
Abstract
Introduction:
Chronic stress causes a variety of physiological and behavioral alterations, including social impairments, altered endocrine function, and an increased risk for psychiatric disorders. Thereby, social stress is one of the most effective stressful stimuli among mammals and considered to be one of the major risk factors for the onset and progression of neuropsychiatric diseases. For analyzing the effects of social stress in mice, the resident/intruder paradigm of social defeat is a widely used model. Although the chronic social defeat stress model has been extensively studied, little is known about the effects of repeated or chronic social defeat stress on the endocannabinoid system (ECS). The present study aimed to understand the effects of chronic social stress on anxiety behavior and the levels of endocannabinoids (ECs) and two N-acylethanolamines (NAEs) in different brain regions of mice.
Materials and Methods:
Two-month-old, male C57Bl/6J mice were exposed to chronic psychosocial stress for 3 weeks. The effects of stress on anxiety behavior were measured using the light–dark box and hole board test. The EC levels of 2-arachidonoyl glycerol (2-AG) and anandamide (N-arachidonoylethanolamine [AEA]), as well as the levels of two NAEs (oleoylethanolamide [OEA] and palmitoylethanolamide), were analyzed by liquid chromatography–tandem mass spectrometry in the hippocampus, cerebellum, and cortex.
Results:
In comparison with control mice (n=12), mice exposed to social defeat stress (n=11) showed increased anxiety behaviors in the light–dark box and hole board test and gained significantly more weight during the experimental period. Additionally, chronic social stress induced differential alterations in the brain levels of 2-AG and AEA. More precisely, 2-AG levels were higher in the cortex and cerebellum, whereas reduced AEA levels were found in the hippocampus. Furthermore, we observed lower OEA levels in the hippocampus.
Conclusion:
The current study confirms that the ECS plays an essential role in stress responses, whereby its modulation seems to be brain region dependent.
Introduction
Social stress is considered to be a major risk factor for the onset and progression of common neuropsychiatric diseases, including mood and anxiety disorders.1,2 In humans, social defeat has been shown to have an increased incidence of depression, anxiety, post-traumatic stress disorder (PTSD), alcohol or drug abuse, and other aberrant physiological and behavioral symptoms.2–4
The resident/intruder paradigm of social defeat is a widely used model to investigate the behavioral and molecular consequences of social defeat stress in rodents. 5 In this paradigm, social defeat is initiated when a male intruder is introduced into the home cage of an older, aggressive, dominant resident male.6,7 Thus, this model is based on an initial occupation and establishment of a territory by a resident male that consequentially defends the territory against unfamiliar intruders. 8
The acute psychosocial stress model involves one fight on a single occasion only, while the chronic model allows fights at several consecutive occasions over days or weeks. 5 The intruder is usually protected by a wire cage from direct physical contact with the resident, but still exposed to visual, auditory, and odor cues of the resident, after having been defeated. Consequentially, the intruder is additionally exposed to chronic stress in the form of threat of attack. 5 It has previously been reported that social defeat causes a wide range of behavioral, physiological, hormonal, and molecular changes.6,9–11 Social stress alters neuroendocrine and neurobiological systems in several brain regions, including the hypothalamic–pituitary–adrenal axis (HPA), the primary neuroendocrine stress response system of mammals.10,12,13
Interestingly, although the specific mechanisms are still not entirely understood, the endocannabinoid system (ECS) is regarded as an important mediator of the interaction between HPA axis activity and stressful conditions. 13 The ECS is a retrograde, feedback messenger system that plays an essential homeostatic role in the central nervous system and regulation of emotional processes, including fear reactions and anxiety, and has also been associated with stress management.14,15
The effects of chronic stress exposure on the ECS seem to be brain region specific and spread over different system levels, that is, cannabinoid type 1 receptor (CB1R) expression and/or activity, degrading and synthesizing enzyme activity, and endocannabinoid (EC, i.e., N-arachidonoylethanolamine [AEA or anandamide] and 2-arachidonoyl glycerol [2-AG]) levels. 13
While different types of chronic stress models (such as social, restraint, unpredictable, and chronic mild stress) led consistently to decreased CB1R expression and/or activity in specific brain regions, contradictory effects have been reported on EC levels. 13
While the chronic social defeat stress model has been extensively studied, little is known about the effects of repeated (7 or 10 consecutive days, respectively) or chronic (3 weeks) social defeat stress on the ECS.16–19
So far, it has been shown that repeated social defeat stress in mice affects the EC-mediated spike-timing plasticity in the nucleus accumbens that controls the vulnerability to anxiety-like behavior. 18 Another study observed a region-specific decrease of AEA (hypothalamus and hippocampus) and an increase of 2-AG levels (hypothalamus, hippocampus, and frontal cortex) that probably modulate the expression of cued fear memory and suggested an involvement of CB1R not only in the control of cued fear expression but also of unconditioned anxiety and increased sucrose preference in mice. 19
Tomas-Roig et al. investigated the effects of chronic social defeat stress in mice on lipid concentrations in the cerebellum only and reported higher 2-AG and unaltered levels of AEA and related N-acylethanolamines (NAEs—palmitoylethanolamide [PEA] and oleoylethanolamide [OEA]) as well as reduced CB1R mRNA and protein expression.
In this study, we aimed to further understand the neurobiological mechanism by which stress is translated into psychopathology such as anxiety behavior. Therefore, we measured the effects of chronic social defeat stress on anxiety behavior in the light–dark box and hole board test and analyzed the EC and related NAE levels in three different brain regions.
Materials and Methods
Animals
Wild-type, male C57Bl6/J (n=23) and FVB/N (n=28) mice used in this study were obtained from Charles River Laboratories (Sulzfeld, Germany). All animals were housed individually under standard conditions (12-h inversed light/dark cycle; 21°C±2°C). Water and food were available ad libitum. After a 1-week habituation period, eleven 2-month-old C57Bl6/J mice were subjected to the resident–intruder paradigm procedure. Thereby, 1-year-old, male FVB/N mice served as residents as these mice display a higher level of offensive behavior in the resident–intruder paradigm than the C57Bl/6 strain. 20 The FVB/N colony was housed in a separate room to prevent olfactory habituation to C57Bl/6J intruder mice.
All animals were handled according to the German guidelines for animal care. Animal experiments were approved by the local animal protection authorities (Niedersächisches Landesamt für Verbraucherschutz und Lebensmittelsicherheit).
Experimental design
Two-month-old, male C57Bl/6J mice (n=11) were exposed to chronic psychosocial stress for 3 weeks. After 3 weeks of daily social defeat, stressed mice as well as control mice (n=12) were subjected to behavioral tests (Fig. 1A). All behavioral experiments were performed during the dark phase (active phase of nocturnal mice) between 8 a.m. and 8 p.m. Furthermore, weight gain of mice was monitored as part of a general physical assessment.

Chronic stress influences body weight.
Mice were sacrificed by cervical dislocation after the behavior tests. Cerebellum, cortex, and hippocampus were rapidly dissected, frozen on dry ice, and stored at −80°C until mass spectroscopy analysis.
Social stress procedure
The chronic social defeat stress model was based on a variant of the resident-intruder paradigm first described in the 1990s and further modified about 10 years later.9,11,21,22 As mentioned above, this common variant involves two phases—physical attack and threat of attack. The resident–intruder paradigm minimizes wounding from physical attack and involves a stronger emotional stress component. The paradigm was performed as previously described.16,23
In brief, an intruder mouse (C57Bl/6J, n=11) was placed into the home cage of a resident mouse (FVB/N, n=28). Mice were allowed to interact freely until the first signs of aggression or fighting behavior. After first signs of aggression, the intruder mouse was placed in a small wire-mesh cage within the resident's cage to protect it from direct attacks and potential physical injuries. Usually, the attack occurred within the first 2 min since only aggressive residents were chosen for the experiment. The nature of the stress paradigm should not be confused with the stress of severe physical injury. Therefore, residents that showed signs of violence that may result in serious physical attacks were excluded from the experiment.
Due to the wire-mesh cage, the intruder mouse remained in auditory, olfactory, and visual contact with the resident. This period of threat after the actual defeat is known to be stressful.24,25 After 1 h, the intruder mouse was placed back into its home cage. Experimental animals (C57Bl/6J, n=11) were subjected to the described social stress daily for 3 weeks. Each day, intruders were confronted with different residents to avoid habituation and ensure similar defeat intensity between stressed animals. Furthermore, the resident–intruder protocol was performed daily at similar time of the day to enhance the stress factor induced by the prediction of social defeat.
Aside from social defeat, the control animals (C57Bl/6J, n=12) were exposed to the same procedures regarding handling and a new environment (novel cage, but no intruder) as the socially stressed animals.
Light–dark box
A light–dark box with two equally divided compartments (26×13×28 cm) was used to assess anxiety-like behaviors. The light–dark box test was performed 24 h before the hole board test. It has been shown that previous testing experience in other anxiety and behavioral tests does not appear to alter behavioral performance in the light–dark box.26–28 The light–dark box test is based on the conflict between the innate aversion to brightly illuminated areas and the spontaneous exploratory activity of rodents. 29 The two chambers were connected by a 10×10-cm doorway in the center of the separating wall to allow free access to the adjacent chamber. Animals (n=11–12) were introduced to the dark chamber facing away from the door. Mice were allowed to explore the light–dark box for 10 min, whereby the first 5 min were regarded as the habituation period. ActiMot video tracking software (TSE, Bad Homburg, Germany) was used to record the time spent in the light and dark chambers as well as the distance traveled and the resting time.
Hole board test
The hole board test was used to measure exploratory behavior in an unfamiliar environment.30,31 Thereby, an altered head-dipping activity can be interpreted as a change in the anxiety state of an animal. 30 Mice (n=11–12) were placed in the center of a transparent hole board chamber (50×50×36 cm), with a nontransparent floor raised 4 cm above the bottom of the chamber that contains 16 equally spaced holes (2.2 cm diameter). Mice were allowed to explore the chamber freely for 5 min. ActiMot software (TSE) was used to record the number of holes explored (head dips).
Lipid extraction from mouse brain samples
ECs (AEA and 2-AG) and two structurally related NAEs (OEA and PEA) were extracted from the cerebellum, frontal cortex, and hippocampus, as previously described. 32 In brief, brain tissues (n=11) were homogenized with methanol containing the following internal standards: [2H4] AEA, [2H8] 2-AG, [2H2] OEA, and [2H4] PEA (Cayman Chemicals, Ann Arbor, MI). Lipids were extracted with methanol–chloroform (2/1 vol/vol). The recovered chloroform phase was evaporated to dryness under a stream of N2 and reconstituted in chloroform. Subsequently, lipids were fractionated by open-bed silica gel chromatography eluted with chloroform–methanol (9/1 vol/vol). The organic phase was concentrated again before liquid chromatography–tandem mass spectrometry (LC-MS-MS) analysis.
LC-MS/MS analysis
AEA, 2-AG, OEA, and PEA were measured by isotope dilution LC-MS-MS 33 performed with an Agilent® 1200 HPLC system (Agilent Technologies, Inc. Headquarters, Santa Clara, CA) coupled to an API 5000 triple quadrupole mass spectrometer (Applied Biosystems®, Foster City, CA), operating in positive electrospray ionization mode. Samples were injected into a 4-μm Synergi Hydro-RP C18 column (150×2 mm; Phenomenex®, Aschaffenburg, Germany) and eluted using a gradient of methanol in water at a flow rate of 0.5 mL/min. Protonated adducts of molecular ions of AEA, 2-AG, OEA, and PEA were quantified in the selected ion monitoring mode. Spectra were acquired at 4.34 sec/cycle and collected with Applied Biosystems Analyst software.
For LC-MS/MS data analysis, the ratio of the peak areas of analyte/internal standard was used to interpolate the tissue concentration of the lipid of interest. This isotope dilution method has the advantage that possible losses of these low-abundance compounds during the extraction do not affect their quantification. Values are expressed as pmol per g tissue.
Statistical analyses
Behavioral and weight differences between groups were tested using unpaired t-tests or two-way analysis of variance (ANOVA), followed by Bonferroni multiple comparisons, as indicated. As for lipid levels, the assumption of normal distribution was violated, Mann–Whitney tests were used for statistical analysis. All data are given as mean±standard error of the mean. Significance levels are given as follows: ***p<0.001, **p<0.01, and *p<0.05. All statistical analyses were done using GraphPad Prism, version 6.07, for Windows (GraphPad Software, San Diego, CA).
Results
Stressed mice show increased body weight
The body weight was measured before and after the social stress experiment at 8 and 11 weeks, respectively. Both control and stressed mice significantly gained weight during the three experimental weeks [Fig. 1B; two-way repeated ANOVA: age: F(1, 21)=58.97; p<0.001; Bonferroni correction: control: p<0.001; stress: p<0.001]. However, stressed mice gained significantly more weight compared with nonstressed control mice [Fig. 1B; two-way ANOVA: treatment: F(1, 21)=21.74; p=0.001; Bonferroni correction: 11 weeks control vs. stress: p<0.001].
Stressed mice show increased anxiety in the light–dark box
The light–dark test was used to evaluate the relative anxiety status of mice. During the habituation period, no significant differences were detected between stressed and nonstressed mice. However, after the 5-min habituation period, stressed mice spent significantly less time in the light compartment compared with control mice [Fig. 2A; two-way repeated ANOVA: treatment: F(1, 21)=15.64; p=0.0007; Bonferroni correction: after habitation: control vs. stress: p<0.001; control: habituation vs. test: p<0.001]. Both groups traveled an equivalent total distance in the light–dark box [Fig. 2B; unpaired t-test: t(21)=1.773; p=0.0907]. Furthermore, no significant difference in the total resting time was detected between stressed and control mice [Fig. 2C; unpaired t-test: t(21)=1.873; p=0.9378].
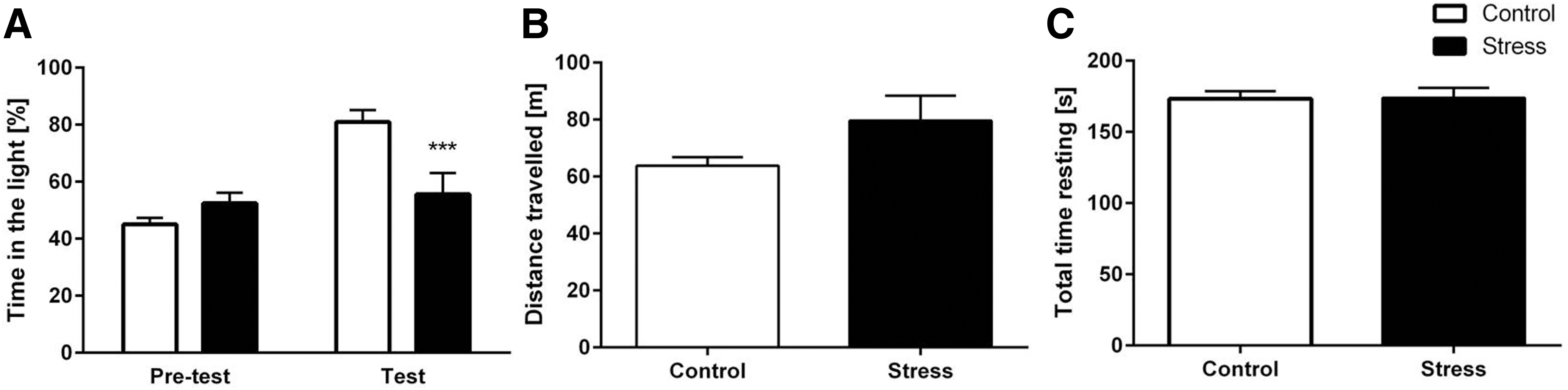
Chronic stress increases anxiety in the light–dark box.
Stressed mice show increased anxiety in the hole board test
To further explore possible changes in the anxiety behavior of socially stressed mice in a new environment, the hole board test was performed (Fig. 3A). Stressed mice showed an increased anxiety behavior in the hole board test [Fig. 3B, two-way ANOVA: treatment: F(1, 21)=3.203; p=0.0291]. Nonstressed control mice showed no preference for a specific area and explored all holes of the board equally. In contrast, stressed mice visited the corner holes significantly more often than the holes at other locations [Fig. 3B, two-way ANOVA: corners: F(3, 63)=9.406; interaction corners×treatment: F(3, 63)=3.694; Bonferroni correction: control: corner vs. back wall: p<0.05; corner vs. front wall: p<0.001; corner vs. center: p<0.001]. The altered exploration was not due to a decrease in overall explorative behavior as stressed and nonstressed mice showed a similar number of total hole visits [Fig. 3C; unpaired t-test: t(21)=0.5989; p=0.5557].
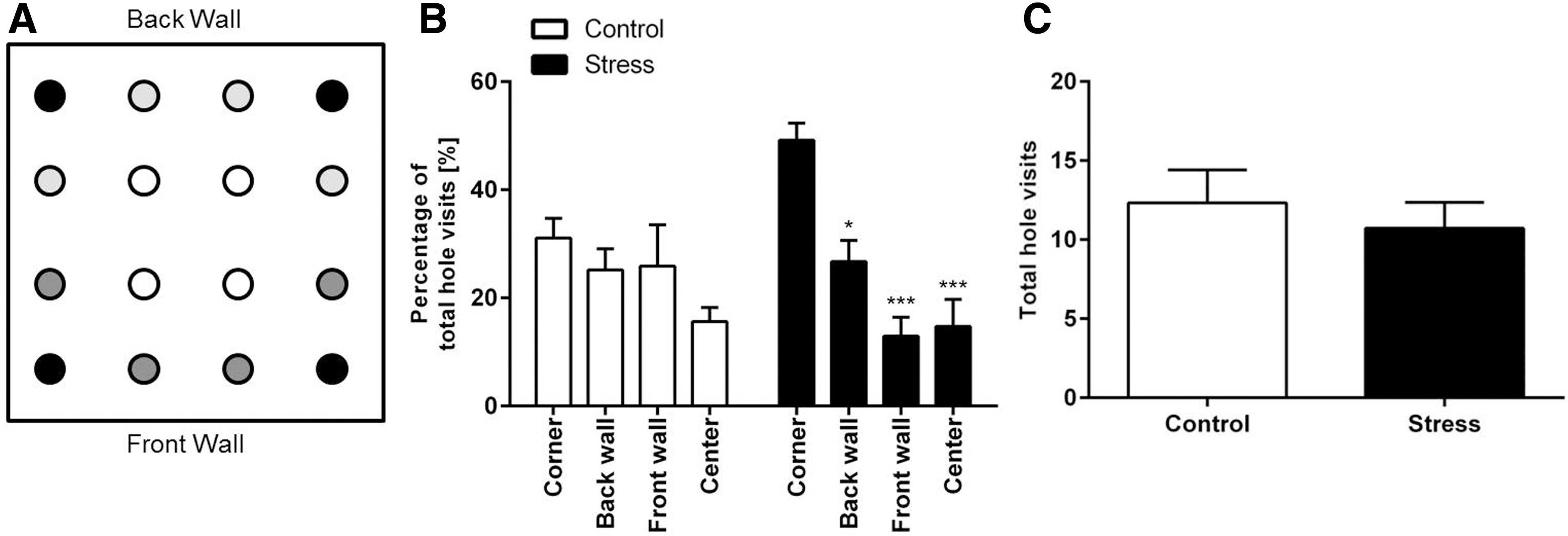
Chronic stress increases anxiety in the hole board test.
Altered EC levels in stressed mice
LC-MS/MS was used to quantify the two most common ECs, namely 2-AG and AEA, and two NAEs, OEA and PEA, in the hippocampus, cortex, and cerebellum of chronically stressed mice (Fig. 4–6).

EC concentrations in the cerebellum.

EC concentrations in the cortex.

EC concentrations in the hippocampus. Social stress led to decreased levels of
Stressed mice showed a higher concentration of the EC 2-AG in the cerebellum and cortex compared with control mice (Mann–Whitney test; cerebellum: p=0.0434; frontal cortex: p=0.0433). In contrast, AEA, OEA, and PEA levels in the cortex and cerebellum of stressed mice were not significantly different from nonstressed mice.
Furthermore, stressed mice displayed lower levels of AEA and OEA in the hippocampus compared with the levels of nonstressed control mice (Mann–Whitney test; AEA hippocampus: p=0.0068; OEA hippocampus: p=0.0294). However, social stress did not affect 2-AG and PEA levels in the hippocampus.
Discussion
Chronic stress causes a wide variety of physiological and behavioral alterations, including social impairments, altered endocrine function, and an increased risk for a variety of psychiatric disorders. Thereby, social stress (e.g., bullying, loss of social status, and changes of social roles) is one of the most potent stressful stimuli among mammals.2,34,35 For analyzing social stress in mice, the chronic social defeat paradigm is a widely used model. The induced stress causes several physiological changes from alterations in immune organs and endocrine function, elevations in corticosterone, and body weight changes, as well as behavioral and social modifications.36–39
Chronic social stress influences body weight
It has previously been shown that chronic stress alters body weight.38,40 Interestingly, whereas most studies using chronic social defeat stress models reported a body weight reduction during the stress period,22,40–43 our stressed mice gained body weight over time. However, other groups also reported increased or unaltered body weight after social stress.38,44,45 In line with our findings, Razzoli et al. observed that defeated mice showed increased body weight after 10 min of daily repeated social defeat over 10 days. 46 Similarly, Patterson et al. demonstrated that chronic exposure to social stress is associated with an increased body weight gain and caloric intake in male C57BL mice. 47
The effects of stress on body weight might also depend on the mouse strain analyzed. Savignac et al. stated an increased total body weight gain in C57BL/6 mice, but not in BALB/c mice, after repeated daily social stress. 48
These inconsistent results suggest that stress-induced body weight fluctuations are complex regulatory processes specific to the duration, nature, and degree of the stress applied. This has also been observed in humans. Clinically, patients with major depressive disorder can be divided into different subtypes, including atypical and melancholic depression. While patients with melancholic depression show reduced appetite and weight loss, patients with atypical depression report increased appetite and weight gain.49,50
We did not measure the daily food intake of the mice; hence, we cannot conclude that the weight gain was due to increased food intake and/or other factors. Interestingly, OEA released by small intestinal enterocytes during absorption of dietary fat is a satiety signal that inhibits eating and controls fat metabolism and energy expenditure, mainly through activation of type-α peroxisome proliferator-activated receptor (PPAR-α). 51 The PPAR-α-mediated signal is relayed to nuclei of the hypothalamus, and it has been suggested that the release of oxytocin in the hypothalamus and/or other regions of the brain may be a key effector of OEA-induced satiety. 52
As we have observed a significant reduction of OEA in the hippocampus, it may be possible that chronic stress also induces a reduction of gut-derived OEA, resulting in a reduced satiety signal and an increase in food intake. However, we did not measure peripheral OEA levels, nor EC and NAE levels, in the hypothalamus (the key brain region responsible for control of appetite and food intake), 53 thus further studies are needed to investigate chronic stress-induced weight gain and the underlying mechanisms more deeply.
Chronic social stress alters the ECS
The ECS is a ubiquitous, modulating neurotransmitter system involved in a variety of neurophysiological processes. Furthermore, the ECS is regarded as a central integrator of external and internal stimuli and corresponding neurophysiological and behavioral outcomes, 15 including anxiety and fear, as well as stress management. It has been shown that the ECS is very sensitive to stress, whereby the induced alterations are brain region specific and depend on the type and duration of applied stress.15,54
In the present work, the two best-studied ECs, 2-AG and AEA, showed opposite alterations in the brains of socially stressed mice in comparison with control mice. While levels of 2-AG were higher in the cortex and cerebellum, AEA levels were downregulated in the hippocampus. This finding is in line with previous reports showing a differential and brain region-specific regulation of 2-AG and AEA after repeated social16,19 or restrained stress.55–57 More precisely, decreased AEA levels have been found in the hypothalamus and hippocampus after 7 days of social stress, but not in the striatum and frontal cortex of C57Bl6/N mice. 19 Furthermore, elevated hippocampal, hypothalamic and frontocortical, 19 and cerebral 16 2-AG levels have been reported.
In contrast, we did not observe increased 2-AG levels in the hippocampus. It seems that the course of time of the applied stress needs to be considered and should be therefore systematically addressed in future studies. Similarly, repeated restraint stress induced a reduction of AEA levels in the amygdala,55–57 hypothalamus, 57 hippocampus, 57 and medial prefrontal cortex (mPFC),56,57 but led to an elevation of 2-AG levels in the mPFC, 56 forebrain, 55 amygdala,55–57 and hypothalamus. 57 Thus, the EC system seems to react to repeated social and restraint stress comparably.
Changes in EC level can be based on an altered transport, synthesis, or degradation enzyme activity. 2-AG and AEA are synthesized from phospholipid precursors through independent enzymatic cascades and degraded by monoglyceride lipase and fatty acid amide hydrolase (FAAH), respectively.55,58
Interestingly, an increased FAAH activity, and thus an elevated AEA degradation, has been reported after repeated restraint stress, 56 resulting in reduced AEA levels. As the FAAH enzyme also metabolizes the related NAEs, OEA and PEA, we expected a reduction not only in AEA levels but also in OEA and PEA levels. However, while Hill et al. found reduced OEA and PEA levels, we observed only reduced PEA levels in the hippocampus, while the OEA levels were unaffected. 59 This finding is in line with Rademacher et al. who also reported alterations in PEA levels only. 56 Therefore, an altered FAAH activity might not be the only contributing factor. In contrast, it has been suggested that increased 2-AG levels are mainly based on an enhanced 2-AG synthesis as no consistent general hydrolysis activity has been found in all brain regions. 56 However, the actual mechanism needs to be investigated in more detail in future studies.
Both ECs contribute to distinct forms of HPA axis adaptation.60,61 More precisely, it has been suggested that the steady-state reductions of AEA following repeated stress contribute to the basal hypersecretion of corticosterone, whereas the stress-specific increase in 2-AG signaling within the amygdala following repeated stress induces habituation of HPA axis activation.” 44
Interestingly, chronic administration of corticosterone dissolved in drinking water resulted in decreased CB1R binding site density in amygdala and hippocampus, as well as in reduced AEA levels, and increased FAAH activity in both brain areas. 62 In contrast, 2-AG levels were elevated in the hippocampus, but not in the amygdala. Furthermore, a cerebellar CB1R downregulation has also been observed in a chronic social stress model accompanied by elevated 2-AG levels. 16
Chronic social stress triggers increased anxiety
ECS had also been linked to anxiety.60,63–65 In this study, we showed that mice exposed to social defeat display increased anxiety behaviors in the light–dark box and hole board test. Defeated mice spent significantly less time in the light compartment of the light–dark box and, accordingly, more time in the dark area when compared with controls. The increased sensitivity to anxiety-like behavior in defeated mice has been previously reported. 66 In this study, this anxiety phenotype was further confirmed in the hole board test as the stressed mice explored the most secure holes more extensively in comparison with the more exposed holes.
Previous studies linked social defeat with decreased locomotion. 67 Such an activity reduction would unquestionably bias the interpretation of anxiety tests. However, our light–dark box and hole board test data demonstrated that social stress does not affect locomotion as both groups covered comparable total distances during the test, suggesting that the overall activity levels did not affect anxiety-like behaviors, or vice versa. Thus, the observed reduction in time spent in the light area of the light–dark box and the center of the hole board test was not just an artifact of decreased mobility, but instead reflects different exploratory patterns of stressed–defeated and nondefeated control mice. More specifically, the mice exposed to social defeat stress chose to spend more time in safer areas of the light–dark box and hole board test arenas (i.e., dark compartment or holes close to the wall, respectively) compared with the nonstressed mice.
Recently, behavioral consequences of reduced 2-AG brain levels have been studied in knockout mice lacking the diacylglycerol lipase α, an enzyme responsible for 2-AG synthesis. 2-AG levels were reduced by 80% to 90% in the hippocampus, cortex, amygdala, and striatum. 14 This substantial reduction of 2-AG concentration was accompanied by significantly decreased hippocampal, cortical, and amygdalar AEA levels. On a behavioral level, these knockout mice showed enhanced anxiety, stress, and fear responses. 14
Several studies also demonstrated the critical role of the ECS in stress and anxiety regulation in humans. Interestingly, lower AEA levels correlate with higher anxiety in healthy as well as mentally ill individuals.68,69 In healthy volunteers, an acute psychosocial stressor (Trier Social Stress Test) also induced a significant increase of serum 2-AG immediately after the offset of the stressor compared with baseline. In addition, serum OEA and PEA levels were significantly reduced during the stress recovery phase (30 min after the offset of the stressor). 59
Several pre-clinical studies also showed that stress inhibits AEA signaling.70,71 Therefore, it could be argued that a pharmacological intervention using ECs could be a promising strategy for treating anxiety disorders. However, a more recent study reported contradictory results. Compared with a control condition, acute psychosocial stress induced an increase of serum AEA and related NAE levels immediately after the stress period in healthy participants. 69 Interestingly, lower AEA baseline levels were correlated with higher anxiety ratings at baseline.
Furthermore, alterations in ECs and NAEs have been reported in PTSD patients, although the results were contradictive. Compared with healthy controls, not only both decreased serum AEA levels 72 and increased 2-AG, AEA, and OEA concentrations 73 but also solely elevated serum OEA levels 74 were observed. These data indicate that the ECS presumably plays a quite complex role in stress-related diseases and might be influenced by various parameters such as different environmental stimuli.
Conclusion
The current study confirms that the ECS plays a vital role in stress responses, whereby modulation of the ECS seems to be brain region dependent. The exact interrelationships are quite complex and therefore additional studies are needed to understand the various interactions during a stress response. In particular, the effect of the duration of stress and the underlying mechanism leading to altered EC and NAE levels in response to stress need to be studied in more detail.
Nevertheless, our findings support the idea that pharmacological alterations of ECS signaling might be a promising approach for treating mood and anxiety disorders.75,76 However, further studies are required to determine both, the validity of EC levels as a potential biomarker for anxiety states and the efficacy of new drugs targeting the ECS for treatment of stress- and anxiety-related disorders.
Footnotes
Acknowledgment
The authors acknowledge support by the Open Access Publication Funds of the Georg-August-University Göttingen University.
Author Disclosure Statement
No competing financial interests exist. The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Funding Information
This work was supported by the DFG Research Center for Nanoscale Microscopy and Molecular Physiology of the Brain (CNMPB). The project received support by intramural funding of CIMH.