
Abstract
Introduction:
Relatively little is known about the molecular pathways influenced by cannabis use in humans. We used a multi-omics approach to examine protein, metabolomic, and lipid markers in plasma differentiating between cannabis users and nonusers to understand markers associated with cannabis use.
Methods:
Eight discordant twin pairs and four concordant twin pairs for cannabis use completed a blood draw, urine and plasma toxicology testing, and provided information about their past 30-day cannabis use and other substance use patterns. The 24 twins were all non-Hispanic whites. Sixty-six percent were female. Median age was 30 years. Fifteen participants reported that they had used cannabis in the last 30 days, including eight participants that used every day or almost every day (29–30 of 30 days). Of these 15 participants, plasma 11-nor-9-carboxy-Δ9-tetrahydrocannabinol (THC-COOH) and total tetrahydrocannabinol (THC) concentrations were detectable in 12 participants. Among the eight “heavy users” the amount of total THC (sum of THC and its metabolites) and plasma THC-COOH concentrations varied widely, with ranges of 13.1–1713 ng/mL and 2.7–284 ng/mL, respectively. A validated liquid chromatography–tandem mass spectrometry (LC-MS/MS) assay measured plasma THC-COOH, THC, and other cannabinoids and metabolites. Plasma THC-COOH was used as the primary measure. Expression levels of 1305 proteins were measured using SOMAScan assay, and 34 lipid mediators and 314 metabolites were measured with LC-MS/MS. Analyses examined associations between markers and THC-COOH levels with and without taking genetic relatedness into account.
Results:
Thirteen proteins, three metabolites, and two lipids were identified as associated with THC-COOH levels. Myc proto-oncogene was identified as associated with THC-COOH levels in both molecular insight and potential marker analyses. Five pathways (interleukin-6 production, T lymphocyte regulation, apoptosis, kinase signaling pathways, and nuclear factor kappa-light-chain-enhancer of activated B cells) were linked with molecules identified in these analyses.
Conclusions:
THC-COOH levels are associated with immune system-related pathways. This study presents a feasible approach to identify additional molecular markers associated with THC-COOH levels.
Introduction
Over the past two decades 42 states, the District of Columbia, and U.S. territories Guam and Puerto Rico have legalized or decriminalized the use of cannabis for medical purposes, including 11 states and the District of Columbia, which have fully legalized recreational use of cannabis. 1 An increasing number of Americans use cannabis recreationally and for medicinal purposes.2,3 A recent National Academy of Sciences, Engineering, and Medicine review of cannabis's effects reported that cannabinoids affect a wide range of organ systems, including the cardiovascular, central nervous, gastrointestinal, immune, reproductive, respiratory, and visual systems.4,5
Controlled trials of cannabinoids and cannabinoid derivatives have shown preliminary efficacy as antiemetic, analgesic, and antiepileptic agents, as well as for appetite regulation, muscle spasticity, and intraocular pressure.6,7 Animal models have shown potential anti-inflammatory effects of cannabinoids, which may be efficacious for rheumatoid arthritis (RA) and inflammatory disease of the gastrointestinal tract.8,9 Conversely, cannabis use has been implicated in chronic bronchitis, metabolic syndromes, cardiovascular disease, abnormal brain development, psychiatric illnesses, and some cancers.5,10
When smoking Δ 9 -tetrahydrocannabinol (THC), the primary psychoactive component is rapidly absorbed from the lungs into the bloodstream. 11 THC is metabolized by oxidation to 11-hydroxy-Δ 9 -tetrahydrocannabinol (11-OH-THC, active metabolite) and 11-nor-9-carboxy-Δ 9 -tetrahydrocannabinol (THC-COOH, inactive metabolite).12,13 THC is preferentially stored in adipocytes and continues to passively diffuse into the blood over several days after discontinuation of cannabis use.14–16 THC-COOH levels increase slowly as released THC is metabolized, and THC-COOH levels remain stable and detectable over several days to weeks. 11
The physiologic effects of cannabinoids are mediated through the endocannabinoid system. Cannabinoid receptors CB1 and CB2, the primary targets of THC, are expressed in the central nervous system (CNS) and peripheral tissues.17,18 Other proposed cannabinoid receptors include the G-protein coupled receptor 55, 19 transient receptor potential vanilloid 1 ion channel, 20 and peroxisome proliferator-activated receptor α and γ. 21 While there is increasing interest in the therapeutic potential of pharmacologic targeting of cannabinoid receptors, 22 relatively little is known about the molecular pathways influenced by cannabis use. We utilized a multi-omics approach to identify plasma proteins, metabolites, and lipids associated with cannabis use in twin pairs discordant and concordant for cannabis use.
Materials and Methods
Participants
Participants consist of eight discordant twin pairs and four concordant twin pairs for cannabis use. Twins were enrolled in the Longitudinal Twin Sample of the Colorado Twin Registry, a population-based registry of monozygotic and dizygotic twins.23,24 Informed consents were obtained following institutional guidelines and according to the Declaration of Helsinki. All participants completed a blood draw, urine, and plasma toxicology testing and provided information about their past 30-day cannabis use and other substance use patterns.
Cannabinoid concentrations in ethylenediaminetetraaceticacid plasma
As previously described, a validated liquid chromatography–tandem mass spectrometry (LC-MS/MS) assay was used to quantify 11 cannabinoids and their metabolites in ethylenediaminetetraaceticacid plasma: THC, 11-OH-THC, THC-COOH, THC-glucuronide, cannabidiol (CBD), cannabinol, cannabigerol, cannabichromene, cannabidivarin, tetrahydro-cannabivarin (THCV), and THCV-COOH. 25 THC-COOH was measured directly, and samples were not pretreated with glucuronidases.
Proteomic analysis
Protein expression levels in plasma were measured using SOMAmersTM (Slow Off-rate Modified Aptamers) and the SomaScan platform (SomaLogic, Boulder, CO). SOMAmer reagents are made from single-stranded DNA that has been chemically modified at the five-position of pyrimidine residues to mimic amino acid side chains. We used the 1310 SOMAmers platform developed to bind to human proteins and assay their concentrations, with a median limit of detection (LOD) of 40 fM or 1.6 pg/mL, a median dynamic range of 8 logs, and a median coefficient of variation of 5% for individual proteins measured repeatedly in replicate runs of serum and plasma samples. 26
Lipidomic analysis
Bioactive lipid mediators were analyzed in plasma samples using a multianalyte high-performance liquid chromatography–tandem mass spectrometry (LC-MS/MS) assay. The assay analyzed 34 lipid mediators, including prostaglandins, leukotrienes, resolvins, and hydroxylated polyunsaturated fatty acid mediators (see Supplementary Data and Supplementary Table S1 for additional details). It is a modification of previously described methods that have been proven to deliver reliable, quantitative, and ultrasensitive results for the analysis of this group of small molecule lipid mediators.27–32
Metabolomic analysis
Plasma metabolomics profiles were assessed using a previously described semiquantitative targeted LC-MS/MS assay platform.33,34 The assay targets 314 small molecule metabolites covering all major metabolic pathways, including, but not limited to, glycolysis; the tricarboxylic acid cycle; the pentose phosphate pathway; and metabolism of amino acids, nucleotides, and so on. A list of the multiple reaction monitoring transitions for the metabolites monitored and their ionization modes is shown in Klepacki et al. 34
Statistical analysis
To examine differences in molecular phenotypes (i.e., protein, metabolite, and lipid levels) associated with THC exposure, we used two types of analyses: one to explore the association of differences in molecular phenotypes with differences in THC exposure within twin pairs (molecular insight analysis) and one to explore the association of molecular phenotypes with THC exposure (marker analysis).
The molecular insight analysis prioritizes differences in molecular phenotypes within twin pairs and, therefore, provides insight into the changes that occur within the body after accounting for genetic differences in the natural starting levels of these molecules. This type of analysis is useful when examining whether THC influences other molecules in the body. However, molecular phenotypes within this category may not be viable markers of THC exposure across individuals because between twin pair variation in the molecular phenotype may be much larger than the THC-associated variation in the molecular phenotype within twin pairs. Therefore, in marker analysis, we also assessed the association between levels of molecular phenotypes and levels of THC exposure without incorporating information about twin pairs.
The molecular insight analysis evaluated differences in protein, metabolite, or lipid levels in discordant twins that were not observed in concordant twins by including within the model the genetic relatedness of the twin pairs. Log-base 2 values for protein, metabolite, and lipids levels were each modeled independently using mixed model linear regression that included a general linear covariance structure to account for shared genetics between twin pairs (1 for monozygotic twins and 0.5 for dizygotic twins). Plasma THC-COOH levels were used as a fixed effect covariate, and the significance of the association between plasma THC-COOH levels and protein/metabolite/lipid levels was tested using t-statistics based on the estimated regression coefficient. The significance of the genetic influence on the protein/metabolite/lipid levels was assessed using a likelihood ratio test. The functional analyses were executed in SAS Studio (University Edition, Release 3.6; SAS Institute, Cary, NC).
The marker analysis excluded the shared genetics of the twin pairs and identified molecules that may have the predictive power to function as a marker for THC exposure. Log base 2 values for protein, metabolite, and lipid levels were used to predict plasma THC-COOH levels in a standard linear regression model. Pairing among twins was not included in the model. The marker analyses were executed in R Statistical Software (version 3.3.3).
Functional enrichment
Functional enrichment was examined for proteins that were associated with THC-COOH in both the mechanistic analysis and the marker analysis (nominal p-value <0.01). Enrichment of individual Gene ontology (GO) terms was determined using a Fisher's exact test executed in R and the GO annotation from the biomaRt package. 35 To account for the bias based on which proteins were targeted by the array, only proteins on the array were included in the background data set for the enrichment analysis. A false discovery rate (FDR) was used for multiple testing across terms. A GO term was significantly enriched if at least two proteins from the GO term were associated with THC-COOH, and the enrichment FDR was <0.10.
Results
Of the 24 participants, all were non-Hispanic Caucasian, and 66% were female (Table 1). The median age was 30 years. Of the 15 participants who reported that they had used cannabis in the last 30 days, 8 used daily or almost daily (29–30 of 30 days). Participants who did not endorse regular cannabis use reported on average using 0.125 days of the past 30, with undetectable urine THC levels (Table 1).
Clinical Characteristics
Days of use in the past 30 days (self-report).
DZ, dizygotic; MZ, monozygotic; THC, tetrahydrocannabinol; CBD, cannabidiol; THC-COOH, 11-nor-9-carboxy-Δ 9 -tetrahydrocannabinol.
THC-COOH is a comparatively stable inactive metabolite that is detectable for several days to weeks after last use. Among the eight “heavy” users, the amount of total THC (sum of THC and its metabolites) and THC-COOH in plasma varied greatly, with ranges of 13.1–1714 ng/mL and 2.7–284 ng/mL, respectively (Fig. 1). Of the 15 participants who reported that they had used cannabis in the last 30 days, 12 had detectable levels of THC-COOH and total THC. CBD was measurable in only one subject and, therefore, CBD levels were not included in any further analyses.
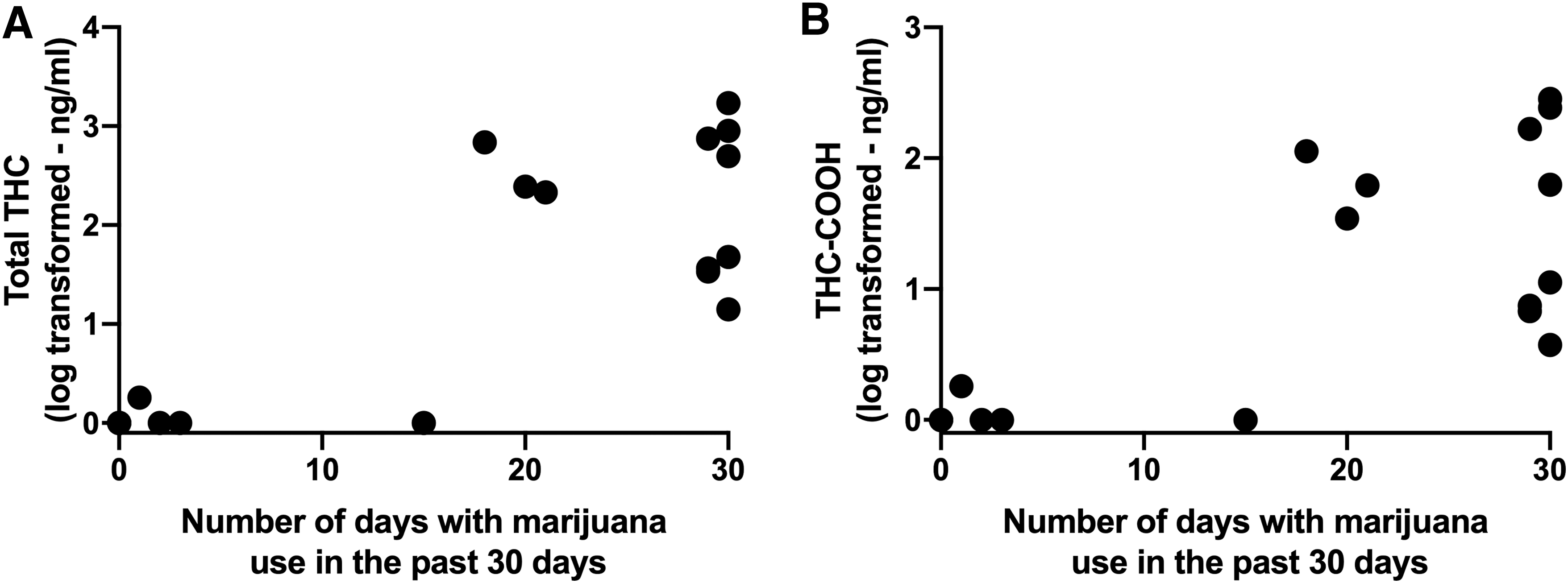
Association of THC and THC-COOH levels in plasma with self-report of number of days with cannabis use in the past 30 days. Each point represents a different subject.
Proteins, metabolites, and lipids associated with THC-COOH levels
Protein levels
In the original proteomics data set, 1310 probes were assessed. Five probes were later determined to be of poor quality by SomaLogic and four probes were identified as a “Rat protein” in the annotation and were therefore eliminated from further analyses. Because this array was specifically designed from human blood proteomics, expression levels were estimable for all proteins and all samples. The quantile–quantile plot (Supplementary Fig. S1) indicates that it is worthwhile to explore the top results in both analyses.
Using a significance threshold of 0.01 (nominal p-value), 19 proteins were associated with THC-COOH in the mechanistic insight analysis, and 22 proteins were associated with THC-COOH in the marker analysis (Fig. 2). Thirteen proteins were significant using both analysis approaches (Table 2). Only 3 of the 13 proteins indicated a negative association with THC-COOH levels: complement C3, clusterin, and proprotein convertase subtilisin/kexin type 7. Myc proto-oncogene protein (MYC) was the only protein that was statistically significant after multiple testing correction (FDR <0.05) in both analyses (Fig. 3A, B).

Identification of proteins, metabolites, and lipids that were associated with THC-COOH levels. Proteins (blue), metabolites (yellow), and lipids (green) were analyzed separate for both mechanistic insight (controlling for shared genetics between twins) and for marker potential (not accounting for shared genetics). All 24 subjects were included in all analyses.
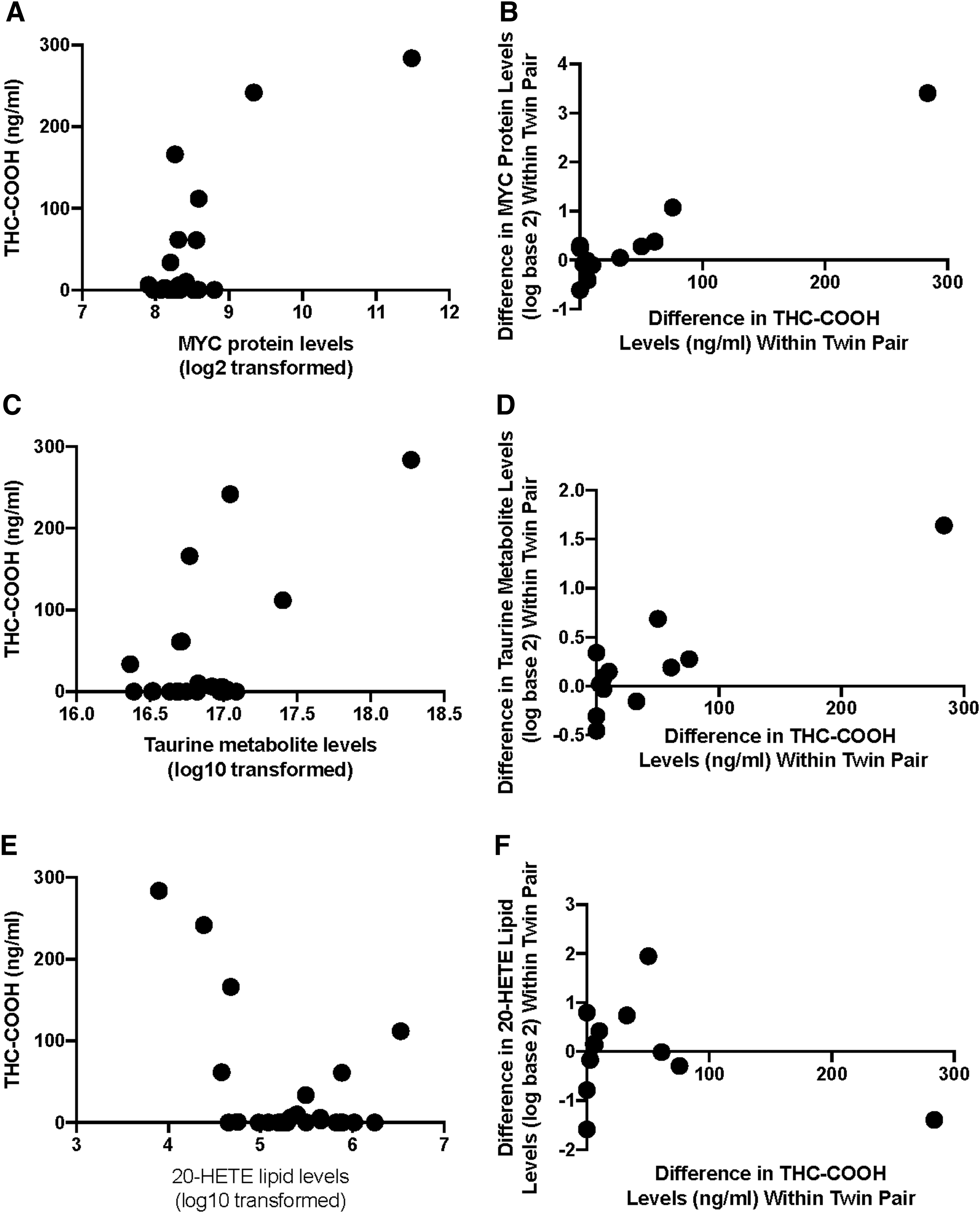
Demonstration of the association between proteins, metabolites, and lipids with THC-COOH levels.
Proteins Associated with 11-Nor-9-Carboxy-Δ 9 -Tetrahydrocannabinol Plasma Levels
JAK-STAT, Janus kinase-signal transducers and activators of transcription; JNK, c-Jun N-terminal kinase; FDR, false discovery rate.
In the marker potential analysis of MYC (Fig. 3A), a clear positive association was observed between Myc proto-oncogene protein expression levels in subjects with at least some detectable level of THC-COOH with the two subjects with extremely high levels of THC-COOH also having extremely high levels of Myc proto-oncogene protein. For the mechanistic insight analysis (Fig. 3B), the twin pairs that differed the most in THC-COOH levels also differed in their Myc proto-oncogene protein levels (e.g., the twin with higher THC-COOH levels also had higher Myc proto-oncogene protein levels).
Metabolite levels
Of the original 316 metabolites targeted, 296 were identified above the LOD in at least 25% of samples. Using a significance threshold of 0.01 (nominal p-value), 10 metabolites were associated with THC-COOH in the mechanistic analysis, and 8 metabolites were associated with THC-COOH in the marker analysis (Fig. 2). Of these, six metabolites were significant using both analysis approaches. Three of the six metabolites could be validated with further analyses (Table 3). Of the three, taurine had the most significant association in both the mechanistic analysis and the marker analysis. Plasma levels of taurine were positively associated with THC-COOH levels in the marker analysis (Fig. 3C), and the difference in taurine levels between twins was also positively associated with the difference in THC-COOH levels (Fig. 3D).
Validated Metabolites Associated (p<0.01) with 11-Nor-9-Carboxy-Δ 9 -Tetrahydrocannabinol Plasma Levels
Parameter estimate (standard error of parameter estimate).
Lipid levels
Of the original 34 lipids examined, all 34 were detected above quantitation limits in at least 25% of samples. Three lipids were associated with THC-COOH in the mechanistic analysis (nominal p-value <0.01), and two lipids were associated with THC-COOH in the marker analysis (nominal p-value <0.01). Both lipids that were significant in the marker analysis were also significant in the mechanistic analysis (Table 4). d12-PJ2 was the most significant lipid in both analyses. In the marker analysis, d12-PJ2 was below limits of quantitation in three of the four subjects with the highest levels of THC-COOH (Fig. 3E). In the mechanistic analysis, this same negative trend was observed (Fig. 3F).
Lipids Associated (p<0.01) with 11-Nor-9-Carboxy-Δ 9 -Tetrahydrocannabinol Plasma Levels
Parameter estimate (standard error of parameter estimate).
Functional enrichment
The 13 proteins that were significantly associated with THC-COOH levels were examined for enrichment of GO terms. Four GO terms were significantly enriched (FDR <0.10) for proteins associated with THC-COOH levels (Fig. 4). Six of the 13 proteins were included in cytokine-mediated signaling pathway (GO: 0019221): CD86, CX3CL1, IL19, IL1RAPL2, IL23R, and MYC, whereas different pairwise combinations of CLU, CX3CL1, and C3 were associated with the other three enriched terms: microglial cell proliferation (GO: 0061518), neuron remodeling (GO: 0016322), and positive regulation of receptor-mediated endocytosis (GO: 0048260).
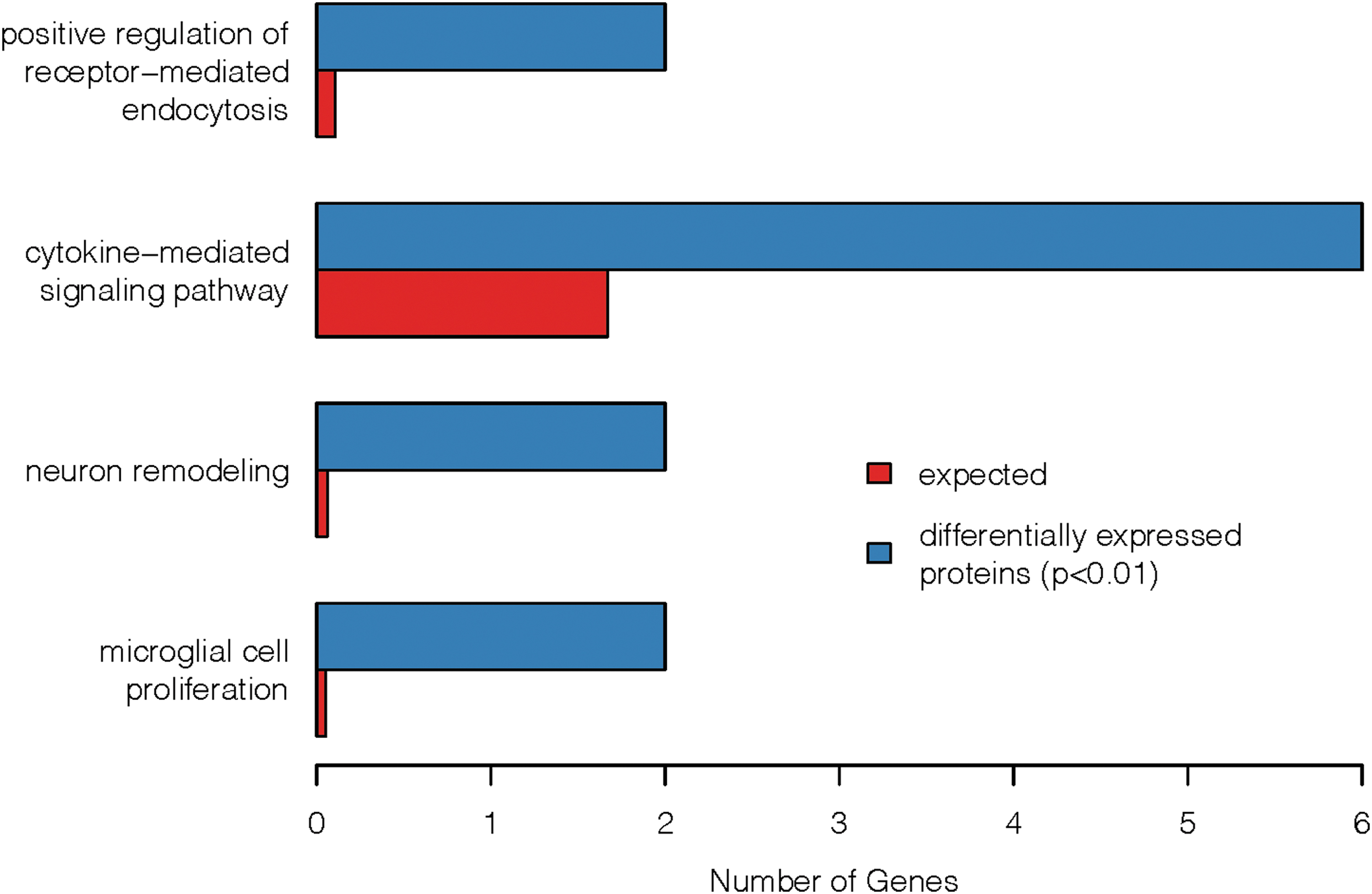
Functional enrichment of GO terms for proteins associated with THC-COOH levels. Included in the graphic are GO terms that were enriched (FDR <0.10) for proteins that were associated with THC-COOH (p<0.01) in both the analysis for mechanistic insight and the analysis for marker potential (13 genes). The blue bars represent the number of proteins that met the criteria for association with THC-COOH that were annotated to that particular GO term. The red bars represent the number of associated proteins that we would expect to be annotated to that GO term by chance. GO, gene ontology; FDR, false discovery rate.
Discussion
In this study, we identified 13 proteins, 3 metabolites, and 2 lipids that were significantly associated with THC-COOH levels using analyses that account for or ignore twin information to assess mechanistic insight or marker potential, respectively. A literature review of the significant molecules identified in this study revealed five pathways linked to these molecules (interleukin [IL]-6 production, T lymphocyte regulation, apoptosis, kinase signaling pathways, and nuclear factor kappa-light-chain-enhancer of activated B cells [NF-κB]) and is summarized in Table 5.
Molecular Pathways of Associated Protein, Metabolite, and Lipid Markers
A literature review of the significant molecules suggests that immunosuppressive effects of cannabis may be modulated through kinase signaling cascades, including MAPK and ERK1/236,37; transcription regulation by NF-κB 38 ; and apoptosis-mediated T lymphocyte regulation.39–41 Δ 9 -THC is a potent inducer of apoptosis leukemic cell lines, including mononuclear cells, 42 through p38 MAPK signaling activation and caspase-dependent apoptosis. 43 Similarly, treatment of macrophage cells with other THC compounds results in CB2 receptor-mediated activation of MAPK-induced apoptosis. One study has evaluated the effects of cannabis administration on p38 MAPK-ERK signaling in healthy subjects, which have been shown to be decreased and decrease overall phosphorylation in T lymphocytes, whereas chronic use may have a paradoxical effect. 44
Phosphorylation of ERK results in activation of transcription factors, including NF-κB, which was initially characterized as a lymphoid-specific transcription factor.45,46 NF-κB is a protein complex transcription factor that regulates gene expression fundamental to innate and adaptive immune responses.47,48 In T lymphocytes Δ 9 -THC results in decreased binding of NF-κB at CD40L promotor, suppressing T lymphocyte function. 38 Another potential mechanism of NF-κB immunomodulation is regulation of apoptosis. 49 In dendritic cells Δ 9 -THC has been shown to induce apoptosis through activation of NF-κB. 50
We also identified proteins significantly associated with THC-COOH levels that may provide further insight into THC-induced lymphocyte apoptosis, including fractalkine, 51 IL-19, 52 MYC, 53 and clusterin. 54 Fractalkine, encoded by CX3CL1, is a cytokine protein that mediates leukocyte differentiation, trafficking, and activation. 55 Expression of CX3CL1 has been shown to be regulated by cannabinoid receptor signaling. 56 Fractalkine induces pro-inflammatory response through activation of multiple pathways identified in Table 5, including MAPK-ERK and c-Jun N-terminal kinase (JNK) signaling cascades and NF-κB-regulated gene transcription.57,58 Through NF-κB-dependent gene expression fractalkine induces an antiapoptotic effect. 59 Fractalkine subsequently acts as a chemoattractant of macrophages when it is released from apoptotic lymphocytes. 51
In addition to fractalkine, prostaglandins activate MAPK-induced NF-κB gene expression and exhibit pro- and anti-inflammatory properties.60,61 Prostaglandin J2 family members, including delta-12PGJ2, are immunomodulatory, in part, through modulation of cell cycle gene expression. 62 15-d-PGJ2 induces production of IL-8 in activated T lymphocytes, whereas pretreatment with 15-d-PGJ2 triggers apoptosis in unstimulated T lymphocytes. In Burkitt lymphoma cells, 15-d-PGJ2 has been shown to suppress constitutive NF-κB activity, promoting tumorigenesis. 63 Delta-12-PGJ2, a related prostaglandin and associated with THC-COOH levels in the study presented herein, has also been shown to induce caspase-dependent apoptosis in human leukemic cells 64 and in eosinophils through inhibition of NF-κB signaling. 65
MAPK and NF-κB pathways may also modulate T lymphocyte differentiation and phenotype expression. IL-23R, the primary receptor of IL-23, forms a heterodimer receptor complex that induces signal transduction through p38 MAPK signaling cascade and NF-κB-dependent gene expression. 66 IL-23 is a member of the IL-6 type family of interleukins that is produced by monocytes. 67 IL-6 and IL-23 promote Th17 lymphocyte development, which increases neutrophil production and recruitment as part of the adaptive immunity response against pathogens.
In vascular endothelial cells 20-hydroxyeicosatetraenoic acid (20-HETE) is a vasoactive eicosanoid that regulates endothelial cell function and increases production of inflammatory cytokines through MAPK signaling cascades, including phosphorylation of p38 MAPK and ERK1/2 and increased nuclear translocation of NF-κB. 68 Although the mechanistic relationship with Δ 9 -THC is not well characterized, 20-HETE has also been shown to mediate p38 MAPK activation 69 and ERK phosphorylation. 70
Further elucidation of the biology of cannabis-induced immunosuppression may also provide insight into autoimmune diseases and new treatment targets. Liu et al. recently reviewed the role of fractalkine in inflammatory and autoimmune diseases, including amylotrophic lateral sclerosis, atherosclerosis, coronary artery disease, liver fibrosis, diabetes, inflammatory bowel disease, RA, and cancer. 71 In RA fractalkine may stimulate inflammation through an NF-κB-dependent pathway. 72 Dysregulation of NF-κB is also implicated in viral illnesses, autoimmune disease, and chronic inflammation,73–75 as well as cancers. 76
Cannabinoid biology may also provide treatment insight into multiple sclerosis (MS), an autoimmune disease of the CNS. In MS, fractalkine recruits CD4+ T-lymphocytes, increasing early inflammation in the CNS. 77 Th17 leukocytes subsequently promote chronic inflammation. 78 In mouse models of MS, Δ 9 -THC and CBD induce suppression of IL-6 and decrease Th17 activation. 78
In addition to fractalkine, IL-6, and Th17 leukocyte regulation, other systemic pathways identified herein may mediate cannabinoid activity in the CNS. In animal models, MAPK-ERK signaling in the brain is responsive to Δ 9 -THC, 79 which is proposed to be integral to neuronal plasticity and implicated in long-term adaptive responses to chronic Δ9-THC exposure.80–82
Interestingly, elevated NF-κB activity has also been associated with schizophrenia 83 and is one of several transcriptional targets implicated in maintenance of addiction in the nucleus accumbens. 84
In addition to identifying potential pathways influenced by cannabis administration, another goal of this study was to identify potential molecular targets that may indicate cannabis consumption. Currently, cannabis consumption is primarily identified through direct testing of THC or its metabolites. The Myc proto-oncogene protein was associated with THC-COOH levels in both mechanistic and potential marker analyses. As this was a cross-sectional study, it could not determine whether this protein may act as a marker of cannabis consumption after consumption has desisted, but this could be examined using longitudinal or treatment samples in a future project.
Limitations
The primary limitation of this study was the small sample size, which limits generalizability of this pilot study. The use of discordant twins partially mitigates this limitation, as twins share many unmeasured background variables (including genetics) that can confound association studies. 85 A larger study that includes detailed cannabis use history may identify the associated markers or a combination of these markers may identify cannabis use with more accuracy.
Footnotes
Acknowledgments
The authors acknowledge Sally Wadsworth, PhD, and Chandra Reynolds, PhD, for their support.
Author Disclosure Statement
No competing financial interests exist.
Funding Information
This project was also supported by National Institutes of Health grants K24DA032555, DA035804, DA042755, and AG04693.
Abbreviations Used
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.