
Abstract
The observation of neuron-like behavior in bacteria, such as the occurrence of electric spiking and extended bioelectric signaling, points to the role of membrane dynamics in prokaryotes. Electrophysiology of bacteria, however, has been overlooked for long time, due to the difficulties in monitoring bacterial bioelectric phenomena with those probing techniques that are commonly used for eukaryotes. Optical technologies can allow a paradigm shift in the field of electrophysiology of bacteria, as they would permit to elicit and monitor signaling rapidly, remotely, and with high spatiotemporal precision. In this perspective, we discuss about the potentiality of light interrogation methods in microbiology, encouraging the development of all-optical electrophysiology of bacteria.
Introduction
The current golden standard for electrophysiology is represented by the patch clamp technique, which permits to measure membrane electrical phenomena precisely and directly. However, patch clamp and, in general, electrical probing and stimulation techniques require physical contact with the target cells and are usually time-consuming. In contrast, optical technologies offer the possibility to target specific cells or even cell compartments at the same time with high spatiotemporal precision, remotely and in a noninvasive manner. Therefore, driven by these motivations, many research efforts have been directed toward the development of noninvasive and high-throughput optical methods in electrophysiology.
Neuroscience has largely benefited from optical technologies so far, as the efficient and reversible modulation of neuronal activity allows studying the function of specific brain circuits at unprecedented time/space scales, thus opening new avenues for the treatment of psychiatric and neurological diseases. 1 Optogenetics is the pioneering technique for eliciting and probing neuronal response by means of light. 2 This approach exploits gene therapy to express exogenous light-sensitive transmembrane proteins, enabling both on–off control of neuronal activity with cell-type specificity and recording of intercellular signaling via genetically encoded voltage reporters. 3
Photopharmacology is an alternative method to confer light sensitivity to cells. 4 In this case, selected light actuators (i.e., azobenzene-tethered ligands) are directly attached to the specific ion channel, 5 at the lipid membrane level, 6 or even to drugs to photomodulate their pharmacodynamics and pharmacokinetics. 4 Both optogenetics and photopharmacology, however, require either genetic modification or covalent attachment of light actuators to the target, thus introducing a set of technical challenges. An alternative and relatively novel paradigm in optostimulation relies on the use of nanostructures and molecular actuators to achieve optical control over bioelectric signaling without the need to modify genetically or chemically the bio-target. Relevant examples include the use of inorganic and organic nanostructures and molecular systems to modulate neuronal activity and to rescue vision.7,8 In general, these methods possess lower specificity and temporal responses than optogenetics and photopharmacology, while ensuring comparable electrophysiological performances with lower invasiveness. 9
Recent findings have demonstrated that membrane potential dynamics and extended bioelectric signaling also occur in bacteria.10,11 This offers a chance for exploiting optostimulation methods to prokaryotes. The study of brain-like signal propagation in bacteria is of fundamental interest as a model system of neuronal network, 12 but it has also direct implication in bacterial behavior, metabolism, and adaptation to antibiotics, among others.13,14 In this regard, the relatively new field of bacterial electrophysiology can benefit extensively from the utilization of optical methods. Light holds the potential to overcome the existing limitations of the electrode-based techniques, mostly arising from the requirement to contact small and motile cells exhibiting thick cell walls. 15 Therefore, the development and engineering of new photoactive systems capable to interface effectively with bacteria and to trigger/probe signaling is highly desirable.
In this perspective, we aim at providing an outlook over the use of optical methods to control and probe biological signaling, with particular emphasis on the possible use of light technologies for the investigation of the bacterial electrophysiology. It is worth adding that while optogenetics and nongenetic optical methods are being applied to trigger and/or investigate signaling in bacteria, photopharmacology is traditionally utilized for the light modulation of antibacterial activity of existing antibiotics, that is, via the covalent attachment of azobenzene ligands to the target molecules (see, for instance, the works of the Feringa's group and collaborators16–18 ). Therefore, we will not include photopharmacology in our discussion, as this method is not directly employed for the photocontrol of bioelectric signaling in bacteria.
Electrostimulation and Probing Methods in Bacterial Electrophysiology
Electrostimulation and probing methods are currently being applied for studying the electrophysiology of bacteria. In this regard, Asally and collaborators have shown recently that membrane potential dynamics can be elicited via stimulation with exogenous electrical signal. 14 For this purpose, the authors designed a bespoke electrode-coated glass-bottom dish, which allowed electrostimulation while ensuring membrane potential probing with a fluorescence membrane-potential indicator. In particular, they showed that electrical stimulation causes hyperpolarization in unperturbed cells. Conversely, when cells were pre-exposed to ultraviolet (UV) light or antibiotics, the same electrical stimulation depolarizes cells instead of causing hyperpolarization. This suggested that external stimulation can be used as a tool for the discrimination between proliferative and nonproliferative bacteria.
Passing to electrical probing, patch clamp is the technique of choice in neuroscience. Although it permits to measure directly and quantitatively the electrical potential, it cannot be applied to living bacteria, due to their small sizes and cell wall. However, patch clamp can still provide some precious information on the role of ion channels in bacteria.19,20 For example, Martinac et al. used patch clamp on giant spheroplasts to study native mechanosensitive channels in Escherichia coli. 21
The most important advantage of electrical techniques resides on the fact that they have been utilized largely in neuroscience for many years. Electrical stimuli can permit to trigger the potential dynamics with spatial precision, both in planktonic cells and in biofilms by using multielectrode arrays. 22 However, optostimulation can surpass electro-based methods in some regards. Specifically, light can be used to target both specific cells and subsets of a bacterial community at the same time and with different colors, also via the use of patterned excitation. For instance, this can be a key advantage to study cell-to-cell signaling heterogeneity in biofilms, which accounts for antibiotic stress response 23 and the xenobiotic metabolism in the gut microbiota. 24 In addition, light stimuli can enable fast and localized excitation at the close proximity or even within the bacterial cell, permitting to potentially disclose and investigate fast (milliseconds) signaling in bacteria. Finally, biomaterials science can permit to optimize the photoactuation process for a given bacterial species or, alternative, can allow engineering of actuators that are well suited for a wide range of bacteria, in the view to utilize optostimulation also in complex communities.
Passing to the membrane potential probing, our idea is that optical methods are intrinsically more advantageous than patch clamp for bacterial electrophysiology. This explains why fluorescent voltage indicators are already used extensively for this purpose. However, these systems do not allow a direct and quantitative measurement of the potential, as it happens in patch clamp. This can be a serious limitation for the development of the field.
Optogenetics
In optogenetics, light sensitivity is conferred to cells via the incorporation of a protein-based light actuator that is fully genetically encoded. In its classical form, optogenetics employs microbial opsins to take photocontrol over neuronal signaling. The first seminal study on the photocontrol of neuron spiking via the use of channelrhodopsin-2 was reported by Deisseroth and collaborators in 2005. 25 This report demonstrated for the first time that temporally precise and noninvasive control of neuronal activity can be achieved at faster timescales (milliseconds) than previous photostimulation methods, such as photopharmacology. 26 Since then, a plethora of genetically encoded light actuators and fluorescent voltage reporters have been developed.1,3,27 These include various natural photoreceptors from animal, plants, and microorganisms.
In regard to microbiology, optogenetics has been employed recently to achieve photocontrol over the swimming speed of bacteria,28,29 a parameter that ultimately depends on the protonmotive force and membrane potential. However, genetically encoded voltage indicators are largely utilized in bacterial electrophysiology. 13 In 2011, Cohen and collaborators reported for the first time on the occurrence of electrical spiking at 1 Hz in E. coli (Fig. 1), which was revealed by using a genetically encoded dye, namely proteorhodopsin optical proton sensor (PROPS). 30 In particular, the indicator showed a fluorescence blinking behavior that was directly related to membrane electrical depolarization. This seminal study opened the field of bacterial electrophysiology, a research area that was usually poorly investigated due to small size of bacteria and a lack of sensitive tools. For instance, Bruni et al. employed a combined calcium–voltage sensor based on PROPS to show that electrical spiking is connected to mechanosensation. 31 More recently, the same indicator has been used to prove that the bactericidal action of aminoglycosides, such as kanamycin, arises from the dysregulated membrane potential. In the absence of transmembrane voltage, aminoglycosides are taken into cells and exert bacteriostatic effects by inhibiting translation. 32
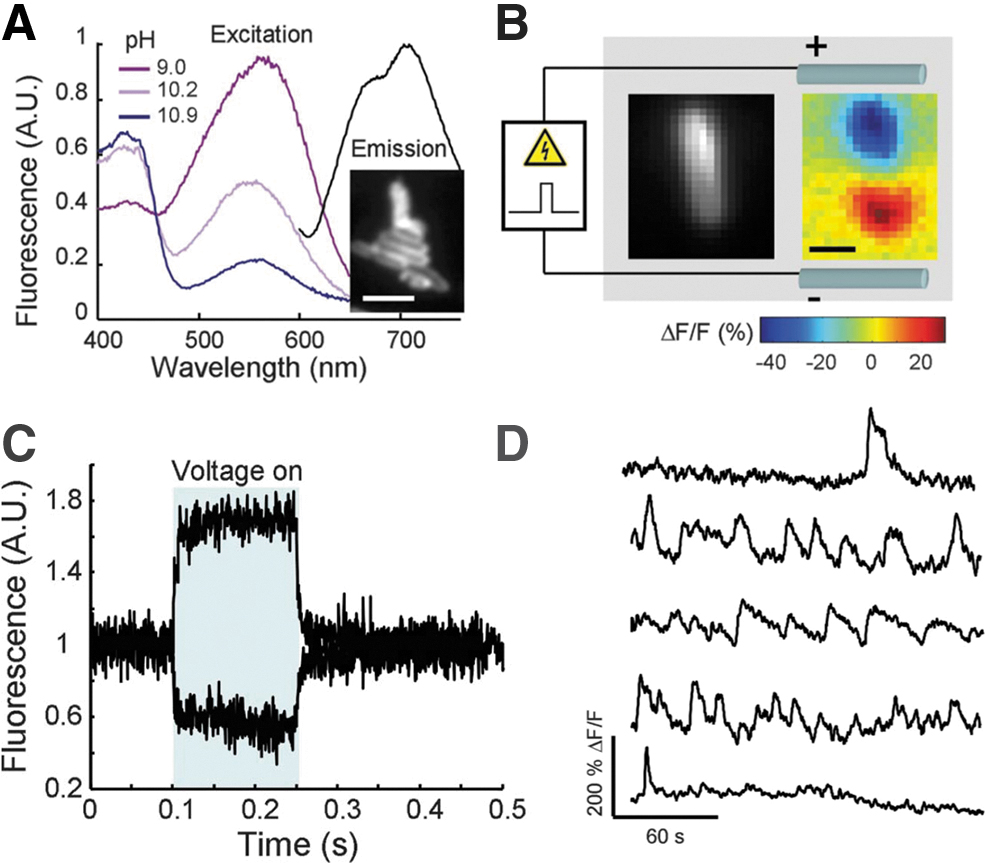
PROPS dye is used as fluorescent indicator to monitor membrane potential dynamics.
To summarize, although optogenetics would allow stimulation and monitoring of membrane potential dynamics at unprecedented length scales, this technique would unavoidably introduce a set of challenges, that is, the poor applicability of the genetic transformation to certain bacterial species and strains, 13 as well as the large use of UV light that already interfere with membrane potential. 33 This might possibly represents a serious limiting factor for the widespread use of optogenetics for bacterial electrophysiology.
Nongenetic Optostimulation
In this strategy, the photoactuator and/or the probe are neither genetically encoded nor chemically linked to the bio-target, as it happens for optogenetics and photopharmacology, respectively. Thus, the abiotic/biotic interface is constituted on the basis of noncovalent affinity or physical contact with the cells. In the last couple of decades, this field has seen a steep rise mostly due to the development of new nanostructured materials and molecular actuators. These systems permit to localize the excitation with a relatively high spatial confinement owing to their reduced dimensionalities, as well as offering the possibility to tune their action spectrum and temporal resolution via top-down processes and chemical synthesis. 8
Photomodulation usually occurs via three main mechanisms, namely: photothermal, photocapacitive, and photoelectrochemical phenomena due to charge generation and photomechanical. Both the photoexcitation properties of the materials and the interface structure (i.e., diffuse or planar interface) dictate which biostimulation mechanism is at work. Detailed information about the most employed materials and interfaces used for nongenetic optostimulation, as well as the relative stimulation mechanisms, is contained in the following recent reviews,7,8,34 and is briefly summarized in Figure 2. Conversely, nongenetic photomechanical stimulation has been introduced for eliciting signaling in neurons only recently. In particular, it has been proposed a new amphiphilic azobenzene that dwells in the plasma membrane in a noncovalent manner, giving rise to a light-induced transient hyperpolarization, followed by a delayed depolarization that triggers action potential both in vitro and in vivo.9,35–37 The stimulation mechanism resides in the trans→cis photoreaction of azobenzenes: molecular dynamics simulations show that in dark the trans isomer can undergo dimerization causing a thinning of the membrane and an increase of its electrical capacitance, while illumination generates the formation of a stable population of cis isomers and, thus, the disruption of the dimers leading to a restoration of membrane thickness and capacitance.

Cartoon illustrating the three main approaches and materials employed for nongenetic optostimulation.
Currently, nongenetic optostimulation methods are largely applied to neuroscience, for instance, for eliciting signaling response in neurons 8 and for vision restoration,41,42 and for developing new biolectronic plaftorms.43,44 However, this approach holds a great potential for delivering transient (from milliseconds to picoseconds), localized, and controllable perturbations to bacterial cells and communities, without the need to use complex wiring or specific stimulation devices. These advantages can make light stimulation a valid and powerful alternative to electrical interrogation methods,14,45,46 that is, permitting to study fast microbial signal transduction in response to transient perturbations. The first evidence of the potentiality of light stimulation methods in microbiology have been demonstrated in a recent article. 47 In particular, Gao et al. have reported on the use of silicon micro/nanostructures to trigger signaling in planktonic bacteria and communities via photothermal stimulation. Interestingly, this effect permits to elicit a previously unidentified form of rapid, photothermal gradient-dependent, intercellular calcium signaling within the biofilm (Fig. 3).
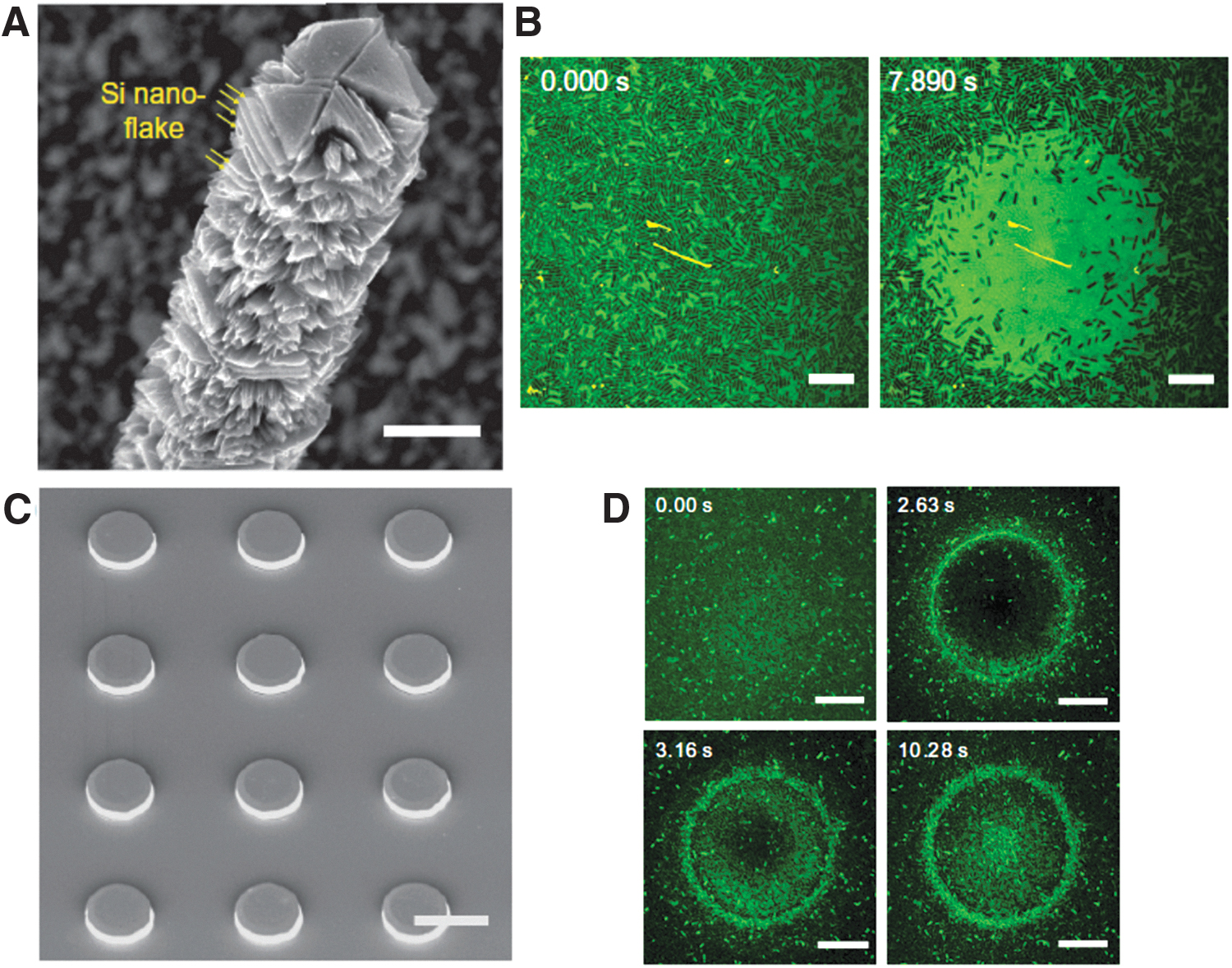
However, the simple extension of the optostimulation tools used in neuroscience to prokaryotes might be still insufficient for a rapid advancement of the field. This is largely due to the differences between eukaryotic and prokaryotic electrophysiology, stemming essentially from the relatively small sizes and presence of cell wall in bacteria. Details about the electrophysiological dynamics in prokaryotes are discussed comprehensively in two review articles.13,15 Briefly, the smaller surface area of bacteria than mammalian cells (6 μm2 vs. 1600 μm2) leads to a lower membrane capacitance in prokaryotes (
Altogether, these profound differences suggest some general criteria for the design of photoactuators ad hoc for bacterial nongenetic optostimulation. Given that membrane capacitance and resistance can be sensitive targets for stimulation, an ideal photoactuator should be able to directly focus on these parameters. In neurons, this is usually accomplished by means of thermal increase of the local temperature via the excitation of metal nanostructures and capacitive coupling with semiconducting materials (Fig. 2), with both methods acting indirectly on the ionic species and electric field in the vicinity of the membrane. In prokaryotes, the presence of cell wall and the small number of extracellular ions would in principle decrease the impact of these approaches. Alternatively, we reckon that molecular materials that directly partition in the plasma membrane would be more effective in these regards. These could act on the geometrical characteristics of the membrane and/or on its polarizability (i.e., donor–acceptor molecules), thus leading to a direct photomodulation of the membrane capacitance or to the opening of mechanosensitive or voltage-gated channels (Fig. 4).19,21 Last but not least, chemical synthesis usually enables fine tailoring over molecular characteristics. This can allow enhancing the affinity of the actuator for bacterial cells and the membrane environment, for instance, via the judicious use of bio-affine charged groups capable to interact with the negative charges of the cell wall, and appropriate lipophilic units that drive partitioning in the membrane interior.
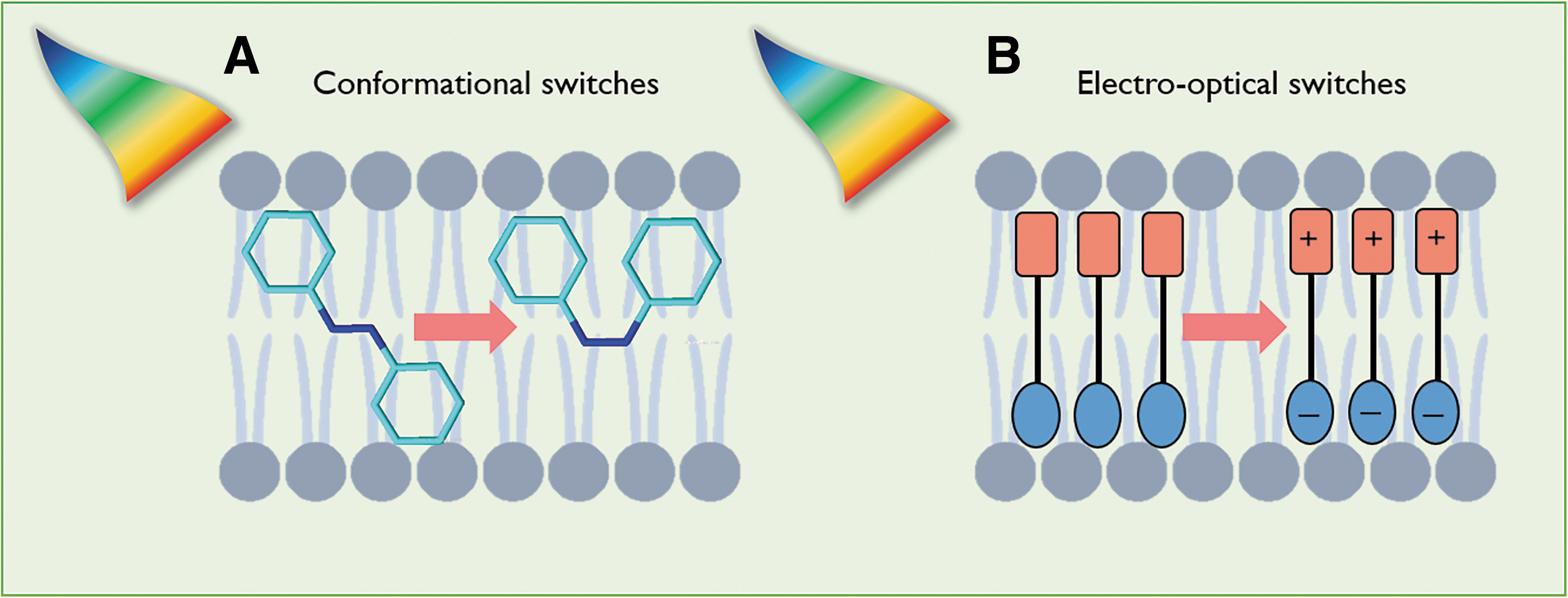
Two possible examples of intramembrane molecular switches that could permit to elicit efficiently the bacterial membrane potential dynamics. These systems would directly modulate the membrane capacitance via geometrical perturbation of the bilayer characteristics (i.e., thickness) by using conformational switches
Together with the introduction of efficient light actuators, the exploitation and development of nongenetic fluorescent indicators for the noninvasive monitoring of cell membrane potential can permit to advance toward all-optical bacterial electrophysiology. Currently, the most used nongenetic membrane potential indicators are Nernstian dyes and membrane-bound dye. The functioning mechanism of Nernstian dyes relies on the different distribution across the membrane of cationic molecules, whose partitioning is regulated by the Nernst potential. Therefore, it is possible determining the transmembrane potential by measuring the fluorescence ratio of the dye inside and outside the cell. The most used Nernstian dye in bacteria electrophysiology are tetramethylrhodamine, methyl ester (TMRM) and bis-(1,3-dibutylbarbituric acid) trimethine oxonol [DiBAC4(3)]. These systems, however, require careful calibration for a quantitative estimation of the membrane potential. 48 In addition, their relatively high permeability hampers the investigation of fast dynamics. 13
Conversely, membrane-bound dyes are well suited for monitoring fast membrane potential dynamics, even though they exhibit a weaker signal-to-noise ratio than Nernstian dyes. The most used membrane-bound dye in bacteria electrophysiology is aminonaphthylethenylpyridinium (ANEP). 49 Membrane-bound dyes shift their excitation spectra according to the membrane potential. The fast developments in the field of membrane potential indicators suggest that optical methods in electrophysiology will be more routinely used in the future. However, to compete effectively with patch clamp technique, these indicators should allow direct measurement of the membrane potential without the need of careful calibration procedures. To this end, a paradigm shift in the recording mechanism is highly desirable. A possible strategy for obtaining absolute information is to translate the spectral data into the time domain, such as by using fluorescence-lifetime imaging microscopy. For instance, the use of indicators that change their excited state lifetime upon potential variation has been demonstrated as a powerful approach to measure absolute voltage in eukaryotes,50–52 while this method has never been applied to prokaryotes.
Conclusions and Outlooks
In this perspective, we have summarized the most popular approaches employed to confer light sensitivity to cells, allowing both photomodulation and monitoring of cells signaling. To date, optical modulation and interrogation techniques have been largely applied in neuroscience. These approaches have allowed gaining insights into extended neuronal signaling and specific brain functions at unprecedented time/length scales.
Interestingly, recent findings have indicated that bacteria possess neuron-like behavior, such as electric spiking and extended bioelectric signaling, opening the way to electrophysiological studies in bacteria. In principle, bacterial electrophysiology can immensely benefit from optical methods, as light can surpass electrostimulation/probing in many regards, thus allowing a paradigm shift analogous to the introduction of optostimulation in neuroscience. To achieve this, the actuation process needs to be tailored specifically for prokaryotes, while optical probing should permit to access the membrane potential directly and quantitatively.
Although this field is at its infancy, we reckon that interdisciplinary research efforts encompassing microbiology, biomaterials science, optical spectroscopy, and imaging will permit to reach the critical mass necessary to develop the tools and knowledge for bacterial all-optical electrophysiology.
Footnotes
Authors' Contributions
All authors contributed to article drafting and revising, and figure creation. The final version of the article was reviewed and approved by all authors. This article has been submitted solely to this journal and is not published, in press, or submitted elsewhere.
Author Disclosure Statement
No competing financial interests exist.
Funding Information
This study was supported by Fondazione Cariplo (Grant No. 2018-0979 and Grant No. 2018-0505).