
Abstract


During my school days I had a passion for biology. I suppose at that time a nagging question—although I would never have phrased it this way—was: “how does life work?” As I progressed through my undergraduate studies my interests focused increasingly on plants and, in particular, on why plants grow where they do. Above-ground factors such as temperature and light obviously play a role, but so do soils. I performed my final-year undergraduate research project at the University of York on acid and calcareous (alkaline) ecotypes of a grass: Festuca ovina. Through hydroponic growth experiments I discovered that the ecotypes are differentially responsive to pH per se and not only, as previously thought, to the effects of pH on the solubility of various soil nutrients and toxic ions.
My interest in plant membrane transport was awakened, and with that the question of how plants energize cellular uptake of nutrients from the relatively dilute solutions that surround them. The models for animal epithelia involving co-transport with Na+ seemed unlikely to apply because most plants are happy growing in a Na+-free environment. As a result of elegant electrophysiological experiments on giant-celled charophyte algae, Kitasato 1 posited that the cell membrane might possess an electrogenic H+ pump, and the possibility therefore arose that the resultant inward-facing H+ electrochemical potential, or protonmotive force (pmf), might be used to energize uptake of solutes. Indeed, H+-driven uptake of sugars 2 and amino acids 3 was subsequently demonstrated for several algal and higher plant species.
H+-Coupled Ion Transport in Plants
There was clearly much more to discover, so in 1975 I decided to undertake graduate work on freshwater charophyte cells with Enid MacRobbie at Cambridge. The length of charophyte internodal cells is several centimeters and their diameter is typically of the order 1 mm (Fig. 1A). 4 This makes them very easy to impale with microelectrodes, and indeed early experiments on charophyte action potentials using this approach were performed almost a century ago. 5

Chloride-induced membrane depolarization in Chara corallina indicative of 2H+:Cl− cotransport.
When I joined the MacRobbie laboratory there was still considerable controversy surrounding the energization mechanism for uptake of inorganic solutes in these model cells. For the major osmotic component, Cl−, several hypotheses for its mode of entry into cells were circulating when I began my PhD work, including the possibility of a KCl “salt pump” and a proton-driven transport system that would have to operate with a stoichiometry of at least 2H+ per Cl− to effect net uptake of the anion. 6 However, isotopic measurements of Cl− influx suggested that any electrical impact of electrophoretic transport would be effectively too small to measure.
A solution to this issue of small Cl− influx was provided by parallel studies I conducted on isotopic Cl− influx. I discovered that if cells are starved of Cl− for a few hours, influx is reliably stimulated by a factor of 3–4. 7 Thus, I took my first dive into electrophysiology: Cl−-starved cells were impaled with a microelectrode and the response of membrane potential to the addition of Cl− then measured. A small and transient, but consistent, membrane depolarization of ∼10 mV was observed 8 (Fig. 1B), and this was independent of the presence of alkali cations in the bathing medium. The obvious conclusion was that each Cl− enters the cell together with at least 2H+.
The pmf would then be sufficient to power net uptake of Cl−, even at alkaline pH, because of the dominance of the highly negative membrane potential (−170 mV). Since that time, the general model has emerged for pmf-driven uptake of most essential ions and solutes across the plasma membrane of plants [although some well-documented examples of Na+ coupling have emerged; see Energizing Plant Plasma Membrane Potassium Transport with H+ (and Na+) section]. Indeed, the dominance of a “proton economy” that energizes the plasma membrane applies to all major nonanimal organisms: fungi and bacteria as well as plants.
Intracellular pH and the Electrogenic Proton Pump in Neurospora
Clearly, if plasma membrane transport is dominated in these organisms by the activity of a H+ pump, we need to understand more of its properties in situ, not only in energizing transport, but also in relation to controlling cytosolic pH. Carolyn and Clifford Slayman—who ran independent but collaborating laboratories at Yale University School of Medicine—had performed groundbreaking research on a fungal plasma membrane H+ pump from Neurospora crassa, with Carolyn working on the biochemistry of this H+-adenosine triphosphate (H+ATPase) 9 and Clifford on the electrophysiology. 10 Building on my results at Cambridge, I obtained a James Hudson Brown Fellowship in 1978 to join Clifford Slayman's laboratory at Yale—initially to study the role of the Neurospora plasma membrane H+ pump in the control of cytosolic pH.
Roger Thomas had recently ingeniously devised so-called “recessed tip” electrodes in which a sealed pH-sensitive glass pipette was fused within an aluminosilicate impalement pipette casing (Fig. 2A).11,12 The possibility of real-time measurements of cytosolic pH in an organism running a proton economy was enticing, although the fabrication of the electrodes and interpretation of the results proved difficult. To penetrate successfully the fungal hyphae, which are about 15 μm diameter (over an order of magnitude less than that of the snail giant neurons on which the electrodes had previously been used), the external tip diameter and dead space of the microelectrodes had to be minimized.
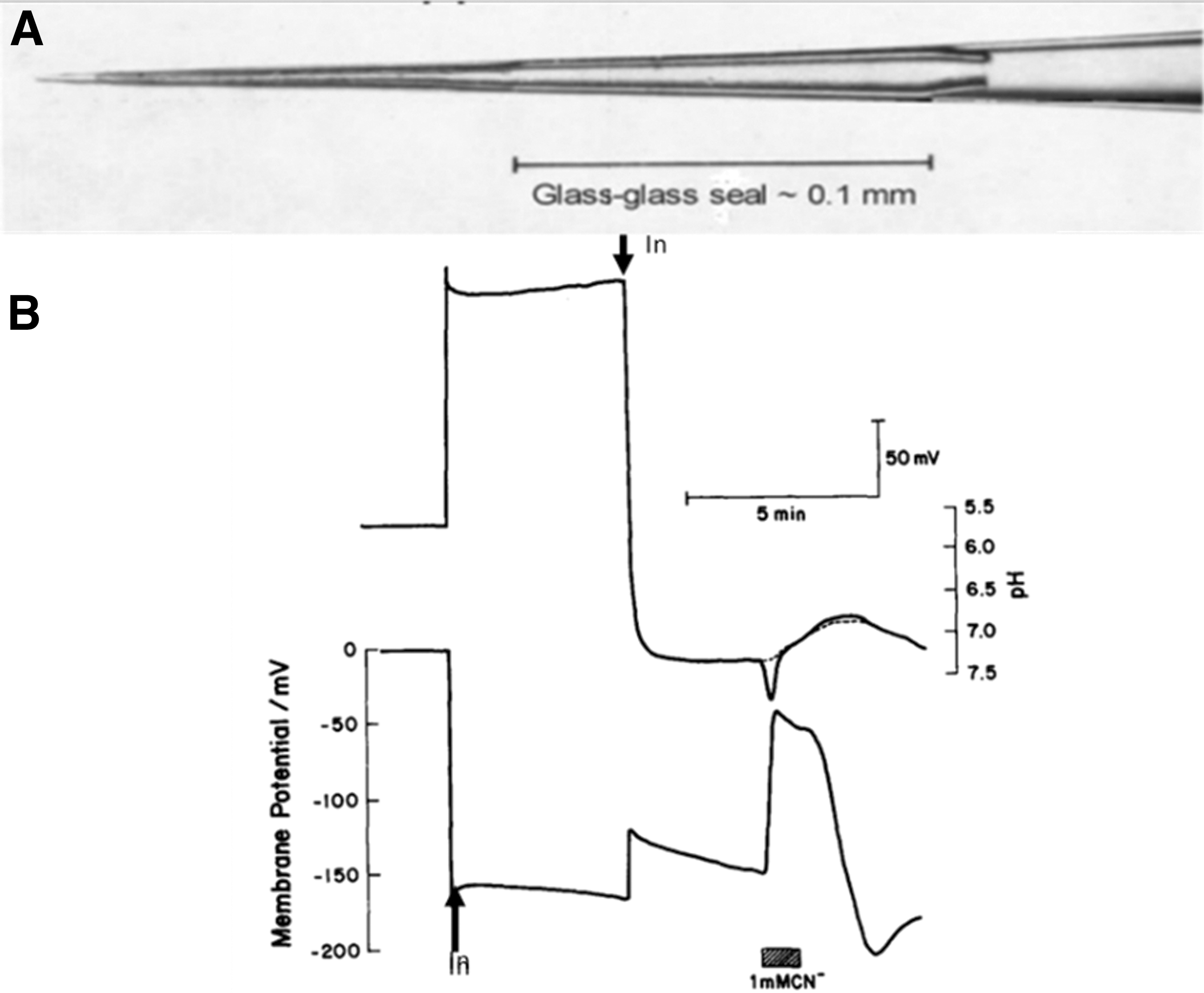
Recessed tip microelectrode measurement of intracellular pH in Neurospora crassa.
This further miniaturization resulted in an increase of the electrical (Resistance × Capacitance [RC]) time constant, which was typically in the region of 5–20 s. Since the signal from the pH electrode had to be passed through a differential amplifier to subtract the contribution of membrane voltage from the output of the pH electrode, rapid changes in membrane potential had to be quantitatively accounted for to derive the bona fide pH signal.
We devised a segmental linear algorithm to correct for the impact of rapid changes in membrane potential on the signal from the intracellular pH electrodes 13 and the results of the correction are shown in the example in Figure 2B. Before insertion of either electrode, the pH electrode registers the pH of the medium: 5.8. Insertion of the voltage-measuring electrode registers a membrane potential of −157 mV, which is accompanied by an equal and opposite deflection in the difference trace. Subsequent insertion of the pH electrode results in a transient depolarization of 47 mV and a measurement of intracellular pH, which stabilizes at pH 7.32.
Neurospora, similar to most other fungi, is an obligate aerobe, and addition of CN− results in rapid depletion of ATP and, with it, inhibition of the electrogenic H+-pumping plasma membrane ATPase and consequent membrane depolarization. The rapid change in membrane voltage is reflected in what appears to be a transient alkalinization, but after correction for the impact of the slow RC time constant of the pH electrode is actually the start of a CN−-induced acidification that is sustained until CN− is completely washed from the recording chamber.
I anticipated acidification of the cytosol in the presence of CN− because export of H+ across the plasma membrane was inhibited. Confirmation of this idea was borne out (Fig. 2B) although my reasoning was not correct. I found that application of the well-known inhibitor of plant and fungal plasma membrane H+ pumps—orthovanadate—had only small and variable effects on cytosolic pH (Fig. 3A). Furthermore, the CN−-induced acidification occurred at the same rate and extent whether or not there was an inwardly directed pmf across the plasma membrane (Fig. 3B). We concluded that in the absence of Krebs cycle activity, buildup of acidic glycolytic end products (e.g., lactate and pyruvate) must be the cause of cytosolic acidification and not uncompensated proton influx from the medium.
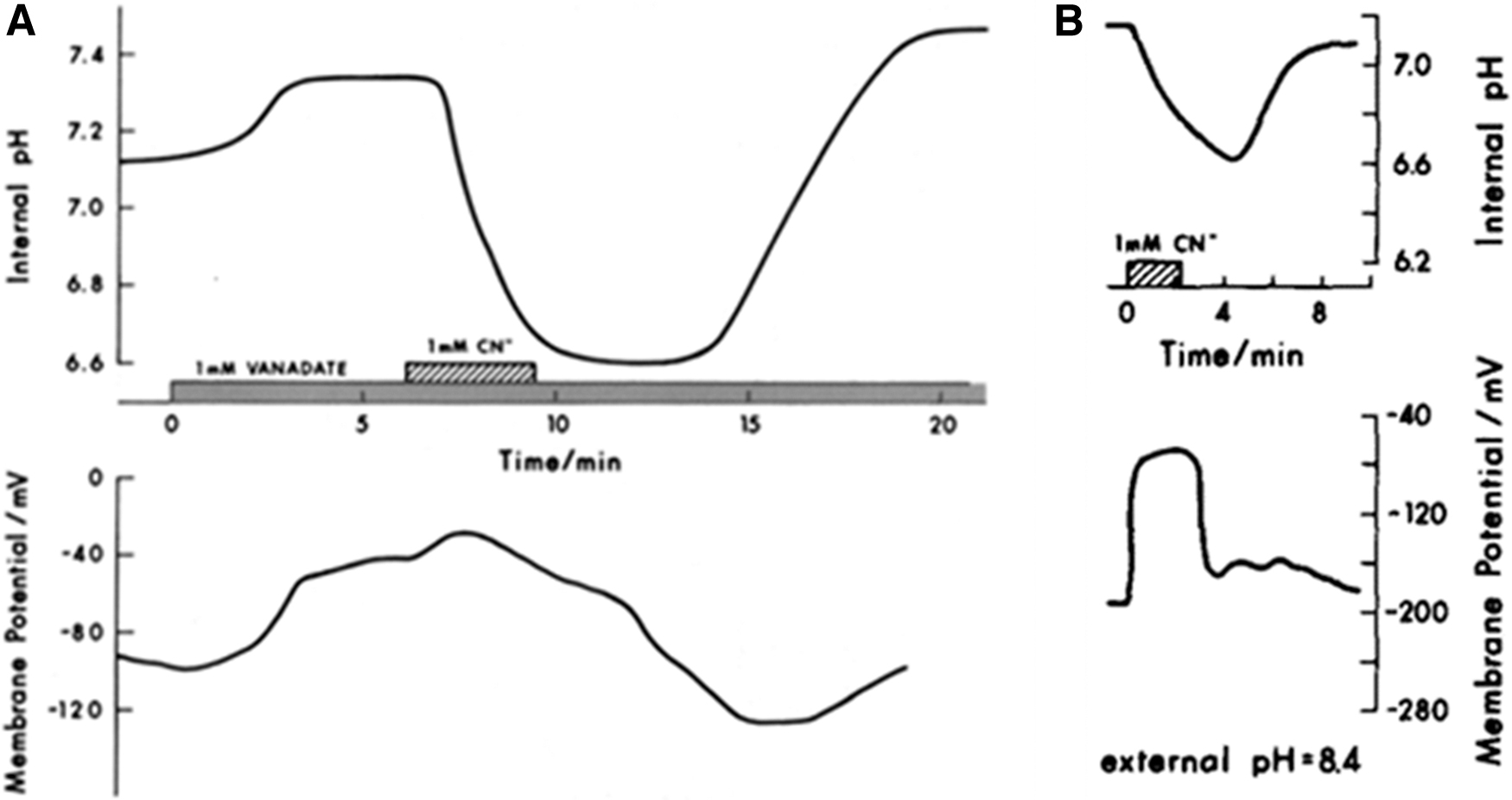
Evidence for a dominant role of oxidative metabolism in controlling intracellular pH in Neurospora crassa.
Cytoplasmic pH was linearly sensitive to changes in external pH, although with a slope of only 0.1 unit internal pH/unit external pH. This suggested strong buffering and/or regulation of cytosolic pH. We were able to use an established method to determine the cytosolic buffer capacity by application of a weak acid or a weak base.14,15 We used butyric acid and procaine as the weak acid and weak base, respectively. These compounds permeate the membrane freely in their nonionized form, and the magnitude of the pH change as a result of ionization in the cytoplasm, together with the calculated concentration of the ionized form in the cytoplasm, then enables estimation of the buffer capacity. A value of 35 mM H+/pH unit was derived, in close agreement with measurements on animal cells.14,15 The steep rise in buffer capacity was observed in both directions away from the control pH, which implied that side chains of acidic and basic amino acid residues are the major source of buffer.
Our ability to manipulate cytosolic pH with butyric acid led me to ask whether and how the electrogenic H+ pump controls pH. I was surprised that the principal electrical effect of butyric acid on the membrane was a depolarization (Fig. 4A). Surely if the H+ pump were activated to counter intracellular acidification, the membrane should hyperpolarize!
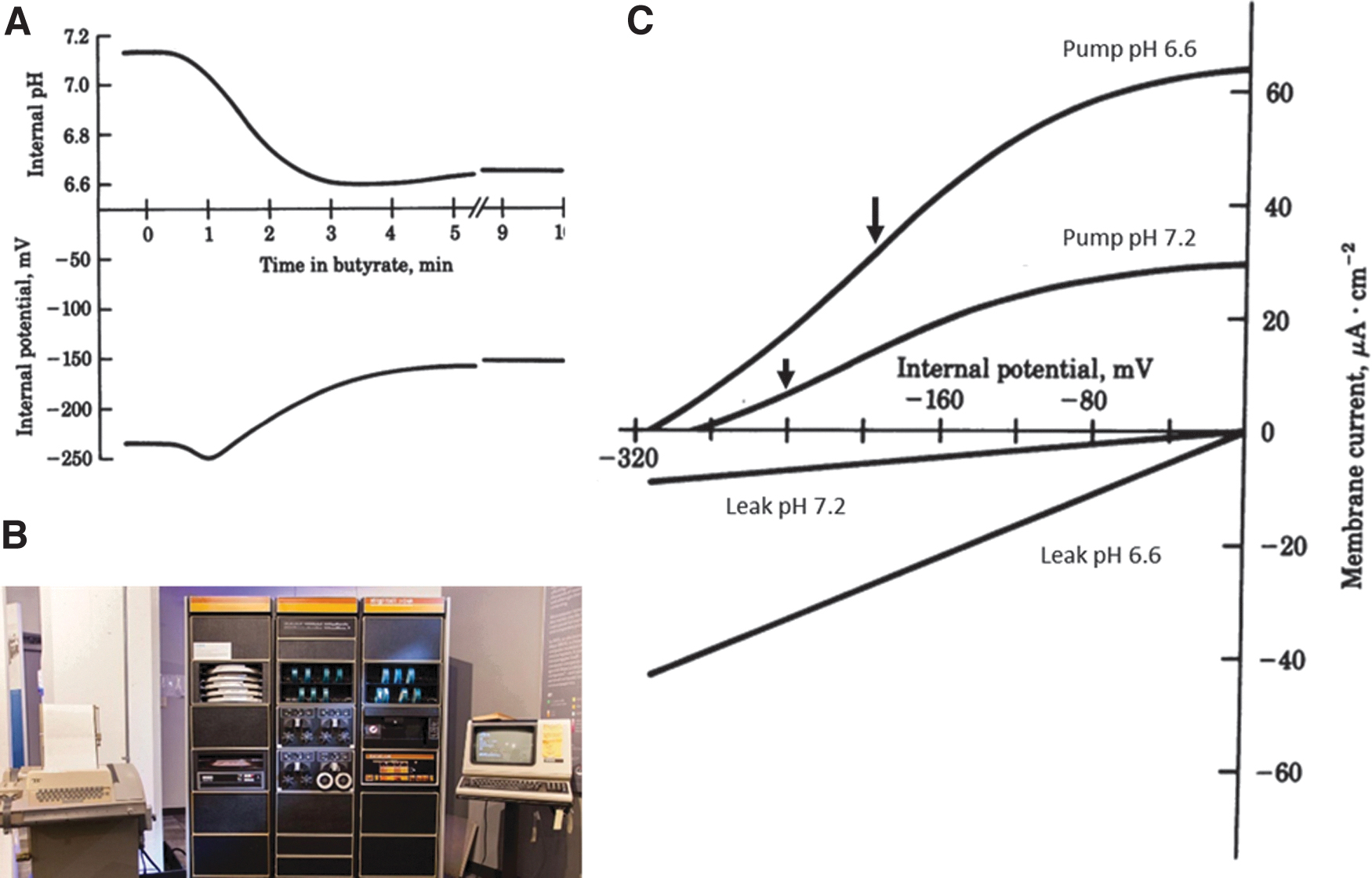
Critical role of the membrane potential in activating the plasma membrane H+ pump in Neurospora during acidosis.
Fortunately, a brilliant biophysicist—Ulf-Peter Hansen—was visiting the laboratory on sabbatical leave from Kiel University during 1980. Peter devised a model consisting of an enzyme-like H+ pump operating in parallel with a linear “leak” (representing the ensemble of other electrically active pathways), which enabled dissection and interpretation of membrane current–voltage (I–V) relationships during acidosis. The I–V relationship was generated by, and recorded on, a PDP8 microcomputer (Fig. 4B). [The computer was about 1.5 m tall and the 10 Mb recording disks about 30 cm diameter, requiring two hands to load. Such was the power consumption of the PDP8 that, even when experiments were not in progress Peter and I would turn the computer on to warm the small laboratory on cold New England days!]
Our findings were intriguing. 16 As pH declined from the control value of about 7.2 to 6.6, the pump was indeed activated by a factor of about 2 (Fig. 4C). However, any effect of pump activation on membrane potential was obscured by a disproportionate increase in “leak” current that depolarized the membrane and in so doing activated the pump still further. Thus, the H+ pump plays a role in controlling cytoplasmic pH not only through kinetic activation by protons but more importantly through its potential dependence as inherently depolarizing conductances are opened.
Calcium Signaling and Photosynthesis
In 1983, I moved from my postdoc position at Yale back to the University of York as a lecturer. As I focused on clearing a backlog of articles for submission and writing undergraduate lectures and grant applications, my days of hands-on experimentation drew to a close. Indeed, I was very fortunate in gaining considerable grant funding for my laboratory and this in turn attracted a series of talented postdocs who performed all of the electrophysiological experiments described in the remainder of this article.
Before leaving Yale, I had become intrigued by the notion that plants might utilize Ca2+ as a signaling ion in the cytoplasm analogously to the then emerging landscape in animal cells. 17 Dynamic changes in cytoplasmic free Ca2+ related to a physiological function had simply not been demonstrated in plants, and I reasoned that if such changes occur, then it should be possible to measure them electrophysiologically with the recently developed liquid membrane-based sensors. 18 Tony Miller (now Professor at the John Innes Centre) joined my laboratory to attempt to do just this, but there were major technical hurdles to overcome.
First, how could turgor-driven forcing of the liquid sensor back into the glass pipette be prevented? Tony devised a method of stabilizing the sensor with poly(vinyl chloride) to overcome this problem. A second challenge was that the cytoplasm of most land plant cells is restricted to a narrow band next to the plasma membrane, with the major fraction of cell volume being occupied by the vacuole into which the microelectrode is normally unavoidably inserted. We decided to return to the giant cells of charophyte algae (H+-Coupled Ion Transport in Plants section), which have a manageable depth of cytoplasm (about 10 μm), despite the presence of a large vacuole.
After many patient attempts with this challenging system, Tony finally began to obtain results, and they were dramatic. Light/dark transitions evoked a sustained but reversible change in free Ca2+ from ∼200 nM (light) to ∼350 nM (dark: Fig. 5). 19 The light intensity dependence of the Ca2+ response mirrored that of carbon fixation and was blocked by the photosynthetic electron transport inhibitor N′-(3,4-dichlorophenyl)-N,N-dimethylurea. We argued that light-induced Ca2+ depletion must represent Ca2+ uptake by the chloroplasts and could represent a signal for activation of sucrose synthesis, which occurs in the cytoplasm when reducing equivalents are being produced within the chloroplast. A subsequent collaboration with Mark Stitt (then at University of Bayreuth) produced biochemical evidence for this suggestion. 20

Elevation of cytosolic free Ca2+ in an internodal cell of a charophyte alga induced by darkness. Inset. Calibration data for the Ca2+-selective electrode before (○) and after (●) impalement. Reproduced with permission. 19
Energizing Plant Plasma Membrane Potassium Transport with H+ (and Na+)
My interest during PhD studies in how transport is energized and controlled at the plasma membrane of plants had not been forgotten. I was keen to apply my thinking to “higher plants” (angiosperms), and in the late 1980s wrote a grant application to the-then Agricultural and Food Research Council proposing to elucidate the mechanism of K+ transport in plants. Classic work by Manny Epstein at UC Davis had shown for cereal species that K+, similar to many other elements, exhibits biphasic kinetics for uptake with two distinct Kms.21,22 I argued in my grant application that we were now able to explore the mechanistic basis of biphasic uptake using electrophysiological approaches.
The species on which I proposed to do experiments was Arabidopsis thaliana—emerging at the time as the plant model for genetic studies. One reviewer of my grant application argued that I was jumping on an Arabidopsis bandwagon and that given the small size of the plant, the cells would not render themselves for electrophysiological studies. Fortunately, these spurious comments were ignored and funding was awarded. This grant enabled recruitment of Frans Maathuis (now a Professor at York), a highly talented electrophysiologist who drove a lot of the subsequent experimental design.
A first step in the investigation of K+ transport into roots was to define the energetic background. Using double-barreled K+-selective microelectrodes, we established through simultaneous measurement of cytoplasmic K+ activity and membrane potential that at micromolar concentrations of K+ externally transport must be energized, whereas above 1 mM passive transport would be possible. 23 We speculated that the high affinity transport mechanism could be effected through H+ cotransport in a 1:1 ratio with K+ and that transport at higher concentrations might then occur through K+ channels.
We investigated the hypothesis that high affinity K+ transport might by energized through H+ coupling using whole cell patch clamp in isolated Arabidopsis root cells after cell wall digestion. 24 Inward currents induced by low concentrations of K+ (Fig. 6A) exhibited a Km = 30 μM (in the range of reported values for whole-plant K+ uptake) and had reversal potentials consistent with our 1H+:1K+ hypothesis (Fig. 6B). Simultaneously with our own work, Daniel Schachtman and Julian Schroeder (UC San Diego) provided important genetic background by isolating a cDNA from wheat that, when heterologously expressed, exhibited properties consistent with H+ and K+ cotransport and which they named HKT1 (for High affinity K Transporter.) 25

Evidence for H+:K+ cotransport into root cells of Arabidopsis thaliana.
I mentioned in the H+-Coupled Ion Transport in Plants section that, although it is now clear that energized uptake of inorganic solutes in plants is normally energized through H+-coupling, exceptions exist. Sodium-dependent inward K+ currents had already been demonstrated in the giant cells of Chara. 26 The Chara genus is alkalophilic, and following a conversation with Alan Walker (then of Sydney University; now sadly deceased), we decided to investigate whether the Na+ dependence is apparent in a more acidophilic charophyte, Nitella translucens, where low environmental proton activity would not compromise H+ energized solute uptake.
Alan visited my laboratory on sabbatical leave, and we discovered significant enhancement of inward currents in the presence of Na+, not only when cells were presented with low concentrations of K+ (Fig. 7) but also with urea and lysine (data not shown). 27 Alan was keen to determine whether this Na+ dependence of K+ inward currents was more widespread among other plants, and in collaboration with Frans Maathuis we found electrophysiological evidence for Na+-coupled K+ transport in aquatic, but not terrestrial, angiosperms. 28 [It should be noted that work on HKT1 subsequent to the original publication 25 has convincingly revealed a capacity in some members of the HKT family to transport Na+, largely in the context of Na+ distribution within the plant.29,30]

Alkali cation-dependent plasma membrane K+ currents in the charophtye Nitella translucens. Potassium was added to alkali cation-free artificial pond water at 20 μM, or in the presence of Na+ or Li+ as shown. Reproduced with permission. 27
Patch Clamp Electrophysiology Opens the Door to Understanding Vacuolar Solute Transport
As all plant scientists know, the appearance of mature plant cells is dominated by a large central vacuole that normally occupies 80–90% of intracellular volume and can attain a diameter of tens of μm. The organelle is relatively structureless and serves—among other functions—as a repository for solutes. These solutes contribute to the turgor of plant cells and, depending on the elasticity of the cell wall, to their volume. The advent of improved membrane separation techniques in the 1980s enabled detailed biochemical characterization of the two dominant H+ pumps at the vacuolar membrane: the V-ATPase and the V-PPase that, respectively, hydrolyze either ATP or inorganic pyrophosphate and which acidify the vacuolar lumen. 31 Alongside biochemical approaches, the application of patch clamp electrophysiology to easily isolated whole vacuoles enables unparalleled insights into the properties of many organellar transporters and channels.
Even questions regarding vacuolar primary pumps can be answered with patch clamp. When the V-PPase was first identified biochemically, it was noted that hydrolytic activity displayed an obligatory dependence on K+. 32 Julia Davies (then a postdoc in the laboratory, and now Professor at Cambridge University) elegantly addressed the sidedness of this dependence in a simple decisive experiment: she measured inorganic pyrophosphate (PPi)-dependent currents in whole vacuoles in the presence or absence of K+ selectively on the cytoplasmic or luminal side of the membrane. The results (Fig. 8) showed unequivocally that the K+ dependence resided on the cytoplasmic side. 33

Membrane currents elicited by PPi in whole vacuoles from sugar beet roots in the presence of K+ selectively on the cytosolic or luminal side of the vacuole. PPi was added at 0.1 mM in the presence of 50 mM K+ or choline. Reproduced with permission. 33 PPi, inorganic pyrophosphate.
Julia went on to determine for both the V-PPase 34 and the V-ATPase 35 the I–V relationships of the two enzymes. In the latter case she was able to demonstrate from analysis of reversal potentials that the H+:ATP coupling ratio could be variable. This would enable the pump to generate considerable pH differences across the membrane in some conditions while maintaining greater energetic efficiency in conditions where a highly acidic lumen is not required.
The patch clamp technique is, of course, more conventionally used to study ion channel permeability and gating. Solute fluxes across the vacuolar membrane are critical in effecting changes in cell turgor. A number of laboratories were conducting pioneering studies on the selectivity and regulation of the vacuolar channels that might be involved in organellar solute exchange, and Gethyn Allen (now tragically deceased) joined the laboratory in 1992 to research in this burgeoning field. Gethyn performed extensive single-channel patch clamp studies on vacuoles from the guard cells that through changes in cell turgor control gas and water exchange through stomata in leaves. The aim was to describe the major pathways for Ca2+ release from vacuolar stores and the manner in which they might be regulated. In brief, our studies resulted in the characterization of two novel voltage-gated channels that potentially mediate Ca2+ release in these important cells, 36 as well as the elucidation of ion channel control by phosphorylation 37 and by cytosolic free Ca2+. 38
Calcium release from plant vacuolar membranes can be activated not only by voltage and cytosolic free Ca2+ but also by inositol 1,4,5-trisphosphate (InsP3). James Brosnan (now director of research at the Scottish Whisky Research Institute) had shown as a graduate student in the laboratory that this InsP3-induced Ca2+ release from plant vacuolar-enriched membrane vesicles was potently inhibited by nM concentrations of the mammalian InsP3 antagonist heparin. 39 Shelagh Muir (now global vice president R&D, Unilever), a graduate student in the laboratory, had just demonstrated that another Ca2+ channel agonist first identified in animal cells, cyclic ADP-ribose (cADPR), 40 also elicited Ca2+ release from vacuolar-enriched vesicles.
Gethyn, Shelagh, and I reasoned that if this release were truly from vacuoles, and not from a contaminating membrane fraction, then we should be able to detect InsP3- and cADPR-triggered currents from single intact vacuoles. Figure 9 shows that this indeed is the case. At low concentrations, both InsP3 and cADPR additively elicit 45 Ca2+ release from membrane vesicles as well as outward current from single intact vacuoles. 41 This finding was revealing not only in identifying one organelle from which Ca2+ can be mobilized, but also in demonstrating that both InsP3 and cADPR can mobilize Ca2+ from a single vacuole.

Additive release by InsP3 and cADPR of Ca2+ from red beet root vacuoles monitored radiometrically
Changes in cytosolic free Ca2+ are central to many signal transduction events in plants. 42 By the early years of the 21st century electrophysiological approaches in many labs, including my own (Fig. 10), had identified a variety of Ca2+-permeable ion channels that might be activated to generate elevation of free Ca2+, yet none of these channels had been characterized at a genetic level. However, some promising candidate genes had been identified based on homology to well-characterized channels in animals. One such gene in plants was TPC1, the sole member of the so-called Two Pore Channel (TPC) family of genes present in Arabidopsis. The TPC1 gene complements the phenotype of a yeast plasma membrane Ca2+ channel mutant and this and other experimental data led to a tentative conclusion that the TPC1 channel was located at the plasma membrane of plants. 43

At a patch clamp setup discussing results with a postdoc in the laboratory, Dr Chris Crotty (left), c. 2005.
I was intrigued by the possibility that the TPC1 channel might be involved in signaling events in plants, and Edgar Peiter (a postdoc in the laboratory who is now Professor at Martin Luther University, Halle) had been conducting experiments to test this idea. The experiments using tpc1 mutants were devised around my strong feeling that TPC1 was located at the plasma membrane, and were not working! At around this time, I was collaborating on a separate project with Frans Maathuis (now running his own laboratory at the University of York) on the proteomics of plant vacuolar and plasma membranes and was convinced that we had cross-contamination of membrane fractions because TPC1 was appearing consistently in the vacuolar membrane fraction.
At a conference in Montpellier in July 2004 I mentioned the problem to Julian Schroeder, who immediately suggested a solution: what if TPC1 was actually the Slow-activating Vacuolar (SV) channel that had first been reported in pioneering experiments as the dominant vacuolar channel 44 back in 1987? After all, the TPC1 channel was predicted to possess a cytoplasmic helix-loop-helix (EF hand) Ca2+-binding domain and SV currents are activated by cytoplasmic Ca2+; SV channels, although relatively nonselective among cations, will pass Ca2+ currents.
On my return to the laboratory, a roadblock on not one but two projects were instantly resolved thanks to Julian's comment. Frans patch clamped the tpc1 mutants that Edgar had obtained and back-crossed, and the dramatic result was that there was no SV current (Fig. 11C). And we could have confidence in the purity of our membrane preparations for the proteomics studies after all. 45 We went on to show visually that TPC1 is indeed located at the vacuolar membrane and that the channel is involved in seed germination and in control of stomatal aperture. 46

TPC1-dependent currents in vacuolar patches from Arabidopsis mesophyll cells. (
Besides the excitement of the biological findings, I took home a strong lesson from the informal discussion I had at the Montpellier conference: when at conferences, discuss the problems with your data as well as your achievements!
Closing Thoughts
I was once on an interview panel for an academic position in plant cell biology and remarked after one of the candidates had left the room: “They are producing some very interesting results but have very few papers”. One of my fellow panelists retorted “Yes, but not all science is like electrophysiology where you can obtain a few milliseconds-worth of data then publish a paper.” The retort was of course made largely in jest, but I hope I have shown in this retrospective that electrophysiology is a little more sophisticated than that.
Experimenting with plants and fungi has been fun. In their lifestyle, plants and fungi are quite different from most animal cells in being sessile if living on land and, therefore, needing to adapt to their environment at a cellular level in a way in which animal cells do not. This adaptation is reflected in the electrical properties of plant and fungal membranes—most notably the highly electronegative plasma membrane and the proton economy it runs.
Perhaps the most fun of all, however, has been to interact with stimulating and friendly colleagues—both within the laboratory and in other laboratories—to advance in just one small part of biology our knowledge of how life works.
Footnotes
Acknowledgments
I thank Dr. Zeenat Noordally and Professor Frans Maathuis for constructive comments on the article.
Author Disclosure Statement
No competing financial interests exist.
Funding Information
The author thanks the National Institutes of Health (USA) and the Biotechnology and Biological Sciences Research Council (UK) and its predecessor, the Agricultural and Food Research Council for financial support of the work reported in this paper.