported clinical advice that explicitly mentions the benchmark FXN mRNA and protein levels that can be consistently applied across the various FRDA-affected tissues needed to predict improved clinical outcomes in the patients. Therefore, it is difficult to forecast the direct clinical benefits of SF’s ability to upregulate FXN levels shown in our study. It is also worth noting that Omav was not designed to increase FXN levels and it was approved as the first treatment for FRDA due to its NRF2-inducing properties and ability to improve modified Friedreich Ataxia Rating Scale (mFARS) scores (Lynch et al., 2021; Lynch et al., 2019; Lynch et al., 2024; Petrillo et al., 2019). While the ability of new drugs to increase FXN levels remains a primary point of interest, the impact on FXN levels should not be the sole determinant in deciding if novel compounds should advance to clinical trials.
Epigenetic modifications are emerging therapeutic targets across various gene-related disorders since the downregulation of the above can facilitate the transcription of downstream genes (Jiang et al., 2016; Zhao et al., 2018). Consequently, DNMTs and HDACs have been investigated in the pathogenesis of FRDA (Quesada et al., 2015; Soragni and Gottesfeld, 2016). No studies have been conducted to investigate the epigenetic effects of SF in the context of FRDA. Here, we report that SF can downregulate HDAC1, HDAC3, HDAC6, and DNMT1 gene expression levels in the FRDA sensory neurons (Fig. 5). These observations echo SF’s DNMT and HDAC inhibitory properties in models of cancer and Alzheimer’s disease (Kaufman-Szymczyk et al., 2015; Zhang et al., 2017). Specifically, SF at micromolar concentrations was found to downregulate HDAC1 and HDAC3 expression in β-amyloid-induced SH-SY5Y cells (Zhang et al., 2017). This downregulation was followed by an elevated expression of the P75 neurotrophin receptor, which protects cells from β-amyloid toxicity and improves binding affinity to neurotrophic factors (Zhang et al., 2017). Thus, the epigenetic modulation effects of SF on HDAC1 and HDAC3 in the present study may have contributed to its neuroprotective effects. SF, Omav, and DMF, at specific concentrations, all appear to have the potential to achieve HDAC6 inhibition, which may have downstream effects on modulating neuronal oxidative stress by reducing peroxides (Simões-Pires et al., 2013). The suppression of DNMT1 expression following SF treatment was also found to be associated with the reactivation of certain genes, such as cyclin D2, for apoptosis transition or NRF2 for redox protection (Kaufman-Szymczyk et al., 2015; Zhao et al., 2018). Thus, the epigenetic modulatory effects of SF may influence the protection against oxidative stress in the sensory neurons studied here. Despite having the ability to reverse aberrant gene expression of disease markers, many HDAC inhibitors (HDACIs) (e.g., Compound 109) are not likely to be considered suitable drug candidates due to their limited ability to penetrate the CNS and they can generate toxic metabolites that are harmful to cardiac tissues (Codazzi et al., 2016). However, SF bears therapeutic promise as an HDACI given its lipophilic nature and its ability to cross the BBB (Sun et al., 2017).
Oxidative stress plays a significant role in the degeneration of specific neurons implicated in FRDA as evidenced by the reduced expression levels of antioxidant enzymes (NQO1, HO-1, and GCL) (La Rosa et al., 2019; Petrillo et al., 2019). To the best of our knowledge, there is only one preclinical study that had investigated the effects of SF, Omav, and DMF on these markers of antioxidant pathways, but using FRDA patient fibroblasts (Petrillo et al., 2019). All three drugs have shown to induce NRF2 gene and protein activation in these cells (Abeti et al., 2018; Petrillo et al., 2019). In contrast, we only observed a significant increase in the NRF2 mRNA levels by SF treatment in the present study. The difference in our observations is likely due to the different cell types used in the two studies (Petrillo et al., 2019). In addition, gene expression levels may not always reflect molecular pathway activities due to the potential involvement of unforeseen post-translational modifications, which further complicates the establishment of drug cause and effect relationships (Ben-Hamo et al., 2020; Michoel and Zhang, 2023). In addition, the absence of an effect of Omav and DMF in the FRDA sensory neurons could be attributed to the main mechanism of action of both drugs, that is, preventing the degradation of the NRF2 protein. Therefore, increases in NRF2 mRNA are not expected (Tiberi et al., 2023).
For the NRF2 downstream Phase II redox enzymes, SF upregulated the expression levels of all genes studied. We showed an increase in NQO1 gene expression of up to 11-fold compared with the untreated control. DMF treatment also enhanced NQO1 gene expression but at a smaller magnitude (7.1-fold increase) compared with SF and Omav treatment. Among the three drugs, Omav resulted in the greatest increase in NQO1 mRNA levels (11.1-fold). A similar drug-induced trend in increasing NQO1 mRNA levels has been demonstrated by others (Petrillo et al., 2019). HO-1 is an attractive biomarker that has been associated with reduced oxidative stress and inflammation in various brain conditions (Vijayan et al., 2018). The upregulation of HO-1 protein expression also contributes to radical scavenging activities and anti-inflammatory and neuroprotective effects in both the peripheral and CNSs (Calabrese et al., 2007). Herein, the mRNA levels of HO-1 were only upregulated by SF but not by the two positive drug controls. Although this finding is contrary to Petrillo et al where both DMF and Omav were found to upregulate HO-1 mRNA levels in FRDA patient fibroblasts, the HO-1 gene expression levels may differ according to the level of cellular stress experienced by different disease models or cell types (Saha et al., 2020; Tonelli et al., 2018). One of the subunits of the GCL complex is GCLM, which plays a key role in the synthesis of GSH and thus modulates GSH and GSSG levels (Petrillo et al., 2017). Here, we found that SF was more potent in regulating GCLM mRNA levels compared with Omav. DMF treatment showed no effect. Despite the lack of studies reporting the effects of the three drugs on GCLM expression in FRDA models concurrently, the gene expression profile of GCL induced by these drugs as shown in Petrillo et al. is similar to that of GCLM in our study. These findings, collectively, indicate that SF may be a superior compound in eliciting positive modulatory effects on GCL and GCLM (Petrillo et al., 2019). Overall, the different modes of action exerted by SF, DMF, and Omav on the dynamic regulation of NRF2 by KEAP1 in different cell types may account for the varied effects of these drugs on the expression of NRF2 and genes encoding downstream redox enzymes, as well as for the possible time-dependent changes in gene expression (Tiberi et al., 2023; Tonelli et al., 2018; Vomund et al., 2017). Furthermore, the gene expression profiles of the redox enzymes modulated by the three drugs reflect the associated increase in the GSH-GSSG ratios (Fig. 6E). The increase in the GSH-GSSG ratio indicates that SF is capable of reversing abnormal redox status in the FRDA patient sensory neurons, thus conferring a protective cellular state against oxidative stress (Tiberi et al., 2023). This finding is in accordance with other studies showing that SF can successfully reestablish glutathione homeostasis in FXN-silenced mouse motor neurons (Petrillo et al., 2017). However, DMF did not exert any effect on improving glutathione levels, which may be related to its lack of gene modulatory effects on GCLM (Fig. 6D). On the contrary, Omav exerted the most effective redox effect among the three drugs, as reflected by the largest increase in GSH-GSSG ratio compared with the untreated control. Collectively, SF has comparable efficacy in exerting its antioxidant properties on FRDA-affected cells against oxidative stress similar to Omav.
Neuroinflammation is a common feature of neurological diseases, including FRDA (Lu et al., 2009). In this study, the TNF-α mRNA levels were found to be suppressed by SF at micromolar concentrations in a dose-dependent manner by up to 2.8-fold (Fig. 7A). This result aligns with a previous study showing SF’s ability to reduce inflammation in a dose-dependent manner in lipopolysaccharide-stimulated mouse microglia cells (Calabrese et al., 2010; Eren et al., 2018). Although DMF treatment was shown to suppress the TNF-α mRNA levels in a mouse model with induced aneurysm and various neurodegenerative diseases, no effect on TNF-α expression was observed in our study (Majkutewicz, 2022; Pascale et al., 2020). The difference in observations may be explained by cell-type-specific response or time-dependent changes in gene expression mentioned above. Herein, Omav exerted a similar anti-inflammatory effect on the gene expression levels of TNF-α as SF. Other studies have also reported the anti-inflammatory effects of Omav in non-FRDA models (Hu et al., 2022; Jiang et al., 2022). Nevertheless, the anti-inflammatory effects of SF, Omav, and DMF have been reaffirmed by the MCP-1 protein expression levels in this study. MCP-1 is a key inflammatory mediator and useful biomarker for predicting disease progression in various inflammatory conditions affecting the CNS (Lu et al., 2009; Singh et al., 2021). Elevated MCP-1 levels were found in various neurodegenerative disorders such as Alzheimer’s disease and Parkinson’s disease (Singh et al., 2021). To date, our study is the first to reveal that FRDA-affected sensory neurons exhibit elevated MCP-1 levels, and all three drugs successfully suppressed MCP-1 protein expression in this context. The anti-inflammatory effects of DMF on MCP-1 were consistent with findings reported in a review article on preclinical neurodegenerative disease models (Majkutewicz, 2022). The dose-dependent inhibitory effect of Omav on MCP-1 protein levels in patient sensory neurons aligns with the negative correlation observed between MCP-1 mRNA levels and Omav concentrations in a mouse leukemia cell line (Probst et al., 2015). Overall, our data highlight the efficacy of SF in downregulating proinflammatory cytokines at both the gene and protein levels, thereby supporting its therapeutic benefits in addressing neuroinflammation in FRDA.
In conclusion, we demonstrate that the FRDA iPSC-derived patient sensory neurons used in this study display the various FRDA characteristics that reflect the disease-related markers observed in real FRDA patient tissues, which makes it suitable for drug screening. Our data suggest that SF had superior efficacy in increasing the proportion of viable patient sensory neurons compared with the positive control drugs, and none of these drugs, at the tested concentrations, reduced the viability of isogenic control or patient sensory neurons. This study also showed that SF treatment could increase the expression of frataxin and various antioxidant markers while inhibiting the expression of epigenetic markers and inflammatory cytokines (Table 1). In contrast, DMF and Omav have only shown selective regulatory effects on some of these markers. It is noteworthy that this study investigated the epigenetic, antioxidant, and anti-inflammatory effects of SF using sensory neurons derived from iPSCs of one patient with FRDA with moderate severity of disease, reflected by 550/830 GAA repeats in the FXN gene in the latter. Therefore, there will be a need to examine the effect of SF in sensory neurons generated from iPSCs derived from other patients with FRDA that carry different numbers of GAA repeats representing less and more severe disease. Conducting whole-genome RNA sequencing and chromatin immunoprecipitation assay with sequencing will also provide comprehensive insight into the genome-wide impact of SF treatment, which will help to identify genes that may be affected upstream or downstream of the GAA repeats, establish epigenetic response to SF treatment, and predict any potential off-targeting effects. These additional studies will help to consolidate the relevance of using SF to treat patients with different onset or severity of FRDA and at various stages of the disease. Rigorous pharmacokinetic modeling and clinical trials will be essential in establishing SF’s full therapeutic potential in FRDA. In summary, this study presents SF as an attractive multipronged therapy to battle the different molecular events driving the symptoms in individuals with FRDA.
Innovation
For the first time, sulforaphane (SF) is being evaluated using Friedreich ataxia (FRDA) sensory neurons generated from induced pluripotent stem cells (iPSCs) carrying 550/830 guanine–adenine–adenine (GAA) repeats in the frataxin gene (FXN gene), and its effects are compared with the clinically approved drug, omaveloxolone (Omav). In vitro FRDA models used in other drug screening studies involved either FRDA fibroblasts and lymphoblastoid cells or mice models carrying a maximum of 230 GAA repeats. This study has also tested the limits of SF dosing, comparable with the in vitro doses of Omav reported in the literature, in generating beneficial effects against oxidative stress, inflammation, and abnormal epigenetic mechanisms. See Graphical Abstract.
Materials and Methods
Generation of FRDA patient iPSC-derived sensory neurons
The FRDA patient iPSCs and CRISPR-corrected isogenic control line were generated from a male FRDA patient, by Marek Napierala and his team (University of Alabama) (Ku et al., 2010; Miellet et al., 2024a). The protocol describing CRISPR-Cas9 gene editing in both iPSC lines to integrate a doxycycline-inducible NGN2 gene cassette into the CLYBL locus was recently published (Miellet et al., 2024a; Miellet et al., 2024b). The successful insertion of the NGN2 gene cassette is indicated by positive mApple expression. The NGN2 FRDA and isogenic iPSC lines were induced to form neural crest progenitor cells, which were further differentiated into peripheral sensory neurons using an established protocol with modifications (Supplementary Fig. S1) (Hulme et al., 2020). Briefly, the iPSC colonies were cultured in TeSR-E8 media (#5990, Stem Cell Technologies, USA) contained in vitronectin XFTM (#07180, Stem Cell Technologies, USA)-coated flasks. Upon reaching 80% confluency, iPSC colonies were harvested with 0.5 mM EDTA (#R1021, Thermo Fisher, USA) and induced into NCCs using the STEMdiff neural crest differentiation kit (#08610, Stem Cell Technologies) on 10 µg/mL laminin-coated plates (#23017015, Thermo Fisher Scientific). Subsequently, the NCCs were harvested using Accutase (#00–4555-56, Invitrogen, USA) and replated to be induced into sensory neurons for downstream assays. All plates for maintaining sensory neurons were coated with 10 µg/mL poly-d-lysine (#A3890401, Thermo Fisher Scientific) for 30 min at room temperature followed by a second coating with 10 µg/mL laminin at 4°C overnight. The NCCs were cultured in a complete neurobasal medium with 2 µg/mL doxycycline (D9891, Sigma-Aldrich, USA) for 3 days to activate NGN2 expression for cellular differentiation into sensory neurons. All sensory neurons were maintained in complete neurobasal medium for a minimum of 14 days postdoxycycline treatment to achieve physiological capacity (Alshawaf et al., 2018; Hick et al., 2013; Hulme et al., 2020). The complete neurobasal medium was made from neurobasal medium (#21103–049, Thermo Fisher Scientific) supplemented with 1× of each N-2 (#17502–048, Thermo Fisher Scientific), B-27 (#12587–010, Thermo Fisher Scientific), insulin–transferrin–selenium-A (#51300–044, Thermo fisher Scientific), and growth factors, including 10 µM Y-27632 (#72302, Stem Cell Technologies) and 10 ng/mL of each of brain-derived neurotrophic factor (BDNF) (#78005, Stem Cell Technologies) and glial cell line-derived neurotrophic factor (GDNF) (#78058, Stem Cell Technologies). neurotrophin-3 (NT-3) (#78074, Stem Cell Technologies) and beta nerve growth factor (ß-NGF) (#78092, Stem Cell Technologies). Cells were maintained at a temperature of 37°C and 5% CO2. Media changes were performed once every 2 to 3 days as required. All experiments conducted using the FRDA patient and isogenic control cell lines were approved by the Swinburne Biosafety and Research Ethics Committee (2021_5740) and the
Henceforth, the FRDA iPSC-derived sensory neurons with and without the GAA repeats are referred to as “FRDA sensory neurons” and “isogenic control sensory neurons,” respectively.
Validation of DRG sensory neurons derived from FRDA-derived iPSCs
Immunofluorescence staining
Immunofluorescence imaging of sensory neurons derived from FRDA iPSCs was performed as previously described (Hulme et al., 2020). All primary and secondary antibodies were prepared in 10% donkey serum in phosphate-buffered saline (PBS) and used at a 1:500 dilution. Antibodies used were as follows: mouse anti-BRN3A (#MAB1585, Millipore, USA), rabbit anti-ISLET1 (#ab20670, Abcam, UK), mouse anti-PERIPHERIN (#MAB1527, Millipore), mouse anti-ß-III TUBULIN (#MAB1637, Millipore), donkey anti-mouse IgG-488 (#ab150109, Abcam), and donkey anti-rabbit IgG-647 (#ab150063, Abcam). Primary antibodies were incubated overnight at 4°C followed by incubation with the secondary antibody and 1 µg/mL DAPI (#D9542, Sigma-Aldrich) at room temperature for 1 hour. Immunofluorescent images were captured using a 63× objective on a Leica confocal SP8 microscope (Leica Stellaris, Wezler, Germany). Exported images were then processed with ImageJ software.
RNA extraction and complementary DNA synthesis
Total RNA was extracted from sensory neurons, which were seeded at a density of 1 × 105 cell/mL in 12-well plates using the RNeasy Plus Mini Kit (#74132, Qiagen, Valencia, CA), following the manufacturer’s instructions.
Complementary DNA (cDNA) was synthesized from 100 ng RNA using the High-Capacity cDNA Synthesis Kit (#4374966, Applied Biosystems, USA). The amplification of cDNA was carried out for 10 min at 25°C (initialization) followed by 120 min at 37°C (reverse transcriptase step) and 5 min at 85°C (termination of reverse transcriptase reaction).
Quantitative real-time polymerase chain reaction
TaqMan® Fast Advanced Master Mix (#4444963, Applied Biosystems) and the following gene expression assays were used in qRT-PCR based on the manufacturer’s protocol. Each reaction was carried out in triplicate on a MicroAmp optical plate, and each reaction consisted of a minimum of 100 ng of cDNA, 1× primer and probe, nuclease-free water, and 1× Master Mix. The plate was sealed with an optical adhesive cover (#4360954, Thermo Fisher), centrifuged at 500 g for 2 min, and subjected to the following qRT-PCR cycling conditions: 50°C for 2 min, 95°C for 20 s followed by 40 cycles of 95°C for 3 s and 60°C for 30 s. The qRT-PCR data were analyzed using the 2−ΔΔCt method with glyceraldehyde 3-phosphate dehydrogenase (GAPDH, Hs99999905_m1) as the housekeeping gene, and the data are shown as changes in fold difference relative to the untreated control (Hulme et al., 2020). The TaqMan gene expression assays (#4331182, Applied Biosystems) chosen to validate the identity of the sensory neurons generated are peripherin (PRPH, Hs00196608_m1), brain-specific homeobox/POU domain protein 3A (BRN3A, Hs00366711_m1), and TRKC (Hs00176797_m1) (Hulme et al., 2020; Viventi et al., 2021). The human adult retinal pigment epithelial cell line, ARPE-19, was used as a negative control for the lack of expression of the sensory neuronal markers. The gene expression of RLBP1 (Hs00165632_m1), a key modulator of the visual cycle, was measured in the ARPE-19 cells to confirm its identity as human retinal cells (Samuel et al., 2017).
Drug treatment of FRDA isogenic control and patient sensory neurons
SF (#S6317, Sigma-Aldrich) and DMF (#242926 Sigma-Aldrich) were dissolved in DMSO (#D5879, Sigma-Aldrich), while Omav, which was kindly donated by Reata Pharmaceuticals, Inc. (USA), was prepared in anhydrous DMSO (#276855, Sigma-Aldrich, USA). DMF and Omav have been included as positive control drugs with NRF2 inducing properties shown in other studies (Abeti et al., 2018; Kalinin et al., 2013; Lee, 2023; Petrillo et al., 2019; Probst et al., 2015; Yang et al., 2022; Zighan et al., 2022).
To establish the response of the sensory neurons to the various drug treatments, both isogenic control and patient sensory neurons were treated with 100 nM to 20 μM SF, 30 μM DMF, or 10 nM to 100 nM Omav for 24 hour. The concentrations chosen for SF and DMF were based on previously published articles (Dulull et al., 2018; Hayashi and Cortopassi, 2016; Jasoliya et al., 2019; Kwa et al., 2019; Petrillo et al., 2019; Yang et al., 2022). The dose selection for Omav was based on the guidance received from Reata Pharmaceuticals, Inc. (now acquired by Biogen, Inc.) and previous in vitro studies that showed therapeutic effects (Probst et al., 2015; Zighan et al., 2022). Untreated cells and 0.1% DMSO (drug vehicle control) served as the negative controls. As there were no significant effects of the drug vehicle on cell viability on the patient and isogenic control sensory neurons, as described later in this article, untreated cells were used as a negative control for the following molecular assays (i.e., gene expression, GSH/GSSG, and enzyme-linked immunosorbent assays [ELISAs]). Since none of the drugs were found to impact the viability of the isogenic control sensory neurons, these cells were not included in further mechanistic studies conducted here. To determine the net effects of the drugs tested, comparisons of these effects between untreated and treated FRDA sensory neurons were made. This data analysis approach is consistent with those reported by others (Codazzi et al., 2016). Drug concentrations (100 nM, 500 nM, 5 μM, 10 μM, 20 μM SF and 30 μM DMF) that induced a statistically significant increase in cell viability were to treat the patient sensory neurons conducted in the following gene expression and protein assays. Although Omav treatment did not alter cell viability, the patient sensory neurons were treated with this drug as the only approved FRDA treatment at 10 nM, 50 nM, and 100 nM to compare its molecular mechanisms of action with those of SF.
Effects of SF on the viability of FRDA isogenic control and patient sensory neurons
The CellTiter 96 Aqueous One Solution Cell Proliferation Assay kit (#G3581, Promega, USA) was used to determine the metabolic activity of cells, which is directly proportional to the number of living cells (Promega, 2023; Yanez et al., 2019). Briefly, 20 μL of the MTS solution was added into each well of a 96-well plate containing 10,000 cells in 100 μL of culture medium and the sensory neurons were incubated for 4 hours at 37°C. The absorbance was measured at 490 nm. The percentage cell viability was calculated using the manufacturer’s formula as follows: (Mean difference treated group − Blank)/(Mean difference untreated group − Blank) × 100%. The data presented are based on the mean value from three independent experiments, each conducted with three technical replicates per experiment.
Effects of SF treatment on the gene expression of biomarkers dysregulated in FRDA tissues and cells
Total RNA and cDNA were obtained as described above. The abovementioned qRT-PCR protocol was repeated with different TaqMan gene expression assays (#4331182, Applied Biosystems, USA) that measure the mRNA levels of the FXN gene, as well as other genes involved in regulating epigenetic mechanisms (HDAC1/3/6, DNMT1), oxidative stress (NRF2, NQO1, HO-1, GCLM), and inflammation (TNF-α) (Bolotta et al., 2020; Jasoliya et al., 2019; Petrillo et al., 2019; Quesada et al., 2015; Soragni and Gottesfeld, 2016; Xu et al., 2009). GAPDH was used as the housekeeping gene. The TaqMan gene expression assays (FXN, Hs00175940_m1; DNMT1, Hs00154749_m1; HDAC1, Hs02621185_s1; HDAC3, Hs00187320_m1; HDAC6, Hs00997427_m1; NRF2, Hs00975961_g1; HO-1, Hs01110250_m1; NQO1, Hs01045993_g1; GCLM, Hs00978072_m1; TNF-α, Hs00174128_m1) were used in this study for investigating the effects of SF on pathological markers involved in FRDA. The qRT-PCR data are represented as changes in fold difference relative to GAPDH and the untreated cells served as calibrator cells (Hulme et al., 2020). The data presented are based on the mean value from three independent experiments, each conducted with three technical replicates per experiment.
Effects of SF on protein expression levels of FXN and markers of oxidative stress and inflammation in FRDA patient sensory neurons
FXN ELISA
The effect of various treatments on the protein expression levels of FXN in the sensory neurons was measured with the Human FXN ELISA kit (#ab176112, Abcam, UK) in accordance with the manufacturer’s instructions. Briefly, the patient sensory neurons were seeded at a density of 1 × 106 cell/mL in 6-well plates and treated with SF, DMF, and Omav at the said concentrations for 24 hour. Cells were harvested with Accutase, rinsed twice with PBS, and then lysed in the cold cell extraction buffer provided in the kit, and total protein concentrations were assayed using the BCA Protein Assay Kit (#ab102536, Abcam, UK). Each protein sample was diluted in 100 μg/mL assay buffer and loaded onto an antitag-coated microplate. This was followed by incubating the sensory neurons with Antibody Cocktail for an hour at room temperature on a plate shaker set to 400 rpm as per the manufacturer’s protocol. Subsequently, each well was washed three times with 1× wash buffer PT and then 100 μL of the tetramethylbenzidine (TMB) development solution was added to produce a colorimetric signal proportional to the concentration of protein. The absorbance was measured at 450 nm. The data presented are based on the mean value from three independent experiments, each conducted with two technical replicates per experiment as recommended by the manufacturer.
MCP-1 ELISA
The effect of the various treatments on the protein expression levels of MCP-1 was measured with the Human MCP-1 ELISA kit (#ab179886, Abcam) in accordance with the manufacturer’s instructions. Briefly, the patient sensory neurons were seeded at a density of 1 × 106 cells/mL in 6-well plates and treated with different concentrations of the test compounds for 24 hour. Cell culture medium was collected and centrifuged at 2000 g for 10 min. This was followed by incubating the sensory neurons with Antibody Cocktail for an hour at room temperature on a plate shaker set to 400 rpm as per the manufacturer’s protocol. Subsequently, each well was washed three times with 1× wash buffer PT and then 100 μL of the TMB development solution was added to produce a colorimetric signal proportional to the concentration of protein. The absorbance was measured at 450 nm. The data presented are based on the mean value from three independent experiments, each conducted with two technical replicates per experiment as recommended by the manufacturer.
GSH-GSSG detection assay
To establish whether SF can reduce oxidative stress in FRDA patient sensory neurons, the concentrations of GSH and GSSG levels after 24-hour drug treatments were measured using the Glutathione Fluorescent Detection Kit (#EIAGSHF, Invitrogen, USA). In brief, cell lysate samples in ice-cold 5% aqueous 5-sulfo-salicylic acid dihydrate (SSA: #S2130, Sigma-Aldrich) were sonicated for 2 min, freeze–thawed three times in liquid nitrogen, and centrifuged at 20,817 g for 10 min at 4°C to release total protein content. To measure the total protein concentrations of GSH and GSSG, the supernatant was collected and further diluted (1:5) in 1× assay buffer provided in the kit. Samples and standards were then loaded to a black 96-well half-area plate with the addition of detection reagent and subsequent reaction mixture containing NADPH concentrate (10×), GSH reductase concentrate (10×), and GSH assay buffer (1×). After incubation at room temperature for 15 min, fluorescence was read at an excitation wavelength of 390 nm and an emission wavelength of 510 nm. An increase in the GSH-GSSG ratio indicates a protective effect against oxidative stress (Dulull et al., 2018).
Statistical analyses
Statistical analyses were conducted using the GraphPad Prism software version 9.3 (San Diego, CA, USA). To determine the significance of differences between the two groups, Student’s t-test was performed (Codazzi et al., 2016). One-way analysis of variance with Bonferroni correction was performed to examine the significance of differences across multiple groups (Abeti et al., 2018). All data presented are based on the mean value from three independent experiments, each conducted with at least two technical replicates per experiment. All data are presented as mean ± standard error of the mean. Statistical significance was defined as *p < 0.05, **p < 0.01, and ***p < 0.001, compared with untreated controls.
Electronic laboratory notebook was not used.
Footnotes
Acknowledgments
This study was supported by funding from the School of Health Sciences at Swinburne University of Technology, Melbourne School of Health Sciences at the University of Melbourne, Friedreich Ataxia Research Alliance, Medical Research Future Fund Stem Cell Therapies Mission, and the University of Wollongong. Omav was kindly donated by Reata Pharmaceuticals, Inc. (USA), which has now been acquired by Biogen, Inc. (USA).
Authors’ Contributions
M.N. established the cell model. F.K., B.T., and M.D. designed the experiments. W.Y., S.M., M.M., and F.K. performed the experiments. All authors analyzed and interpretated the data. W.Y. and F.K. wrote the initial draft of the article. All authors were involved in the editing, reviewing, and approving the content of the article.
Author Disclosure Statement
The authors have no conflict of interest to declare.
Funding Information
The funding to support the publication costs of this article was supported by the Melbourne School of Health Sciences.
Supplementary Material
Supplementary Figure S1
Abbreviations Used
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.