
Abstract
Selective serotonin reuptake inhibitor (SSRI) drugs, targeting serotonin transport, are widely used. A puzzling and biomedically important phenomenon concerns the persistent sexual dysfunction following SSRI use seen in some patients. What could be the mechanism of a persistent physiological state brought on by a transient exposure to serotonin transport blockers? In this study, we briefly review the clinical facts concerning this side effect of serotonin reuptake inhibitors and suggest a possible mechanism. Bioelectric circuits (among neural or non-neural cells) could persistently maintain alterations of bioelectric cell properties (resting potential), resulting in long-term changes in electrophysiology and signaling. We present new data revealing this phenomenon in planarian flatworms, in which brief SSRI exposures induce long-lasting changes in resting potential profile. We also briefly review recent data linking neurotransmitter signaling to developmental bioelectrics. Further study of tissue bioelectric memory could enable the design of ionoceutical interventions to counteract side effects of SSRIs and similar drugs.
Introduction: Long-Lasting Effects After Exposure to Serotonin Transporter Inhibitors
Selective Serotonin Reuptake Inhibitor (SSRI) drugs are widely used to treat depression and anxiety. Introduced around 1990, between 10% and 15% of the population of most Western countries now take them. 1 Close to 100% of takers of a SSRI have a degree of genital sensory change within 30 min of taking. These effects consist primarily of a reduced sensitivity, often termed “numbing” by those affected but others have genital arousal (irritability). The reduced sensitivity is accompanied by an immediate delay of ejaculation in men and muting of orgasm in both men and women. After a period of treatment, orgasm may stop and there may be a loss of libido. 2
The “numbing” effect produced by SSRIs has similarities to the effect of rubbing lidocaine into the genital area, which was a prior treatment for premature ejaculation, and SSRIs in single doses are used for premature ejaculation now. The effect is also described in terms of a loss of pleasurable sensation. In some cases, there is an actual genital numbing equivalent to that produced by lidocaine. These immediate onset sexual effects ordinarily lift when treatment stops. In 2006, reports appeared of a condition now termed Post-SSRI Sexual Dysfunction (PSSD), in which the genital numbing, pleasureless or absent ejaculation/orgasm, and loss of libido remain and may become more pronounced after treatment stops.3,4 PSSD can persist for decades afterward.2,5
In 2001, persistent genital arousal disorder (PGAD), an enduring disorder of irritable genital sensation, was described. 6 This condition is not linked to enhanced libido and does not stem from psychological issues. At present PGAD appears to affect women more than men. This condition seems more likely to happen around the menopause and, while closely related to discontinuation from SSRI medication, can also occur following trauma to the genital area. 2 These genital effects do not occur on antidepressants that do not inhibit serotonin reuptake; other antidepressants and psychotropic drugs can cause erectile dysfunction but not the syndromes of numbness, pleasureless orgasm, loss of libido, or persistent arousal.
Two other syndromes have been described which appear closely related to PSSD. One is postfinasteride syndrome (PFS). First described in 2011, this occurs in young men taking finasteride to stall hair loss. 7 It also happens with other 5-alpha reductase inhibitors—dutasteride and saw palmetto. Genital anesthesia, loss of libido, and sexual dysfunction are features of this syndrome. Initial finasteride treatment can produce some sexual dysfunction, but this is less common than with SSRIs. It is unclear if the sexual dysfunction that appears on treatment is continuous with PFS or distinct from it.
A postretinoid sexual dysfunction (PRSD) has also been described. 8 This also includes genital anesthesia, sexual dysfunction, and loss of libido. There can be some sexual dysfunction on initial treatment in patients taking isotretinoin for acne, but it is not clear what continuity there may be between this and PRSD. These enduring post-treatment syndromes may interface with tardive dyskinesia linked to antipsychotic drugs in the 1960s. Antipsychotics can cause dyskinesias on treatment, which ordinarily resolve when treatment is stopped. Dyskinesias can also emerge on withdrawal but clear up in time. Tardive dyskinesia is a syndrome that involves dyskinetic movements centered on the jaw and lower facial area, which can emerge on treatment but become more marked when treatment stops. The syndrome can endure for years or decades afterward.
These legacy effects of antidepressants and antipsychotics have some interface with withdrawal syndromes linked to these drugs. Withdrawal to opioids and alcohol is viewed as limited to a few weeks, having features not found during administration of the drug and as ordinarily responding to reinstitution of treatment. Antidepressant and antipsychotic withdrawal, however, is linked to dysthymia, which may appear continuous with the original problem but can be demonstrated in healthy volunteers given these drugs, as well as to other sensory and autonomic disturbances. These states can last for months or longer, opening up a possible link between enduring sexual syndromes and other legacy effects of antidepressants and antipsychotics. 9 There are variations among antidepressants and antipsychotics in their likelihood of causing withdrawal problems and likelihood of causing tardive syndromes, but the basis for these differences is not understood.
PSSD happens in all ages, both sexes, and all ethnic groups. It can begin after a few doses of treatment or only become apparent after years of exposure. There are two issues to account for. One is the original sensory changes. These almost certainly extend beyond the genital area, but are more salient there perhaps because of the functional consequences. SSRIs also produce a more general dampening of reactivity—commonly termed emotional numbing. This may be linked to the pronounced sensory features that characterize the SSRI withdrawal syndrome, which can include spontaneous orgasms and can result in PGAD.
In this study, we hypothesize about a possible mechanism of this effect and provide data in a physiologically-amenable model system consistent with this possibility.
Possible Mechanisms: A Hypothesis
At present, there is no agreement as to how the sensory changes on SSRIs come about. Lidocaine, which also produces genital numbing, appears to do so through an action on late sodium currents, 10 and serotonin reuptake inhibitors also have effects on late sodium currents. 11 It is also the case that antidepressants with effects on sodium currents are used to treat neuropathic pain.
Aiming at finding a treatment, PSSD sufferers have tried a wide range of agents active on various dopamine and serotonin receptors along with phosphodiesterase inhibitors and other drugs, but these have no therapeutic effect for PSSD, PFS, or PRSD. PFS sufferers have focused on evidence for androgen insensitivity. It is also the case that SSRIs reduce testicular volume and sperm counts, but these effects appear to happen in the absence of PSSD. At present, no endocrine manipulations appear to make a difference in PFS, PSSD, or PGAD.
The treatment approaches adopted to date have been largely targeted at reversing the acute sexual effects rather than reversing the mechanism that leads to enduring effects. This is similar to research efforts on tardive dyskinesia which for four decades have focused on the dopamine system without finding an answer. A second issue therefore is one of pinpointing a mechanism that might underpin enduring effects like these. It does appear that with time (several years) a degree of spontaneous recovery happens in some cases. In other cases, there are brief remissions (days), often triggered by stopping a brief course of another drug such as an antibiotic. There are grounds to think therefore that these enduring effects do not stem from permanent damage.
Is this problem best seen as a physiological (bioelectric) or a pharmacological matter? Is the site at which the original sensory changes are affected central or peripheral? Do they arise in a central nucleus, at the dorsal root ganglion level, or do from local treatment effects on C-fibers? In this study, we explore one possible mechanism: ion channel- and pump-driven circuits that maintain tissue bioelectric state as a kind of persistent physiological memory.
Developmental bioelectricity is a field which studies how cells, neural and non-neural, propagate, store, and process information through the propagation of electrical states—specifically resting potential or Vmem.12–14 Recent work in this field has revealed the importance of bioelectric networks not only in embryogenesis but also in adult regeneration, stem cell biology, cancer, and immune system function.13–27 Importantly, recent computational models predicted that some bioelectric circuits exhibit a kind of memory, in which induced changes of Vmem are actively maintained.28–31
Research in model systems, such as regenerative planarian flatworms (an important model for human neurophysiology and pharmacology32–37 ), has revealed a remarkable long-term memory that can be induced by alterations to endogenous ion flows. Planaria whose bioelectric circuits are briefly modulated by small molecule drugs experience an alteration in bioelectric patterns within their tissues that are apparently permanent and persistently affect their cell- and tissue-level functions long after the original drugs are withdrawn.38–40 This occurs over many months and is (as far as is known) permanent, despite the rapid turnover of all of their somatic cells within a few weeks.
Importantly, however, the ion channel and gap junction networks that implement bioelectric control circuits are ubiquitous, being present and function across taxa, from bacterial biofilms to mammals.41–52 Effects on cell migration, 53 proliferation, 54 migration/pathfinding,55,56 and stem cell differentiation 57 have been observed after modulation of neurotransmitter pathways. Even the microbiome, with many known roles in regulating mood and functioning of many organ systems, has been affected by neurotransmitter signaling.58,59 Taken together, these data suggest the possibility that persistent behavioral and physiological states, affecting the brain and numerous other organs, could be explained by long-lasting modulation of the bioelectricity-neurotransmitter axis through transient modulation by drugs. Thus, we hypothesized that SSRI application may alter Vmem in vivo, resulting in persistent changes to bioelectric circuit parameters in cells. This could be due to effects on the electrogenic serotonin transporter SERT or perhaps upon one or more of several ion channels.60–64 Because the basic bioelectric circuitry is highly conserved, animal models can serve as an important context within which to understand clinically relevant persistent physiological states induced by transient drug exposure. Thus, we examined the possibility of SSRI-induced long-term changes, which could provide a mechanism for the post-SSRI syndrome in human patients, in a tractable model system: planaria.
Experimental Test: Short-Term SSRI Soak Alters Long-Term Bioelectric Properties of Planarian Tissue
We experimentally examined the possibility of long-term effects of transient SSRI treatment on bioelectric state in intact planarian flatworms. We utilized the SSRI fluoxetine, because it has previously been shown in the literature to impact planarian regeneration. 65 The serotonin neurotransmitter system in planaria has been characterized in numerous studies: an ortholog of the gene encoding the enzyme tryptophan hydroxylase which catalyzes the rate-limiting step in serotonin synthesis has been identified in Dugesia japonica, 66 planarian serotonin receptors have been characterized,67,68 and multiples studies have found that SSRI treatment impacts planaria in numerous ways, including locomotion, light/dark preference (photophobic tendencies), DNA damage, and regenerative polarity.65,69–71
To assess the impact of transient fluoxetine treatment on bioelectric states, intact wild-type D. japonica were soaked in a 2 μM fluoxetine solution for 3 days, at which point the solution was washed out and the samples were placed in Poland Spring water for 1 week at 13°C to prevent fissioning (termed “1wk washout” group). Results are shown in Figure 1. Membrane polarization was then imaged using the voltage-sensitive dye bis-[1,3-dibarbituric acid]-trimethine oxonol [DiBAC4(3)].39,40,72 The 1wk washout group was compared directly to samples, which had been soaking in fluoxetine for 3 days (“fluoxetine soak” group) before imaging, and an age-matched H2O (“control”) group, which was never treated. Both the 1wk washout treated group and the 3d fluoxetine soak group were significantly depolarized relative to the controls (****p < 0.00001, Student's t-test, n = 14), while there were no differences in relative polarization between the 1wk washout group and the fluoxetine soak group (p = 0.16, Student's t-test, n = 14). This indicates that the depolarizing effect of fluoxetine is persistent far after the drug treatment has ended.
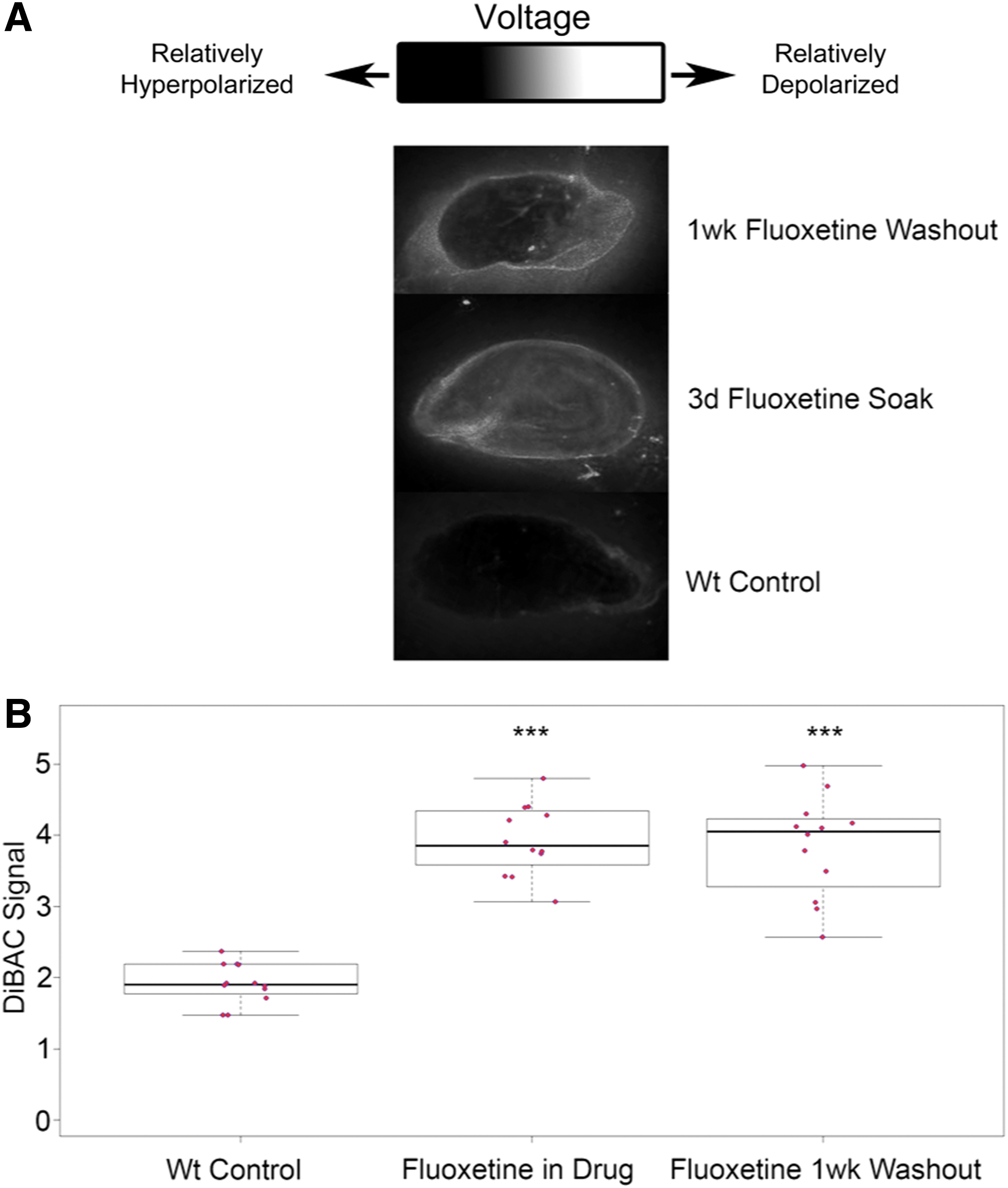
Fluoxetine exposure results in long-term physiological changes in planaria.
Conclusion
SSRIs' main target is the serotonin transporter, SERT. The well-known role of neurotransmitters in brain function has been suggested 73 to be an evolutionary extension of a more ancient and ubiquitous role in developmental (preneural) morphogenetic systems.74–76 A number of neurotransmitter pathways have been identified as functioning in vertebrate development, for example, in the embryogenesis of the face, 77 eye, 78 and heart,79–87 as well as invertebrate regeneration 65 and development.88,89 SERT, and serotonin signaling more broadly, has been identified as being part of bioelectric circuits in prior work. For example, voltage differences in early embryos drive the consistent left-right asymmetry of maternal serotonin molecules, which in turn control lateralized gene expression and visceral organ situs.90–93 Likewise, bioelectric controls of serotonin movement through gap junctions mediate the effect of ion channel drugs on ectopic innervation from transplanted organs94–96 and conversion of cells to a metastatic phenotype.95,97–99 However, prior work placed SERT downstream of voltage changes, and it was not known that SSRIs could also function upstream to alter Vmem of non-neural cells (relevant effects on neurons have been observed however100,101). In this study, we show this in planaria; moreover, the changes are persistent long after the SSRI is withdrawn.
Bioelectric circuits can maintain long-term and stable changes of state after relatively brief alterations of Vmem, and we have previously suggested bioelectric state to be a target of SSRIs and other psychoactive drugs in the context of developmental defects. 102 Such alterations can plausibly affect neural (and non-neural) responses such as could be important for human sexual function, either directly on somatic cells or through indirect effects acting through the microbiome, immune system, or brain.12,103–105 Bioelectric memory has not yet been demonstrated in human patient tissues, representing an important area for subsequent work, which could be addressed in vivo and in human organoid systems in vitro. 106 Paralleling the development of ion channel modulator drug cocktails, guided by computational models of bioelectric circuits to induce desired pro-regenerative states, it's possible that the negative effects of SSRI exposure could someday be mitigated by rationally designed cocktails of already human-approved drugs acting as ionoceuticals.107,108
Materials and Methods
Planaria husbandry
A clonal colony of D. japonica maintained in Poland Spring water at 13°C was used for the experiments in this study. All samples were starved for >7 days and continued to be starved throughout all experiments to control for metabolic variance in individual planaria. Colony care was performed as described in Oviedo et al. 109
DiBAC membrane voltage assay
DiBAC4(3) (Invitrogen, Carlsbad, CA) was utilized to visualize membrane potential in samples. Whole intact samples were treated with a 2 μM fluoxetine solution for 3 days, at which point they were rinsed thrice to remove residual solution and placed in water. One week after the fluoxetine treatment, these samples were imaged along with planaria that had been soaked for 3 days in fluoxetine, as well as H2O controls. Samples were soaked in DiBAC, which had been dissolved in the appropriate solution for the treatment, for half an hour before imaging. Planaria were immobilized using a 2% low-melting point agarose and mounted on microscopy slides using a cold plate. Groups of fluoxetine 1wk washout, fluoxetine in drug, and H2O control samples were mounted in a single slide, ventral side up. Analysis of relative membrane potential was done using the measure function in ImageJ software.
Footnotes
Acknowledgments
The authors thank Catharina Faber for helpful discussions and ideas.
Authors' Contributions
All authors wrote the text; J.L. performed the experiments. All authors have reviewed and approved of the article before submission. The article has been submitted solely to this journal and is not published, in press, or submitted elsewhere.
Author Disclosure Statement
No competing financial interests exist.
Funding Information
M.L. and J.L. gratefully acknowledge support by an Allen Discovery Center award from the Paul G. Allen Frontiers Group (No. 12171).