
Abstract
Aims:
Periprosthetic osteolysis (PPO), a leading cause of aseptic loosening in joint replacement, arose from complex interactions among osteoblasts, osteoclasts, and osteocytes. Given the pivotal role of connexin 43 (Cx43) in osteocyte communication and bone remodeling, investigating its function was essential for understanding the mechanisms of osteolysis. Our previous studies showed that titanium (Ti) particles increased Cx43 expression in osteocytes. However, the role of Cx43 in osteolysis remained unclear. This study investigated the role of Cx43-mediated regulation of osteocytes on osteoclastogenesis in wear debris-induced osteolysis.
Results:
Using Dmp1-cre conditional Cx43 knockout mice and the MLO-Y4 osteocyte cell line, we demonstrated that Cx43 deficiency reduced bone resorption and osteoclastogenesis, thereby improving bone remodeling in a Ti particle-induced osteolysis model. Sequencing analysis revealed that Cx43 gene expression changes might be linked to oxidative stress and the Janus Kinase (JAK)-STAT pathway. Elevated Cx43 expression in osteocytes stimulated by Ti particles increased STAT1 protein phosphorylation, induced oxidative stress, elevated the Receptor Activator of Nuclear Factor Kappa-Β Ligand (RANKL)/Osteoprotegerin (OPG) ratio, and promoted osteoclast activation and bone resorption. Conversely, Cx43 gene knockout decreased STAT1 protein phosphorylation and enhanced Nuclear Factor Erythroid 2-Related Factor 2 (NrF2) protein expression. Blocking the JAK-STAT signaling pathway activated by Cx43 increased NrF2 expression, reduced reactive oxygen species levels, and subsequently decreased the RANKL/OPG ratio.
Innovation and Conclusions:
This study identified a novel mechanism where Cx43 in osteocytes promoted osteoclastogenesis through JAK-STAT pathway activation and oxidative stress in wear debris-induced osteolysis. These findings highlighted the critical role of Cx43 in bone resorption and suggested targeting Cx43 or the JAK-STAT pathway as potential therapeutic strategies to mitigate osteolysis and improve implant longevity. Antioxid. Redox Signal. 43, 215–238.
Introduction
Total hip arthroplasty is extensively utilized for treating end-stage joint disorders. As the number of surgical patients rises, aseptic loosening of joint prostheses is becoming more prominent. The pathogenic factors of aseptic loosening involve biological reactions due to wear particles, which lead to periprosthetic osteolysis (PPO) eventually. However, the mechanism of wear particle-induced PPO remains to be elucidated. Previous studies have demonstrated that wear particles promote inflammatory cells and mediators, subsequently activating osteoclasts and leading to increased bone resorption (Wu et al., 2022). Concurrently, osteoblast function is inhibited, reducing bone formation and ultimately exacerbating PPO (Chai et al., 2023).
Osteocytes are located within the lacunae of bone tissue, constituting 90% of the bone cell population. They communicate with each other and with other cell types through dendrites that extend to the bone surface (Bonewald, 2007). Accumulating evidence suggests that osteocytes play a crucial role in regulating osteoblast and osteoclast function (Clarke, 2008). Osteocytes release Receptor Activator of Nuclear Factor Kappa-Β Ligand (RANKL) and produce inflammatory cytokines, stimulating osteoclast differentiation (Nakashima et al., 2011). Some studies indicate that osteocytes play a pivotal regulatory role in the process of osteolysis by influencing key pathways and cellular interactions involved in bone resorption (Yoshimoto et al., 2022).
Connexin 43 (Cx43) is predominantly found in cartilage and bone tissue and participates in the formation of membrane hemichannels and gap junctions (GJs) between cells (Donahue et al., 2017). As the principal protein of GJs, Cx43 plays a vital role in substance transmission and signal exchange among osteocytes, osteoclasts, and osteoblasts. Some studies have shown that Cx43 is closely associated with bone cell differentiation, skeletal metabolism, bone remodeling, and maintaining osteocyte activity (Hoppock et al., 2023; Moorer et al., 2017; Takemura et al., 2019). Inhibition of Cx43 reduces the early inflammatory response induced by implants (Calder et al., 2015). Knocking down Cx43 in osteocytes leads to a reduction in Sclerostin (SOST) expression (Hoppock et al., 2023), which can promote the formation of osteoclasts and enhance their resorptive activity (Wijenayaka et al., 2011). Emerging evidence suggests that GJs formed by Cx43 play a pivotal role in regulating oxidative stress (Gao et al., 2015; Yan et al., 2012). Reactive oxygen species (ROS) have been shown to affect osteocyte signaling and bone remodeling processes, known to significantly contribute to PPO (Sies et al., 2022; Wei et al., 2022).
Given these considerations, connexin 43 has captured our focus. Our recent research demonstrated that inhibiting osteocytic Cx43 gap junctions enhances bone formation and mitigates wear particle-induced osteolysis (Chai et al., 2022, 2023). However, the role of Cx43 in regulating bone resorption and its underlying mechanisms during osteolysis remains unclear. In this study, we found that titanium (Ti) particles increased Cx43 expression in osteocytes, enhanced osteoclast activity, and exacerbated osteolysis. Cx43 deficiency reduced osteoclastogenesis and improved bone remodeling. Further results demonstrated Cx43 in osteocytes played a critical role in osteoclastogenesis through the Janus Kinase (JAK)-STAT pathway and the regulation of oxidant stress.
Innovation
This study revealed that Cx43 regulated oxidative stress and osteoclast activation through the JAK-STAT pathway, contributing to wear debris-induced osteolysis by suppressing NrF2, increasing ROS, and promoting bone resorption. These findings highlighted Cx43 as a potential therapeutic target (Fig. 10).
Results
Ti particles increased Cx43 expression and the markers related to osteoclastic activity in mice femur osteolysis model
Microcomputed tomography (micro-CT) analysis revealed significant bone loss, thinned bone cortex, and enlarged marrow cavity in mice treated with Ti particles compared with control mice. Specifically, the bone volume fraction (BV/TV), a commonly used micro-CT parameter that measures the ratio of mineralized bone volume to total tissue volume and serves as an indicator of bone density and structural integrity, decreased by 1.9%, and trabecular number (Tb.N) decreased while trabecular separation (Tb.Sp) increased, all with statistically significant differences (p < 0.05, Fig. 1A), indicating Ti particle-induced osteolysis.
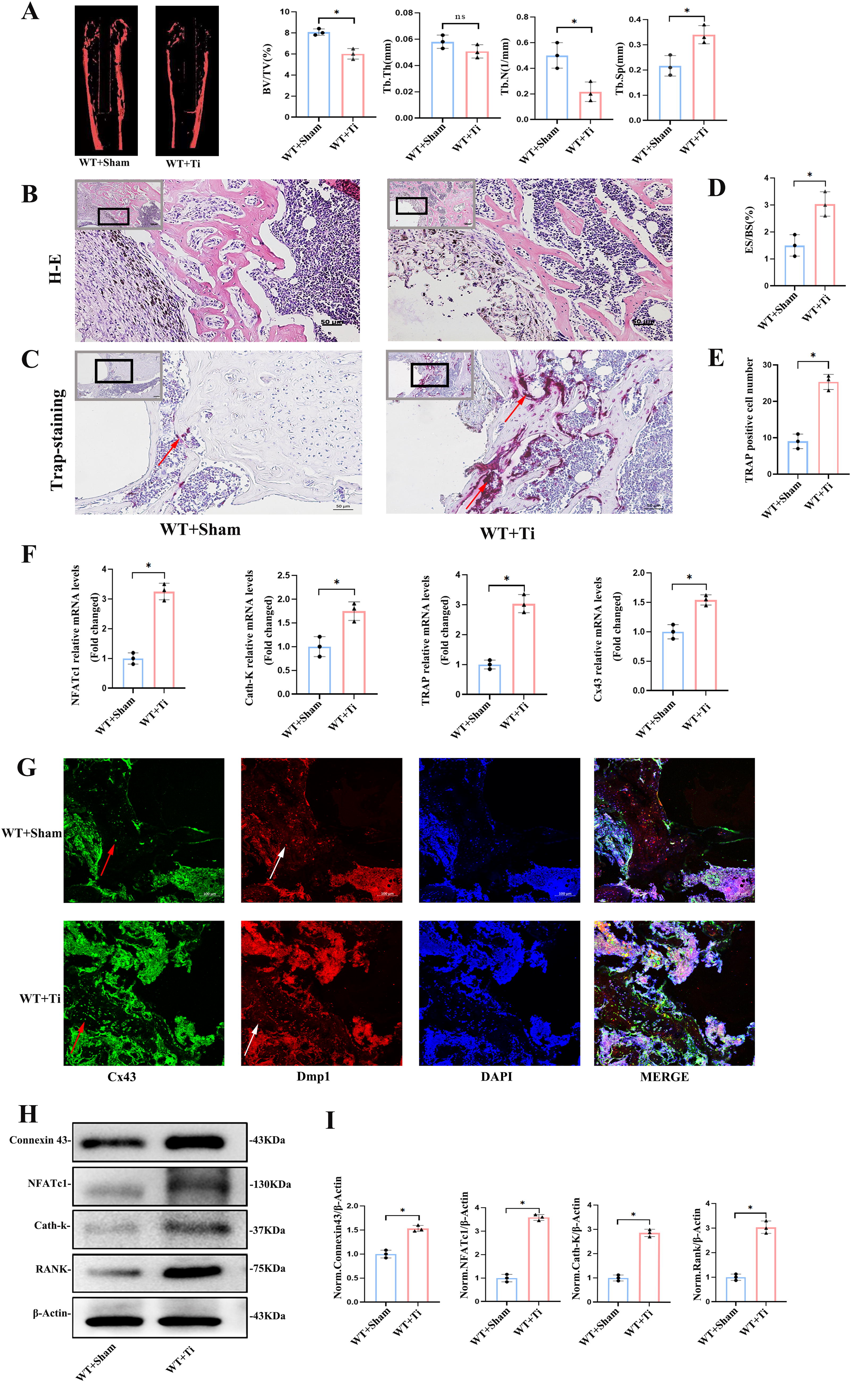
Histological analysis through H&E staining showed an obvious inflammatory reaction, macrophage-like cell infiltration, interruption of trabecular bone continuity, and aggravation of bone destruction at the interface between the hydroxyapatite (HA)-coated Ti rod prosthesis and the cancellous trabecular bone in the WT + Ti group (Fig. 1B). Bone histomorphometric analysis showed that the eroded surface/bone surface percentage was 1.5% ± 0.6% in the WT + Sham group and 3.0% ± 0.7% in the WT + Ti group, with a significant difference between the two groups (p < 0.05, Fig. 1D).
Tartrate-resistant acid phosphatase (TRAP) staining revealed more TRAP-positive osteoclasts in the Ti group, indicating enhanced osteoclastic activity (Fig. 1C), which was confirmed by elevated mRNA levels of osteoclast markers NFATc1 and Cath-K in the distal femur (p < 0.05, Fig. 1F). Additionally, Cx43 expression was significantly upregulated at both the mRNA and protein levels (p < 0.05, Fig. 1F and H).
As a characteristic marker, Dmp1 can facilitate to locate osteocyte in trabecular bone. The results of immunofluorescence staining of femoral tissue indicated an increased expression of Cx43 in trabecular bone following the intervention with Ti particles (Fig. 1G). This observation was supported by Western blot analysis of femoral tissue protein extracts, which revealed that the protein expression levels of Cx43, NFATc1, Cath-K, and RANK were significantly increased after the intervention with Ti particles. The differences were statistically significant in the WT + Ti group compared with the WT + Sham group (p < 0.05, Fig. 1H, I).
Osteocyte Cx43 deletion increased bone mass of cancellous bone and alleviated osteolysis induced by Ti particles in mice distal femur
The results above showed Ti increased Cx43 expression in the femur osteolysis model. To evaluate the effect of osteocyte-specific Cx43 deletion on bone mass and osteolysis, we scanned and three dimensionally reconstructed the distal femur using micro-CT. The results demonstrated that cancellous bone mass in the distal femur of CKO mice was significantly greater than that of WT mice within the same region of interest (ROI) (Fig. 2A). Statistical analysis showed a significant increase in BV/TV in the CKO group compared with the WT group (p < 0.05, Fig. 2B). Although trabecular thickness (Tb.Th) in the CKO group increased slightly, the difference was not statistically significant (p > 0.05, Fig. 2B). Additionally, compared with the WT group, the Tb.N in the CKO group increased, and Tb.Sp decreased, with both differences being statistically significant (p < 0.05, Fig. 2B). These findings indicated that osteocyte-specific Cx43 deletion increased cancellous bone mass in the distal femur.
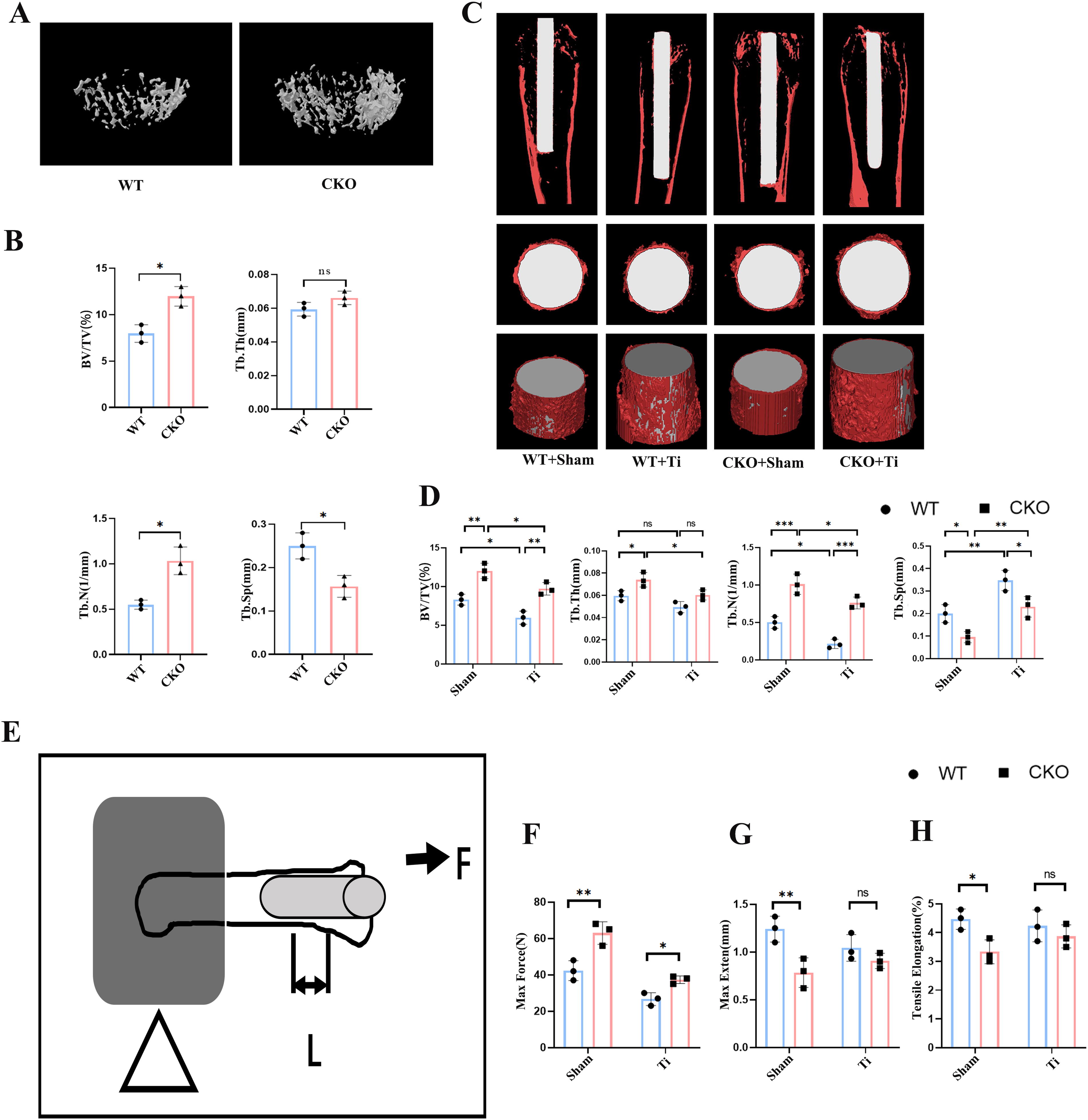
After establishing the osteolysis model, micro-CT scans and reconstructions revealed that bone mass in CKO + Sham mice was higher than in WT + Sham mice. In the WT + Ti group, BV/TV significantly decreased compared with the WT + Sham group, and a similar reduction was observed in the CKO + Ti group compared with CKO + Sham. However, BV/TV in the CKO + Ti group remained significantly higher than in the WT + Ti group (p < 0.05). Ti particle intervention also resulted in a significant decrease in Tb.Th in the CKO + Ti group compared with the CKO + Sham group (p < 0.05), but no significant difference was found between the WT + Ti and CKO + Ti groups (p > 0.05). Cancellous bone Tb.N increased and Tb.Sp decreased significantly in the CKO + Ti group compared with WT + Ti (p < 0.05, Fig. 2C and D).
To further assess the effect of Cx43 deletion on the integration between the Ti implant and the distal femur, biomechanical pull-out tests were performed after 2 weeks of implant placement (Fig. 2E). The maximum pull-out force in the WT + Sham group was 43.6 ± 3.5 N, whereas, in the CKO + Sham group, it was significantly higher at 66.5 ± 3.9 N (p < 0.05, Fig. 2F). Similarly, the pull-out force in the CKO + Ti group was higher than in the WT + Ti group (35.5 ± 3.1 N vs. 26.3 ± 2.8 N, p < 0.05). Maximum extension and tensile elongation were lowest in the CKO + Sham group, and the difference between WT + Sham and CKO + Sham was significant (p < 0.05, Fig. 2G and H). However, no significant differences in maximum extension or tensile elongation were observed between the WT + Ti and CKO + Ti groups (p > 0.05, Fig. 2J and K). These biomechanical results suggest that Cx43 deletion enhanced the pull-out resistance and improved the integration of the cancellous bone with the Ti implant.
These findings demonstrated that conditional knockout of the Cx43 gene alleviated bone loss around the prosthesis and improved cancellous bone remodeling in the Ti particle-induced osteolysis model.
Osteocyte Cx43 deletion decreased osteoclast differentiation and the expression of osteoclastic markers in mice osteolysis model
To investigate the role of osteocyte-specific Cx43 deletion in osteoclast differentiation and bone resorption in a mouse osteolysis model, bone marrow macrophages (BMMs) from the femoral marrow cavity were induced to differentiate into osteoclasts. F-actin rings, a key cytoskeletal component, serve as functional markers of osteoclast activation, closely associated with osteoclast migration and bone resorption. Immunofluorescence analysis revealed a significant reduction in the area and density of F-actin rings (green) in the CKO + Sham group compared with the WT + Sham group (Fig. 3A). Similarly, TRAP staining indicated fewer TRAP-positive osteoclasts in the CKO + Sham group after 7 days of induction, with a statistically significant difference (p < 0.05, Fig. 3B and C). Furthermore, the bone resorption area on tooth slices caused by osteoclasts was significantly smaller in the CKO + Sham group compared with the WT + Sham group (p < 0.05, Fig. 3B and D), indicating reduced osteoclast activity following Cx43 knockout in osteocytes.
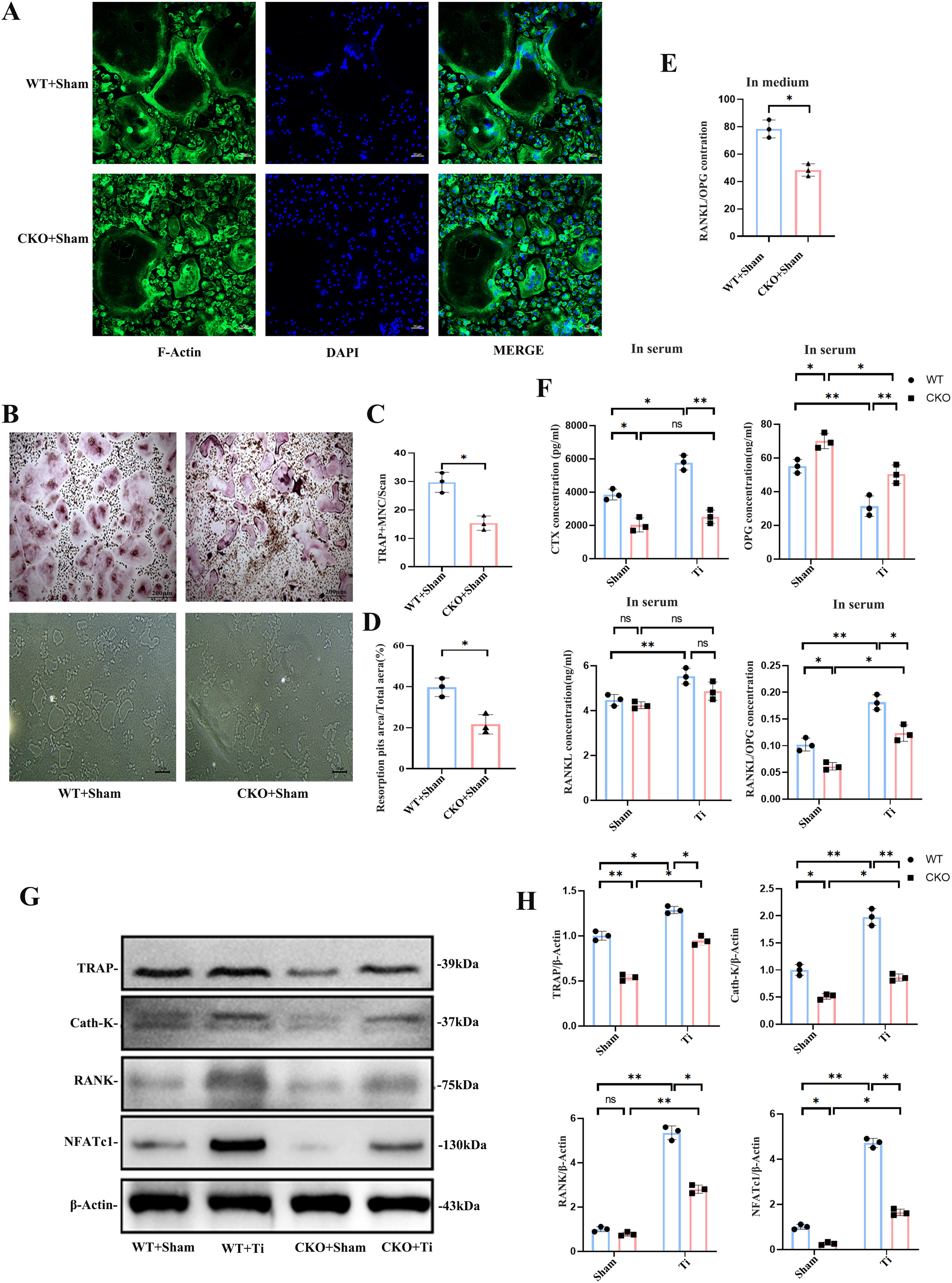
The RANKL/Osteoprotegerin (OPG) system plays a critical role in osteoclast activation. ELISA results showed that, compared with the WT + Sham group, the OPG content in the CKO + Sham group culture medium extracted from BMMs of femurs significantly increased on the 6th day, whereas the RANKL content increased slightly, leading to a significant decrease in the RANKL/OPG ratio (p < 0.05, Fig. 3E).
Peripheral blood analysis revealed that the C-terminal peptide of type I collagen (CTX), a marker of bone resorption, was significantly elevated in the WT + Ti group compared with the WT + Sham group. In contrast, CTX levels significantly decreased in the CKO + Sham group and in the CKO + Ti group compared with their respective controls (p < 0.05). The RANKL/OPG ratio also increased with the Ti particle challenge, but Cx43 knockout predominantly affected OPG levels, with a significant decrease in the RANKL/OPG ratio in the CKO + Ti group compared with the WT + Ti group (p < 0.05). This suggests that Cx43 knockout partially reversed the increase in CTX and the RANKL/OPG ratio during Ti particle-induced osteolysis (Fig. 3F).
Western blot analysis of distal femur protein expression further supported these findings. The expressions of TRAP, NFATc1, Cath-K, and RANK, key markers closely associated with osteoclast differentiation, were significantly upregulated in the WT + Ti group compared with the WT + Sham group (p < 0.05, Fig. 3G). However, these proteins were significantly downregulated in the CKO + Sham group compared with the WT + Sham group and similarly downregulated in the CKO + Ti group compared with the WT + Ti group (p < 0.05, Fig. 3G and H). These results indicate that osteocyte-specific Cx43 knockout reduces osteoclast activity and bone resorption in Ti particle-induced osteolysis.
Increased Cx43 induced by Ti particles in MLO-Y4 cells promoted osteoclast differentiation in vitro
To further confirm the effects of osteocytic Cx43 on osteoclast function under Ti particle stimulation, MLO-Y4 cells were treated with Ti particles in vitro. The conditioned medium was then collected and added to BMMs from C57BL/6 wild-type mice for 7 days to induce osteoclastic differentiation. TRAP staining and bone resorption assays on tooth slices were performed to evaluate osteoclast function.
The TRAP staining results showed that the Ti-treated group had significantly more TRAP-positive osteoclasts than the negative control (NC) group, which did not receive Ti particle intervention. However, after silencing Cx43 in MLO-Y4 cells, there was a significant reduction in the number of TRAP-positive osteoclasts, even in the presence of Ti particles. This difference was statistically significant among the three groups (p < 0.05, Fig. 4A and B). Furthermore, the bone resorption area caused by osteoclasts increased significantly in the Ti-treated group. Low expression of Cx43 in MLO-Y4 cells reduced the resorption area even under Ti particle intervention conditions (p < 0.05, Fig. 4A and C). These results suggest that the ability of BMMs to form osteoclasts decreased significantly after Cx43 silencing in MLO-Y4 osteocytes.
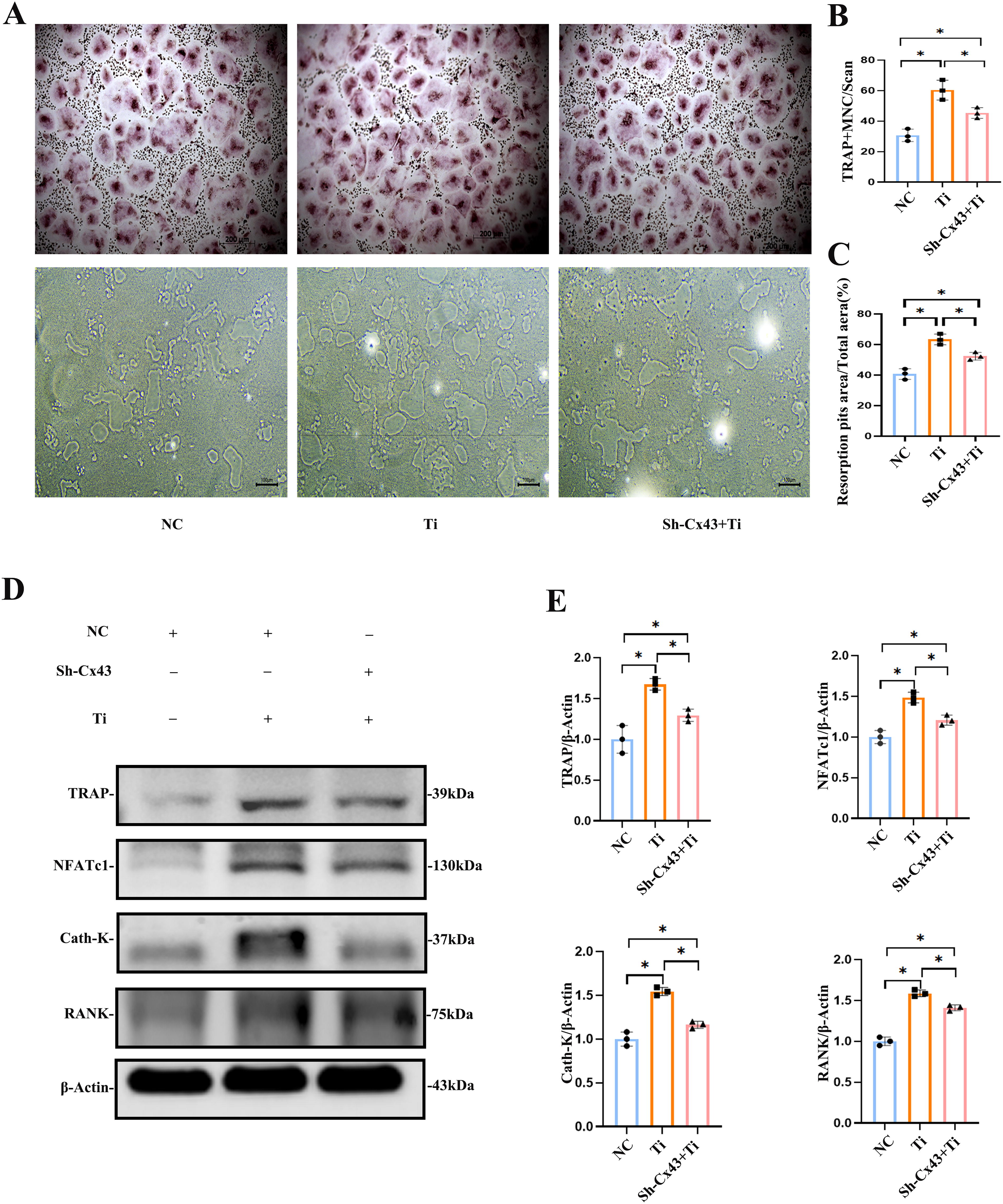
Similarly, the medium from MLO-Y4 cells, with or without Ti particle treatment, was added to BMMs for osteoclast induction. After 7 days of culture, the protein expressions of TRAP, NFATc1, Cath-K, and RANK were detected. The results showed that the expressions of these proteins were significantly upregulated in the Ti-treated group compared with the NC group (p < 0.05, Fig. 4D and E). In contrast, when Cx43 expression was low in MLO-Y4 cells treated with Ti particles, the expressions of TRAP, NFATc1, Cath-K, and RANK were significantly downregulated compared with the Ti-treated group (p < 0.05, Fig. 4D and E). These findings indicate that changes in MLO-Y4 cells induced by Ti particles stimulated osteoclast activation, whereas low expression of Cx43 in MLO-Y4 cells inhibited this activation.
Transcriptomic sequencing and KEGG pathway enrichment analysis in osteocytes revealed that Cx43 changes induced by Ti particles are linked to oxidative stress and the JAK-STAT pathway
To explore the molecular mechanisms underlying Cx43 knockdown in osteocytes, we performed transcriptomic sequencing on Sh-Cx43 MLO-Y4 cells and MLO-Y4 cells treated with 0.2 mg/mL Ti particles (high Cx43 expression) for 2 days. Oxidative stress was identified as a key factor in both conditions. Principal component analysis (PCA) showed clear separation between the treatment groups. In Figure 5A, PC1 and PC2 accounted for 50.21% and 17.73% of the variance, respectively, with the Sh-Cx43 group clearly separated from the NC, indicating significant gene expression changes. Similarly, in Figure 5B, PC1 and PC2 explained 34.91% and 22.16% of the variance, respectively, with the Ti-treated group distinct from the NC group. The tight clustering of samples in both groups confirmed the high reproducibility of the results. The PCA results highlighted significant transcriptomic changes induced by Cx43 knockdown and Ti particle treatment, providing insights into the underlying molecular mechanisms.
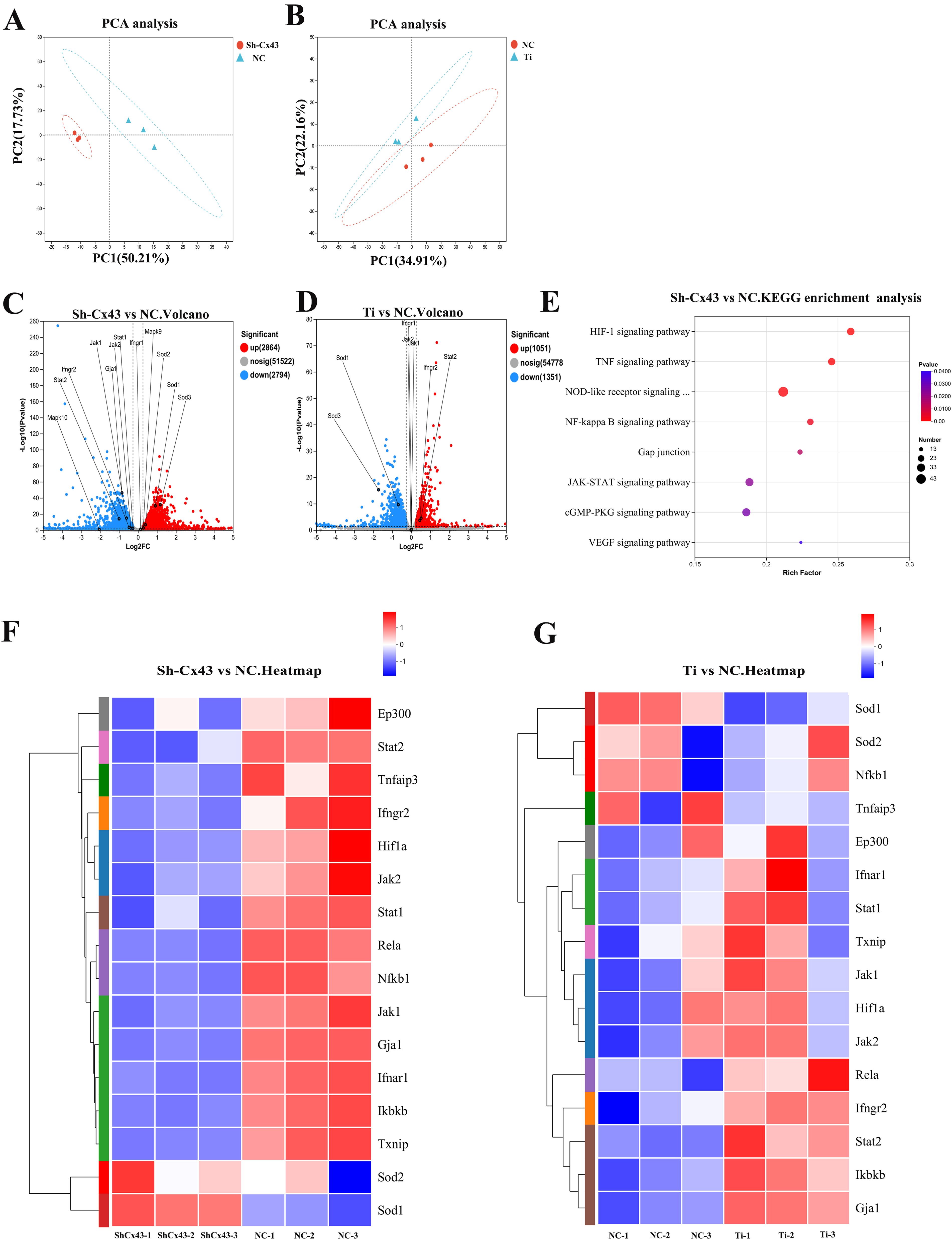
Volcano plots in Figure 5C and D depicted the differentially expressed genes (DEGs) between Sh-Cx43 vs. NC and Ti vs. NC, respectively. Significantly altered genes were highlighted, with the x-axis representing log2 fold change and the y-axis indicating statistical significance (−log10 p value). In the Sh-Cx43 group, the expression of STAT1, JAK1, JAK2, and IFNGR2 was reduced, whereas superoxide dismutase (SOD)1, SOD2, and SOD3 were upregulated. In contrast, Ti particle treatment decreased SOD1 and SOD3 expression while increasing IFNGR2 and STAT2 expression. Previous studies have shown that inhibiting IFNGR2 weakens the JAK-STAT pathway by reducing STAT1 phosphorylation, aligning with our findings (Xia et al., 2022).
Heatmaps in Figure 5F and G presented hierarchical clustering of differential gene expression between the various groups. Key genes involved in oxidative stress and the JAK-STAT pathway, such as STAT1, STAT2, JAK1, JAK2, IFNGR2, GJA1, SOD1, and SOD2, were highlighted in red ellipses, further supporting the role of these pathways in Cx43 knockdown and Ti particle treatment.
KEGG pathway enrichment analysis in Figure 5E revealed significant enrichment of pathways related to oxidative stress. The pathways enriched in both groups include HIF-1, PI3K-Akt, TNF, and NOD-like receptor, NF-kappa B, cGMP-PKG, VEGF, and JAK-STAT signaling pathways. Of course, we prioritized pathways with more significant enrichment. For detailed information, please refer to Supplementary Figure S2. However, upon investigating the DEGs enriched from these pathways, we found that, under the condition where Ti particles inhibit osteocyte antioxidant stress ability, only the JAK-STAT pathway can effectively enhance osteocyte antioxidant stress ability when Cx43 is downregulated. By analyzing the enriched pathways alongside the volcano plots and heatmaps, we identified that the JAK-STAT pathway plays a critical role in the oxidative stress response induced by Cx43 knockdown and Ti particles. This hypothesis was verified in subsequent studies.
Ti particles increased the Cx43 expression and the ratio of RANKL/OPG and induced oxidative stress in MLO-Y4 osteocytes
To investigate further the effects of Cx43 expression induced by Ti particles on osteoclast differentiation, the RANKL/OPG ratio was detected. We treated the cells with different concentrations of Ti particles for 2 days. The results showed an increase in Cx43 expression in MLO-Y4 osteocytes with 0.1 mg/mL and 0.2 mg/mL Ti particle stimulation. Notably, the expression of ITGα5 and ITGβ1, which regulate the opening of Cx43 channels, also increased with Ti particle stimulation. Further analysis demonstrated a gradual increase in RANKL/OPG expression with increasing concentrations of Ti particles, indicating that osteocytes may influence bone resorption during osteolysis (Fig. 6A and B).

To investigate the impact of Ti particles on oxidative stress resistance in MLO-Y4 osteocytes, Western blot, and immunofluorescence staining were performed. The results revealed that Ti particle stimulation significantly decreased the levels of NrF2 and HO-1, markers of cellular antioxidant defense, indicating a reduction in the cells’ ability to resist oxidative stress (Fig. 6A–C). To assess the production of ROS, we used dihydroethidium (DHE) staining, a superoxide anion indicator. DHE staining showed an increase in red fluorescence in the nuclei of osteocytes with Ti particle stimulation, indicating elevated levels of ROS (Fig. 6D). This suggests that Ti particles stimulate an increase in intracellular ROS levels in osteocytes.
Ti particles not only decreased the antioxidant capacity of osteocytes but also increased ROS production. The upregulation of Cx43, ITGα5, and ITGβ1 suggests a potential correlation between the change in Cx43 channels and the altered oxidative stress response in osteocytes under Ti particle stimulation.
Cx43-mediated GJ communication increased ROS levels and reduced antioxidant capacity in osteocytes
Cx43, the principal protein of GJs, plays a vital role in substance transmission and signal exchange among cells. 18α-GA, the common Cx43 channel inhibitor, was used to assess the GJ communication. To further explore the impact of Cx43 expression levels and 18α-GA treatment on GJ communication in MLO-Y4 osteocytes, we conducted dye transfer assays combined with immunofluorescence staining and flow cytometry analysis. We compared the effects of Cx43 knockdown (Sh-Cx43), overexpression (OE-Cx43), and 18α-GA treatment on GJ functionality. The results from immunofluorescence staining and flow cytometry consistently demonstrated that Cx43 expression levels significantly influence GJ communication in MLO-Y4 osteocytes (Fig. 7A–C). Treatment with 18α-GA and knockdown of Cx43 both resulted in a marked reduction in GJ functionality, as evidenced by decreased dye transfer. Conversely, overexpression of Cx43 enhanced GJ communication, indicated by increased dye transfer. These findings underscore the crucial role of Cx43 in regulating intercellular communication.
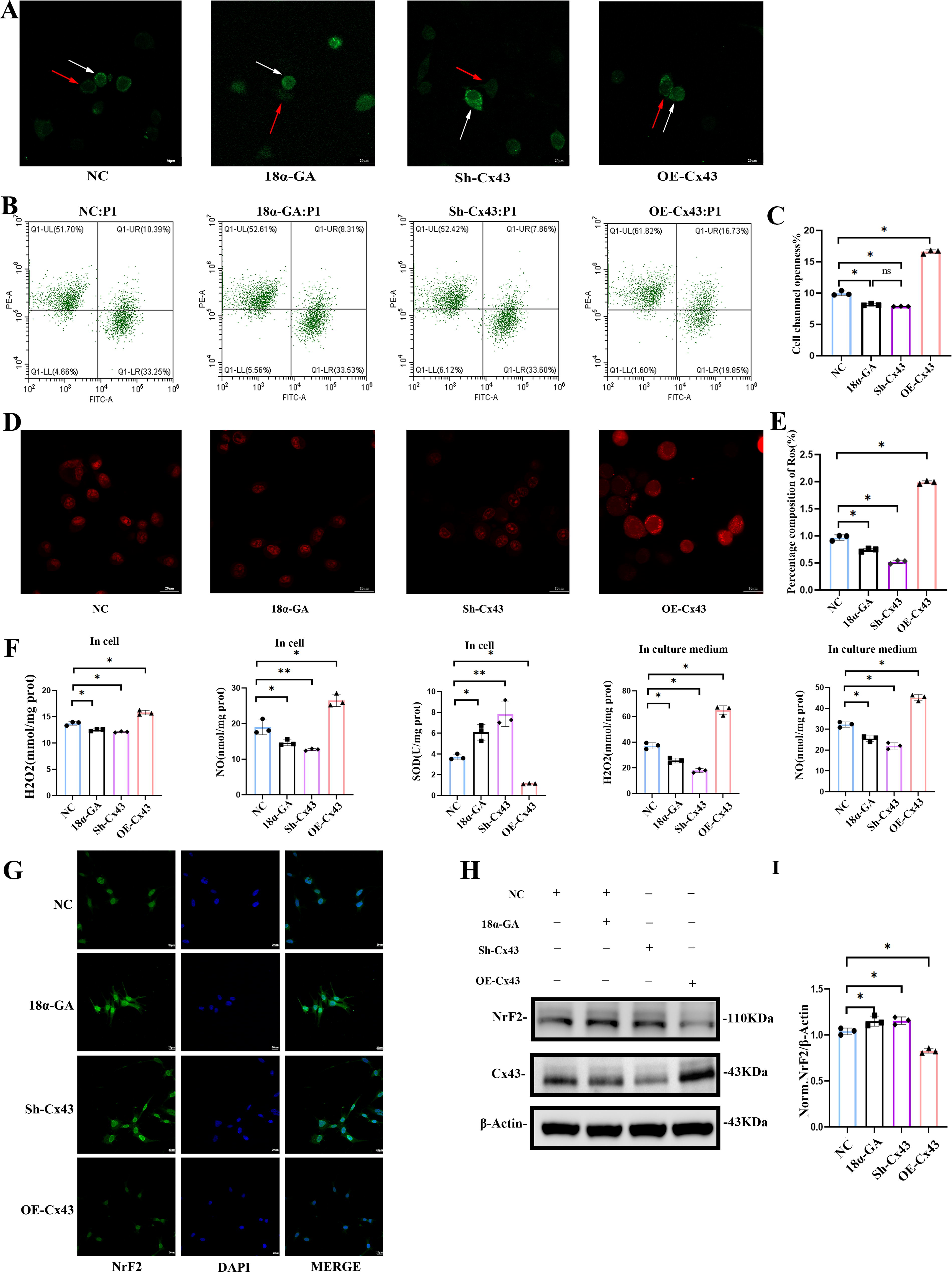
We further investigated the relationship between Cx43 and ROS in osteocytes. Flow cytometry and immunofluorescence analysis of DHE-stained cells revealed that reduced Cx43 expression and decreased channel opening both led to a reduction in intracellular superoxide levels. Conversely, overexpression of Cx43 increased intracellular superoxide levels (Fig. 7D and E, Supplementary Fig. S1).
Further analysis using H2O2, NO, and SOD assay kits demonstrated changes in various ROS indicators in both the cells and their culture media, as shown in Figure 7F. Increased Cx43 expression elevated intracellular and extracellular H2O2 and NO levels while reducing intracellular SOD content. In contrast, Cx43 knockdown and 18α-GA blocking exhibited opposite effects. These results suggest that Cx43 plays a crucial role in modulating ROS levels in osteocytes. NrF2 is a critical transcription factor regulating cellular antioxidant defenses. Immunofluorescence staining and Western blot analysis showed varying levels of NrF2 protein expression in different groups of osteocytes. Blocking Cx43 channel opening or Cx43 knockdown strengthened the antioxidant capacity of osteocytes, as evidenced by increased NrF2 protein levels, whereas Cx43 overexpression inhibited the antioxidant capacity (Fig. 7G–I).
Cx43 expression and channel blockade mediated STAT1 phosphorylation activates osteoclastogenesis by reducing NrF2 and enhancing RANKL/OPG ratio in osteocytes
As mentioned above, we speculated that the JAK-STAT pathway might play a role in the oxidative stress induced by Cx43 and Ti particles through transcriptomic and pathway enrichment analysis. To investigate this, we examined the role of phosphorylated STAT1 in MLO-Y4 cells under different conditions: 18α-GA treatment, Cx43 knockdown, and Cx43 overexpression. Western blot results revealed that phosphorylated STAT1 levels were significantly reduced in Cx43 knockdown and 18α-GA-treated cells compared with the control group, whereas Cx43 overexpression led to a significant increase in phosphorylated STAT1 levels (Fig. 8A and B). Notably, after 18α-GA treatment to block GJs, the total expression level of Cx43 in osteocytes did not show a significant change. However, phosphorylated STAT1 levels were markedly reduced, and NrF2 expression increased (Fig. 6H), which suggests that the GJs formed by Cx43 play a critical regulatory role in the activation of the JAK-STAT pathway.
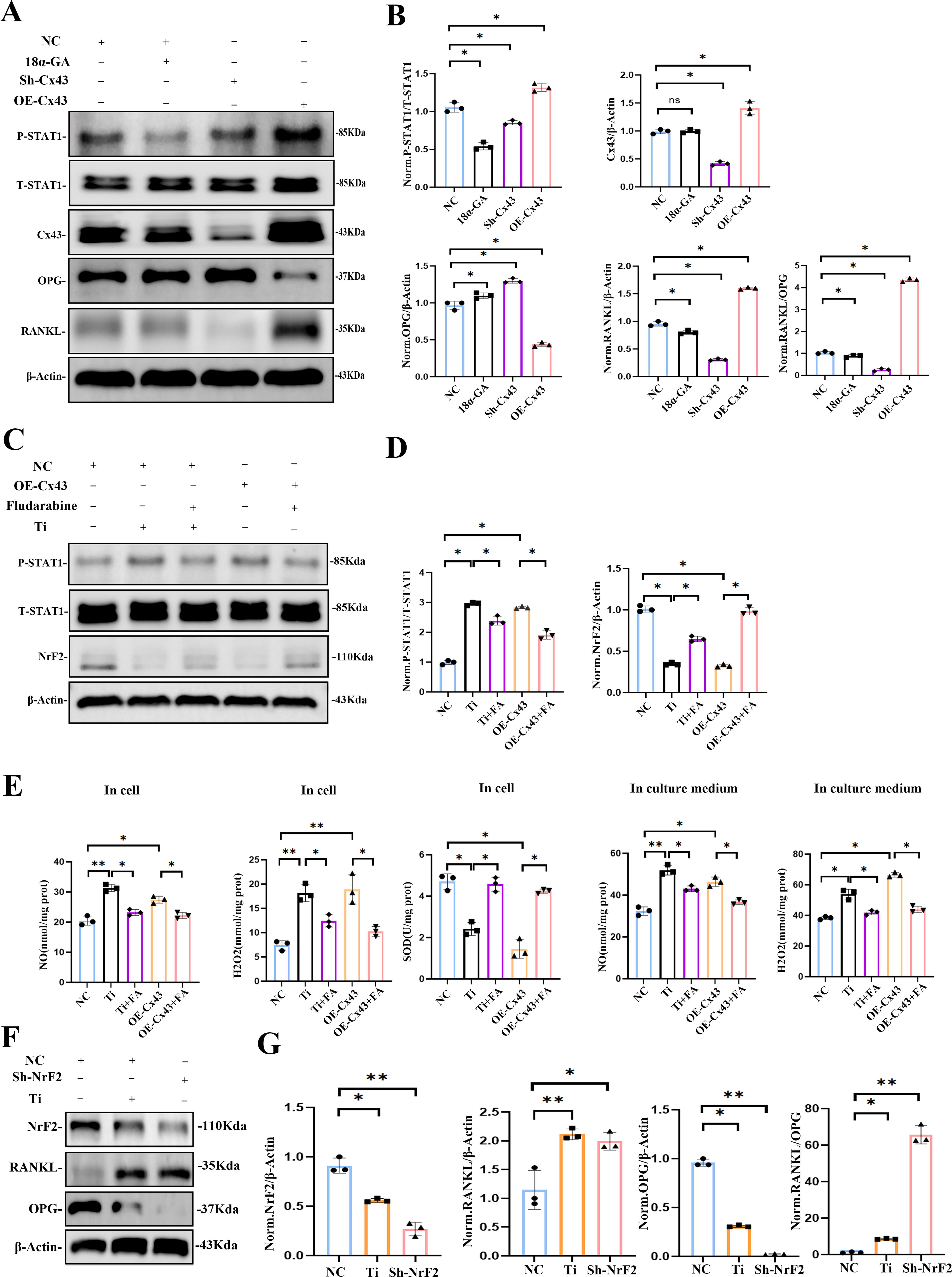
We also assessed the RANKL/OPG ratio, a key indicator of osteoclast activity regulation by osteocytes. Elevated phosphorylated STAT1 levels corresponded with a significant increase in the RANKL/OPG ratio, whereas reduced phosphorylated STAT1 levels decreased the ratio (Fig. 8A and B).
To further clarify the impact of phosphorylated STAT1 on the antioxidant capacity of osteocytes, we treated Cx43 overexpressing and Ti particle-stimulated MLO-Y4 cells with fludarabine (10 µM), an inhibitor of STAT1 phosphorylation. The results showed that both Ti particles and increased Cx43 expression promoted STAT1 phosphorylation and reduced NrF2 levels in osteocytes. However, fludarabine treatment significantly reduced phosphorylated STAT1 levels and increased NrF2 levels in both OE-Cx43 and Ti particle-treated cells, suggesting that STAT1 phosphorylation plays a key role in oxidative stress modulation (Fig. 8C and D).
Next, we quantified H2O2, NO, and SOD levels in both the culture medium and intracellularly under five different treatment conditions. Ti particle treatment and Cx43 overexpression elevated H2O2 and NO levels, whereas intracellular SOD levels were reduced. Fludarabine treatment effectively decreased H2O2 and NO levels while increasing intracellular SOD levels, with significant differences (p < 0.05; Fig. 8E). These results suggest that reducing STAT1 phosphorylation can inhibit ROS production induced by Ti particles and Cx43 overexpression.
Our findings also showed that Ti particles increase Cx43 expression, reduce NrF2 expression, and elevate the RANKL/OPG ratio. To further explore how oxidative stress affects osteoclast regulation, we performed NrF2 knockdown in MLO-Y4 cells. Western blot analysis indicated that NrF2 knockdown led to an increase in the RANKL/OPG ratio, consistent with previous results from Ti particle treatment and Cx43 overexpression (Fig. 8F and G).
Therefore, we concluded that the diminished antioxidant capacity in osteocytes, due to lower NrF2 levels, led to an increased RANKL/OPG ratio, which in turn promoted osteoclast activation.
Cx43 knockout decreased the phosphorylation levels of STAT1 and increased the expression of NrF2 in vivo
We further explore the impact of Cx43 on the antioxidant capacity in vivo through a calvarial osteolysis model of Cx43 conditional knockout mice. H&E staining confirmed the erosive impact of Ti particles on the calvaria, and immunohistochemical staining showed the changes in NrF2 and phosphorylated STAT1 (Supplementary Fig. S3). There were significant differences in NrF2 and phosphorylated STAT1 expression between WT + Sham and CKO + Sham mice, as well as between WT + Ti group and CKO + Ti group. In CKO mice, the increase in NrF2-positive osteocytes indicated an amplified oxidative stress response due to the absence of Cx43. Conversely, NrF2 expression decreased in Ti-treated WT mice. Additionally, the reduction in phosphorylated STAT1 levels in CKO mice underscored the role of Cx43 in regulating STAT1 activation. In both WT and CKO mice, Ti particle stimulation significantly elevated phosphorylated STAT1 levels, further demonstrating the critical role of STAT1 in oxidative stress and osteolysis (Fig. 9A and B).
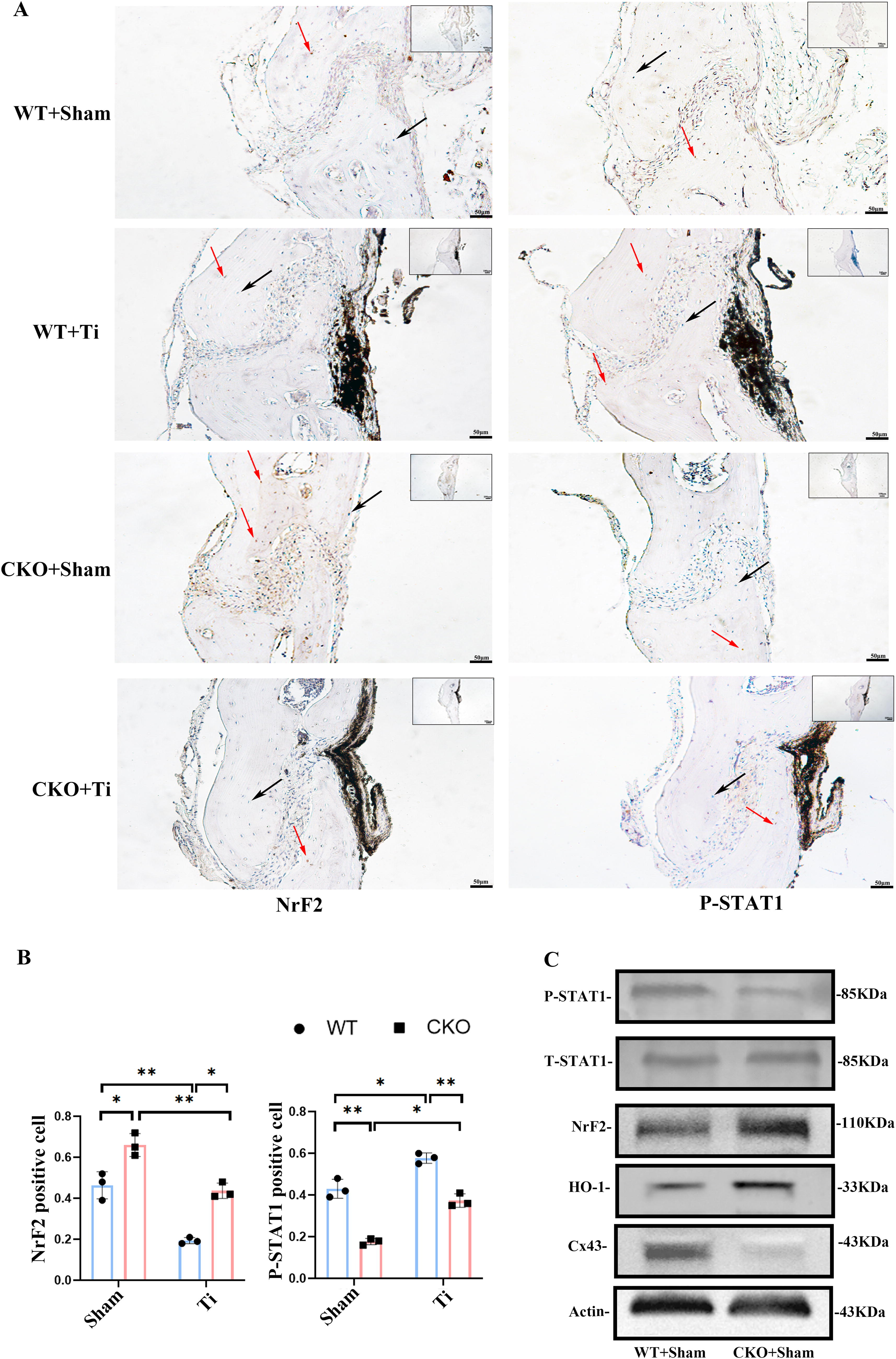
To further verify the changes in NrF2 and phosphorylated STAT1 in vivo, we extracted proteins from the femoral tissues of WT + Sham and CKO + Sham mice. Western blot analysis revealed significantly higher levels of NrF2 and HO-1 in CKO mice compared with WT mice, indicating that selective deletion of Cx43 in osteocytes enhanced the antioxidant capacity of the femur. Phosphorylated STAT1 levels were reduced in the femurs of the CKO + Sham group compared with the WT + Sham group, consistent with the known regulatory role of phosphorylated STAT1 in oxidative stress and inflammation (Fig. 9C).
Discussion
Numerous studies have shown that reduced osteogenic activity, combined with increased osteoclastic activity, leads to a dominance of bone resorption over formation, disrupting the balance of bone remodeling in wear particle-induced osteolysis (Xie et al., 2023; Zhang et al., 2020a). Our previous research demonstrated that osteocytes regulate osteoblast activity by binding Cx43 to β-catenin, thereby inhibiting Wnt pathway activation and suppressing osteoblast differentiation (Chai et al., 2022; Jiao et al., 2023; Zhang et al., 2020b). However, the role of osteocyte Cx43 in regulating osteoclast function remains unclear. This study investigates the mechanisms by which Cx43 in osteocytes influences osteoclast activity.
Osteocytes, the most abundant cells in bone, connect extensively with other osteocytes, osteoblasts, and osteoclasts via dendritic processes, playing a crucial role in bone remodeling (Bonewald, 2007; Clarke, 2008). Cx43, the most prevalent GJ protein in bone, forms hemichannels and GJs that mediate the transmission of small molecules and ions such as ATP, Ca2 +, cAMP, and ROS between cells and their environment, coordinating intercellular signaling and metabolic regulation (Herve and Derangeon, 2013). In a Ti particle-induced osteolysis model, we observed elevated Cx43 expression in regions marked by Dmp1, alongside increased osteoclast markers. Similarly, Ti particle treatment of MLO-Y4 cells in vitro resulted in heightened Cx43 expression and an elevated RANKL/OPG ratio, which is critical for osteoclast differentiation (Cheng et al., 2022; Xiong et al., 2022; Yasuda, 2021; Zaidi et al., 2023). These findings suggest a link between Cx43 expression and osteoclast activity. In Cx43 knockout mice, we observed improved trabecular bone regeneration, reduced serum CTX levels, and a lower RANKL/OPG ratio, which improved femoral implant biomechanical properties. In vitro, Cx43 knockout inhibited osteoclast formation from BMMs, providing further evidence that osteocytes regulate osteoclast function through Cx43, reducing bone resorption and mitigating Ti particle-induced osteolysis.
To explore the underlying mechanism, we performed transcriptome sequencing on Cx43-knockdown MLO-Y4 cells and Ti particle-treated cells (Cx43 overexpressed). KEGG enrichment analysis indicated that Cx43 expression influenced oxidative stress-related signaling pathways. The central role of oxidative stress has been widely recognized in neurodegenerative diseases and aging research. For instance, Calabrese et al. highlighted that Nrf2 and its downstream signaling pathways, including the vitagene family, play a crucial role in protecting cells from oxidative damage (Calabrese et al., 2005, 2010). Jiandong et al. also showed that Ti particles elevate ROS levels and suppress NrF2 in osteoblasts (Dong et al., 2022). Itgα5 and Itgβ1, which are associated with Cx43 channel opening (Batra et al., 2012, 2014), were also upregulated in Ti-treated cells, suggesting that Ti particles modulate Cx43 channel activity, linking Cx43 with oxidative stress. Previous studies revealed that increased Cx43 hemichannel opening promotes ROS production in fibroblasts (Fang et al., 2011), and Cx43 inhibition can protect tissues from ischemia-reperfusion injury by reducing oxidative stress and apoptosis (Theodoric et al., 2012). Further, we confirmed that Ti particles reduced NrF2 and HO-1 expression while increasing ROS levels, highlighting the role of Cx43 in oxidative stress modulation. Additionally, we demonstrated through dye diffusion and Western blotting that 18α-GA reduced the opening of Cx43 GJs in osteocytes without affecting the overall expression of Cx43. Furthermore, we found that low-expression Cx43 in osteocytes and treatment with 18α-GA effectively blocked Cx43 GJs, resulting in an increase in NrF2 expression and a reduction in ROS accumulation.
In our study, increased Cx43 protein in osteocytes promotes oxidative stress. However, the relationship between Cx43 and oxidative stress is still debated. Blocking Cx43 GJs has been shown to decrease intercellular ROS transmission, thereby preventing acute kidney injury, which suggests that Cx43 may exert a protective effect against oxidative stress (Yuan et al., 2019), whereas the study of Jiang et al. suggests that Cx43 low expression reduces oxidative stress (Zhang et al., 2022). Additionally, Jiang et al. found that Cx43 facilitates the exchange of oxidants and antioxidants in lens epithelial cells under oxidative stress, ultimately reducing intracellular ROS (Quan et al., 2021). These conflicting findings might be associated with different cell types and various microenvironments, and further research is needed to clarify Cx43′s exact role under different conditions.
Sequencing analysis also found that IFNGR2 expression, which regulates STAT1 phosphorylation in the JAK-STAT pathway linked to oxidative stress (Lin et al., 2017), was reduced in Cx43-knockdown cells but elevated in Ti-treated cells. We thought that the JAK-STAT pathway might play a significant role in osteocytes’ response to oxidative stress induced by wear particles. By inhibiting STAT1 phosphorylation with fludarabine, we observed increased NrF2 levels and decreased ROS in MLO-Y4 cells. This was corroborated in vivo, where phosphorylated STAT1 and NrF2 levels in mouse cranial and femoral tissues confirmed STAT1’s role in oxidative stress regulation. Other studies also showed that inhibiting STAT1 phosphorylation decreases RANKL secretion and increases OPG expression (Yan et al., 2019). However, their study did not clarify the specific relationship between STAT1 and RANKL/OPG secretion in osteocytes. Huang et al. reported that the accumulation of ROS can activate p38 MAPK and JNK phosphorylation in osteoblasts, leading to increased RANKL secretion (Yu et al., 2017). In our study, we observed that 18α-GA treatment, while not altering the total expression level of Cx43 in osteocytes, inhibited the opening of GJs between adjacent osteocytes. This treatment also reduced STAT1 phosphorylation, increased NrF2 expression, and ultimately led to a decrease in RANKL/OPG secretion, inhibiting osteoclast activation. Osteocytes stably transfected with low-expressed Cx43 also exhibited a reduction in the opening of GJs between osteocytes, showing similar effects to those observed with 18α-GA treatment. This finding further supports our conclusion that the increased Cx43 expression and opening of Cx43 GJs activate the JAK-STAT pathway to suppress NrF2 expression, ultimately reducing the antioxidant stress capacity of osteocytes. Although 18α-GA primarily blocks GJs, some studies have reported that it also inhibits Cx43 hemichannels. The observed changes in extracellular NO and H2O2 levels before and after 18α-GA treatment may suggest that Cx43 hemichannels also play a role in this process. To confirm that Ti particle-induced overexpression of Cx43 in osteocytes could drive the increase in RANKL/OPG by inhibiting NrF2, we generated NrF2 knockdown MLO-Y4 cells. Our study found that suppression of NrF2 in these cells resulted in an elevated RANKL/OPG ratio, providing a reasonable explanation for the increased RANKL/OPG secretion and enhanced osteoclastic activity observed in osteocytes following Ti particle stimulation in our previous experiments. Zhang et al. found that NrF2 inhibition in osteocytes downregulates DNA methyltransferase 3a (Dnmt3a) at the transcriptional level, ultimately upregulating RANKL expression in osteocytes and promoting osteoclast activation (Jiang et al., 2024). This is consistent with our findings.
In sum, our findings demonstrate that in Ti particle-induced osteolysis, high-expression Cx43 and the increased opening of GJs enhance the intercellular exchange of small molecules between adjacent osteocytes, which activates the JAK-STAT pathway, further suppressing NrF2 expression and enhancing ROS levels. Ultimately, these changes lead to an increase in RANKL/OPG secretion by osteocytes, promoting osteoclast activity and exacerbating bone resorption. This study is the first to reveal that osteocytes regulate oxidative stress and the RANKL/OPG ratio through Cx43 GJs, thereby influencing osteoclast activity. However, our study has limitations, as it primarily focused on GJs without fully exploring the potential role of Cx43 hemichannels, which may also contribute to oxidative stress modulation (Shahidullah and Delamere, 2014). Additionally, the involvement of specific second messengers, such as ROS or other small molecules, in JAK-STAT pathway activation and the potential direct effects of elevated ROS on osteoclast activation warrant further investigation.

Materials and Methods
Preparation of Ti particles
Ti nanoparticles were purchased from Nanjing Emperor Nano Materials Company. The particles that were 24.51–233.58 nm in size were sterilized by baking at 180°C for 6 h, followed by treatment with 70% ethanol for 48 h. The particles were then stored in PBS at 4°C at a concentration of 40 mg/mL. Prior to the experiments, the stock solution was sonicated for 10 min and diluted with a medium to 0.1 mg/mL or 0.2 mg/mL for the in vitro study, and to 40 mg/mL for the in vivo study. When used in cell cultures and animal experiments, such particles have been shown to effectively mimic wear particles retrieved from periprosthetic tissue (Chai et al., 2023). The particles were prepared as previously described by Chen et al. (2014). The absence of endotoxins was confirmed using the QCL-1000 Endotoxin Assay Kit (Biowhittaker). The sensitivity of this method is 0.005 EU/mL, which enables the detection of low levels of endotoxin in the preparations. The upper limit of detectable endotoxin content using this method is typically around 1 EU/mL.
Preparation of conditional Cx43 knockout mice
All procedures were approved by the Ethics Committee for Animal Care and Use of the Research Center for Experimental Medicine of Second Affiliated Hospital of Soochow University. The global knockout of Cx43 is embryonically lethal. In the in vivo experiment, we utilized mice with a conditional deletion of the Cx43 encoding gene Gja1 in osteocytes. Knockout mice were provided by GemPharmatech Co. Ltd. The obtainment of CKO mice proved to be effective, and the detailed protocol was described in our previous study (Chai et al., 2022). Briefly, dentin matrix acidic phosphoprotein 1 (Dmp1) as a marker of osteocytes, we obtain Cx43 fl/wt (WT) and Dmp1-cre-Cx43 flox/wt mice by mating Dmp1-cre mice with Cx43 flox/wt mice. Finally, Cx43 flox/flox target mice were obtained by mating Dmp1-cre-Cx43 flox/wt mice with Cx43 flox/wt mice. Thirty 6–8 weeks old male C57BL/6 wild-type mice (WT group) and thirty 6–8 weeks old Dmp1-cre-Cx43 flox/flox mice (CKO group), each weighing an average of 20–25 g, were included in the study. All mice were fed a standard laboratory diet and provided tap water ad libitum under climate-controlled conditions.
Mouse femoral and calvarial Ti-particle osteolysis model
We established concurrent femoral and calvarial osteolysis models in the same group of mice. A total of 60 mice, including 30 WT and 30 CKO mice, were randomly allocated into four groups: WT + Sham, WT + Ti, CKO + Sham, and CKO + Ti, with 15 mice in each group (Table 1).
Experimental Groups for Mouse Femoral and Calvarial Titanium-Particle Osteolysis Model
Based on the experimental design and random assignment principles, it is estimated that 12 mice are required per group. To prevent uncontrollable factors such as mice death or errors in operation from affecting the process of the experiment, we have set the number of mice at 15 for each group (with an additional 3 mice per group).
For the femoral model, the mice received an implant insertion into the left distal femur (HA-coated Ti rod, diameter 0.8 mm, and length 5 mm). HA, with a hexagonal structure, mimics natural bone and supports cell adhesion and osteogenesis. Nano-sized HA enhances osteoblast activity, ideal for bone repair. Composed of calcium phosphate (Ca10(PO4)6(OH)2) with a Ca/P ratio of 1.67, HA adsorbs ions such as Sr2 +, Mg2 +, and Zn2 + to modulate bioactivity. It dissolves slowly, aiding bone regeneration. For details, refer to the study by Hacking et al. (2010). Prior to implant insertion, the WT + Ti and CKO + Ti groups were administered 50 µL of Ti particle solution (40 mg/mL) into the medullary cavity of the left distal femur, whereas the WT + Sham and CKO + Sham groups received 50 µL of PBS. Subsequently, for the calvarial osteolysis model, the skull surfaces were exposed and the periosteum was completely removed. The WT + Ti and CKO + Ti groups received subcutaneous injections of 40 µL of Ti particles (40 mg/mL) diluted in PBS around the middle suture, whereas the WT + sham and CKO + sham groups received 40 µL of PBS. Two weeks postsurgery, femoral and calvarial specimens, along with peripheral blood samples, were collected for subsequent analysis. In our study, C57BL/6 mice were anesthetized via intraperitoneal injection of 2% pentobarbital at a dose of 60 mg/kg body weight. After ensuring deep anesthesia, euthanasia was performed using cervical dislocation to ensure humane sacrifice.
Micro-CT analysis
Femoral specimens of surgery mice (n = 4 per group) were fixed in 4% paraformaldehyde and were analyzed by a Micro-CT (SkyScan 1076) set with 9 µm per layer. The X-ray parameters were set at 100 kV and 100 µA. Micro-CTs were taken of a hollow cylindrical region of interest (ROI of 3.14 × [62–42] × 10 mm3) located from the epiphyseal line of the distal femur to the proximal end with the Ti rod as the center. Meanwhile, the femoral specimens without surgery were treated the same as above. A cylindrical region was selected as ROI from the epiphyseal line of the distal femur to the proximal end with the medullary cavity as the center (ROI of 3.14 × 62 × 10 mm3). The following data were collected: bone volume/tissue volume (BV/TV), thickness of trabeculae (Tb.Th), number of trabeculae (Tb.N), and space of trabeculae (Tb.Sp). Three micro-CT scan results of specimens were selected for data analysis for each group (n = 3).
Biomechanical testing
The distal femurs were wrapped in normal saline-soaked gauze, and the specimens were stored in a plastic bag in refrigerator at −20°C for biomechanical test within 1 h. The prosthesis-implanted femoral specimens of WT + Sham group, CKO + Sham group, W + Ti group, and CKO + Ti group (n = 3) were fixed on the support of the tester, the distal implant end was fixed with the test collet, the femoral position was adjusted to make the implant axis consistent with the pull-out force line, and the implant was pulled out at a uniform speed until it was pulled out. The extraction–deformation curve was drawn by computer. The pull-out measurement accuracy is 0.01 N, the displacement measurement accuracy is 0.001 mm, and the loading speed is 2 mm·min−1. Relevant experimental data including maximum force (N), maximum extension (mm), and elongation (%) were read and analyzed from the extraction–deformation curve.
H&E staining and IHC
Following a 2-day incubation in formalin, femur and calvariae samples from osteolysis models of four mouse groups (WT + Sham, WT + Ti, CKO + Sham, and CKO + Ti; n = 3 per group) underwent decalcification in ethylenediaminetetraacetic acid (EDTA; Sigma-Aldrich, Burlington, MA, USA) for 1 month. The specimens were subsequently trimmed and selected. After dehydration and paraffin embedding, the calvariae samples were sectioned into 2 µm slices, whereas the femur samples were sectioned into 5 µm slices for H&E staining. The calvariae sections underwent both H&E and immunohistochemical staining according to standard protocols. For immunohistochemistry (IHC), primary antibodies against NrF2 (Proteintech, 16396-1-AP) and P-STAT1 (Cell Signaling Technology, 9167) were utilized. The sections were subsequently examined under a light microscope (Zeiss, Oberkochen, Germany). Three slides from each group were chosen for quantitative analysis. For the calvariae samples, ImageJ software was employed to calculate the proportion of positive (brown) cells, determined as follows: proportion of positive cells = (number of positive cells in cranial bone plate/total number of cells in cranial bone plate) × 100%.
ELISA test for OPG and RANKL
The first group of samples from culture media of the BMMs extracted from the femurs of WT + Sham and CKO + Sham mice (n = 3 per group). The media were collected on the 6th day of culture for analysis. The second group of samples was peripheral blood serum collected from WT + Sham, WT + Ti, CKO + Sham, and CKO + Ti mice (n = 3 per group). The blood samples were centrifuged to separate the serum.
OPG and RANKL were quantified with ELISA kits (Immunodiagnostic Systems Inc) according to the manufacturer’s protocols and using an epoch Microplate Spectrophotometer (BioTek Instruments, Inc). The peripheral blood serum collected from WT + Sham, WT + Ti, CKO + Sham, and CKO + Ti mice was tested for CTX (BioTek Instruments, Inc.), RANKL, and OPG using ELISA kits.
Cell cultures and Ti particle intervention
MLO-Y4 cells, an osteocyte cell line, were procured from the Cell Bank of the Chinese Academy of Sciences. The cells were cultured on collagen-coated dishes (rat tail collagen type I, Sigma-Aldrich) and maintained in αMEM (Gibco) supplemented with 5% fetal bovine serum (FBS, Gibco), 5% calf serum (Gibco), and 1% penicillin-streptomycin in a 5% CO2 atmosphere at 37°C. Twenty-four hours postseeding, the MLO-Y4 cells were treated with 0.1 and 0.2 mg/mL Ti particles, whereas the control group remained untreated. Fresh medium was replenished every 3 days for all cell cultures. The addition of Ti particles to the cells was designated as day 0. Subsequent tests were conducted 24 h after culture initiation.
To evaluate osteoclastic differentiation and function, we employed a modified version of a previously described protocol for extracting and inducing differentiation of primary bone marrow-derived monocyte-macrophages (BMMs), for detailed protocols, please refer to our previous study (Jiao et al., 2023). Cells were cultured for 7 days and then stained for TRAP using the Leukocyte Acid Phosphatase kit (Sigma-Aldrich, St. Louis, MO), and the characteristic markers of osteoclasts were detected.
MLO-Y4 osteocytes were treated with Ti particles before and after Cx43 silencing. Twenty-four hours post-Ti intervention, the medium was collected and centrifuged to remove cellular debris, Ti particles, and other impurities. The medium was then added to BMMs (half-medium changed every 3 days), inducing osteoclastic differentiation for 6–8 days, after which the characteristic markers of osteoclasts were detected.
Cx43/NrF2 silencing or overexpression in MLO-Y4
Following the manufacturer’s instructions, MLO-Y4 osteocytes were subjected to Cx43 gene silencing using short hairpin RNA (shRNA) lentiviral particles. The sequences for Cx43 and NrF2 genes were first retrieved from GenBank, and three shRNA interference sequences targeting Cx43 and NrF2 were designed using Invitrogen’s RNA interference technology and synthesized by Jima Gene Co., Ltd. The cells were transduced with lentiviral particles carrying either scrambled shRNA or specific shRNA targeting Cx43. The efficacy of the shRNA was evaluated by measuring Cx43 protein expression via Western blotting. The most effective Cx43-shRNA sequence, 5′-GGTGTCTCTCGCTCTGAATAT-3′, and the most effective NrF2-shRNA sequence, 5′-GCAACTGTGGTCCACATTTCC-3′, were selected. MLO-Y4 osteocytes with silenced Cx43 and NrF2 were then used as the Cx43-silenced and NrF2-silenced groups in subsequent in vitro experiments. Similarly, Cx43 overexpression lentivirus was obtained from Jima Gene Co., Ltd. The Cx43 overexpression cell line was established in the same manner as GJA-1 silencing.
RNA-seq and DEG analysis in MLO-Y4
Total RNA was extracted from MLO-Y4 cells transfected with Sh-Cx43 and NC, as well as from MLO-Y4 cells pretreated with Ti particles (0.2 mg/mL) for 2 days, using Trizol reagent (Invitrogen). RNA-seq analysis was then conducted at Majorbio (Shanghai, China) according to the standard Illumina protocol. The RNA products were purified using the AMPure XP system, and library quality was assessed using the Agilent Bioanalyzer 2100 system. Clusters were generated using the TruSeq PE Cluster Kit v3-cBot-HS, and sequencing was performed on the Illumina HiSeq 2500 platform. Data analysis was carried out in the R programming environment. Differential expression analysis was conducted using the DESeq2 package, with significantly DEGs identified based on an adjusted p value <0.05 and a log2-fold change >1.2. Subsequently, PCA was conducted to evaluate the correlations among the samples. Separately, differential gene expression analysis was performed, and volcano plots were generated. Target gene sets were created from the DEGs for subsequent clustering analysis and KEGG pathway enrichment analysis.
TRAP staining and dentine resorption assay
The femurs and tibias of mice were immediately harvested postsacrifice and embedded in paraffin according to previously described protocols (Loiselle et al., 2013). Sections (n = 3 per group) were stained using a TRAP kit (Sigma-Aldrich) following standard protocols. The TRAP staining experiment comprised three groups: (1) distal femur sections from WT + Sham and WT + Ti group (n = 3 per group); (2) osteoclasts induced from BMM cells in the femoral marrow cavity of WT and CKO mice (n = 3 per group); and (3) BMM cells from the bone marrow cavity of WT mice cocultured with MLO-Y4 conditioned medium (n = 3 per group). All cells were cultured and induced for osteoclastogenesis according to previously described methods. The relative abundance of TRAP staining was assessed at a magnification of 100× using Image-Pro Plus 6.0 software (Image-Pro Plus 6.0, Media Cybernetics, MD, USA).
For the bone resorption dentine disc assay, primary bone marrow cells were isolated from mice. On the second day, nonadherent BMM cells were collected and seeded into a 24-well bone resorption dentine disc culture plate (Corning, USA) at a density of 5 × 103 cells per well. Osteoclastogenesis was induced as described previously, with the indirect coculture groups supplemented with conditioned medium from the corresponding groups of MLO-Y4 cells. After 5 days of culture, the medium was adjusted to pH 7.0 and then discarded. The bone resorption dentine discs were washed with PBS, air-dried, observed under an inverted microscope, and photographed to capture images of bone resorption lacunae. Analysis was conducted using the microscope computer image analysis system (Image-Pro Plus 6.0), and the bone resorption lacuna area of each well (%) was calculated as follows: bone resorption lacuna area/culture well area (resorption lacuna area/total area).
Immunofluorescence staining
IF of femur sections and F-actin in osteoclasts
A standard immunofluorescence method was used on decalcified sections of femur and osteoclast culture dish. Briefly, the sections and culture dishes were blocked with 5% normal goat serum and incubated with a 1/100 dilution of Cx43, Dmp1 antibody at 4°C overnight, followed by DyLight 594 goat anti-rabbit IgG secondary antibody. After cellular nuclei were counterstained with DAPI, the digital images were captured using a confocal microscope (Leica TCS SP8 MP, Wetzlar, Germany).
BMMs were seeded into confocal culture dishes and subsequently induced to differentiate into osteoclasts. The differentiated osteoclasts were then fixed and permeabilized following established protocols. The osteoclasts were incubated with ActinGreen™ 488 ReadyProbes Reagent (Invitrogen, R37110, two drops per 1.5 mL PBS) for 30 min at room temperature in the dark. Following PBS washes, the nuclei were stained with DAPI (1:1000 dilution) for 5 min. The F-actin rings were subsequently visualized using an immunofluorescence microscope (ZEISS).
IF detection of NrF2 and HO-1 in MLO-Y4 cells
MLO-Y4 osteocytes were fixed in 4% paraformaldehyde for 10 min, permeabilized with 0.1% Triton X-100 for 5 min, and blocked with 5% goat serum for 1 h in 0.1% PBS-Tween (PBST) containing 5% bovine serum albumin (BSA). The cells were then incubated with NrF2 (Proteintech, 16396-1-AP) primary antibody diluted 1:300 and HO-1 (Abcam, AB223349) primary antibody diluted 1:100 in 2% goat serum overnight at 4°C. After incubation, the cells were washed three times with PBST and incubated with goat anti-rabbit IgG H&L (Alexa Fluor 488, Abcam, AB150077) and goat anti-mouse IgG H&L (Alexa Fluor 647, Abcam, AB150115) secondary antibodies (both at 1:500 dilution) for 1 h in the dark. The nuclei were stained with DAPI (Beyotime, C1002) for 5 min after being washed three times with PBST. The stained cells were observed under a laser scanning confocal microscope LSM880 (Zeiss, Oberkochen, Germany) using excitation wavelengths of 488 and 647 nm.
Parachute dye-coupling assay for GJ evaluation
Functional GJIC was examined with “Parachute” dye-coupling assay as described (Fonseca et al., 2006; He et al., 2009): DiIC18(3), a lipophilic dye that labels cell membranes, and BCECF-AM, a dye that is easily retained in the cytoplasm after esterase cleavage of the acetoxymethyl ester inside the cell. The resulting compound has a molecular weight of 520 Da, allowing it to pass through GJs (Fonseca et al., 2006).
The donor cells were incubated with BCECF-AM (0.5 µM; DOJINDO; B262) at 37°C for 30 min. Recipient cells were labeled with the lipophilic dye DiIC18(3) (10 µM; Thermo Fisher; D282) at 37°C for 1 h. After incubation, the cells were washed five times with PBS and trypsinized with Trypsin-EDTA for 5 min. The donor and recipient cells were cocultured at a 1:3 ratio in α-MEM medium containing 10% FBS for the same duration. The cells were allowed to attach to the monolayer of recipient cells and form GJs at 37°C and pH 7.4 for 4 h and then observed under a flow cytometer. The average number of recipient cells containing BCECF-AM per donor cell was considered an indicator of the degree of GJIC.
RNA extraction and qPCR
Total RNA was extracted from distal femoral bones that were flushed of bone marrow and subsequently flash-frozen in liquid nitrogen, as described previously. The bones were then pulverized using a tissue pulverizer. Total RNA was extracted using Trizol reagent (Invitrogen) and subsequently purified with Phase-Lock gel tubes and the Qiagen RNA Easy kit, following the manufacturer’s instructions. Contaminating DNA was eliminated using RNase-free DNase treatment. RNA concentration and purity were determined using a NanoDrop spectrophotometer (with A260/A280 ratios between 1.8 and 2.0 indicating acceptable quality), and RNA integrity was confirmed by agarose gel electrophoresis. One microgram of total RNA was reverse transcribed using the Superscript VILO cDNA synthesis kit (Invitrogen). Real-time PCR analysis was conducted on an Applied Biosystems 7500 Fast detection system using the SYBR Green PCR method, according to the manufacturer’s instructions (Applied Biosystems). PCR cycling conditions were as follows: an initial denaturation at 95°C for 10 min, followed by 40 cycles of 95°C for 15 s, 60°C for 30 s, and 72°C for 30 s; a dissociation step was performed to verify the specificity of the amplification. The mean cycle threshold (Ct) value from triplicate samples was utilized to calculate gene expression, and PCR products were normalized to GAPDH levels in each reaction. Relative gene expression levels were determined according to the protocol described in the User’s Bulletin (P/N 4303859) from Applied Biosystems. TaqMan Gene Expression Assays (Applied Biosystems) were employed for the quantification of Cx43, NFATc1, and Cath-K, following the manufacturer’s instructions (Applied Biosystems).
Western blot of cell and femoral tissue proteins
MLO-Y4 cells were cultured in 6-well plates and exposed to varying concentrations of Ti particles for 48 h. Similarly, BMMs were evenly seeded into 6-well plates and subjected to the same treatment protocol. The cells were washed twice with PBS, lysed with lysis buffer, and incubated on ice for 20 min. The lysates were subsequently centrifuged at 14,000 rpm for 10 min. The supernatant was collected, and protein concentrations were determined using the BCA protein assay kit (Beyotime, Shanghai, China, P0010). Approximately 30 µg of protein samples was separated by 10% SDS-PAGE and transferred onto PVDF membranes preactivated by methanol soaking. The membranes were blocked with 5% BSA (Sangon Biotech, Shanghai, China, 4240GR100) for 1 h, followed by overnight incubation with primary antibodies diluted (see Table 2).
Primary Antibodies Used for Western Blot Analysis in MLO-Y4 Cells and Bone Marrow Macrophages Treated with Titanium Particles
The membranes were washed four times with Tris-buffered saline with Tween (TBST) and incubated with horseradish peroxidase (HRP)-conjugated goat anti-mouse IgG (Proteintech, China, SA00001-1) and goat anti-rabbit IgG (Proteintech, China, RGAR001), followed by further washing with TBST. Protein signals were detected using enhanced chemiluminescence and subsequently analyzed using the E-BLOT image analysis system.
Fresh femoral tissues from different groups of mice (n = 3 per group) were carefully dissected and immediately snap-frozen in liquid nitrogen. The tissues were then pulverized using a mortar and pestle under liquid nitrogen to preserve protein integrity. The powdered tissue was collected into 1.5 mL Eppendorf tubes, followed by the addition of lysis buffer. The mixture was incubated on ice for 30 min to ensure thorough lysis. Subsequent protein concentration measurement using the BCA assay and Western blot procedures was carried out as described previously for cell protein analysis.
ROS, SOD, NO, and H2O2 detection in cells/media
Cells were seeded into 6-well plates to detect ROS levels. The cells were washed twice with PBS and incubated in serum-free DMEM containing 10 µM DHE (Sigma-Aldrich) at 37°C for 30 min. The intracellular ROS staining was observed using a FACS Calibur flow cytometer (BD Biosciences), and the stained cells were examined using a laser scanning confocal microscope, LSM880 (Zeiss, Oberkochen, Germany), with an excitation wavelength of 647 nm. The cells were uniformly seeded into 6-well plates and cultured for 2 days. The cell culture medium and cells were then separated, and the intracellular levels of SOD, NO, and H2O2 were measured using the respective assay kits: NO (Beyotime, S0021S), SOD (Beyotime, S0109), and H2O2 (Beyotime, S0038). The levels of NO and H2O2 in the culture medium were also determined using the same kits. Assay Principles: NO Assay: This kit is based on the Griess reaction, which measures the stable metabolites of NO, primarily nitrites, by reacting with reagents to form a pink azo dye. The absorbance of the dye at 540 nm correlates with the NO levels in the sample. SOD Assay: This kit employs the classical nitroblue tetrazolium (NBT) assay. Superoxide anions (O2 −·) are generated by the xanthine and xanthine oxidase system, which reduces NBT to a blue formazan product. SOD activity inhibits formazan formation by scavenging superoxide anions, and the degree of color change is inversely proportional to SOD activity. H2O2 assay: This kit utilizes HRP to catalyze the reaction of H2O2 with a specific substrate, producing a colored or fluorescent product. The intensity of the color or fluorescence is directly proportional to the H2O2 concentration in the sample.
Statistical analysis
We performed a normality test on the data using SPSS 25.0 software, and the Shapiro–Wilk test was used to assess normal distribution. For data that conformed to a normal distribution, GraphPad Prism 9.5 software was used for statistical analysis. The resulting data were analyzed using one-way or two-way ANOVA, followed by Tukey’s post hoc test. Results were expressed as the mean ± standard deviation based on at least three independent experiments. For statistical comparisons between two groups, we used Student’s t test, and for comparisons among three or more groups, one-way ANOVA or two-way ANOVA was performed. A p value of <0.05 was considered statistically significant (*p < 0.05, **p < 0.01, ***p < 0.001).
Footnotes
Authors’ Contributions
W.X. designed and directed the study. J.O. wrote the article. J.O. and H.C. performed most of the experiments. J.O. and C.S. conducted data analysis. S.W. and C.S. intellectually contributed throughout the project. W.X. and D.G. guided the project and obtained funding. All authors contributed to the article and approved the submitted version.
Data Availability
Data are available upon reasonable request. All data and materials generated in this study are available upon request from the corresponding author.
Ethics Approval and Consent to Participate
All animal experiments and tests were guided and approved by the Ethics Committee of Soochow University (SUDA20210320A05).
Author Disclosure Statement
The authors declare no competing interests.
Funding Information
This research was supported by the innovation team project of the second affiliated hospital of Soochow University (XKTJ-TD202405) and the second affiliated hospital of Soochow University preresearch project of the National Natural Science Foundation of China (SDFEYGJ2004).
Supplementary Material
Supplementary Figure S1
Supplementary Figure S2
Supplementary Figure S3
Supplementary Figure S4
Abbreviations Used
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.