
Abstract
Introduction:
In this study, the effects of the cannabinoid CBD were assessed on cyclooxygenase (COX)-1 and COX-2 expression and activity in resting and activated human neutrophils (polymorphonuclear [PMN] leukocytes).
Methods:
COX expression was measured at the mRNA levels, whereas COX activity was assessed by enzyme-linked immunosorbent assay measurement of prostaglandin (PG)E2. In vitro experiments in a standard commercial acellular assay of COX-1/COX-2 activity completed the study.
Results:
Results show that CBD profoundly inhibits expression of COX-1 and COX-2 mRNA in activated PMN, however, without any significant consequences for PGE2 production. CBD, however, was able to induce a slight but significant direct inhibition of COX-2 in the acellular model.
Conclusion:
The effects of CBD occur in the μM concentration range, which is attained in humans with therapeutic doses of the drug, suggesting the clinical relevance of these findings.
Introduction
Cannabis and cannabinoids are used to treat chronic pain associated with several different medical conditions; however, the evidence of their clinical efficacy is still limited to a small number of clinical studies, and above all that the understanding of the relevant mechanisms involved in the analgesic effects induced by cannabinoids is still limited.1–6
The endocannabinoids and the prostaglandins (PG) are key players in nociception and several lines of evidence support the existence of a strong connection between endocannabinoids and PG systems in the modulation of pain perception. 7 In particular, PG are important mediators in the inflammatory process and their production is catalyzed by cyclooxygenase (COX) enzymes, which exist in at least two isoforms: a constitutively expressed enzyme, COX-1, and an inducible enzyme, COX-2. 8 COX are the main targets of nonsteroidal anti-inflammatory drugs (NSAIDs), which are the most frequently consumed drugs worldwide. There is now general acceptance that NSAIDs exert their anti-inflammatory, analgesic, and antipyretic effects by inhibiting to different extents COX-1 and COX-2.8–10
Interestingly, preliminary evidence exists that cannabinoids isolated from Cannabis sativa may result in inhibition of COX activity. 11 In particular, cannabidiolic acid (CBDA) selectively inhibited COX-2 activity with an IC50 value (50% inhibitory concentration) ∼2 μM, having ninefold higher selectivity in comparison with COX-1 inhibition, suggesting that naturally occurring CBDA in C. sativa is a selective inhibitor for COX-2. 12 Assessing the effects of cannabinoids on COX-1 and COX-2 in comparison with NSAIDs could help both in clarifying the mechanisms involved in CBD-induced antinociception as well as in the clinical development of CB for the treatment of chronic pain.
Human neutrophils (polymorphonuclear [PMN] leukocytes) express COX enzymes that produce PGE2, the main PG involved in nociception.13–16 We have previously shown that the nonpsychoactive cannabinoid CBD effectively inhibits human PMN functions, including migration, oxidative metabolism, and tumor necrosis factor (TNF)-α production. 17 This study was devised to assess the effects of CBD on COX-1 and COX-2 expression and activity in resting and activated human PMN. COX expression was measured at the mRNA level, whereas COX activity was assessed by enzyme-linked immunosorbent assay (ELISA) measurement of PGE2. In vitro experiments in a standard commercial acellular assay of COX-1/COX-2 activity completed the study. The effects of CBD were compared with those of reference NSAIDs such as indomethacin, nimesulide, paracetamol, and ibuprofen, which were tested at clinically relevant concentrations (Table 1).
Reference Nonsteroidal Anti-Inflammatory Drugs Used in the Study
Methods
Test drugs and control agents
CBD power (white/off-white or slightly yellow powder, Adven® batch no. EP20/054-07) was kindly provided by CURALEAFINT Curaleaf International Limited (London, United Kingdom). Certificates of analysis provided by CURALEAFINT shows that CBD constitutes 99.8% of the dried substance. Stock solutions of CBD 0.01 M were prepared in dimethylsulfoxide (DMSO), tightly covered with tinfoil and stored for up to 1 month at −20°C. From stock solutions, appropriate dilutions were prepared daily in RPMI medium as required by the experimental procedures.
Measurement of PGE2 production
To test the ability of CBD to affect PGE2 production both under resting and stimulated conditions, we used cells obtained from human blood buffy coats provided by the local blood bank (Ospedale di Circolo, Fondazione Macchi, Varese, Italy). Cells were separated by density gradient centrifugation as previously described. 17 Finally, freshly isolated PMN were resuspended at 1×107 cells/mL in RPMI 1640 medium supplemented with 10% fetal bovine serum and 1% penicillin/streptomycin and placed in sterile 5 mL test tubes. Cells were then stimulated with lipopolysaccharide (LPS) from Escherichia coli serotype O127:B8 or formylmethionine-lecyl-phenylalanine (fMLP) (0.1 μM) in presence of test substances at 37°C under a 5% CO2 atmosphere.
The concentration and incubation time of the test substances were different according to the experimental design and detailed for each condition. After incubation, cells were centrifuged (1500 g, 10 min, 20°C) and pellets and supernatants were harvested and stored at −80°C until further analyses were performed. Cell pellets were used for quantification of mRNA levels of COX-1 and COX-2 using real-time reverse transcription polymerase chain reaction as described hereunder. PGE2 protein levels were measured in cell culture supernatants using commercial ELISA kit according to the protocol supplied by the manufacturer.
Experimental design
As detailed earlier, CBD was dissolved in DMSO. For all the methods, and whatever the experimental conditions used, a sample of DMSO as vehicle alone (VH), prepared at the same expected concentration of CBD, was tested. CBD was used in the concentration range of 0.01–1 μM or at the fixed concentration of 1 μM, as appropriate.
To test the effect of CBD we selected two experimental conditions that through the text were named coincubation and preincubation. More in detail are as follows.
Coincubation
For this experimental set, CBD was always added to the culture medium together with stimulating agents (LPS or fMLP) when present, or at the beginning of the culture for the resting conditions.
Preincubation
In this experimental condition, CBD was added to the culture medium 1 h before the activating stimuli (LPS or fMLP) or to resting conditions.
COX-1 and COX-2 mRNA expression in cultured cells
To test the ability of CBD to affect COX expression, we measured mRNA expression of both isoform named COX-1 and COX-2 in cells cultured in both conditions detailed in the experimental design section.
COX-1 and COX-2 gene expression was assessed from cell pellets by real-time PCR. In brief, total mRNA obtained from PMN cells was reverse-transcribed with random primers, by using High-capacity cDNA RT kit (Applied Biosystems). cDNA was amplified with SsoAdvanced™ Universal Probes Supermix (BIORAD) for analysis of COX-1 and COX-2 gene expression. cDNA was assayed on StepOne® System (Applied Biosystems). Linearity of real-time PCR assays were tested by constructing standard curves by use of serial 10-fold dilutions of a standard calibrator cDNA for both genes and regression coefficients (r 2 ) were always >0.999 (data not shown). Gene expression level in each sample was represented as 2−ΔCt where ΔCt=[Ct (sample) − Ct (housekeeping gene)]. Relative expression was determined by normalization to 18S cDNA (analyzed by StepOne Software™ 2.2.2; Applied Biosystems).
Enzyme-based inhibition assay
We used an acellular model aimed at testing the ability of CBD to affect COX activity. The COX (human) Inhibitor Screening Assay directly measures PGF2α by SnCl2 reduction of COX-derived PGH2 produced in COX reaction. The prostanoid product is quantified through ELISA using a broadly specific antiserum that binds to all major PG compound. This assay includes human recombinant COX-1 and COX-2, allowing the user to screen specific inhibitors. COX-1 and COX-2 activity was measured using the COX (human) Inhibitor Screening Assay kit according to the protocol supplied by the manufacturer (No. 701230; Cayman Chemical, MI, USA). The concentrations and incubation times of the test substances are in accordance with the experimental design.
Statistics
Data are shown as mean±standard error of the mean (SEM), unless otherwise indicated, with n showing the number of replicates. Differences between groups were assessed by Student's t-test or by ANOVA using GraphPad Prism (9.3.1) as appropriate.
Results
Effect of CBD on COX-1 and COX-2 mRNA expression in human cultured PMN
The ability of CBD to affect COX-1 and COX-2 mRNA expression was tested in the two experimental conditions described in the Methods section.
Coincubation
Human PMN were cultured for 24 h alone or in the presence of different concentrations of CBD (0.1–10 μM) and COX-1 and COX-2 mRNA levels were measured. CBD did not affect either COX-1 or COX-2 mRNA expression in resting cells, and neither did indomethacin (1 μM), which was chosen for comparison (Fig. 1).
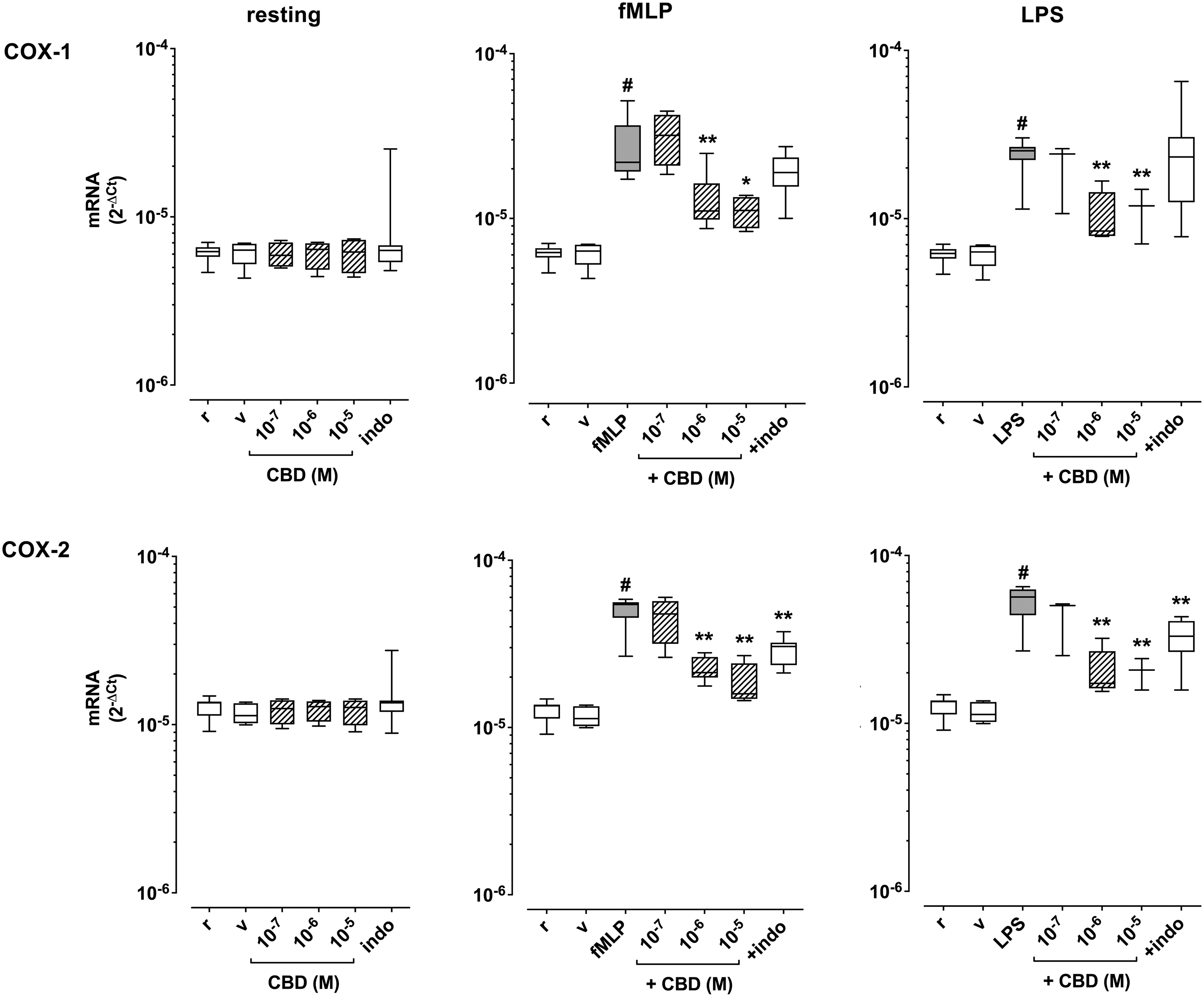
Effect of coincubation with CBD on COX-1 and COX-2 mRNA expression in human PMN under resting conditions (r) and after activation with fMLP (0.1 μM) or LPS (1 μg/mL). Indomethacin (indo, 1 μM) was used for comparison. Data are medians with 25°–75° percentiles (boxes) and min–max values (whiskers). #p<0.01 versus r, *p<0.05 and **p<0.01 versus fMLP or LPS. COX-1, cyclooxygenase 1; COX-2, cyclooxygenase 2; fMLP, formylmethionine-lecyl-phenylalanine; PMN, polymorphonuclear; LPS, lipopolysaccharide.
Treatment of PMN for 24 h with fMLP (0.1 μM) or LPS (1 μg/mL) significantly increased both COX-1 and COX-2 mRNA levels, and CBD concentration-dependently reduced the effects of both fMLP and LPS (Fig. 1). For comparison, indomethacin exerted a weaker effect in comparison with CBD, and the statistical significance was reached only with LPS (Fig. 1).
Preincubation
The ability of CBD to counteract LPS- or fMLP-induced COX-1 and COX-2 mRNA expression was also evaluated in preincubation. As detailed in the section “experimental design,” experiments were performed by adding CBD 1 h before the activating stimuli and finally culturing cells up to 24 h. In this set of experiments, we chose for comparison indomethacin (1 μM), nimesulide (1 μM), paracetamol (1 μM), and ibuprofen (5 μM) (Fig. 2).

Effect of 1 h preincubation with CBD (1 μM) on COX-1 and COX-2 mRNA expression in human PMN under resting conditions (r) and after activation with fMLP (0.1 μM) or LPS (1 μg/mL). Indomethacin (indo, 1 μM), nimesulide (nime, 1 μM), paracetamol (para, 1 μM), and ibuprofen (ibu, 5 μM) were used for comparison. Data are medians with 25°–75° percentiles (boxes) and min–max values (whiskers). #p<0.091 versus r, *p<0.01 versus fMLP or LPS.
Under these conditions, CBD (1 μM) profoundly inhibited both LPS- and fMLP-induced increase of COX-1 and COX-2 mRNA expression, an effect that was fully mimicked only by paracetamol and ibuprofen, whereas the indomethacin effect was only statistically significant on COX-2 mRNA, but it was in response to both fMLP and LPS; finally, nimesulide did not affect fMLP-induced increase of COX-1 mRNA expression (Fig. 2).
Effect of incubation with CBD on PGE2 production in human cultured PMN
The ability of CBD to affect PGE2 production was tested in the two experimental conditions described in the Methods section.
Coincubation
Human PMN were cultured for 24 h alone or in the presence of different concentrations of CBD (0.1–10 μM) and PGE2 was finally measured in supernatants. CBD did not affect PGE2 levels in resting cells, and neither did indomethacin (1 μM) (Fig. 3). Treatment of PMN for 24 h with fMLP (0.1 μM) or LPS (1 μg/mL) significantly increased PGE2 levels in PMN supernatants, however, and CBD did not affect such response. For comparison, indomethacin effectively prevented both fMLP- and LPS-induced increase of PGE2 (Fig. 3).
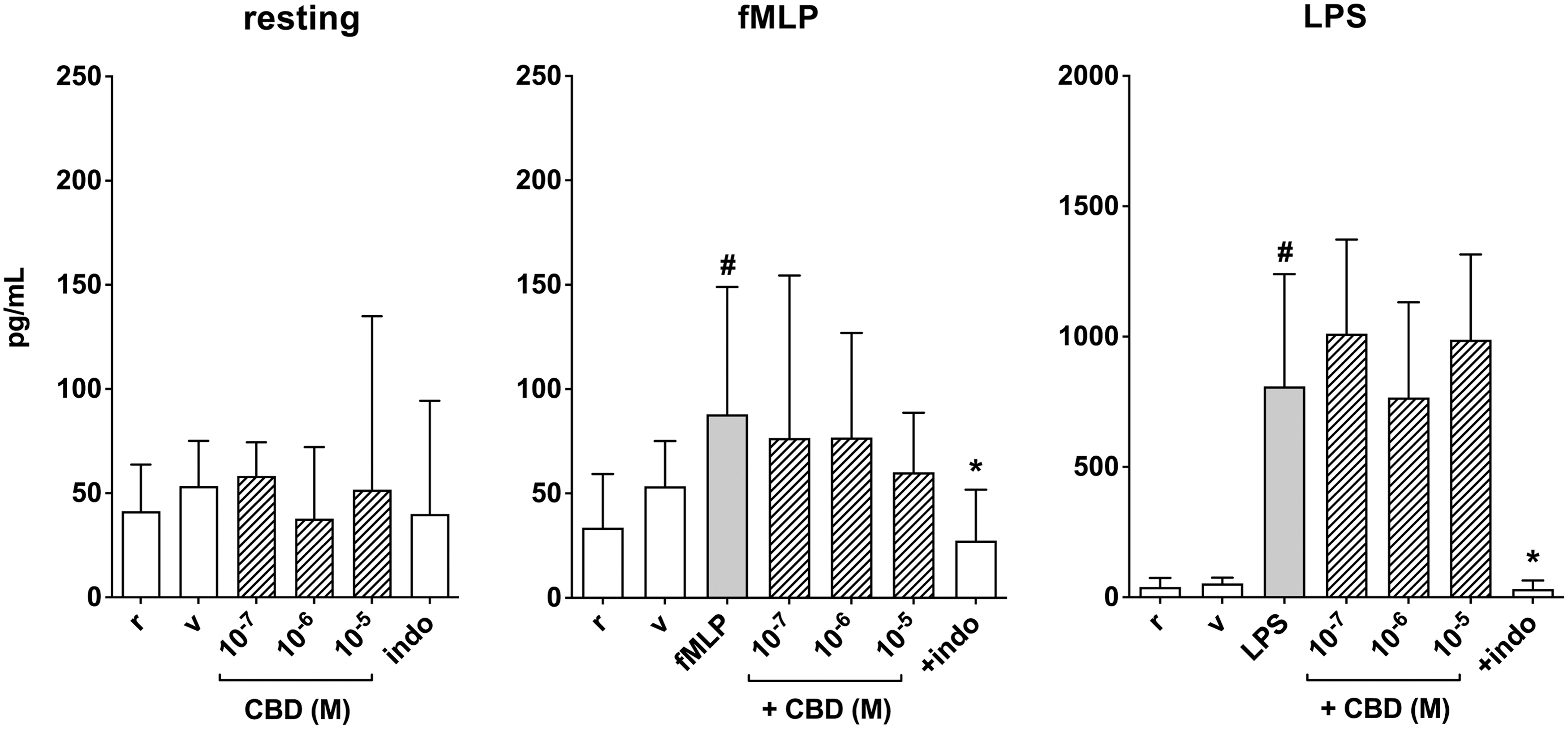
Effect of coincubation with CBD on PGE2 levels in supernatants from human PMN under resting conditions (r) and after activation with fMLP (0.1 μM) or LPS (1 μg/mL). Indomethacin (indo, 1 μM) was used for comparison. Data are medians with 25°–75° percentiles (boxes) and min–max values (whiskers). #p<0.01 versus r, *p<0.05 versus fMLP or LPS. PGE2, prostaglandin E2.
Preincubation
The ability of CBD to counteract LPS- or fMLP-induced increase of PGE2 in PMN supernatants was also evaluated in preincubation. To this end experiments were performed by adding CBD 1 h before the activating stimuli and finally culturing cells up to 24 h. Similarly, to the experimental design used to test PGE2 production in the condition of preincubation, also in this case we used indomethacin (1 μM), nimesulide (1 μM), paracetamol (1 μM), and ibuprofen (5 μM) as comparison drugs (Fig. 4).
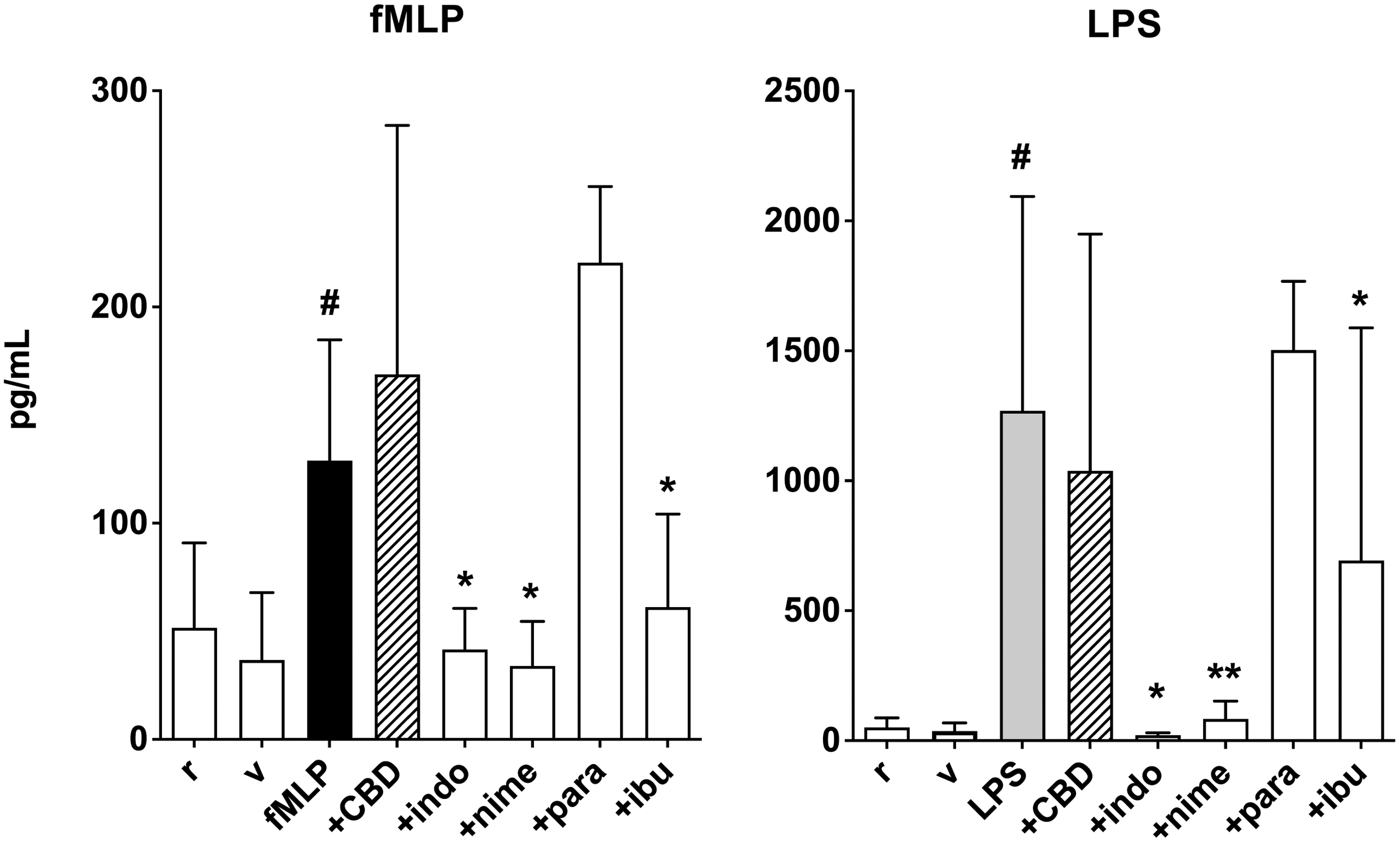
Effect of 1 h preincubation with CBD (1 μM) on PGE2 levels in supernatants from human PMN under resting conditions (r) and after activation with fMLP (0.1 μM) or LPS (1 μg/mL). Indomethacin (indo, 1 μM), nimesulide (nime, 1 μM), paracetamol (para, 1 μM), and ibuprofen (ibu, 5 μM) were used for comparison. Data are medians with 25°–75° percentiles (boxes) and min–max values (whiskers). #p<0.091 versus r, *p<0.05 and **p<0.01 versus fMLP or LPS.
CBD (1 μM) had no effect on LPS- or fMLP-induced increase of PGE2 in PMN supernatants, even if at least the effect of LPS was slightly reduced. Indomethacin, nimesulide, and to a lesser extent ibuprofen all inhibited LPS- and fMLP-induced increase of PGE2 levels, whereas paracetamol was ineffective (Fig. 4).
Direct inhibition of COX-1 and COX-2 activity
The ability of CBD to affect COX-1 and COX-2 activity was also investigated in an acellular assay. We used as reference drugs indomethacin to evaluate the effect of CBD on the COX-1 activity and nimesulide to evaluate the effect of CBD on COX-2 activity. The reference NSAID indomethacin (1 μM) effectively reduced COX-1 activity down to 19% of control values, and the reference NSAID nimesulide (1 μM) effectively reduced COX-2 activity down to 45% of control values (Fig. 5). CBD (1 μM) did not affect to any extent COX-1 but reduced COX-2 down to 78% of control values (Fig. 5).
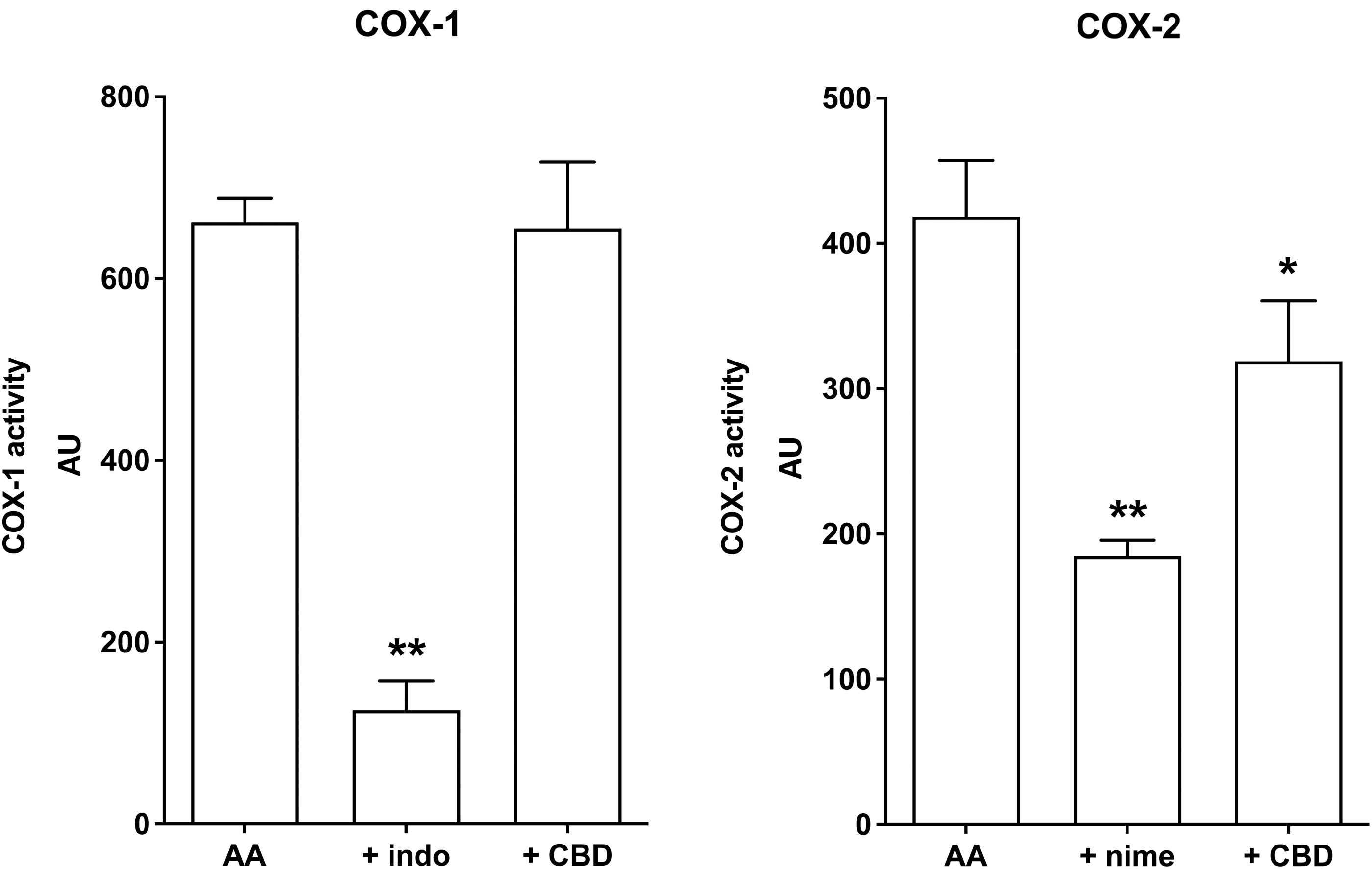
Direct effect of CBD (1 μM) on COX-1 and COX-2 activity measured as AU. Indomethacin (indo, 1 μM) and nimesulide (nime, 1 μM) were used for comparison. Columns are means±SD of n=3–4 separate experiments. *p<0.05 and **p<0.01 versus AA alone. AA, arachidonic acid; AU, arbitrary unit; SD, standard deviation.
Discussion
The main findings of this study can be summarized as follows: in human PMN, CBD profoundly inhibited activation-induced expression of COX-1 and COX-2 mRNA, however, without affecting PGE2 production in these cells. Nevertheless, the ability of CBD to induce a slight but significant direct inhibition of COX-2 in the acellular model suggests that CBD may affect not only expression but also activity of COX-2.
Despite the well-characterized connection between endocannabinoids and PG pathways in pain perception 7 only fragmentary evidence was so far available about the effects of CBD on COX activity and PG production. Ruhaak et al 11 studied CBD and other cannabinoids in an in vitro enzyme-based COX-1/COX-2 direct inhibition assay, similar to the one used in our study, and in a PGE2 production radioimmunoassay in the human colon adenocarcinoma cell line HT29. They reported no effect of CBD in the COX-1/COX-2 direct inhibition assay and only slight inhibition of PGE2 production in the cellular assay. 11 In a rodent model of acute inflammation induced by intraplantar injection of carrageenan in the rat, however, CBD administration resulted in decreased PGE2 plasma levels and tissue COX activity, which accompanied a prominent antihyperalgesic effect. 18 Quite remarkably, CBD has been also reported to enhance COX activity, for example, in rodent femoral arteries 19 and in human lung cancer cells. 20
Our study adds to existing evidence, showing for the first time the ability of CBD to inhibit activation-induced expression of both COX-1 and COX-2 mRNA in human PMN. Such results further expand available knowledge about the ability of CBD to exert a range of anti-inflammatory effects in these cells and are in line with our previous findings showing CBD-induced inhibition of human PMN migration, oxidative metabolism, and TNF-α production. 17 Remarkably, the effects of CBD occur even in this study in the μM concentration range, which is usually attained in humans with clinically relevant doses of the drug, 21 suggesting their translational potential to the clinics.
In view of the marked inhibition exerted by CBD on activation-induced increase of COX mRNA levels, the lack of effect of CBD on PGE2 production in the same experimental conditions was unexpected, also considering the findings in the acellular model where CBD exerted a slight but nevertheless significant direct inhibition of COX-2. A simple explanation could be that CBD does not cross the cell membrane in human PMN, thus failing to reach intracellular targets including COX or because the intracellular mechanisms involved do not affects the PGE2 synthesis.
Nevertheless, in other cell types intracellular carriers for CBD as well as for other cannabinoids have been reported. 22 Another possibility could be that the isolated PMN model used in this study is not sensitive enough to detect the effects of CBD on COX activity. Finally, as in the acellular model CBD inhibited COX-2 but not COX-1, it could be also proposed that even if CBD slightly reduces COX-2 activity in human PMN, COX-1 alone is still able to afford PGE2 production under the conditions chosen for our experiments.
In our model, stimulation of PMN with either fMLP or LPS increased both COX-1 and COX-2 mRNA levels and resulted in enhanced production of PGE2. These results agree at least in part with previous findings showing that stimulation of human PMN with either fMLP or LPS, as well as with many other pro-inflammatory stimuli, results in upregulation of COX expression and increased PGE2 production.15,23 Nonetheless, such studies point to a major role of COX-2, while COX-1 expression and function in human PMN is usually reported as negligible. The prominent role of COX-2 in human PMN during inflammation is also indicated by experiments in volunteers who received bolus injections of LPS resulting in increased temperature, heart rate, and plasma cortisol. Under such conditions of experimental endotoxemia, circulating PMN showed increased expression of COX-2, which is apparently an early effect occurring 1.5–3 h after LPS injection. 24
Available evidence shows that incubation of human PMN with NSAIDs such as nimesulide effectively inhibits PGE2 production, however, without major effects on COX-2 expression. 15 Our results on the contrary clearly show that all NSAIDs tested effectively reduced both fMLP- and LPS-induced enhancement of COX-1 and COX-2 mRNA levels, with the notable exception of indomethacin that had no effect on either fMLP- or LPS-induced increase of COX-1 mRNA and of nimesulide that did not affect fMLP-induced increase of COX-1 mRNA. In our experiments, all NSAIDs were tested at clinically relevant concentrations, thus the occurrence of nonspecific effects is highly unlikely.
Interestingly, indomethacin, nimesulide, and ibuprofen, but not paracetamol, also effectively inhibited both fMLP- or LPS-induced increase of PGE2 production. The lack of effect of paracetamol is consistent with is the notion that this drug is a very weak inhibitor of COX, resulting in no significant anti-inflammatory effects, and it does not exert major effects on PMN activation. 25 Fragmentary evidence also exists suggesting the ability of paracetamol to affect at least some PMN functions such as the oxidative burst. 26
Paracetamol-induced inhibition of both fMLP- and LPS-induced enhancement of COX-1 and COX-2 mRNA levels observed in our experiments may thus add to the range of effects exerted by this drug on PMN. The data on the lack of paracetamol to affect stimuli-induced PGE2 production can also help to understand the same effect of CBD, that even able to affect COX mRNA expression, remain ineffective on PGE2-induced production, suggesting that PGE2 production has a more complicated regulation and different intracellular mechanisms are involved in their production.
In summary, these findings show the ability of CBD to prevent activation-induced increase of COX-1 and COX-2 mRNA levels. Although the ability of CBD to directly inhibit COX-2 also in human PMN remains to be established, results provide novel insights into the effects of CBD on human PMN, and may be relevant for the clinical use of CBD as an adjuvant drug to treat pain, especially during inflammatory conditions involving PMN. Several lines of evidence strongly support the immunomodulatory properties of CBD, which is an established anti-inflammatory agent endowed even with some immunosuppressive properties.27–29 Also, in view of its well-established safety and tolerability profile CBD appears thus as a highly promising drug with multiple effects potentially beneficial to treat disease conditions characterized by inflammation and pain.
Footnotes
Acknowledgment
Warm gratitude is expressed to CURALEAFINT (Curaleaf International Limited, London, United Kingdom) for providing the test materials as well as for an unconditioned grant that was used to perform most of the experiments.
Authors' Contributions
Conception and design of the study by M.C., F.M., B.P., and M.S.; measurement of PGE2 production and enzyme-based inhibition assay by A.L.; assay of COX-1 and COX-2 mRNA expression by M.L.; data analysis by A.L., M.L., and F.M.; interpretation of results by F.M., M.C., M.F., B.P., and M.S.; drafting of the article by M.C. and F.M. All authors were involved in revising it critically for important intellectual content, and all authors approved the final version to be published. All authors agree to be accountable for all aspects of the study in ensuring that questions related to the accuracy or integrity of any part of the study are appropriately investigated and resolved, and declare to have confidence in the integrity of the contributions of their coauthors.
Funding Information
The study was supported by funding of our University (FAR 2019-20 MC and FM).