
Abstract
Introduction:
Medicinal cannabis has proliferated around the world and there is increasing interest in the therapeutic potential of individual plant-derived cannabinoids (phytocannabinoids). Preclinical evidence suggests the phytocannabinoid cannabigerol (CBG) could be useful in treating brain disorders, including stress and anxiety-related disorders. In this study, we aimed to explore whether CBG disrupts various contextually conditioned fear memory processes and trauma-induced anxiety-related behavior in a mouse model of post-traumatic stress disorder (PTSD).
Materials and Methods:
All mice underwent contextual fear conditioning. CBG was administered between 1 and 60 mg/kg intraperitoneally (i.p.). We first assessed the effects of repeated CBG exposure on long-term fear memories. We also examined whether acute CBG affected various fear memory processes, namely expression, acquisition, consolidation, and reconsolidation of conditioned fear. Finally, the effect of acute CBG administration on stress-induced anxiety in the light/dark test was assessed.
Results:
Repeated CBG exposure did not affect long-term conditioned fear that was observed 24 days after the conditioning session. Moreover, acute CBG administration did not influence the acquisition, consolidation, reconsolidation, or expression of contextually conditioned fear. Acute CBG treatment also did not affect stress-induced anxiety-related behaviors in the light/dark test.
Conclusions:
CBG was ineffective in disrupting long-term fear memories, various conditioned fear memory processes, or stress-induced anxiety-related behavior in mice. These preclinical data suggest CBG may have limited scope in the treatment of PTSD and stress-related anxiety.
Introduction
Medicinal cannabis has proliferated rapidly around the world in the last decade and is frequently used to treat post-traumatic stress disorder (PTSD).1–4 PTSD is a complex mental disorder, arising from a chronic impairment of memory processes in response to a traumatic event. 5 Some PTSD patients choose to use medicinal cannabis products because conventional therapies have limited efficacy.6,7 Unlike conventional drugs that require rigorous preclinical and clinical testing before they are introduced to patients, medicinal cannabis products have a direct pathway to market and are being used in the absence of sufficient evidence. Thus, there is an urgent need for research to validate the uses of medicinal cannabis products, while also refuting spurious applications.
Cannabis contains approximately >100 phytocannabinoids, of which Δ 9 -tetrahydrocannabinol (THC) and cannabidiol (CBD) are the best characterized. 8 CBD has recently been approved by drug regulatory bodies to treat intractable childhood epilepsies, but it is also used to treat a myriad of medical conditions. The hype around CBD has led to speculation on which individual phytocannabinoid might be next exploited to treat medical conditions. One of these is cannabigerol (CBG).9,10 CBG is the decarboxylation product of cannabigerolic acid (CBGA), which is the major precursor to the enzymatic synthesis of various phytocannabinoids in the plant.11,12 Many medicinal cannabis growers are developing strains of cannabis with high CBG content, and CBG isolates can be purchased from numerous vendors. There are suggestions that these products can be used for therapeutic purposes.
Accumulating evidence suggests neuroinflammation may contribute to PTSD pathophysiology.13,14 In mouse models of PTSD, long-term persistent fear memories have been associated with the upregulation of various inflammatory-related genes, as well as long-term microglial cell abnormalities.15–17 Treatment with fluoxetine, a drug used to treat PTSD, reduced fear-related behavior, which was associated with a reduction in neuroinflammation in a mouse model of PTSD. 15 Moreover, depletion of microglial cells reduces PTSD-related behaviors in mice. 18 Preclinical research suggests CBG has robust anti-inflammatory effects across an array of disease models.19–21 Thus, our first aim in this study was to assess whether repeated exposure to CBG might similarly reduce long-term fear memories in mice.
Another compelling mechanism of action of CBG that is highly relevant to fear memory and PTSD is its ability to potently activate α2-adrenoreceptors (α2R). 22 α2R agonists such as clonidine and guanfacine have been suggested as potential treatments of PTSD,23,24 and α2R agonists disrupt acquisition, consolidation, and reconsolidation of conditioned fear in rodents.25–29 However, the effects of CBG on these forms of fear memory are unknown. There is some evidence to suggest that CBG may have anxiolytic effects. 30 Additional aims of this study were then to examine whether CBG disrupts various fear memory processes, as well as to observe whether CBG reduces stress-induced anxiety-related behavior in mice.
Materials and Methods
Animals and housing
Male C57BL/6 mice (Animal Resources Centre, Perth, Australia) 10–12 weeks of age were used in this study. All mice were housed in groups of 4–5 per cage and maintained on a reverse 12-h light/12-h dark cycle (lights on at 7 pm). The mice had ad libitum access to food and water, and environmental enrichment in the form of an igloo, a toilet roll, and small wooden block. All research and animal care procedures were approved by the University of Sydney Animal Ethics Committee and were in agreement with the Australian Code of Practice for the Care and Use of Animals for Scientific Purposes.
Mice were handled for 3 min for 3 consecutive days before experimentation. Mice were also habituated to a holding room for at least 1 h before all behavioral tests and were transferred to each procedural room in individual transport cages containing bedding.
Drugs
CBG (THCPharm, Germany, purity 99%) was administered at doses of 1, 3, 10, 30, and 60 mg/kg, and dissolved in a standard vehicle of ethanol, Kolliphor EL™, and 0.9% NaCl at a ratio of 1:1:18, respectively. Doses 1, 3, 10, and 30 mg/kg dissolved into a clear solution, and 60 mg/kg was a suspension solution. Mice were randomly allocated to experimental groups and administered the drug or vehicle through i.p. injection at a dosage of 10 mL/kg. For the acute studies, CBG was administered such that the Tmax (2 h) was during the time of testing. 31
Contextual fear conditioning
The mice underwent fear conditioning in commercially available chambers sourced from CleverSys, Inc.16,32–34 Chambers consisted of transparent Plexiglas walls, with a floor composed of stainless steel bars, which elicited an inescapable electric foot-shock. Each individual chamber was contained within its own sound attenuating isolation box. A video camera was positioned above the center of the box.
All experiments utilized contextual fear conditioning to establish a fear memory associated with the context. Mice were placed into the fear conditioning chamber and after 198 sec, a single inescapable electric foot shock was elicited (duration: 2 sec and intensity: 1.5 mA).15,32,35 Mice remained in the chamber for a further 60 sec, and then were returned to their home cages. All freezing data were retrieved during re-exposure sessions, where mice were placed into the fear conditioning chamber for 180 sec without application of foot shock. After each session, the chambers were cleaned using 70% ethanol.
Freezing behavior was used as an index of contextually conditioned fear memory. Freezing was defined as any time the mouse was vigilantly immobile, that is, with an absence of movement, excluding respiratory-related movements. 36 Manual scoring was performed, where freezing behavior was rated every 2 sec and determined to be either “freezing” or “not freezing” by an observer blind to experimental conditions (studies 3 and 4). 32 Total freezing behavior was calculated as a percentage score as the proportion of “freezing” to “not freezing” observations for each mouse. For all other experiments, freezing was measured using automated Freezescan video tracking software (CleverSys, Inc.), which detected freezing behaviors and quantified these events by calculating total time spent freezing as a % of total session time.16,34
Light/dark test
The light/dark test is well-validated model for measuring anxiety-like behavior in rodents. 37 The opaque Perspex box is divided into two chambers: the enclosed dark chamber with black Perspex walls (28 cm wide×18 cm long×30 cm high) and a larger, open light chamber with blue Perspex walls (28 cm wide×27 cm long×30 cm high). Mice were placed in the center of the light side at the beginning of testing (150±10 lux). A video camera was positioned above the light box and distance travelled in the area, time spent in the area, and entries into the dark box were recorded using TopScan™ software (CleverSys, Inc.).
Experimental design
Study 1: Assessment of whether repeated CBG exposure disrupts long-term contextual fear memory
Mice underwent contextual fear conditioning and 12 h later were administered the first injection of vehicle or 10 or 30 mg/kg of CBG. We tested these doses as they were ≥ the CBG dose of 10 mg/kg, which reduces neuroinflammation in mice. 20 Twelve hours after the first injection, mice were administered the same drug, daily, for an additional 20 injections. A control group was not fear conditioned and treated with vehicle. Seventy-two hours after the last injection, mice were returned to the fear conditioned context and freezing was measured (Fig. 1a). We wished to washout CBG to address whether repeated administration during fear memory incubation was critical to any fear disrupting effects, rather than the acute effects it might have on fear memory expression. CBG's elimination half-life in mouse brain was ∼4 h, so 72 h was deemed sufficient to remove CBG from the brain. 31

CBG did not disrupt long-term conditioned fear memory or conditioned fear memory expression.
Study 2: Assessment of whether acute CBG disrupts expression of contextual fear memory
Mice underwent contextual fear conditioning. A control group of mice was placed in the fear conditioned context without foot shock being applied. Twenty-four hours later, mice were administered vehicle or 30 mg/kg of CBG and then returned to the fear conditioned context where freezing was measured (Fig. 1b).
Study 3: Assessment of whether acute CBG disrupts the acquisition of contextual fear memory
α2R agonists disrupt acquisition, consolidation, and reconsolidation, and CBG potently activates α2R with an EC50 of 200 pM.22,25–29 Therefore, in Study 3, 4, and 5, we included a lower dose of CBG (1 mg/kg) to more selectively activate α2R. Mice were administered vehicle or 1, 10, or 30 mg/kg of CBG, 2 h before undergoing contextual fear conditioning. Twenty-four hours later, mice were re-exposed to the conditioned context and freezing was measured (Fig. 2a).
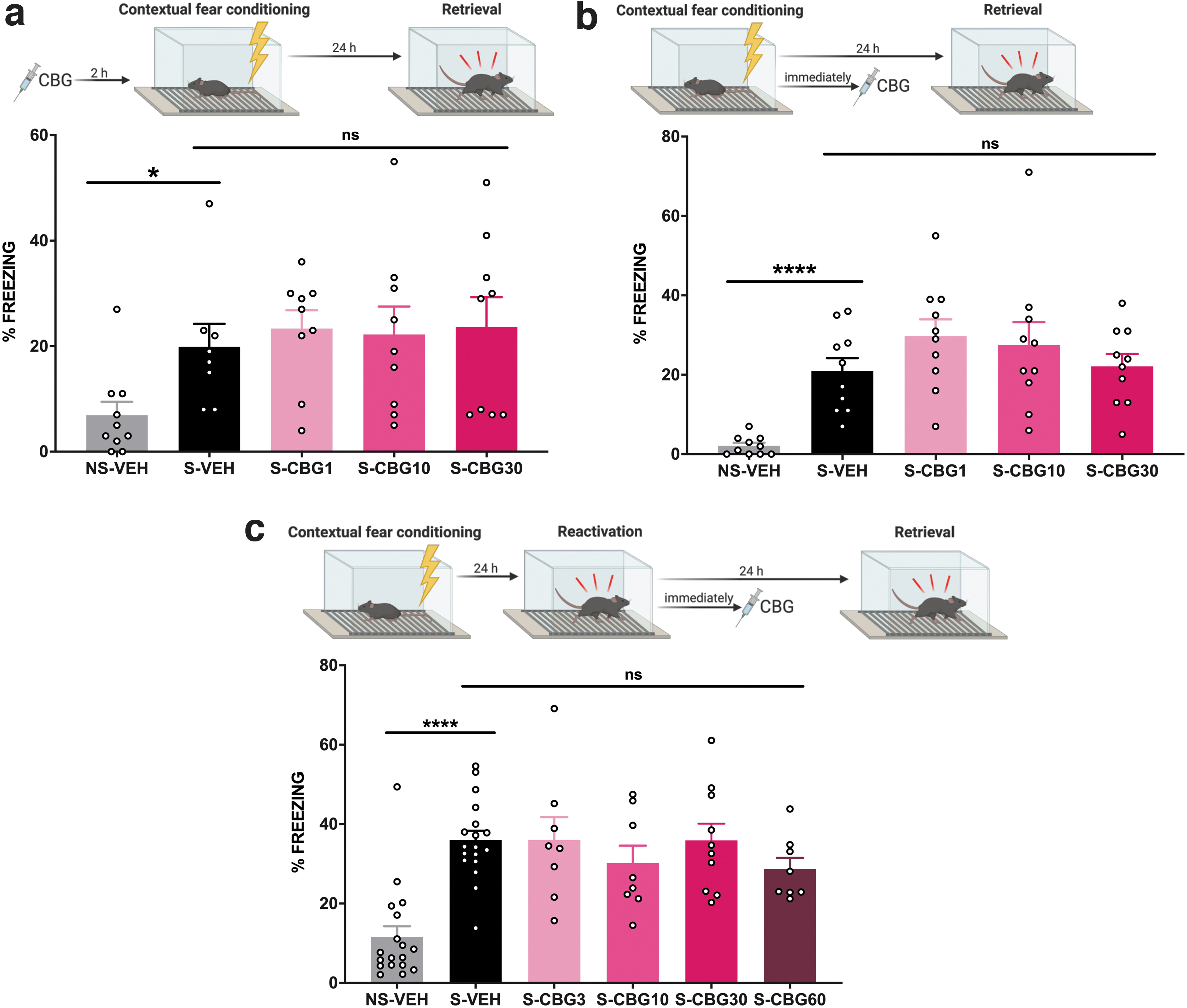
CBG did not disrupt acquisition, consolidation, or reconsolidation of contextual fear memory.
Study 4: Assessment of whether acute CBG disrupts the consolidation of contextual fear memory
Mice were administered vehicle or 1, 10, or 30 mg/kg of CBG, immediately after contextual fear conditioning. This time point was chosen as consolidation preserves the conditioned fear memory into long-term memory, and is usually completed within 6 h of the initial fear conditioning, 38 and CBG has a brain half-life of ∼4 h. 31 Twenty-four hours later, mice were re-exposed to the conditioned context and freezing was measured (Fig. 2b).
Study 5: Assessment of whether acute CBG disrupts the reconsolidation of contextual fear memory
Mice underwent contextual fear conditioning and were returned to their home cages. Twenty-four hours later, mice were returned to the fear conditioned context for 180 sec in the absence of foot shock to reactivate the fear memory. Immediately after, mice were administered vehicle, 3, 10, 30, or 60 mg/kg of CBG. We used a CBG dosing regimen here that delivered higher doses, as the previous CBG dose series on acquisition and consolidation or fear memory was ineffective. Mice were injected immediately after the reactivation session as CBG has a brain half-life of ∼4 h and reconsolidation of fear memory occurs within 6 h of reactivation.31,39 Reactivation of the fear memory renders it labile and vulnerable to pharmacological disruption. Twenty-four hours later, mice were re-exposed to the conditioned context and freezing was measured to assess reconsolidation of the fear memory (Fig. 2c).
Study 6: Assessment of whether acute CBG affects trauma-induced anxiety-related behavior in the light-dark test
Traumatic foot shock exposure alters anxiety-related behaviors in the light-dark test. 32 Mice underwent contextual fear conditioning and 24 h later were administered vehicle, or 10 or 30 mg/kg of CBG, 2 h before being placed in the light-dark test apparatus (Fig. 3a). Higher CBG doses were used here, consistent with CBG activity at higher concentrations on other targets, rather than specifically α2R, including the anxiety-relevant receptors 5-HT1A and CB1.22,40–42 The 60 mg/kg CBG dose was abandoned due to it being poorly soluble in our vehicle.

CBG did not disrupt trauma-induced generalized anxiety.
Statistical analyses
Statistical analyses were performed using GraphPad Prism (Version 8, GraphPad Software, CA). The acquisition of fear memory was assessed by comparing the non-shock-vehicle (NS-VEH) and shock-vehicle (S-VEH) groups using Mann–Whitney U tests where the data were not normally distributed, and unpaired t-tests when the data were normally distributed. To investigate the impact of CBG administration on fear memory, the S-VEH mice were compared to CBG-treated shock groups. An unpaired t-test was used in the cases where only two groups were compared. One-way analysis of variance (ANOVA) tests with Dunnett's multiple comparisons were used for comparison of more than two groups, and Kruskal–Wallis tests with Dunn's post-hoc tests were used when data violated normality or equality of variance assumptions of ANOVA. Post-hoc tests were performed with the S-VEH group as control. Statistically significant results were determined when p<0.05.
Results
Study 1: Repeated CBG exposure did not disrupt long-term contextual fear memory
We first examined whether repeated CBG exposure disrupted long-term contextual fear memories (Fig. 1a). Twenty-two days after the CS-US pairing, vehicle-treated shocked mice displayed a significantly increased % freezing compared to the control mice that were not shocked, consistent with long-term contextual fear conditioning (Mann–Whitney U test, p=0.012). However, repeated treatment with either 10 or 30 mg/kg of CBG did not affect the long-term fear conditioned freezing behavior [one-way ANOVA; F(2,27)=0.081 p=0.922].
Study 2: Acute CBG did not affect the expression of conditioned fear
We then wished to observe whether acute CBG inhibited the expression of conditioned fear (Fig. 1b). Contextual fear conditioning was observed as vehicle-treated mice that were shocked displayed significantly higher % freezing than control mice that were not shocked (Mann–Whitney U test, p=0.001). However, acute CBG treatment did not disrupt the expression of conditioned fear, as mice treated with 30 mg/kg of CBG did not differ from vehicle-treated mice in the shock group [unpaired t-test; t(18)=0.581, p=0.569].
Study 3: Acute CBG did not disrupt acquisition of contextual fear memory
We then assessed whether acute CBG exposure had any effect on the acquisition of contextual fear memory (Fig. 2a). Contextual fear conditioning was observed as vehicle-treated shocked mice had significantly higher freezing than their non-shocked counterparts (Mann–Whitney U test, p<0.0001). However, the acute administration of CBG did not diminish freezing compared to the vehicle-treated group [one-way ANOVA; F(3,36)=0.992, p=0.408], consistent with acute CBG not disrupting the acquisition of contextual fear memory.
Study 4: Acute CBG did not disrupt consolidation of contextual fear memory
Contextual fear conditioning was observed as vehicle-treated mice that were shocked had significantly higher freezing than control mice that were not shocked [Fig. 2b; unpaired t-test, t(16)=2.689, p=0.016]. CBG-treated mice did not have significantly different freezing compared to the vehicle-treated group at any of the doses tested [one-way ANOVA; F(3,31)=0.121, p=0.947], consistent with CBG failing to disrupt the consolidation of contextual fear memory.
Study 5: Acute CBG did not disrupt reconsolidation of contextual fear memory
Contextual fear conditioning was observed as vehicle-treated mice that were shocked had significantly higher freezing than the control group that did not receive shock [Fig. 2c; unpaired t-test, t(34)=6.763, p<0.0001]. Fear conditioned mice treated with CBG did not significantly differ from vehicle-treated mice [one-way ANOVA; F(4,47)=0.847, p=0.503], consistent with acute CBG failing to disrupt the reconsolidation of fear memory.
Study 6: Acute CBG did not affect trauma-induced anxiety-related behavior
Foot shock decreased the total distance travelled in the light area compared to mice that did not receive shock [Fig. 3b; unpaired t-test; t(37)=3.602, p=0.0009], although there was no change in the time spent in the light box (Fig. 3c; Mann–Whitney U test, p=0.465). Foot shock also significantly reduced the number of entries into the dark box [Fig. 3d; unpaired t-test, t(37)=4.408, p<0.0001], and increased the latency for first entry into the dark box (Mann–Whitney U test, p=0.002; Fig. 3e).
Acute CBG treatment did not alter the effects of foot shock on the total distance travelled in the light area [one-way ANOVA; F(2,54)=0.578, p=0.564], the number of entries in the dark box [one-way ANOVA; F(2,54)=0.318, p=0.729], or the latency to enter the dark box [Kruskal–Wallis test, H(54)=3.068, p=0.216].
Discussion
CBG is being touted as the “next big phytocannabinoid” and it has been suggested that CBG might have a place in the treatment of neuropsychiatric disorders.9,10,30 However, the evidence base around the pharmacology of CBG is limited and further research is needed to validate its potential medicinal properties. In this study we examined whether CBG disrupts contextual fear memory processes and shock-induced anxiety-related behavior in mice. Repeated CBG exposure did not influence long-term fear memory. Moreover, acute CBG administration failed to disrupt the acquisition, expression, consolidation, or reconsolidation of contextual fear memory. Finally, acute CBG did not affect stress-induced anxiety-related behavior.
It is highly unlikely that our negative findings are explained by chemical degradation of our CBG stock. First, the CBG active pharmaceutical ingredient we tested had a purity of 99%. Furthermore, pilot experiments confirmed adequate CBG exposure in mice using high performance liquid chromatography and tandem mass spectrometry (HPLC-MS/MS). In one study, we dosed adult male C57BL/6 mice with CBG 30 mg/kg i.p. daily for 3 days consecutively. Two hours after the last injection, which is the reported Tmax, we collected plasma of the mice and analyzed it for CBG concentrations. 31 The average plasma concentration was 23.64 μg/mL (75 μM). These data highlight the presence of our target compound CBG at μM levels in experimental mice following i.p. injection.
In this study we utilized a validated mouse model of PTSD in which mice exposed to foot shock display persistent fear memory, as well as other maladaptive behaviors such as hyperarousal, increased anxiety-related behavior, and disrupted rapid-eye-movement sleep.16,35 Fluoxetine, a drug used to treat PTSD patients, reduced long-term fear memory and associated neuroinflammation in this model, suggesting anti-inflammatory agents might be explored as novel treatments for PTSD. 15 CBG has robust anti-inflammatory effects in cellular and animal models of brain disorders20,40,43–45; thus, we sought to examine whether CBG reduces long-term fear memory in mice. Repeated CBG administration, however, was ineffective at reducing long-term contextual fear memory at 10 and 30 mg/kg doses. These encompass doses of CBG that were reported to reduce inflammation in mouse models of Huntington's disease and inflammatory bowel disease.19,20 For example, four daily i.p. injections of 10 mg/kg of CBG robustly decreased inflammatory and oxidative stress markers COX-2, TNFα, IL-6, and iNOs, as well as microgliosis in the brain of a 3-nitropropionate-induced model of Huntington's disease. 20
We next focused on whether CBG affected different memory processes, some of which are disrupted by CBD.32,46,47 Again, we did not observe CBG to have any activity, even when testing as high as 60 mg/kg. Our experimental fear conditioning paradigm produced consistent and statistically significant fear conditioned responses across all studies, but CBG was not able to reverse these effects. CBG is a very potent α2R agonist (EC50=200 pM) 22 and α2R agonists have been suggested to be promising novel PTSD therapeutics.23,24 Indeed, the α2R agonists clonidine and dexmedetomidine are known to disrupt acquisition, consolidation, and reconsolidation of conditioned fear in rodents.25–29 The doses of CBG we tested likely attained brain concentrations sufficient to exceed the low concentrations necessary to activate murine α2R 31 , therefore, it was surprising to observe no effect of CBG on acquisition, consolidation, and reconsolidation of contextual fear memory. CBG was characterized as an α2R agonist in murine isolated vas deferens and brain tissue, where its α2R activity was inferred using the α2R receptor antagonist yohimbine. 22 It would be of interest to observe whether CBG directly activates murine α2R in mammalian cell lines. We have screened CBG at 3 μM for its agonist activity at human α2A, α2B, and α2C adrenoceptors in human embryonic kidney 293 (HEK-293) cells, and there was no appreciable agonist activity at these receptors (Eurofins gpcrMAX panel; data not shown).
There are increasing claims made on the internet that CBG has anxiolytic effects and that it might reduce stress-induced anxiety. In this study, we could not find any effect of CBG on trauma-induced anxiety-related behavior in a model sensitive to the effects of the phytocannabinoid cannabidiolic acid. 32 One study recently reported a subtle anxiolytic effect of CBG, with CBG-treated mice spending more time in the center area of an open-field test at a dose of 10 mg/kg. 30 However, acute or repeated daily exposure of 2.5 mg/kg CBG did not affect anxiety-related behavior in rats in the light/dark test. 48
The lack of effects of CBG in this study contrasts with data showing CBD is anxiolytic and decreases the acquisition, expression, consolidation, and reconsolidation of contextual fear memory in rodents.32,46,47,49 This might be explained by the distinctive molecular actions that these phytocannabinoids have on the brain. For example, CBD's anxiolytic and disrupting effects on fear expression are, at least partly, mediated by 5-HT1A receptor activation, whereas CBG is a moderately potent 5-HT1A antagonist.22,50–52 The disruptive effects of CBD on reconsolidation involve cannabinoid CB1 receptors and might be explained by CBD indirectly activating these receptors by fatty acid amide hydrolase (FAAH) inhibition, which would increase concentrations of the endocannabinoid anandamide. 53 However, unlike CBD, CBG did not inhibit FAAH in rat brain. 53 CBG appears to directly activate CB1 receptors in cellular assays, but with much lower efficacy than THC. 54
While we could not find any effect of CBG in this study, it remains possible that CBG might affect other PTSD-relevant endpoints that were not addressed here. We did not examine whether CBG facilitates the extinction of fear memory. This might be anticipated, given reports that CBG weakly activates CB1 receptors, which are known to facilitate extinction.30,54 However, agonist activity of CBG at CB1 receptors must be functionally distinct from the effects of THC, as CBG tested up to 80 mg/kg did not produce the classical tetrad effects of THC in vivo, that is, hypothermia, reduced locomotor activity, catalepsy, and antinociception. 55 Perhaps, the subtle activity of CBG at CB1 receptors might be leveraged for therapeutic benefit without the liability of intoxication. CBG's potent α2R agonist activity might also be more relevant to managing hyperarousal and nightmares associated with PTSD, as α2R agonists, like clonidine, have been suggested to treat these PTSD symptoms. 23 Future studies might explore these potential applications of CBG in treating PTSD patients.
In conclusion, CBG was ineffective in disrupting various conditioned fear memory processes and stress-induced anxiety-related behavior in mice. These data suggest CBG has limited scope in the treatment of PTSD and stress-related anxiety.
Footnotes
Acknowledgments
The author gratefully acknowledges Barry and Joy Lambert for their continued support of the Lambert Initiative for Cannabinoid Therapeutics.
Author Disclosure Statement
J.C.A. is Deputy Academic Director of the Lambert Initiative for Cannabinoid Therapeutics, a philanthropically funded research initiative at the University of Sydney. He has served as an expert witness in various medicolegal cases involving cannabis and cannabinoids and as a temporary advisor to the World Health Organization on their review of cannabis and cannabinoids.
Funding Information
J.C.A. receives research funding from the Australian National Health and Medical Research Council (NHMRC) and the Lambert Initiative. J.C.A. reports the patents WO2019227167 and WO2019071302 issued.