
Abstract
Primary fatty acid amides (PFAMs) are interesting lipid-derived signaling molecules with wide-ranging actions, including bioelectric. In this perspective, we evaluate a possible triangular relationship: PFAMs—ion channels—cancer. The primary emphasis is on oleamide and its metabolic associate, oleic acid (OA). As ion channels, we have focused on voltage-gated sodium channels (VGSCs) and voltage-gated and inward rectifier potassium channels in a variety of cell types, including cancer cells. As cancers, we evaluate the evidence for breast and colorectal cancers. Cis-oleamide blocked VGSCs and potassium channels. In parallel, but not always concurrently, oleamide has been shown to produce antiproliferative and anti-invasive effects, consistent with the known control of these cellular behaviors by potassium channels and VGSCs, respectively. Importantly, there is also evidence that oleamide can potentiate chemotherapy. In contrast to oleamide, the effects of OA on the ion channels of interest and, correspondingly, cancer cell behaviors are much less consistent. The perspective is concluded with three short sections. First, we outline the actions of some other PFAMs, especially anandamide and linoleamide. Second, we give an overview of some other targets that have been associated with oleamide, especially cannabinoid receptors and gap junctions. Finally, we cover the anticancer effects of oleamide on some other neoplasms.
Introduction
Primary fatty acid amides (PFAMs) are lipid-derived signaling molecules, which include oleamide, anandamide, palmitamide, stearamide, erucamide, and linoleamide. 1 In general, they are generated from hydrolysis of fatty acids in the presence of ammonia. 2 Most work has been done on oleamide, especially as regards (1) ion channels as targets and (2) cancer as an associated disease state. Oleamide, cis-9-octadecenamide [CH3(CH2)7CH = CH2(CH2)7CONH2], comprises an 18-carbon acyl chain with a single cis-paired link at C9. Following the discovery by isolation from the cerebrospinal fluid of sleep-deprived cats (next paragraph), experiments showed that oleamide is present also in rat and human plasma, indicating that its biosynthesis could occur in the body.
Indeed, using rat brain microsomes, Sugiura et al. synthesized oleamide from oleic acid (OA) and ammonia as a substrate in vitro by the reversible catalytic action of fatty acid amide hydrolase (as illustrated in Fig. 1). 3 OA is an omega-9 fatty acid and can be made by the body. It is also found in foods (highest levels in olive oil and other edible oils). Oleamide may also be derived from N-oleoylglycine by the action of the neuropeptide processing enzyme, peptidylglycine α-amidating monooxygenase. In addition, there are other routes for the production of oleamide. 1 All these are probably dependent on cell type and its biochemical status.
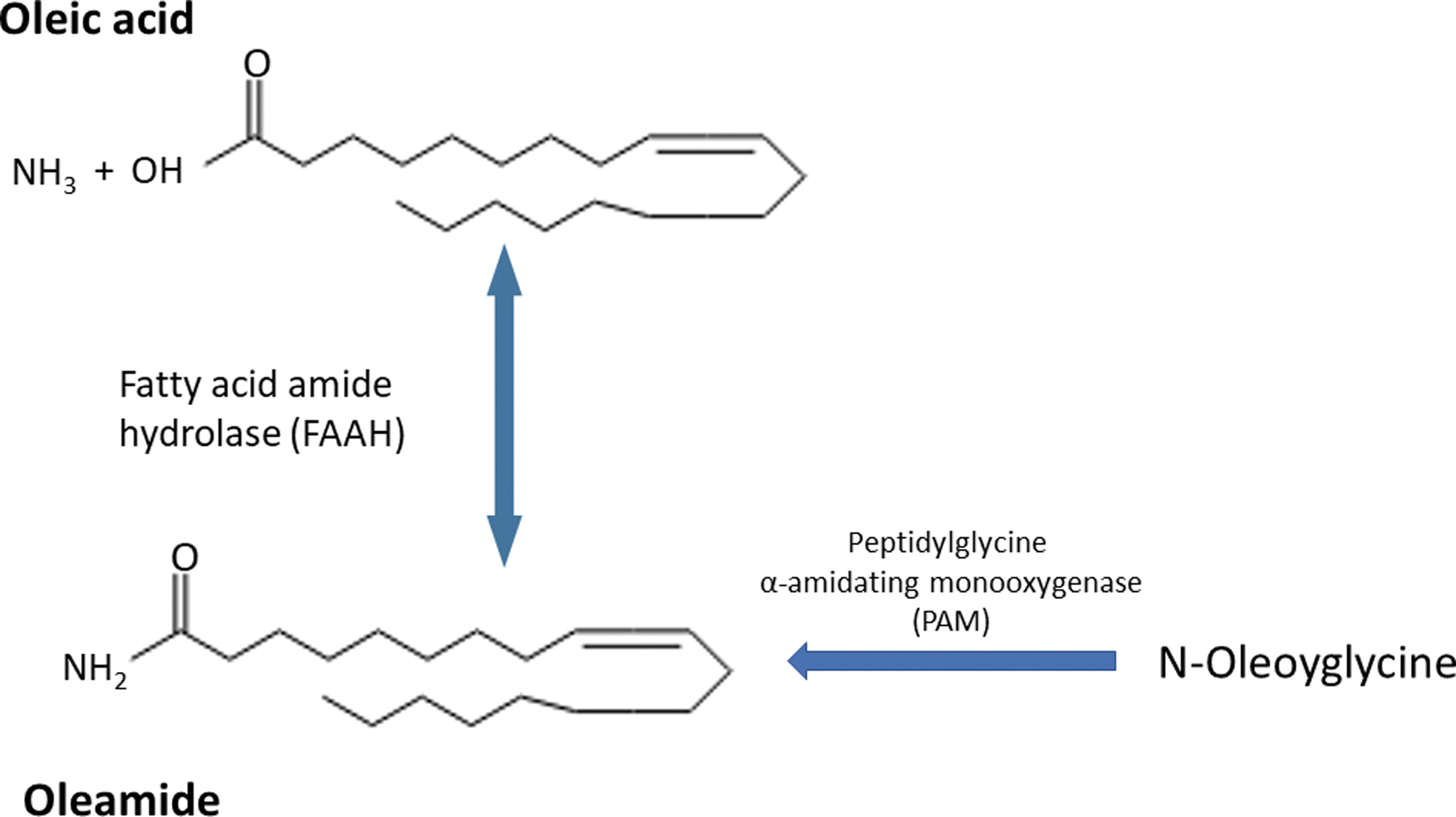
Schematic illustration of synthesis of oleamide from oleic acid and NH3 substrates by the reversible catalytic action of FAAH. FAAH, fatty acid amide hydrolase; PAM, peptidylglycine α-amidating monooxygenase.
Oleamide was first isolated from the cerebrospinal fluid of sleep-deprived cats.4,5 Subsequent work showed that it is involved also in a wide range of physiologies, including locomotion, angiogenesis, inflammation, thermoregulation, and vasoregulation. 6 A range of targets and signaling mechanisms has been associated with the action of oleamide, including receptors (e.g., cannabinoid-like receptor-1) and enzymes (e.g., cyclooxygenase, metalloproteinase). 7
There is significant evidence for involvement of oleamide and OA in the cancer process. For example, it was reported that oleamide analogs (including linoleamide) can cause apoptotic, antiproliferative, antiangiogenic, and antimetastatic effects on human bladder, urinary, and breast cancer (BCa) cells in vitro and in vivo.8–10 Low doses (50 and 100 μM) of oleamide were found to exert antiproliferative effects and caused apoptosis in rat glioblastoma RG2 cells. 11
The effects of OA on cancer appeared somewhat more variable. On the whole, OA produced antitumor (antiproliferative and proapoptotic) effects.12,13 Treatment of human lung cancer A549 cells with different doses of OA caused cytotoxicity by the release of pulmonary surfactant protein B into the tumor microenvironment. 14 On the other hand, OA-induced NADPH oxidase family 4, a mitochondrial energy biosensor, and promoted the metastatic ability of human colorectal cancer (CRCa) cells by increasing the level of reactive oxygen species (ROS). 15
The apparent physiological (especially neurophysiological) and pathophysiological roles of oleamide and OA raise the possibility that their molecular targets may involve ion channels. The latter are known generally to contribute to a wide range of cellular activities, from the most basic (e.g., gene expression) to whole cell (e.g., secretion, motility). Furthermore, ion channel dysfunction can give rise to range of pathologies including cancer. 16
The main aim of this perspective is to evaluate the effects of oleamide (and to a lesser extent other PFAMs) on key Na+ and K+ channels and associated signaling mechanisms. Indeed, Na+ and K+ channels frequently work in concert. Furthermore, the concerted activities of these two channels have been suggested to drive the “celex” mechanism of metastasis. 17 In essence, we endeavor to inquire whether there might be a basis for a possible triangular relationship, involving PFMAs (oleamide), ion (Na+ and K+) channels, and cancer, as illustrated in Figure 2.

A conceptual representation of the potential triangular relationship involving PFAMs, ion channels, and cancer. In this study, we cultivate this relationship for oleamide, Na+ and K+ channels, and breast and colorectal cancers. PFAMs, primary fatty acid amides.
Sodium Channels
There are two main types of sodium channels: voltage-gated sodium channels (VGSCs) and epithelial sodium channels. In addition, some “transient receptor potential” channels also permeate Na+. As regards oleamide, most work has been done on VGSCs.
VGSCs are composed of one α-subunit (VGSCα) and 1–2 β-subunit(s) (VGSCβ). VGSCαs (220–260 kD) form the Na+ pore region and also include the various posttranslational modification sites. 18 The β-subunits (30–40 kDa) are auxiliary and contribute to the channel's intracellular trafficking and stability in plasma membrane. VGSCαs comprise a multigene family with nine functional subtypes (Nav1.1–1.9) with varying tissue expression patterns. 19 Functional VGSCs are expressed in classically “excitable” cells, such as nerves and muscles. However, increasing evidence suggests that VGSC expression also occurs in so-called “nonexcitable” cells, for example, fibroblasts, glia, and cells of the immune system.19,20 Importantly, cancer, including carcinoma, cells also express functional VGSCs, which play a significant role in driving, possibly even initiating, metastasis.21–23 The basic functional mode of VGSCs is generation of all-or-none action potentials (APs) although nonconducting roles also occur. 24 VGSC activity can be blocked selectively by the natural toxin, tetrodotoxin (TTX). Indeed, TTX-sensitive APs have been recorded in human strongly metastatic BCa cells in vitro and in vivo.25,26
Behavioral effects of oleamide were initially studied in mice. 27 Thus, intraperitoneally applied oleamide was found to significantly suppress a range of motor functions, including rearing, grooming, and locomotion. These effects were dose dependent with some gender dependence. Interestingly, oleamide also demonstrated an antidepressant-like property. These results were confirmed, and it was also shown that oleamide potentiated the inhibitory effect of diazepam and antagonized the stimulatory effects of ethanol, methamphetamine, and caffeine. 28 From such diverse neurophysiological/pharmacological effects, it was suggested that oleamide could interact with a multiplicity of neurotransmitter receptors and ion channels in the central nervous system (CNS). 27 The molecular basis of such effects of oleamide was studied on cultured pyramidal neurons. 29 Cis-(but not trans-) oleamide reversibly suppressed the sustained repetitive firing but had no effect on the amplitude or time course of APs (Fig. 3a).

Effects of oleamide and oleic acid on VGSCs in different cell types.
This effect was dose dependent with an average EC50 value of 4.12 μM (Fig. 3b). The mode of action was consistent with a frequency- and/or state-dependent block of VGSCs. Such a modulatory effect seemed similar to the actions of anticonvulsant barbiturates and a variety of anesthetics, which also serve as blockers of VGSCs. 29 Further experiments were undertaken with radiolabeled batrachotoxin A 20-alpha-benzoate (BTX-B), a VGSC ligand. 30 Thus, Nicholson et al. confirmed that oleamide blocked the binding of BTX-B to VGSCs in mouse CNS synaptosomes and cultured NIE115 murine neuroblastoma cells. 31 More recently, leaf extracts of several species of Ipomoea and Dillenia, containing high levels of oleamide, similarly inhibited several subtypes of recombinant VGSCs. 32 In human, strongly metastatic BCa MDA-MB-231 cells, VGSC (neonatal Nav1.5) activity was also inhibited strongly by oleamide (Fig. 3c).
In contrast, from the available evidence, OA appeared to produce inconsistent effects. On VGSCs expressed in cultured human bronchial smooth muscle cells, OA was inhibitory. 33 On the other hand, intracellularly applied OA potentiated VGSC (presumed Nav1.4) currents recorded from adult human skeletal muscle biopsies. 34
In conclusion, oleamide (synthetic or natural) is consistent in inhibiting VGSC activity in a variety of cellular preparations, but further work is required to understand the action of OA.
Potassium Channels
There are three main types of K+ channel: (i) voltage-gated K+ (Kv) channels; (ii) inward-rectifier K+ (Kir) channels; and (iii) two-pore K+ channels. 35 These subserve a range of cellular functions dependent on the physiology of the tissue of origin. Studies on Caenorhabditis elegans suggested the expression of over 100 potassium channel genes. More than 80 genes of potassium channels were found in human genome. 36 As regards oleamide, most work has been done on Kv and Kir channels.
Kv channels
In human genome, there are some 40 different genes encoding for Kv channel subunits and these are subdivided into 12 subfamilies (Kv1–Kv12). 37 The molecular structure of given Kv channel subunits comprises six transmembrane (TM) helices (S1–S6) and one pore-forming P-region. Functional Kvs occur as tetramers mainly of given subfamily members. The different channel subtypes vary in their voltage sensitivity and kinetic profiles. 38 Their cellular functions are varied, including generation of the membrane potential (Vm) and patterned signal transmission. Kvs contribute to signal generation in “excitable” cells but can also occur extensively in epithelial cells. 39
Effects of oleamide (as the main hexane extract from leaves of Ipomoea and Dillenia species) were studied on three different Kv channels expressed in Xenopus oocytes (Fig. 4a). Thus, activities of Kv1.1 and Kv10.1 channel were shown to be inhibited while the Kv11.1 (hERG) channel was not affected. These effects were linked to an anti-inflammatory property of oleamide. 32
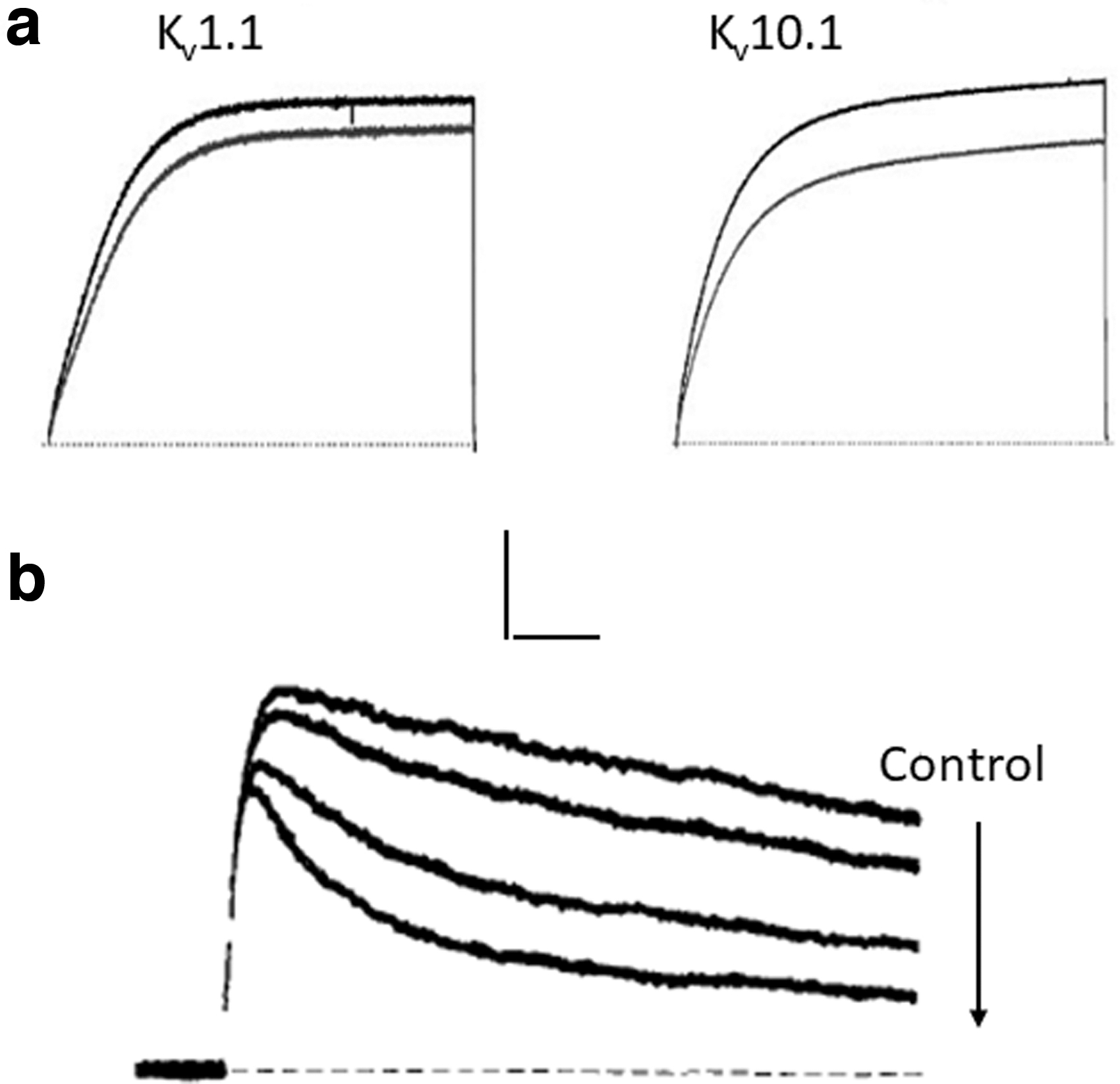
Effects of oleamide and oleic acid on depolarization-induced potassium currents.
OA also inhibited Kv (“delayed rectifier”) currents measured in NG 108-15 neuroblastoma cells (Fig. 4b). Two effects of OA were apparent: (i) time-dependent suppression of peak current and (ii) acceleration of the current inactivation. 40
Some work has also been done on calcium-activated K+ (KCa) channels. These can be considered as a member of the Kv family due to their molecular similarity, although KCa channels are opened primarily by a rise in intracellular Ca2+ and are only weakly voltage sensitive. Effects of some unsaturated fatty acids, including OA, on KCa channels were studied on gastric myocytes of guinea pigs. OA significantly increased the KCa current dose dependently by up to 13%. 41 This could be due to OA mobilizing intracellular Ca2+. 42
Kir channels
These channels also occur as tetramers, each subunit being formed from two TM helices and one P-region. 43 As well as contributing to membrane electrogenesis in some cells, their classic role is in tissue K+ homeostasis as in neuro–glia interactions in the CNS 44 and vasodilatory effects in brain. 45
Liu and Wu showed originally that oleamide inhibited an E-4031-sensitive presumed Kir current in pituitary GH3 cells and in neuroblastoma IMR-32 cells in a dose-dependent manner. 46
A variant of Kir is the ATP-gated K+ (KATP) channel. This channel opens (and Vm hyperpolarizes) when the cellular level of ATP falls. Kir (in particular Kir 6.1/2) is an integral part of the KATP channel complex. OA was shown to inhibit the KATP channels in pro-opiomelanocortin (POMC) neurons and cause excitation in hypothalamus. 47 Such an effect could be associated with metabolic aspects of food intake and insulin signaling. 48 Interestingly, in another study, OA was found to suppress the expression of Kir6.1 protein in human umbilical artery smooth muscle cells. 49 This raises the possibility of long-term effects of oleamide/OA with possible feedback interactions inherent in the metabolism (Fig. 1).
Effects on Cancer Cells
In this section, we review some highlights of the effects of oleamide and OA on cancer cells and attempt to link these to possible involvement of ion channels, especially sodium and potassium channels, covered in Sodium Channels and Potassium Channels sections, respectively. While most experiments have been carried out using these agents exogenously, we should note that cancer cells themselves can also synthesize oleamide. 50 Furthermore, oleamide may be released from cells in “extracellular vesicles” and thus mediate cell–cell interactions within the tumor microenvironment. 51 Overall, our intention is not to give an exhaustive account of all cancers. Instead, we highlight the best understood cases, as follows.
Breast cancer
There is significant experimental evidence showing that oleamide can suppress various aspects of BCa cell behavior, including proliferation and invasion. Indeed, using the MDA-MB-231 cell model of triple-negative human BCa, Zibara et al. showed that oleamide inhibited proliferation (cell cycle arrest) as well as invasiveness in vitro. 52 Similarly, oleamide inhibited the proliferation of human BCa EFM-19 cells. 50 Furthermore, Wisitpongpun et al. showed that oleamide-rich leaf extracts of Moringa oleifera showed anticancer activities by inducing cell cycle arrest and triggering apoptosis in MDA-MB-231 cells. 53 The underlying mechanism was suggested to be suppression of Bcl-2 expression and subsequent activation of caspase 3. 53 Importantly, oleamide also reduced metastasis in a murine xenograft model in vivo. 52 These effects were generally dose and time dependent.
In conclusion, oleamide could suppress both primary and secondary tumorigenesis. These effects of oleamide may be explained by its blocking actions on VGSCs and potassium channels, which are well known, respectively, to control such behaviors in cancer cells. While these observations would support, at least initially, the proposed triangular relationship “oleamide—ion (Na+ and K+) channels—cancer,” much more work remains to be done to verify it more directly.
Importantly, oleamide has also been shown to enhance the anticancer effects of chemotherapy. Thus, Jiang et al. demonstrated that oleamide caused enhanced antiproliferative effects on MCF-7 BCa cells pretreated with doxorubicin (Adriamycin). This effect was suggested to be mediated by uncoupling of gap junctions (GJs). 54 Indeed, oleamide is also a strong inhibitor of GJs (see section on Additional Targets of Oleamide) and the role of cytochrome c in biosynthesis of oleamide could contribute to initiating the apoptotic pathway from mitochondria. Thus, GJ communication could be blocked by mitochondrial metabolic elements activated by oleamide. 1 Nevertheless, there may be an ionic component since GJs are well known to be controlled by Ca2+ and pH. A nanotechnological study also showed that oleamide could increase the antiviability effects of cytotoxic drugs on MCF-7 cells. 55 Further details of this approach are given in Colorectal Cancer section.
In contrast to oleamide, OA and most of its other derivatives appear to have mixed effects and could promote BCa cell behavior. 56 Thus, treatment of MCF-7 cells with OA increased their viability and increased proliferative activity. 57 It is not clear if these effects involve Kv and/or Kir channels, but other fatty acids are known to promote K+ channel activity. 58 In another study, on MDA-MB-231 cells, oleate, a derivative of OA, caused an increase in the level of cytosolic Ca2+ and induced phosphorylation of protein kinase B. 59 Calcium signaling and protein kinase modulators contribute to the proliferation capacity of BCa cells. Furthermore, in media containing OA, adhesiveness of MDA-MB-435 BCa cells increased while in the presence of calphostin C, an inhibitor of Ca2+-dependent protein kinase C (PKC), cellular adhesiveness decreased. It was concluded that the effects of OAs on cellular invasiveness and adhesion were PKC dependent. 60
In conclusion, oleamide consistently suppressed while OA could (also) enhance BCa cell behaviors. Further work is required, however, to substantiate these findings and determine the degrees to which ionic mechanisms may contribute to such effects.
Colorectal cancer
Effects of oleamide on CRCa have been studied mainly through plant extracts. In one study, oleamide-rich fruit of Ziziphus jujuba was shown to delay development and progression of colon cancer along with suppression of tumor inflammation in a mouse model. 61 The effect of oleamide on CRCa has also been investigated in relation to ulcerative colitis (UC), which is known to have a positive association with CRCa. 62 Acetic acid-induced UC in a rat model indeed was associated with decreased levels of oleamide. 63 Treatment with a nanoformulation of a butanol extract of Acacia saligna restored metabolite levels in the inflammatory cascades to normal. 63 In an in vivo rat model, conjugated linOA inhibited the growth of CRCa (Caco-2) cells and this was suggested to occur, at least in part, through increased production of oleamide. 64
Some work has also been done with OA and has produced mixed effects. A high level of OA was observed in plasma membranes of cells from CRCa patients and this could be related to diet. 65 OA increased proliferation and mitogenic activity of CRCa Caco-2 cells. 66 In contrast, another study suggested that OA could inhibit the proliferation of CRCa cells by increasing both release of Ca2+ from intracellular stores and intracellular ROS production or caspase 3 activity. 67 An antiapoptotic effect has also been described. 68 An earlier study had also shown that OA could inhibit development of colon cancer in a rat carcinogenic (1,2-dimethylhydrazine) model. 69
OA has also been used in combination with cancer drugs. In an interesting nanotechnological approach, OA nanoparticles (NPs) were coated with hyaluronic acid (HA), which could then be coupled to two drugs, AZD6244 (a mitogen activated protein kinase [MAPK] inhibitor) and cisplatin (a DNA damaging drug). Release of the drugs from the NPs was much greater in an acidic environment (pH = 5.5), compatible with the low pH of the tumor microenvironment. Importantly, application of OA–HA NPs to human colon cancer (HTC-116) cells with and without the drugs suppressed cell viability. However, compared with the drug cocktail alone, the potency of the combination involving OA was more than 10-fold greater. 55
From the available evidence, similar to BCa, we conclude that oleamide produced a range of inhibitory effects on CRCa cells. On the other hand, the effects of OA, again, were mixed.
Broader Aspects
In this section, we outline a number of issues related to oleamide, ion channels, and cancer, for further awareness.
Other PFAMs
As noted in the Introduction, there are several other PFAMs. Of these, the “cannabinoid” anandamide is the best known. Independent of cannabinoid receptor-1 (CB1) and cannabinoid receptor-2 (CB2) receptors, anandamide acts on a variety of targets, including transient receptor potential vanilloid channel subfamily 1, peroxisome proliferator-activated receptor gamma, K+ channels, VGSCs, and Ca2+ channels. 70 Cellular effects included inhibitions of adhesion and migration of BCa cells 71 and proliferation of CRCa cells. 72 Anandamide can also associate with oleamide functionally as in sleep. 73 In addition, the apoptotic effect of oleamide on neural crest cells was enhanced by anandamide. 74 Linoleamide, a structural analog of oleamide, was shown to produce apoptotic effects through activation of Bax and DNA fragmentation in MDA-MB-231 BCa cells. 10
In addition, linoleamide, together with oleamide, showed potent inhibition of sarco/endoplasmic reticulum Ca2+-ATPase (SERCA) in primate kidney (COS-1) cells. 5 Finally, as already noted, natural sources have contributed significantly to the understanding of PFAM effects. Thus, palmitamide, erucamide, linoleamide, and oleamide were identified in extracts of Ericaria amentacea and Ericaria crinita plants and shown to have strong antioxidant properties, including synergistically “entourage effect”. 75
Additional targets of oleamide
In addition to sodium and potassium channels, oleamide has been shown to act on a variety of other targets including, importantly, G-protein coupled (CB1 and CB1) receptors for cis-oleamide. 76 Downstream signaling pathways included adenylyl cyclase, focal adhesion kinase, MAPK, and nitric oxide synthase. 77 Biological responses to oleamide also could involve GJs, serotonergic receptors, and gamma aminobutyric acid type A receptors. 78 Interestingly, as mentioned in Potassium Channels section, in contrast to oleamide, OA stimulated POMC neurons in the hypothalamus and decreased food intake through KATP channel inhibition. Such opposite effects of OA and oleamide may be explained, at least in part, by impact on ion channels and association with contrasting activities of cis- and trans-oleamide. Another well-known target of oleamide is SERCA and downstream Ca2+ signaling. 5 In RG2 glioblastoma cells, oleamide produced antiproliferative/apoptotic effects, which were suppressed by Ca2+ chelators. 11 In glial cells, on the other hand, the effect of oleamide was independent of Ca2+ and appeared, instead, to involve change in membrane fluidity. 79
GJs have also received considerable attention as potential targets of oleamide, several studies showing inhibitory effects. GJs can contribute to the cancer process in a wide range of ways, including controlling cellular proliferation, epithelial–mesenchymal transition and apoptosis, including in relation to chemotherapy.53,80 Finally, nitric oxide could be an intermediary in the action of oleamide, as in vasorelaxation. 81 In conclusion, oleamide, and probably other PFAMs, are associated with a wide range of targets and intracellular signaling mechanisms. It is highly likely that these are connected by common factors like Vm and Ca2+.
Effects on other cancers
Earlier (Effects on Cancer Cells section), we described in some detail the effects of oleamide on breast and CRCa. Various effects of oleamide have also been described on other cancers. For example, in pheochromocytoma and T24 bladder cancer cells, oleamide (and linoleamide, a structural analog) increased the level of intracellular Ca2+ by release of Ca2+ from internal stores as well as influx from outside. 8 However, the possible impact of these events on the cancer process was not studied. Two oleamide derivatives (MI-18 and MI-22) suppressed viability of metastatic Bl6 melanoma cells by blocking their GJ coupling. 9 Interestingly, oleamide also enhanced significantly the sensitivity of p53-mutant lung cancer cells to irradiation and increased antiproliferative and apoptotic effects through caspase activation. 82 Thus, oleamide and probably other PFAMs can affect a range of cancers and cell behaviors associated with the cancer process, but more work is required to broaden this knowledge.
Conclusions and Future Perspectives
Our overall conclusion is that there is indeed a possible broad basis for a functional triangular relationship involving PFAMs, ion (Na+ and K+) channels, and cancer cell behavior (Fig. 2). As regards PFAMs, we covered mainly oleamide for which most evidence relates to its acute effects. These depended on dose and exposure time as well as the nature of the cell under investigation. Thus, VGSC and potassium channels were shown to be significant targets, and these could be linked to the anticancer effects of oleamide.
Importantly, and not surprisingly, long-term effects are also possible. Indeed, application of oleamide to mouse neurons caused changes in levels of transcription factors, resulting in induction of c-fos messenger RNA. 83 Expression of c-fos in neurons is known to regulate synaptic activity, including during sleep.84,85 In addition, ion channel (including VGSC) activity itself can lead to gene expression changes, including autoregulatory effects.86,87 As well as determining the possible long-term effects of oleamide, more work is needed to elucidate its effects in vivo.
As noted in the Introduction, oleamide can be found in diet as a derivative of OA and also occurs in high amounts in various plants. Its anticancer effects can occur at least partially through modulation of ion channels, in common with several other natural agents.88,89 More broadly, dietary and nutraceutical compounds are important components of integrated management of difficult-to-treat cancers, for example, pancreatic cancer.90–93 Accordingly, understanding their mechanisms of action, including in relation to possible interaction with existing clinical treatment modalities, could ultimately be of significant benefit to human health.
Footnotes
Author Disclosure Statement
The authors declare no conflicts of interest.
Funding Information
No funding was received for this article.