
Abstract
Introduction:
The endocannabinoid system (ECS) plays a key modulatory role during synaptic plasticity and homeostatic processes in the brain and plays an important role in the neurobiological processes underlying drug addiction. Impaired endocannabinoid (eCB) signaling contributes to dysregulated synaptic plasticity, increased stress responsivity, and craving that propel addiction. Therefore, we hypothesized that boosting the ECS by exogenous administration of selective eCBs will attenuate cocaine-induced behaviors.
Materials and Methods:
The behavioral paradigms included psychomotor sensitization (PS) and conditioned place preference (CPP). Liquid chromatography-mass spectrometry analysis was used for quantitative profiling of eCBs in mouse brain.
Results:
We first measured the levels of eCBs in different brain areas of the reward system following chronic cocaine treatment. We found that following daily administration of cocaine, the levels of N-oleoyl glycine (OlGly) were significantly elevated in the nucleus accumbens (NAc) in a region-specific manner. We next tested whether administration of OlGly will attenuate cocaine-induced behaviors. We found that administration of OlGly during withdrawal, but not during acquisition of PS, attenuated the expression of cocaine sensitization. In addition, the administration of OlGly during the acquisition of cocaine CPP, but not during withdrawal, attenuated the expression of cocaine-conditioned reward. To enhance the stability of OlGly and its duration of action, two methylated derivatives of OlGly were synthesized, the monomethylated OlGly (HU-595) and dimethylated OlGly (HU-596). We found that the effect of administration of HU-595 or HU-596 during cocaine conditioning did not differ from the OlGly-induced decrease in the expression of CPP.
Conclusion:
Our findings suggest that the ECS is involved in the common neurobiological mechanisms underlying the development and expression of cocaine reward and drug-seeking. Boosting the ECS exogenously has beneficial effects against cocaine-induced behaviors.
Introduction
Drug addiction is a chronically relapsing disorder involving complex interactions among brain circuits, genetic background, environment, and an individual's life experiences. 1 It develops from an initial recreational drug use, evolves toward compulsive drug-seeking behavior despite negative consequences. 2 Substances of abuse such as cocaine act on the brain reward system, a neural circuitry that includes three principle areas, the ventral tegmental area (VTA), nucleus accumbens (NAc), and prefrontal cortex (PFC). 3
Growing evidence has demonstrated that the endogenous cannabinoid system (ECS) plays a significant modulatory role in synaptic transmission and homeostatic processes in the brain and it has been found to play an important role in the neurobiological processes underlying drug addiction.4,5 The ECS is composed of endogenous cannabinoids (eCBs), cannabinoid receptors, and the enzymes responsible for the synthesis and degradation of eCBs. 6 Moreover, N-acyl amino acids (NAAAs), eCB-like molecules, belong to the endocannabinoidome (eCBome) family of lipids with structural similarities with the eCBs. Basal levels of eCBs and eCB-like molecules differ in specific brain regions and the ECS is upregulated in certain disorders as indicated by an “autoprotective” increased endogenous release of those molecules. 7 Such increases have, for example, been observed in nicotine addiction. 8 This upregulation often appears to reduce or abolish nicotine-rewarding effects. 8
This discovery has raised the possibility of developing a treatment based on boosting the ECS to attenuate cocaine-induced behaviors by exogenous administration of eCBs. This may be especially important in the treatment of cocaine abuse and dependence, for which no proven effective pharmacotherapies exist. 9
We aimed to observe how chronic cocaine exposure will alter the levels of eCBs- and eCB-like molecules in specific brain areas. We also evaluated the pharmacokinetic profile of selected molecules and investigated whether the administration of the endogenous N-oleoyl glycine (OlGly)- and of eCBs-like molecules will exert neuroprotective effects against cocaine-induced behaviors using two behavioral paradigms; psychomotor sensitization (PS), which measures drug-induced neuroadaptations, and conditioned place preference (CPP), which measures drug reward.
Materials and Methods
Animals
All the animals used in this study were housed under specific pathogen-free conditions, up to five per cage, in standard plastic cages with natural soft sawdust as bedding. Four-week-old male C57Bl/6 mice (Harlan [Envigo] Laboratories, Jerusalem, Israel) were used. Animals were housed at an ambient temperature of 22–24°C, humidity at 30–70%, and alternating 12 h light/dark cycles (lights were on between 7:00 a.m. and 7:+00 p.m.) and provided with food and water ad libitum. During the treatment, body weight and food intake were monitored every day. All animal care and experimental protocols are approved by the Institutional Animal Care and Use Committee (IACUC; Reference: MD-20-16375-3) of the Hebrew University (Jerusalem, Israel).
Psychomotor sensitization
PS is most commonly assayed by measuring increases in locomotor activity after repeated experimenter-administered (no contingent) drug delivery.10,11 All mice were assigned to saline and cocaine treatment groups after a week of acclimation to their home cage environment.
One day before the first cocaine or saline injection, mice were habituated to the behavioral testing procedure by placement in photocell cages (MED Associates) for 30 min after saline injection. For chronic cocaine treatment (Fig. 1A, B), mice were injected with cocaine (20 mg/kg, IP.) or saline (1 mL

Levels of eCBs following repeated administration of cocaine.
On day 18, all mice/groups were challenged with cocaine (20 mg
Conditioned place preference
The CPP paradigm is a standard preclinical behavioral model used to study the rewarding or aversive effects of a stimulus. 13 The CPP apparatus (Med Associates) consists of two conditioning chambers of different designs and an intermediate chamber between them. One contains white-colored walls and wire mesh flooring (28×21 cm), while the other has black-colored walls and steel rod flooring (28×21 cm). The compartments are connected by a smaller center compartment (12×21 cm). Infrared beams located at the bottom of the wall allow assessment of animal preference for each compartment. CPP experiments were carried out at predefined times of the day.
Following a 3 day acclimation, a biased CPP design was conducted as follows: the animals were placed in the central gray compartment for 5 min, and then they were free to explore all three compartments for 15 min. The time spent in each compartment was analyzed by automated software and the results were used to determine the initial preference. The least preferred compartment for each subject was then assigned to be the drug-paired compartment. The conditioning period began 1 day following the habituation session. Cocaine or saline injections were given each day. The animals received four saline (1 mL
Thus, a total of 8 days of training were conducted. To evaluate the establishment of cocaine-induced CPP, we tested the animals 1 day following the last conditioning day, unless otherwise stated in the text. Each animal was placed in the central compartment for 5 min followed by a 15 min period of free access to all compartments. CPP score was defined as the percentage determined by 100
Endocannabinoid measurements
One milliliter of 1:1 mixture cold methanol and acetonitrile were added to each tissue followed by d4-PEA as an internal standard. The samples were maintained on ice and homogenized using a Polytron homogenizer (Kinematica). The homogenates were centrifuged for 20 min at 14,000 g at 4°C. Supernatants were collected and transferred to clean tubes and subsequently were evaporated to dryness under nitrogen and resuspended in 200 μL of 70:30 H2O:MeOH. Solid phase extraction (SPE) of the resultant solution was carried out using Bond Elute-C18 solid phase extraction columns (200-mg). The SPE columns were conditioned with 1.0 mL methanol and equilibrated with 1.0 mL water before sample loading.
After the extract was loaded, 0.5 mL of LC-MS-grade water was used to wash the cartridge and remove salt and other polar materials. The analytes were eluted using 100% MeOH (2×0.8 mL), and fractions were evaporated to dryness under nitrogen and stored at −80°C until analysis. Before analysis, samples were reconstituted in 50 μL MeOH.
Liquid chromatography-mass spectrometry analyses
Liquid chromatography-mass spectrometry (LC-MS/MS) analyses were conducted on a Sciex (Framingham, MA) Triple Quad™ 5500 mass spectrometer coupled with a Shimadzu (Kyoto, Japan) UHPLC System. Liquid chromatographic separation was obtained using a Kinetex (Phenomenex) column (C18, 2.6 μm particle size, 100×2.1 mm). The autosampler was set at 10°C and the column was maintained at 40°C during the entire analysis. Gradient elution mobile phases consisted of 0.1% formic acid in water (phase A) and 0.1% formic acid in acetonitrile (phase B). Gradient elution (250 μL/min) was held at 10% B for the first 0.5 min, followed by a linear increase toward 70% B in 0.5 min and a linear increase toward 75% B in 5 min.
The elution was maintained at 75% B for 3.5 min, then increased linearly to 99% B in 0.5 min and maintained at 99% for 13.5 min. eCBs were detected in positive and negative ion mode using electron spray ionization and multiple reaction monitoring mode of acquisition, using d4-Palmitoyl Ethanolamide as internal standard. The TurboIonspray® probe temperature was set at 650°C with the ion spray voltage at 4500 V. The curtain gas was set at 40.0 psi. The nebulizer gas (Gas 1) was set to 50 psi, the turbo heater gas (Gas 2) was set to 70 psi, and the collision gas (CAD) was set to 8 psi. The collision energy, declustering potential, and collision cell exit potential (CXP) for the monitored transitions are given in Supplementary Tables S1 and S2.
The dwell time was 30 ms. The levels of anandamide (AEA), 2-arachidonoyl glycerol (2-AG), PEA, OEA, OlGly, and OS in the samples were measured against standard curves. The values are expressed as pg/mg tissue. LC-MS/MS analyses were conducted as above, however, with few differences. The Kinetex (Phenomenex) column used was C18, 2.6 μm particle size and 50×2.1 mm. The Gradient elution (300 μL/min) was held at 10% B for the first 0.5 min, followed by a linear increase toward 70% B in 0.5 min and a linear increase toward 75% B in 4 min. The elution was maintained at 75% B for 4 min, then increased linearly to 99% B in 0.5 min and maintained at 99% for 3.5 min.
Statistical analysis
All data are presented as mean±SEM. For multiple comparisons between groups, one-way ANOVA was applied. For pairwise comparison one-tailed Student's unpaired t-test was used. Significance was tested using the one-tailed Student's unpaired t-test as indicated (GraphPad Prism v.8). A probability of p<0.05 was considered statistically significant, indicated by asterisks in the figures.
Results
Repeated cocaine administration increases the levels of OlGly in the NAc
To evaluate the role of the eCBs in cocaine-induced behaviors, we tested the effect of repeated cocaine administration on the levels of different eCBs- and eCB-like molecules, some were shown to have neuroprotective properties.7,14 Therefore, mice received daily repeated injections of cocaine (20 mg
As shown in Figure 1A, we found a significant increase in the levels of OlGly 15 in the NAc, but not in the PFC (Fig. 1B) or the hippocampus and cerebellum (data not shown) of cocaine-treated mice. AEA, N-oleoyl serine (OS), N-palmitoyl ethanolamide (PEA), 2-AG, and N-oleoyl ethanolamide (OEA) did not undergo any significant change in the NAc of cocaine-treated mice. These results raise the hypothesis that the region-selective endogenous increase of OlGly levels in the NAc may represent a self-defense protective mechanism against cocaine insult. We, therefore, speculated that elevating OlGly levels by administration of synthetic OlGly will reduce cocaine-induced behaviors.
OlGly attenuates the expression of PS and the acquisition of CPP
First, we conducted a pharmacokinetic experiment to assess whether synthetic OlGly can cross the blood–brain barrier (BBB). Therefore, mice received an acute injection of either vehicle or the maximally effective dose of OlGly (60 mg
We next tested the ability of OlGly to affect cocaine-induced behaviors. Cocaine has been shown to produce sensitization of locomotor behavior (PS) in animal models, which represents core aspects of addiction.
10
We tested the effect of OlGly on the acquisition or expression of PS. To test the effect of OlGly during the acquisition of PS, mice were pretreated with either vehicle or OlGly (60 mg
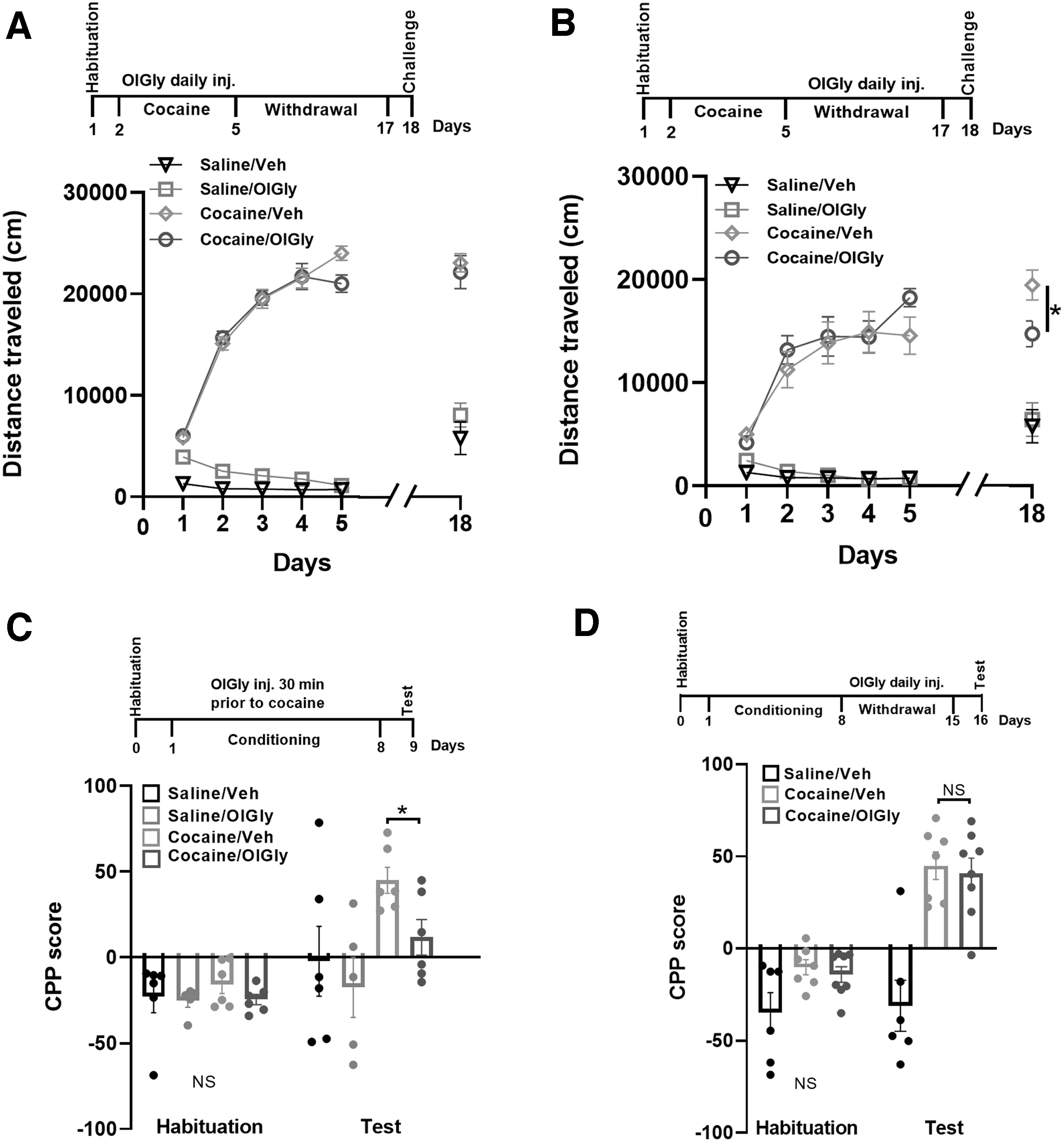
OlGly attenuated the expression of PS and CPP.
To test the expression of PS, mice were treated with cocaine as described above, and following termination of cocaine injection, mice were daily injected with OlGly (60 mg
Next, we tested the ability of OlGly to affect cocaine conditioned reward using CPP
13
during the acquisition phase and following withdrawal. Mice received a systemic injection of OlGly (60 mg
Animals received a daily injection of OlGly (60 mg
Evaluating the effect of OlGly derivatives on cocaine-induced PS and CPP
OlGly is a fatty acid amide susceptible to rapid degradation by fatty acid amide hydrolase (FAAH). 15 It was previously shown that introducing a methyl group in the α-position of the ethanolamine component of AEA 16 or to the glycine carbon atom in OlGly next to the amide bond (Oleoyl Alanine, OlAla, HU-595) 17 stabilizes the molecule resulting in remarkable metabolic stability and prolonged duration of action. 17 We, therefore, tested the ability of the monomethylated OlGly (HU-595), as well as dimethylated OlGly (HU-596), in which two methyl groups were introduced to the carbon atom in glycine next to the amide bond, to interfere with PS or CPP. Like OlGly, HU-595 (oleoyl alanine) is found, together with other NAAAs, in both invertebrate and mammalian tissues, including the mouse brain.18,19
We systemically injected these compounds during withdrawal from PS or during the acquisition of CPP, when OlGly exerts its behavioral effect. We first tested whether acute administration of HU-595 affects locomotor activity. Mice received either vehicle or increasing doses of HU-595 (6, 30, 60 mg
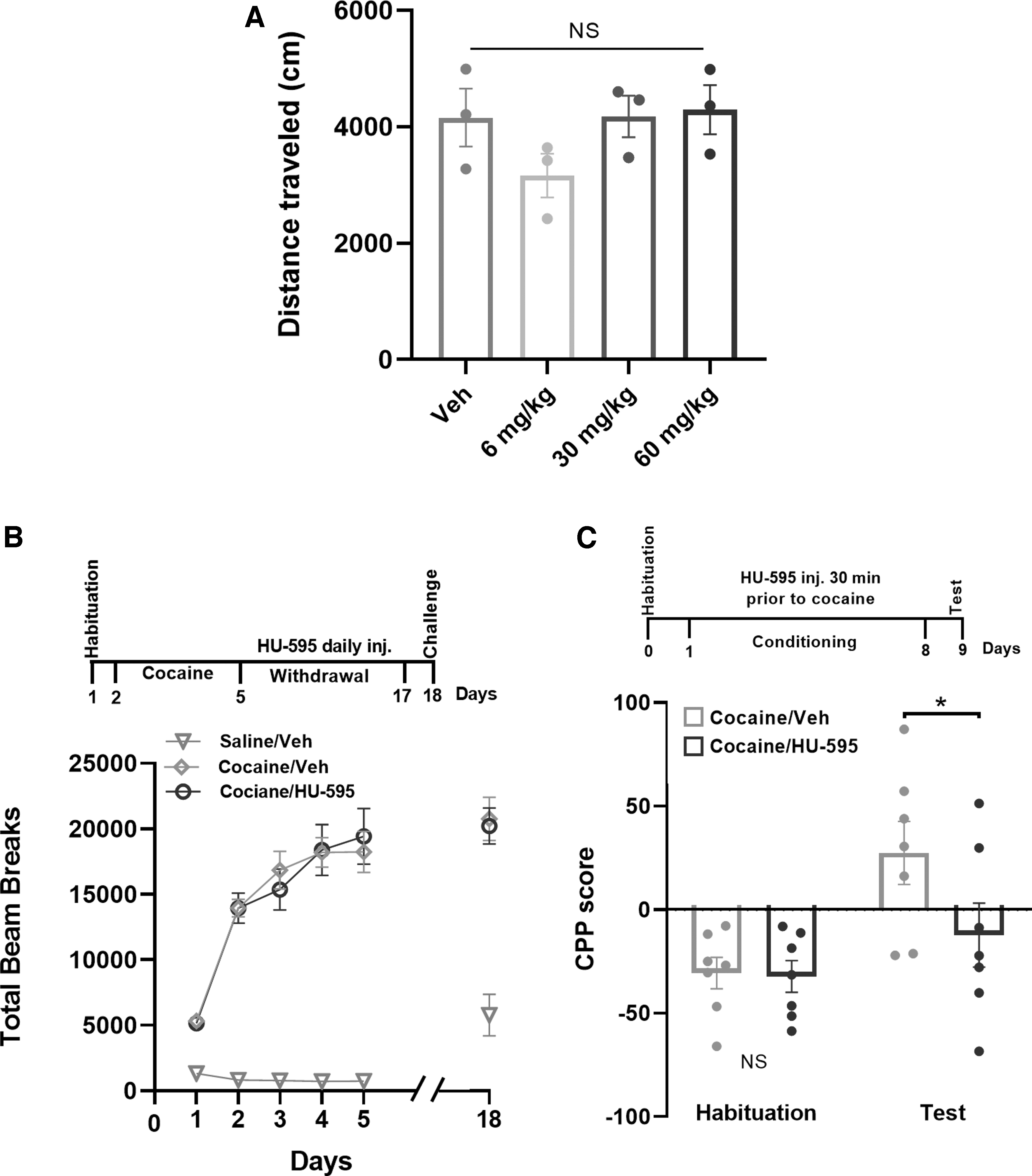
Oleoyl alanine (HU-595) decreased the expression of cocaine-induced CPP but not PS.
To investigate the effect of HU-595 on cocaine condition reward, we performed CPP experiment. Since OlGly negatively affected the acquisition of CPP when given during conditioning but not when injected after withdrawal (Fig. 2C, D), we tested the ability of HU-595 to affect cocaine CPP during acquisition. Administration of HU-595 (60 mg/kg) during the acquisition of CPP, similar to OlGly, abolished the expression of CPP (Fig. 3C).
We next tested the effect of dimethylated OlGly (HU-596) on PS and CPP at the same time schedule as for HU-595. As shown in Figure 4A, HU-596 did not modify the locomotor activity and the levels of HU-596 were significantly higher in the brain in a dose-dependent manner (data not shown). When HU-596 was administered during withdrawal from PS, no effect on the expression of PS was found (Fig. 4B). However, when given during conditioning in the CPP paradigm, abolished the expression of CPP (Fig. 4C), similar to HU-595. Together these results suggest that both compounds act similarly on the acquisition of cocaine reward similar to OlGly.

Dimethylated OlGly (HU-596) decreased the expression of cocaine-induced CPP but not PS.
Discussion
We hypothesized that the ECS will exert neuroprotective effects against cocaine insult. Therefore, in the current study, we evaluated whether elevating ECS exogenously will affect cocaine-induced sensitization and rewarding properties, two core components of addiction. We found that chronic administration of cocaine results in a region-selective endogenous increase of OlGly in the NAc. Chronic administration of OlGly during withdrawal from noncontingent cocaine administration reduced significantly the expression of PS, without affecting the acquisition phase. In addition, the administration of OlGly or its derivatives during the acquisition but not following withdrawal from CPP, attenuate the preference for cocaine. Together, these results suggest that OlGly and its derivatives are effective in interfering with cocaine-induced neuroadaptations and reward processing.
ECS was shown to play a beneficial role in disease states such as inflammation and pain, 20 epilepsy, 20 traumatic brain injury (TBI), 14 addiction, 21 and more. The concept that ECS exerts neuroprotective potential against brain disease is widely accepted. For example, it was shown that the levels of the endocannabinoid 2-AG were significantly elevated following TBI in a brain region-specific manner. 22 Exogenous administration of synthetic 2-AG to mice after closed head injury significantly reduced brain edema, increased clinical recovery, reduced infarct volume, and reduced hippocampal cell death. 22 In addition, it was shown that TBI elicited substantial increases in OlGly in the insular cortex of mice. 23 Chronic administration of synthetic OlGly ameliorates the behavioral alterations associated with mild-TBI. 14
The role of ECS in reward-related brain areas has been extensively studied in the last two decades. 24 In particular, evidence for the role of OlGly in reward was recently explored. Administration of OlGly during conditioning to nicotine, but not morphine, dose dependently reduced the development of nicotine CPP. 23 OlGly-mediated activation of peroxisome proliferator-activated-receptor-α (PPAR-α) resulted in reduced nicotine CPP, and the PPAR-α antagonist GW6471 prevented the OlGly-induced reduction of nicotine CPP in mice, suggesting that this lipid acts as a functional PPAR-α agonist to specifically attenuate nicotine reward. 23 Consistent with its “anti-addictive” properties, PPAR-α stimulation showed selectivity for reducing nicotine reward over the reinforcing effects of cocaine, cannabinoids, and morphine. 21
The selectivity of OlGly in blocking nicotine CPP may be explained by its negative modulation of β2-nAChR via activation of PPAR-α. This activity ultimately inhibits nicotine-induced firing and bursting activity in VTA dopamine neurons. 25 In the current study, we found that chronic treatment with cocaine increased the levels of OlGly in the NAc, but not in other reward-related areas such as PFC or the VTA, and exogenous administration of OlGyl attenuated cocaine CPP, suggesting that the NAc may mediate OlGly action. However, since OlGly was administered systemically, we cannot rule out the possibility that OlGly may act in other brain regions to mediate its protective effect.
Moreover, since other fatty acid amines, such as OEA and PEA, attenuated nicotine CPP through activation of PPAR-α in VTA DA neurons, 26 it is reasonable to speculate that OlGly may act through the same pathway in attenuating cocaine-mediated reward and sensitization. Further investigation (e.g., administration of OlGly to specific reward-related areas) will shed light on its precise site of action and its underlying mechanism of action.
Previous studies have shown the involvement of eCBs in drug-induced PS. It was demonstrated that hyperactivity induced by the D2/3 dopamine receptor agonist is attenuated by inhibitors of endocannabinoid degradation in mice. 27 It was also shown that acute administration of oleoyl ethanolamide (OEA) reduced spontaneous locomotor activity and attenuated psychomotor activation induced by cocaine. 28 Acute administration of the chemically related palmitoyl ethanolamide (PEA) before challenge cocaine injection was able to attenuate the expression of cocaine-induced PS. 29 In addition, it was shown that treatment of rats with CBD during repeated injection of cocaine/caffeine prevented the expression of PS. 30
Together, these results suggest that increasing the ECS tone plays a critical role in attenuating drug-induced adaptations. In the current study, we found that administration of OlGly during acquisition did not affect the development of PS but significantly decreases its expression when given during withdrawal. Since the NAc is known to be critical for the expression of PS, 3 together with our findings demonstrating an increase in OlGly in the NAc, we speculated that the attenuating effect on the expression of PS in the withdrawal stage will likely occur in the NAc and therefore is not evident during acquisition, although we did not test this possibility directly.
Previous studies have shown that the eCBs are also involved in the acquisition of conditioned reward. It was shown that coadministration of cocaine with OEA resulted in attenuation of CPP expression in a dose-dependent manner. 28 Similarly, it was shown that the administration of OlGly before each nicotine injection abolished the expression of nicotine CPP. 23 Furthermore, CBD treatment reduces cocaine-induced CPP, but not PS 31 and intra cranio ventricular injection of CBD during acquisition, decreased the methamphetamine-induced CPP. 32 Together, these results suggest that CBs may interfere with the acquisition of conditioned reward, preventing the association between the drug and the context in which it was administered. Consistent with these studies, we found that OlGly interferes with the acquisition of cocaine reward when given during acquisition.
However, when OlGly was administered daily following conditioning to cocaine, the expression of CPP, tested a week later, remained unchanged suggesting that OlGly does not affect acquired associations and treatment targeting the acquisition is more efficient.
OlGly is a fatty acid amide susceptible to rapid degradation by FAAH. 15 It was shown previously that monomethylation of OlGly, leading to HU-595, results in prolonged response to acute naloxone-precipitated morphine withdrawal. 17 Dimethylation of OlGly, leading to HU-596, was also synthesized. Likewise, we synthesized and assayed these two derivatives. We hypothesized that these modifications will decrease hydrolysis of these molecules and thus enhance their stability and duration of action as well as their ability to penetrate the BBB, as a result of increased lipophilicity. Indeed, both compounds are found in high levels in the brain following systemic administration. When administered during the acquisition of CPP, the expression of CPP was attenuated similarly to OlGly.
However, unlike OlGly, neither compounds affected the development of sensitization to cocaine. These results may implicate that these methylated positions are less likely to be an essential part of OlGly's pharmacophore in attenuating OlGly-induced CPP, and therefore, the behavioral outcome remains unchanged. Since the mechanism which underlies PS differs from the mechanism for CPP, it is likely to assume that these modifications interfere with the signaling pathway mediating OlGly ability to attenuate the expression of PS. Further investigation is required to clarify the effective dose of these compounds in interfering with the acquisition of CPP.
Although the molecular mechanism of the action of these compounds is yet to be determined, the results of the current study suggest that the eCB-like molecules such as OlGly and its derivatives may serve as a new neuroprotective approach for the treatment of cocaine-induced behaviors.
Conclusions
Chronic cocaine treatment resulted in an increase in OlGly in the NAc. Exogenous administration of OlGly attenuates the expression of PS and in the acquisition of cocaine reward. Increasing the stability of OlGly by monomethylation (HU-595) or dimethylation (HU-596) of OlGly did not strengthen the effect of OlGly. We, therefore, conclude that OlGly may serve as a new drug target to attenuate cocaine-induced behaviors.
Footnotes
Author Disclosure Statement
No competing financial interests exist.
Funding Information
This work was supported by the Israel Innovation Authority (R.Y. and R.M.), the Israel Science Foundation (ISF; 1283/16 R.Y.), the David R. Bloom Center for Pharmacy at the Hebrew University of Jerusalem (R.Y.).
Abbreviations Used
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.