
Abstract
Bioelectricity has been studied since the 18th century in every branch of the tree of life. Bioelectricity is involved in cell growth, proliferation, and behavior, which at a tissue-scale level translates in dramatic tissue rearrangements during embryology and somatic development. Although ion fluxes can be measured in single cells, it is not unanimous that a single cell may create and sustain different bioelectrical states within itself by means of electrochemical nonequilibrium phenomena. We address this possibility, with a focus on the pollen tube as a biological model. Pollen tubes are the subject of intense research given its unique properties, evolutionary streamlined to very fast apical growth and sensitivity to external cues that affect chemotropic responses. Pollen tube's functions rely on conspicuous tip-focused ions dynamics, involving the formation of steep ion gradients, with ion concentration differences over an order of magnitude when compared with the shank cytosol. These gradients are thought to be based on the spatial segregation of ion transporters, channels, and pumps along the cell, creating distinct electrochemical environments at the tip and the shank. But how polarity is generated and maintained is still a matter of debate. In the past we hypothesized that opposing electrochemical forces of depolarization at the tip and hyperpolarization in the shank could create a membrane potential gradient spanning from the tip to the shank that would be part of feedback mechanisms essential to cell polarity in these and likely other cell types. In this study, we review the latest progress on understanding apical growth from the perspective of bioelectricity-driven morphogenesis.
Introduction
Bioelectricity is as old as life. Since the beginning of the life primordia as we know it, when cells separated from the environment by a thin lipid bilayer impermeable to ions, an electrochemical imbalance of charges must have formed with the outside of the cell, giving rise to a bioelectrical potential stored as a membrane potential. Throughout evolution, ion channels have evolved to tap into this electric potential to perform work in the form of transport in and outside of the cell, but also as a way of controlling the bioelectrical state of the cell, that is, the amount of energy stored that way. Bioelectricity is carried by simple inorganic ions such as H+, K+, Na+, Ca2+, and Cl−, rather than by electron-based electricity because membranes are permeable to electrons and could not easily confine and concentrate electrons in the cytosol. All living systems have this self-generated electrical dimension and losing the ability to realize it eventually leads to cell death. 1
It is not surprising that from an evolutionary perspective, bioelectricity can be hypothesized to be one of the first signals used by primitive cells to monitor membrane integrity. When considering unicellular stages, in which organisms are challenged by environmental fluctuations, the maintenance of the plasma membrane integrity is a matter of life and death. Any small rupture in the plasma membrane, even if repaired in time to allow survival, would always imply the loss of some intracellular materials and the dissipation of charges from the intracellular space to the exterior, attenuating the transmembrane potential. Over time, these small depolarization events might have been the common origin and the first signals for membrane repair after injury mechanisms. 2
Bioelectricity in Plant Physiology and Development
The earliest documented bioelectric phenomenon was the electrical discharge of certain types of fish, with the first reports of these events dating back to Aristotle (384-322 BC) in Historia animalium (cited in Ref. 3 ). In the 18th century, bioelectrical phenomenon started to be formally studied by Luigi Galvani, who conducted experiments with dead frog legs, where muscle contraction could be triggered by external electrical induction. 4 Galvani thought that the nature of the contraction was caused by a sort of “animal electricity,” different from the “frictional” electricity. 5
Later, Alessandro Volta repeated these experiments, to conclude that the frog contractions were more evident when using a bimetallic conductor, and that the electricity involved in this phenomenon was of the same kind as the one produced by the electrostatic machines, concluding that the frog anatomy was just a very sensitive detector for electrical phenomena. 6 In 1873, Burdon-Sanderson performs the first extracellular measurement of an extracellular action potential in the Venus flytrap, 7 inspired by the correspondence with Charles Darwin that had made many observations on insectivorous plants. In 1901, Jagadish Chandra Bose, a physicist in training but a biologist by heart, started experimenting with bioelectric potentials and movements in plants, believing that the conserved essence between the physical inorganic world and the world of living was to be found in the action of electromagnetic waves. 8
For the next 30 years, Bose performed hundreds of experiments on electrical signaling in plants and gifted posterity with works such as “Research into the Irritability of Plants” 9 and “The Nervous Mechanisms of Plants.” 10 One of the main conclusions from these experiments using Mimosa pudica (telegraph plant), known by the drooping movement of the leaves after mechanical stimulation (thigmotropism), was that electric stimulation alone triggers the same movement. 9 Bose proposed that plant's electrical communication was done through the phloem and that electrical stimulation mimicked mechanical stimulation.
Once it became established that electric responses are evoked by certain stimuli, the quest to understand the electrical signals that trigger movement shifted to understanding the biophysical and chemical components that electrical propagation at a cellular level require. Biophysicist Julius Bernstein, which is largely recognized by his Membrane Hypothesis, correctly proposed that excitable cells are surrounded by a membrane selectively permeable to K+ ions at rest, and during excitation permeable to other ion species that now we know to be mainly Na+ in animals and H+ in plants. 11 In 1930, the first ever intracellular measurement of an action potential is recorded in the green algae Nitella cells, 12 two decades before the first recording in an animal cell. 13
As the studies of electrical propagation advanced, other views on bioelectricity branched out to morphogenesis and to processes that do not require communication at a long distance but rather local cell-to-cell communication. Lund's seminal work showed that the establishment of an electrochemical polarity was fundamentally associated to the development of morphological polarity in the brown algae Fucus spp. egg, because it could be controlled and directed by the application of a difference of electrical potential from external origin. 14
Between the 1930s and 1940s, H.S. Burr focused on measuring and correlating changes in voltage gradients with future developmental patterns in a wide range of systems, including root growth in corn. 15 His views were aligned with Lund, in which electrical phenomena in the living systems are fundamental and are determiners of pattern organization. A landmark experiment, worth mentioning for the ingenuity of its conception, was performed by Lionel Jaffe, using Fucus spp. eggs (Fig. 1). 16 Fucus eggs germination is triggered by the directionality of light, followed up by the bulging and outgrowth of what will become the rhizoid after the first cell division.

Measuring extracellular ion fluxes of Fucus eggs in series.
However, Fucus eggs are relatively small to be studied with the electrodes at the time so, Jaffe created a small electrical circuit with a capillary glass filled with 200 eggs (Fig. 1A), such as dry batteries in a serial stack, as to amplify the electrochemical signal during morphogenesis to the point of being measurable by the available methods in those days. By shining a light from one side, all the eggs in the glass capillary synchronized their development and orientation toward one side (Fig. 1B), and by measuring at both ends of the capillary, it was possible to measure up to −70 μV coming from this developmental process (Fig. 1E). 16
Generating data about extracellular electric field at single-cell level is difficult because the biological signal can be overridden by technical problems such as insufficient signal/noise ratio, artificial ion movement in the medium, Johnson–Nyquist electric noise and instrumental drift. These limitations led to the development of noninvasive vibrating probes to measure extracellular electric currents 17 and, subsequently, ionic fluxes surrounding cells.17,18 These techniques were instrumental to confirm the hypothesis that electric currents and ion fluxes precede pattern formation and accompany polarized growth and cell migration. 19
These type of electric currents were mapped in fungi, water molds,20,21 root hairs 22 and pollen tubes 23 undergoing polarized tip growth, where ion currents were shown to traverse the cell parallel to their polarization, creating an asymmetric front-rear pattern, in which currents enter the back of the cell and exit the polarized site, accordingly to their growth. These patterns can also be found at a multicellular level, where bioelectrical currents are always associated with large-scale tissue behavior, for example, in coleoptile elongation, 24 root growth, 22 root gravitropism,25,26 secondary root growth, 27 and root wounding.28,29
These pioneer studies in bioelectrical patterns showed that spatially organized currents surround plant cells and tissues when morphogenesis and/or polarity take place, leading to the speculation that the resulting endogenous electrical fields could guide cellular processes and tissues during development. A possible mechanism underlying this hypothesis would be that the generated steady electrical patterns during these phenomena arise from synchronous action and spatial segregation of ion channels, which in turn create areas of influx and efflux of different ion species. As a result, electrochemical imbalances would be created at morphogenetic hotspots that are mechanistically regulating cellular processes.
Electrical Morphogenesis in Pollen Tubes
Pollen is angiosperm's male gametophyte. When pollen grains germinate, the vegetative cell emerges creating a cylindrical outgrowth, growing only on its tip (apical growth) and forming the pollen tube. They are arguably one of the fastest growing cells in nature and became central models when it comes to study processes involved in apical growth and polarity. Pollen tubes' function is to grow rapidly, navigating through the female tissues toward the ovules and discharge the sperm cells they carry to allow fertilization and seed formation. Pollen tubes are able to germinate at rates of up to 50 μm/s for a few seconds 30 and maintain prolonged sustained growth rates up to 3.0 μm/s 31 while steering through the female tissues and transducing anatomical and chemical cues into chemotropic responses.
The fast cell growth and the precise targeting of ovules require that growth and cell polarity are well coordinated through cellular mechanisms based on electrochemical phenomena that are expressed in the form of polarized ion fluxes of Ca2+, H+, K+, and Cl− 32 (Fig. 2). The tip-focused ion gradients are formed through the spatial distribution of ion fluxes: Ca2+ and H+ enter the cell at the apex and exit in the tube shank, K+ and Cl− move in the opposite direction, entering the shank and leaving the cell at the tip. 31 In vitro, the magnitude of these fluxes depends on the species, growth conditions, and frequently shows sustained temporal oscillations on growth rate.33–35
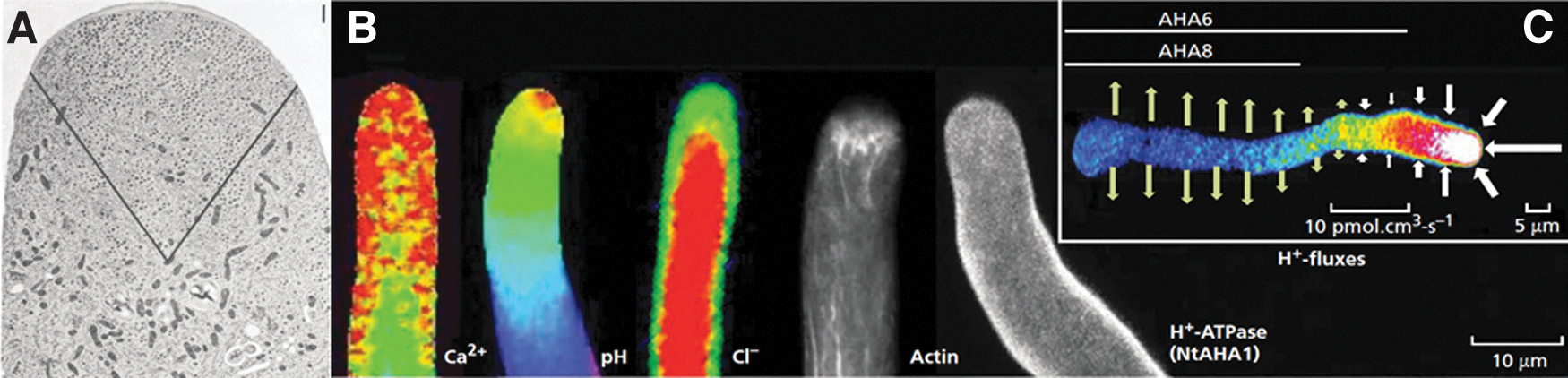
The ionic basis of pollen tube growth.
In all species tested to date, the magnitude of the Ca2+ and H+ apical influx range between 1 and 50 pmol·cm−2·s−1,35,39,40 whereas K+ efflux varies between 30 and 400 pmol·cm−2·s−1 (Dias and Feijó, unpublished). For anions, the apical fluxes exceed those of Ca2+ and H+ by three orders of magnitude, oscillating between 50 and 8000 pmol·cm−2·s−1.39,41 Thus, in the pollen tube tip, these fluxes could be coordinated not only to reduce the ionic strength of the apical cytoplasm by means of the high anionic currents, but also to depolarize the plasma membrane due to the influx of positive charges (Ca2+ and H+) and the efflux of negative charges (Cl−/anions).39,42 While the net ion currents at the tip are predicted to depolarize the membrane, the pollen tube (PT) shank is under hyperpolarization forces, because H+ and Ca2+ are leaving the cell and anions are going in, supporting the notion that this region of the cell may be more negative.2,32
The net ion flux profiles along the PTs show that the higher fluxes at the tip decrease toward the shank and at a certain point reverse direction, usually between 15 and 25 μm from the tip (depending on the species). The spatial distribution of ion fluxes and the reversal point is thought to be formed by the spatial segregation of ion channels/pumps along the membrane, potentially creating an anisotropic electrochemical potential as though the tip and the shank are contiguous compartments rather than a continuous one. The best example to date of this spatial segregation is the distribution of the autoinhibitory H+-ATPases (AHA) pumps, both in Tobacco and Arabidopsis PTs, where AHA pumps are evenly distributed in the shank and never progress to the tip area (Fig. 2C).36,37
Mutations in three specific and highly expressed AHAs in pollen tubes show a severe fertility phenotype because the PTs stop growing prematurely, soon after entering the ovary. Analysis of membrane potential reveal that without the electrogenic AHAs localized in the shank, pollen tubes are depolarized, H+ fluxes are compromised, which consequently affects anion influxes at the PT tip too. This study showed not only the importance in keeping the spatial distribution of ion fluxes for the correct development and morphogenesis in pollen tubes but also highlighted the interconnectedness of the ion homeostasis with the bioelectrical state required for growth. 37
Early on we have speculated around a model in which the combined effect of these electrochemical conditions could self-organize by membrane electrophoretic separation of protein's superficial charge. 43 Later we detailed the model for the pollen tube using Hodgkin–Huxley formalisms and incorporating the localization of the H+ Pumps that we meanwhile described to propose the possible existence of a cytosolic electric field located at the tip (Fig. 3).

Model for ion fluxes around the pollen tube: an active transport system at the tube, and a passive transport system at the tip. The top scheme shows the inorganic ion fluxes around the pollen tube of tobacco (Nicotiana tabacum) as recorded using the ion-specific vibrating probe technique. Ca2+ and H+ leave the pollen tube at the shank, through ATPases and enter the tip, through channels for Ca2+, and nonidentified transporters for H+. Cl− enters the tube through transporters and leaks from the tip through channels. Bottom scheme shows a Hodgkin–Huxley formalism for the electric equivalent circuit of the hypothesized polarization of transporters in the same system. Depending on the resistivity values of cytosol (Gx) this model could sustain a cytosolic electrostatic field within the apical volume of the tip (adapted from Michard et al. 39 ).
A conceptual prediction of this model is the existence of a standing gradient of membrane voltage, a concept generally considered not to exist in dimensions over 10 nm, 44 given that fast rectification either by voltage sensitive channels or cytosolic buffers should occur.
At a cytological level, the ion reversal point correlates with the clear zone (Fig. 2A), the most dramatic point of cytoarchitectural polarization, in which the bigger organelles in the cell are excluded from the volume of the tip, reverting to the back of the cell by cytoplasmic streaming. Although the cytoplasm in PTs is subjected to a reverse fountain streaming where cell components move toward the tip along the cortex, and stream backward through the lumen, there is a clear sorting of organelles based on their size, and the clear zone consists mostly of small secretory vesicles (Fig. 2A).38,45–47
Spatial-temporal image correlation spectroscopy showed that in the apical region, these small vesicles exhibit irregular dynamics pointing to a vesicle transport mediated by diffusion and/or advection by the cytosol rather than the motor proteins trafficking vesicle movement in the shank. 45 Transmission electron micrographs show evidence that the level of vesicle compaction and fluidity at the tip are higher than in the shank where the cytosol is subjected to streaming.38,48 Although there is no physical separation between the volumes of the tip and the shank, there are levels of cellular organization that create a clear separation in the cytosol and possibly in medium resistivity. 49
In connection with these facts, we hypothesized that the ion flux density and the cytosolic architecture can result in a longitudinal membrane potential gradient (MPG). This hypothesis has been proposed before any theoretical approaches have demonstrated that it is plausible.2,32 When taking into consideration the dominant anion fluxes at the pollen tube tip and assuming an average flux density of 1000 pmol·cm−2·s−1, 41 the estimation of current density is about 100 μA·cm−2, comparable with that of an action potential in muscle and neuronal cells.2,50 Such current density traversing the pollen tube's apex can result in a longitudinal gradient similar to that observed in nerve impulses, but in PTs would cause a standing membrane depolarization at the PT tip.
To put into perspective, an action potential depolarizes the membrane when Na+ channels open letting Na+ into the cell and while it propagates, the cell repolarizes using K+ efflux to get to the resting state. 51 While in action potentials, these successive events happen in time, in PTs the similar mechanism might happen in space, where the tip is subjected to depolarizing currents while the shank is subjected to hyperpolarizing/repolarizing currents, thus creating two spatial distinct electric polarizations along the same cell. The co-option of the same molecular machinery in terms of space or time have also been considered when comparing PTs with stomatal functions, in which the hyperpolarization and depolarization of guard cells is usually triggered by day and night and other environmental factors (Fig. 4). 32
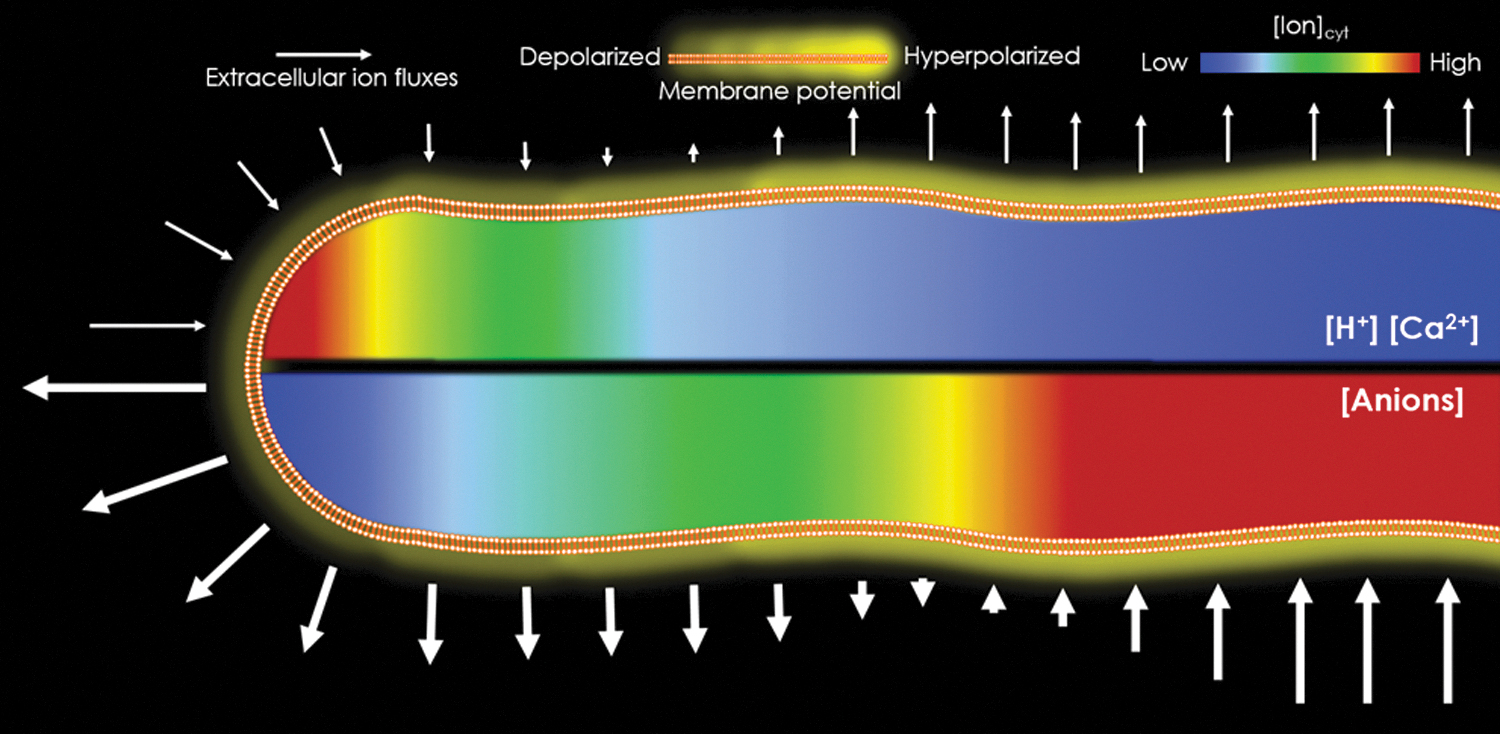
Electrochemical profiles of ion fluxes and cytosolic gradient during pollen tube growth sustains the notion of the existence of an MPG along the membrane. Ion fluxes at the pollen tube tip would sustain depolarizing currents, whereas at the shank the currents would hyperpolarize. Consequently, changes in ion concentrations in the cytosol would be created, generating tip-focused gradients. Energization for this spatial separation would be essentially based on the segregation of H+-ATPases from the tip. MPG, membrane potential gradient.
There have been multiple attempts of measuring membrane potential in the pollen tube shank by using intracellular microelectrode recordings, with measured values ranging from −55 to −130 mV in Zea mays, 52 Agapanthus umbellatus, 53 Lilium longiflorum, 54 and Arabidopsis thaliana. 55 Unfortunately, microelectrode recordings at the pollen tube apical region are not feasible because of the rapid expansion and friable nature of the nascent cell wall, provoking the tubes to burst when impaled due to the high turgor pressure. 32
Nevertheless, preliminary evidence has been reported for membrane potential depolarization at the pollen tube tip by employing voltage sensitive dyes.56–60 With the aid of DiBAC43 and Di-4-ANEPPS dyes, a steep longitudinal gradient of membrane potential was observed at a distance between 4 and 20 μm from the tip in tobacco PTs; unfortunately, calibration and quantitative imaging precision were not adequately addressed to date, and the data from these studies, although pioneer, should be read as a qualitative work by relative comparison of fluorescence. 57
An apical depolarization in tip-growing cells has been demonstrated with microelectrode impalement in hyphae of the fungus Neurosopora crassa. 61 Neurospora hyphae are divided into “compartments” by incomplete septa, and the cytoplasm streams continuously toward the growing tips, similarly to pollen tubes. In these studies, authors have shown that the tip compartment is usually 10–30 mV more depolarized than the contiguous compartment. Although cytologically similar to PTs, the electric potential differences are measured between compartments at a longer distance of ∼100 μm, likely attributed to the slower growth rates and consequently the lower ion current magnitude required for hypha tip growth.
The requirement of bioelectrical phenomena in pollen tube morphogenesis led various teams to delve on the effects of exogenous electric fields in cell polarity and growth. Pollen from Agapanthus,62,63 tobacco and tomato, 64 and Camellia 65 were incubated under the effect of electric fields to analyze their galvanotropic response. All species grow toward the anode side when on small volumes, but larger chamber designs resulted in Agapanthus pollen tubes growing toward the closest electrical terminal, and at random in the middle of the chamber where the electric field wanes, raising the issue that various effects provoked by the currents may be sensed, namely the formation of specific ion gradients in the liquid medium.
Consistently, attraction/repulsion by ion concentrations/ionophores can mimic some of these effects 66 and modifying cytosolic pH by pressing gramicidin to the plasma membrane provokes orthogonal turns of the tip. 36 Genetic screen for differential galvanotropic responses in the fission yeast found that cells mutated for a H+-ATPase (PMA1) would lose their galvanotropic reaction, which usually bends orthogonal to the electric field, further suggesting that either cytosolic H+ concentration or localized plasma membrane potential differences are essential in defining the growing axis of tip growing cells. 67
These conclusions are currently interpreted as an epiphenomena, because the mechanisms underlying cellular responses is elusive, and often speculated rather than experimentally demonstrated. A recent hypothesis posits that membrane potential plays an upstream role in galvanotropism 68 because one of the most immediate effects on cell physiology upon exposure to electric field (EF) is an asymmetric change in membrane potential that can be observed using voltage-sensitive dyes. 69 In a cell electrically silent and with low conductance the side facing the anode hyperpolarizes, whereas the side facing the cathode depolarizes. This hypothesis places membrane potential asymmetries in the spotlight to explain galvanotropism, but how would that explain different directions in tropism toward the anode or the cathode is unknown.
In cells expressing voltage-gated channels, the membrane potential asymmetry would trigger currents that could redefine the default tropism pattern, providing a possible explanation by assuming that the direction of pollen tube growth depends on the balance between contrasting forces created by the different ion currents driven either by passive diffusion and/or voltage-gated ion channels. The effects observed in pollen tube galvanotropism responses could be mimicking the natural processes of having a longitudinal tip-focused MPG.
The Possible Effects of a MPG in Morphogenesis
The existence of a standing MPG could affect many other cellular processes because it presupposes the creation of an electric field at the PT tip. Any sustained electric field such as the one generated by an MPG would have an immediate effect in many cellular components possibly triggering galvanotropic responses.69,70 Pollen tube growth requires exocytosis from the small apical vesicles that accumulate in the apical cytoplasm. There is evidence showing that these vesicles when isolated and exposed to an EF move toward the anode side. 71 The reason of this movement to the anode side comes from the lipid content of these vesicles that is negatively charged.
Apical vesicles contain high amounts of enzymes of the phosphoinositide metabolism that are implicated in PtdIns4P biosynthesis, with negative nominal charge.72–74 Thus, vesicles may be drawn to the tip just based on superficial charge, where the Ca2+ inward currents occur and can trigger exocytosis. This mechanism resonates with presynaptic neuron, in which the depolarizing action potential triggers voltage-activated Ca2+ channels that in turn trigger exocytosis of neurotransmitter vesicles into the synaptic cleft. 75 The transduction of the Ca2+ elevations is thought to occur through two proteins, Synaptogamin-1 and complexin, which interact with the Soluble N-ethylmaleimide-sensitive factor Activating protein REceptor (SNARE) proteins on vesicles. 76
Synaptogamin orthologs are highly expressed in PTs and RNA interference studies suggest that they have a role in pollen tube germination and growth. 77 Voltage-activated Ca2+ channels were never convincingly described in PTs, but in root hairs, another tip growing cell, there is evidence showing that such channels exist, and elicit Ca2+ inward currents at imposed voltages in the range of the root hair physiological conditions. 78 Thus, the existence of an MPG-generated EF at the PT tip may be important for sustaining growth by electrostatically carrying the small vesicles to the apex. This effect may be critical because there is evidence that the vesicle transport in the apical region of the tube is not mediated by motor proteins but can be driven by a combination of other physical forces such as diffusion, electrophoresis, and osmophoresis.2,45,46,48
An MPG-generated EF would not only act on the cytoplasmic components but also would have a direct effect on the charged membrane components that, if free to move laterally on the plane of the membrane, should undergo redistribution in response to an EF applied along the membrane. 79 The phenomenon of lateral electrodiffusion at the plasma membrane level has long been postulated as a possible mechanism underlying the effects of biogenic electric fields in developing organisms.80,81 Other studies revealed that when subjected to EF, acetylcholine receptors accumulate in clusters at the cathodal side in the muscle membrane, 82 and positively charged artificial fluorescent lipids (DiI) migrate toward the cathode side.83,84
Lateral electrodiffusion may explain partly the segregation of ion channels at the plasma membrane that gives rise to the flux profiles with different directions in the tip and in the shank. This way, the different ion channels with either positive or negative nominal charges could be maintained in certain parts of the membrane without invading each other's domains due to the effect of attractive or repelling forces from the EFs. Moreover, the reversal point of the ion fluxes may be an area where the intracellular EF is at the weakest point, which would make this volume an effective barrier for electrodiffusion.
The difference in voltage between the tip and the shank is also another possible ion channel regulatory mechanism, in which channels at the tip are subjected to a different voltage than in the shank. Even if some channels are not under the effect of electrodiffusion due to their nominal charge, the membrane potential difference can be sufficient to have them active at the tip and not in the shank and vice versa. For example, such effect is predicted to occur with SPIK K+ channels based on their voltage dependence. 55 The effect of electrodiffusion in anionic lipids is also an important factor for the control of ion channels because the lipidic microenvironments can confer stability to particular allosteric states of ion channels. 85
Some ion channels have an obligate requirement for one species of phosphoinositide, whereas others may accept a broader range of phosphoinositides or anionic lipids. 86 In some extreme cases, for example, the potassium channel Kv7.1, specific lipid species are required to couple the voltage sensing domain to the pore opening, resulting in pore closing in the absence of a specific lipid. 87
In conclusion, the MPG concept can give rise to new and testable hypotheses that will further our understanding of apical morphogenesis processes that rely on bioelectrical phenomena at a single-cell level (Fig. 5).
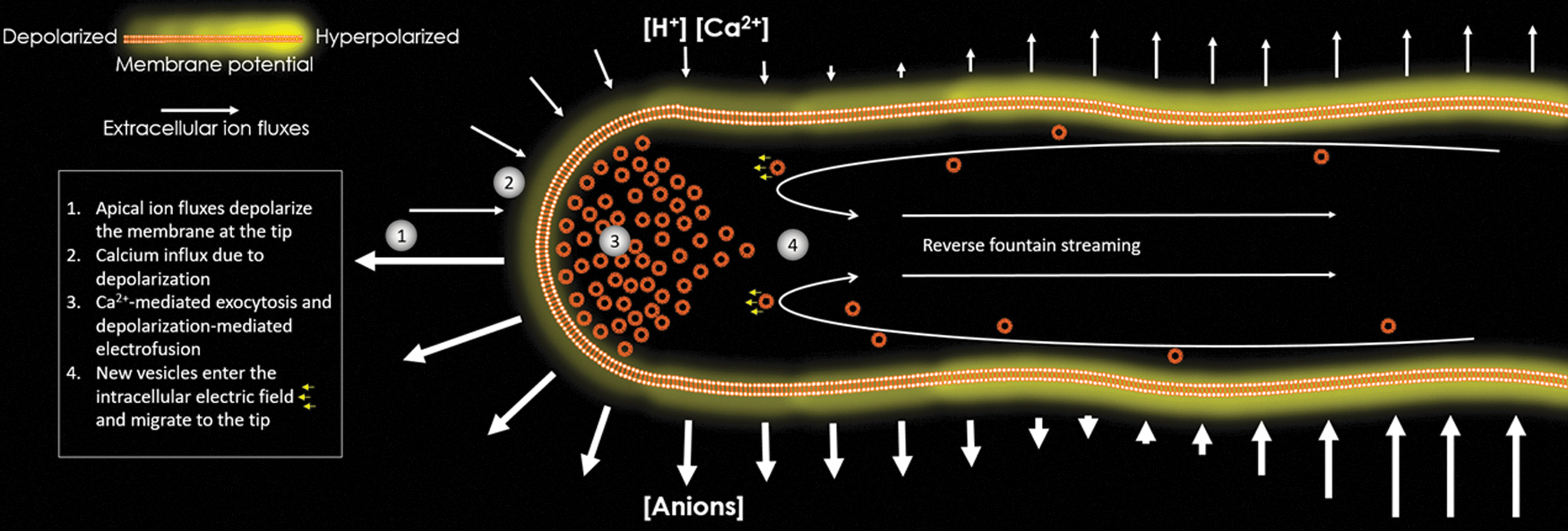
Model for the effects of electrochemical and voltage differences in cellular apical growth. (1) The apical ion fluxes and gradients create an MPG, which in turn, given the geometries of the ion concentration gradients, should mirror an electric field at the PT tip. (2) Ca2+ enters the tip either by depol-activated channels or other antiport/symport transporters. (3) The population of compacted vesicles at the PT tip undergoes Ca2+ mediated exocytosis and membrane depolarization facilitates this by electrofusion, a phenomena in which the membrane loses integrity and becomes destabilized, optimal conditions for fusion. (4) The MPG-induced pollen tube electric field at the tip recruits more vesicles from the fountain streaming movement because the lipid content of these vesicles is mostly anion lipids. Voltage-gated K+-channels along the pollen tube are activated according to their position along the MPG.
Co-Option of Depolarization Signals from Membrane Repair Mechanisms into Apical Growth
Our assumptions being correct, this model would also have ramifications into membrane repair. One of the first challenges that unicellular life faced was preserving membrane integrity and so, it stands to reason that one of the first signals transducing membrane integrity should be intracellular ion signals and bioelectrical fluctuations. When the plasma membrane is breached, the cellular electrochemical gradient is partly dissipated by electrochemical equilibrium toward the extracellular medium. This creates a local depolarization at the injury site. The ions that go out of the cell are mainly K+, Cl−, and other organic anions that are at high concentrations inside the cell thus they move outward in favor of the chemical gradient.
In contrast, Ca2+ is maintained at very low intracellular levels because of its cytotoxicity inside the cell, so it is actively pumped out creating a very steep chemical gradient from cytosolic submicromolar to extracellular millimolar concentrations. When the integrity of the plasma membrane is compromised, a swift influx of Ca2+ generates transient increases in cytosolic free Ca2+ concentration, localized to the injury site.88,89 Ca2+ influx is established as an absolute requirement for plasma membrane repair, namely by using the sea urchin egg model. 88
Molecular mechanisms decoding the Ca2+ signal in membrane repair mechanisms overlap in animals and plants, namely through (1) Ca2+-dependent cysteine protease calpain (DEKK1 ortholog in plants); (2) annexins, which react to Ca2+ by binding to phospholipids creating membrane-associated scaffolds; (3) Synaptotagmins,76,77 regulators of Ca2+-regulated membrane fusion events; (4) SNARE complex, which helps in vesicle docking and fusion.88,89 The influx of Ca2+ inevitably triggers these membrane-bound components to fuse small vesicles that are near the plasma membrane in a Ca2+ regulated exocytosis, subsequently patching the injury.
The consensus that membrane repair involves a Ca2+ transient ignores the bioelectrical component of membrane injury, but local depolarizations happening at injury sites create a transient electric field until repaired. Such integrated view would argue for a parallel evolution of depolarization and Ca2+ signals in relation to plasma membrane repair mechanism, possibly co-opted for other functions such as exocytosis-driven apical growth in pollen tubes or in synapses.
Analogies between pollen tubes and neuronal mechanisms have been elaborated. 90 Both are highly specialized and polarized cells with coincidental mechanisms dictating their functions, namely membrane repair mechanisms. Both PTs and neurons rely on depolarization signals to perform their functions; PTs may sustain an MPG at the growing tip, whereas neurons propagate a depolarization signal for action potential function. PTs and neurons rely on Ca2+ signals too, in one case to deliver vesicles that can create cell expansion, in the other case to fuse vesicles containing neurotransmitters.
Both systems require annexins, synaptogamins, and the SNARE complex to perform the fusion events that determine their functions. The similarities in these two processes are comparable with membrane repair mechanisms, except for the bioelectrical signal origin. While the depolarization during membrane injury comes from the breaking of the membrane, in PTs and neurons there are ion channels that create the depolarizing currents as though to mimick what would happen upon injury. In neurons it is well established that the depolarization triggers depolarization-sensitive Ca2+-channels that can trigger the downstream events of vesicle fusion, 91 but, in plants, such channels have not been identified yet.
Nevertheless, the Ca2+ oscillations at the pollen tube tip could produce the same effect of activating downstream fusion events as similar as the membrane repair mechanisms.53,63 In neurons, cells create depolarization signals to activate the relay of the chemical signal through vesicles, whereas in pollen tubes, spatially organized ion channels create constant depolarizing currents at the morphogenesis hotspot as to recruit a constant repair mechanism at the PT tip that will lead to apical cell expansion.
Depolarization signals are not all the same, and do not promote growth in every cell, but in pollen tubes—a highly specialized cell for fast growth—the local depolarization signals may constitute an organizing event at a cellular scale as strong and as well conserved as Ca2+ signals, which may activate downstream repair mechanisms leading to growth and polarity establishment.
Footnotes
Author Disclosure Statement
No competing financial interests exist.
Funding Information
J.A.F. Lab is supported by the National Institute of Health (R01-GM131043) and the National Science Foundation (MCB-1930165).