
Abstract
Introduction:
The antidepressant properties of ketamine have been extensively demonstrated in experimental and clinical settings. However, the psychotomimetic side effects still limit its wider use as an antidepressant. It was recently observed that endocannabinoids are inolved in ketamine induced reward properties. As an increase in endocannabinoid signaling induces antidepressant effects, this study aimed to investigate the involvement of cannabinoid type 1 receptors (CB1R) in the antidepressant and psychostimulant effects induced by ketamine.
Methods:
We tested the effects of genetic and pharmacological inhibition of CB1R in the hyperlocomotion and antidepressant-like properties of ketamine. The effects of ketamine (10–20 mg/kg) were assessed in the open-field and the forced swim tests (FSTs) in CB1R knockout (KO) and wild-type (WT) mice (male and female), and mice pre-treated with rimonabant (CB1R antagonist, 3–10 mg/kg).
Results:
We found that the motor hyperactivity elicited by ketamine was impaired in CB1R male and female KO mice. A similar effect was observed upon pharmacological blockade of CB1R in WT mice. However, genetic CB1R deletion did not modify the antidepressant effect of ketamine in male mice submitted to the FST. Surprisingly, pharmacological blockade of CB1R induced an antidepressant-like effect in both male and female mice, which was not further potentiated by ketamine.
Conclusions:
Our results support the hypothesis that CB1R mediate the psychostimulant side effects induced by ketamine, but not its antidepressant properties.
Introduction
The endocannabinoid system comprises the endogenous ligands (endocannabinoids), the enzymes responsible for their synthesis and degradation, and the cannabinoid receptors.1,2 Anandamide and 2-arachidonoylglycerol (2-AG) are the best-characterized endocannabinoids, whose effects are mediated mainly by the cannabinoid type 1 (CB1) (cannabinoid type 1 receptors [CB1R]) and CB2 (CB2R) metabotropic receptors.1,3 CB1R are responsible for the psychotropic effects of Δ9-tetrahydrocannabinol, the primary psychoactive compound from Cannabis sativa.4,5 CB1R is expressed in distinct brain regions, including the hippocampus, pre-frontal cortex, and nucleus accumbens. 6 These structures are involved in motivational, rewarding, and emotional processes directly modulated by endogenous and exogenous CB1R ligands.7–9
Preclinical and clinical data fully support the involvement of the CB1R in the neurobiology of mood disorders, including depression.9–11 Manipulating endocannabinoid signaling alters depressive-like symptoms in animal models. For instance, stress-induced models of depression cause imbalances in endocannabinoid levels and downregulation of CB1R in various brain regions involved in emotional processing, reward, and cognition. 12 Accordingly, CB1R-deficient mice display depressive-like behavior,13–15 which is often observed in animals treated with CB1R antagonists.16–18 Moreover, the behavioral effects of antidepressant drugs are abolished by CB1R silencing or pharmacological blockade.11,19 Evidence in humans also supports the involvement of the endocannabinoid system in the neurobiology of depression since the CB1R antagonist rimonabant, developed as an antiobesity medication, was withdrawn from the market due to an increased risk for depression, anxiety, and suicidality.11,20
CB1R signaling is also essential for establishing addiction to cannabinoid drugs. 21 Genetic and pharmacological silencing of CB1R prevent the rewarding effects of synthetic and natural cannabinoids.21–23 Moreover, a controlled study in healthy cannabis users showed that the effects of the drug (i.e., cannabis) are mediated by CB1R. 24 Furthermore, genetic and pharmacological approaches suggest a critical role for CB1R in mediating the responses to other drugs of abuse, including psychostimulants. 25 Indeed, blockade of CB1R inhibits the motor hyperactivity and rewarding-related behaviors induced by cocaine and amphetamine.26,27 In line with the behavioral evidence, CB1R modulate the psychostimulant-activated signaling pathways. Both phosphorylation of extracellular signal-regulated kinase (ERK) and the increase in extracellular dopamine levels promoted by psychostimulants in mesocorticolimbic structures were prevented by CB1R silencing.26,28
Ketamine is a dissociative anesthetic derived from phencyclidine and is widely used in clinical practice.29,30 The discovery that ketamine induces rapid and sustained antidepressant effects revolutionized the perspectives for depression treatment since all monoaminergic-modifying antidepressants require weeks of treatment for their therapeutic effects to emerge.31,32 Moreover, ketamine is effective in patients who are resistant to the monoaminergic drugs, which corresponds to ∼30% of cases, and significantly decreases suicidality.33,34 However, the use of ketamine as an antidepressant is limited by undesired side effects, such as increased liability for abuse and dependence, which is observed even at therapeutic doses.29,35
Recent evidence indicates that the reinforcing and psychostimulant effects induced by ketamine involve increased endocannabinoid levels in the striatum and CB1R activation. 36 On the other hand, the antidepressant effect caused by ketamine was facilitated by AM251, a CB1R antagonist/inverse agonist.37–39 Since AM251 is also an agonist of GPR55 receptors, the precise involvement of CB1 receptors in the antidepressant effect induced by ketamine requires further investigation. 40
In this work, we used pharmacological and genetic tools to investigate the specific role of CB1R in different aspects of ketamine-induced behaviors (antidepressant and psychostimulant) under the same methodological conditions to facilitate direct comparison. By examining the psychostimulant and antidepressant-like effects of ketamine in the same animal, we aimed to demonstrate how much CB1R activation is involved in ketamine-induced effects. Moreover, we explored the possibility that blocking CB1R could attenuate ketamine psychostimulant side effects, while maintaining its antidepressant properties. We observed that motor hyperactivity promoted by ketamine was impaired in CB1R male and female knockout (KO) and wild-type (WT) mice treated with a CB1R antagonist. Moreover, genetic silencing of CB1R did not modify the antidepressant effect of ketamine in male mice, while the pharmacological blockade of CB1R induced an antidepressant-like effect per se in male and female mice.
Methods
Animals and housing
Adult male and female mice (7–8 weeks of age) of CD1 and Swiss strains were used. We used CB1 KO mice on a CD1 background, while the Swiss mice were used to perform pharmacological experiments with a CB1R antagonist. The mice were kept in the animal house in transparent polycarbonate boxes (20 cm width; 12 cm height; and 30 cm length; four animals per cage) with 2 cm of wood shavings. The room had a controlled temperature (24°C±1°C) and a 12-h light/12-h dark cycle (lights on at 6:00 AM). The animals had free access to food and water, except during testing.
All animal protocols were approved by the Ethics Committee on Animal Experimentation for the Use of Animals of the University of Sao Paulo (protocol no. 15.1.536.60.8) and by the Indiana University Bloomington Institutional Animal Care and Use Committee. Experiments were performed under the ethical principles adopted by the National Council for the Control of Animal Experimentation (CONCEA), which complies with international laws and policies for animal experimentation.
Drugs
The following drugs were used in the experiments: rimonabant (Tocris) at 3 and 10 mg/kg 27 and ketamine (Cristália, Brazil) at 10 and 20 mg/kg41,42; rimonabant was dissolved in cremophor–ethanol–saline (1:1:18, v/v). 27 S-ketamine was dissolved in sterile isotonic saline. The doses chosen were based on previous studies describing ketamine levels in the brain of mice treated with S-ketamine after 40 min, 43 and in our previous publication. 42 The solutions were prepared immediately before use and injected intraperitoneally in a volume of 10 mL/kg.
Forced swim test
The forced swim test (FST) was conducted as previously described. 42 Briefly, the mice were individually placed in a cylinder (18-cm diameter) with water at a depth of 10 cm for 5 min and allowed to swim freely. The water temperature was maintained at 24°C±1°C. Behavior was recorded, and the time spent immobile was registered during the test. The same experimenter analyzed all animals and was blind to the treatments to avoid individual bias. The water was changed after each trial to prevent the influence of alarm substances.
Following a previous study, 42 to avoid potential false positive or negative results due to increased individual variability in baseline immobility, all animals were submitted to a pre-test swim session (day 1) and randomized into the different treatment groups according to their immobility time in the session (high or low immobility, HI or LI, respectively). The average immobility of all animals in the pre-test was calculated, and an animal with higher immobility than the average was considered HI, whereas below average was considered LI. The HI and LI animals were randomly allocated to the different treatment groups without any exclusion. This strategy decreased the experimental variability and allowed the antidepressant effect to be observed with fewer animals per group. 42
Open-field test for locomotor activity
The locomotion experiments were conducted in a circular arena (40-cm diameter with a 50-cm high Plexiglas wall). The animals received the treatments and were individually exposed to the open field for 5 min, located in a separate experimental room with attenuated background noise and soft illumination (<200 lux). The exploratory activity was recorded, and the total distance travelled was measured by the software ANYMAZE (Stoelting). The arena was cleaned between each test using alcohol 70% to avoid interference by the smell of the previously tested animal.
Experimental design
To minimize variability in baseline immobility between animals, each animal was initially pre-exposed to the FST for 5 min (pretest session, day 1) (Fig. 1). Then, 24 h later, the same mice were submitted to the open-field test and, immediately after, to the FST for 5 min (test session). The short interval between the open field test (OFT) and FST was used to avoid a significant decrease in drug plasma concentration due to the prolonged exposure to the OFT or long interval between tests, which could confound data interpretation. 43 S-ketamine was administered 30 min before the open-field test, considering its pharmacokinetic profile observed in mice plasma and brain. 43 The CB1R antagonist was administered 10 min before the ketamine injection.

Schematic representation of the experimental procedure and treatment schedule.
Statistical analysis
Immobility time data obtained from the FST and the distance travelled observed in the OFT were analyzed by two-way analysis of variance (ANOVA) followed by Sidak's post-hoc tests. All results are presented as mean±standard error of the mean. The post-hoc tests were only applied when ANOVA results were significant (p<0.05).
Results
Effects of ketamine in CB1R male and female KO on locomotor activity
Figure 2A and B show that treatment with ketamine 20 mg/kg, but not 10 mg/kg, increased locomotor activity in WT male and female mice during the open-field assay. This effect was not observed in CB1R KO mice (male: significant effect of interaction [F(2,69)=7.02, p<0.05], but no effect of genotype [F(2,69)=0.62, p>0.05] or treatment [F(2,69)=1.760, p>0.05]; female: significant effect of treatment [F(2,58)=7.218, p<0.05] and genotype [F(2,58)=4.02, p<0.05], but no significant interaction [F(2,58)=1.93, p>0.05]; the post-hoc analyses revealed that ketamine increased locomotor activity only in WT mice). These data suggest that activation of CB1R is necessary for the psychomotor effects induced by ketamine in both male and female mice.
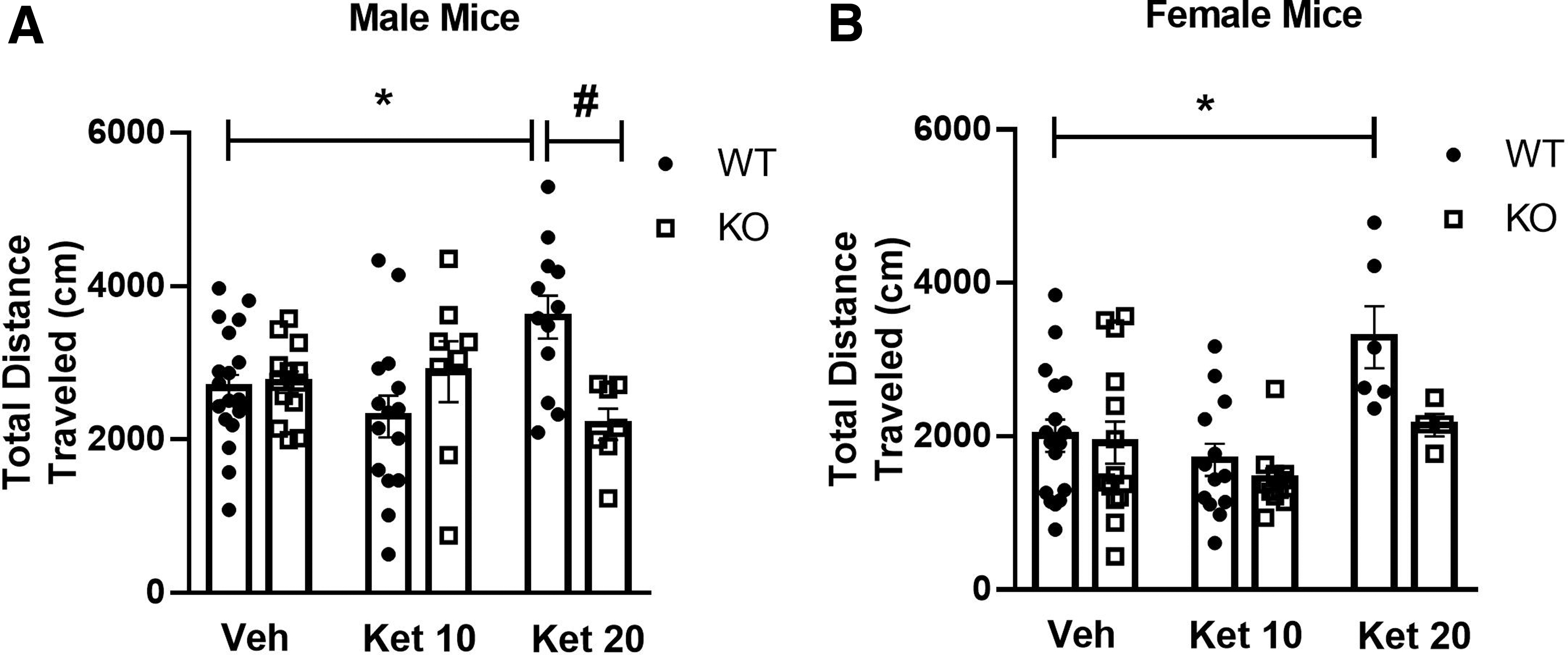
Effects of ketamine (10 and 20 mg/kg) in male
Effects of ketamine in CB1R male and female KO mice in the FST
There were no significant baseline differences in the immobility time (before any treatment) between CB1-R KO and WT, in male and female mice (interaction: F(1,59)=0,212; sex: F(1,59)=1,708; genotype: F(1,59)=0,13; p>0.05; Supplementary Fig. S1). However, from day 1 to 2, there was a noticeable increase in the immobility time in the male CB1-R KO, but not in the WT mice (interaction: F(1,60)=4.148; day: F(1,60)=9.636; p<0.05; data not shown), whereas no change was detected in females from day 1 to 2 (p>0.05; Supplementary Fig. S1).
On the test day, the treatment with ketamine (20 mg/kg) induced an antidepressant-like effect (decreased immobility time) only in CB1-R KO male mice (panel A) (significant effect of treatment [F(2,69)=3.43, p<0.05], and interaction [F(2,69)=2.43, p<0.05], but no effect of genotype [F(2,69)=0.40, p>0.05]). The post-hoc analyses confirmed that ketamine reduced immobility time only in male CB1-R KO mice, as depicted in Figure 3A and B. In females, no significant effect was observed (treatment [F(2,59)=0.32, p>0.05], genotype [F(2,59)=0.31, p>0.05], and interaction [F(2,59)=0.10, p>0.05]).
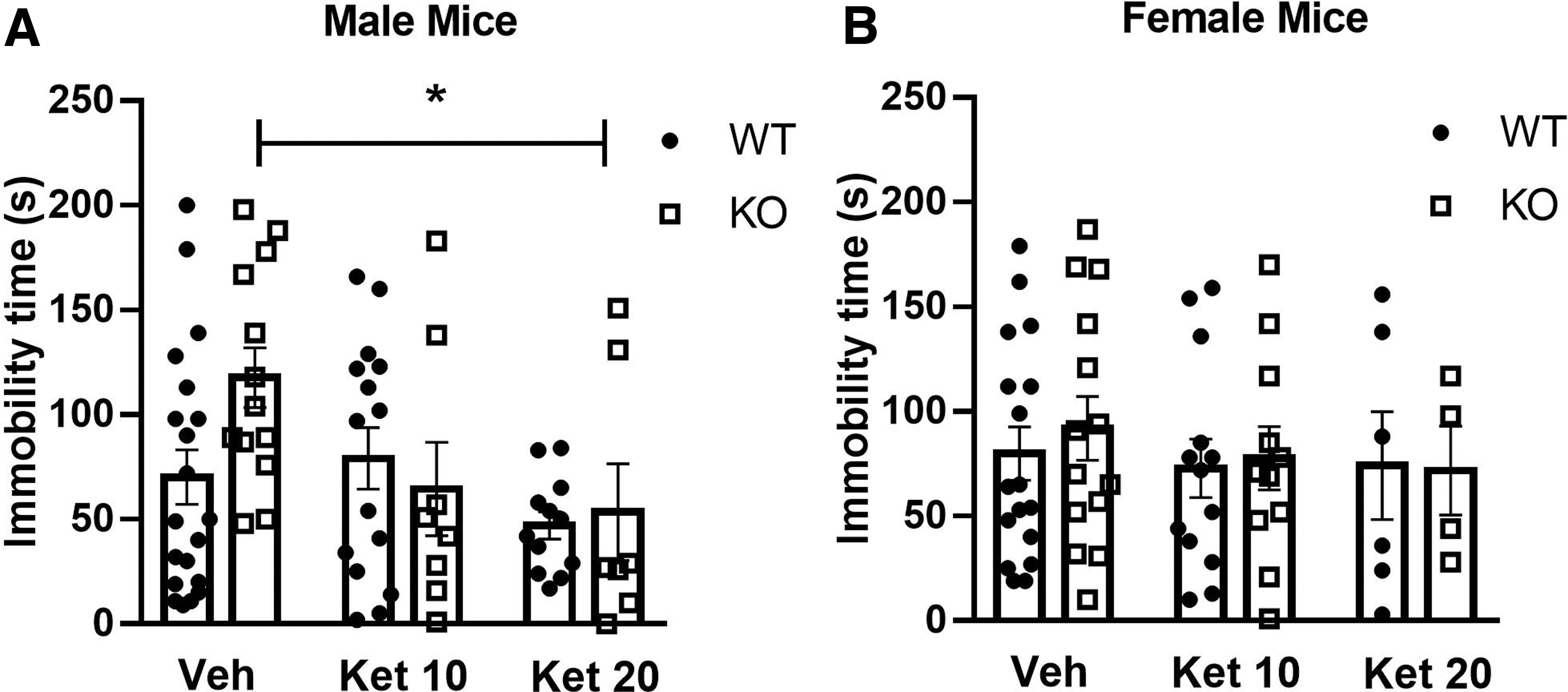
Effects of ketamine (10 and 20 mg/kg) in male
Altogether, these data suggest that the absence of CB1R facilitates the development of learned immobility in the FST in males, but not in females. Moreover, the lack of signaling through CB1R does not impair ketamine antidepressant effects, but rather facilitates it in male mice.
Effects of treatment with ketamine following pharmacological blockade of CB1R in male and female mice on locomotor activity
Figure 4A and B show that treatment with ketamine (20 mg/kg) increased locomotor activity, and this effect was prevented by previous acute administration of rimonabant, a CB1R antagonist/inverse agonist in both male and female mice. The two-way ANOVA indicated an effect of ketamine (F(3,34)=6.549, p<0.05) and of interaction (F(3,34)=5.957, p<0.05), but no effect of rimonabant was detected (F(3,34)=3.82, p>0.05), demonstrating that ketamine (20 mg/kg) induced hyperlocomotion in male mice when administered alone (Sidak's post-test, p<0.05 vs. control), but not when the CB1R antagonist was previously administered (Sidak's post-test, p>0,05 vs. control, Fig. 4A).

Effects of ketamine (20 mg/kg) and rimonabant (10 mg/kg) in male
A similar effect of interaction (F(3,27)=7.384, p<0.05) and of ketamine injection (F(3,27)=4.509, p<0.05), but not of rimonabant (F(3,27)=3.106, p>0,05), was observed in female mice, evidencing that treatment with ketamine (20 mg/kg) increased locomotor activity (Sidak's post-test, p<0.05 vs. control), and that this effect was inhibited by previous administration of the CB1R antagonist (Sidak's post-test, p>0.05 vs. control, Fig. 4B).
Effects of treatment with ketamine following pharmacological blockade of CB1R in male and female mice in the FST
Figure 5A and B show that both treatment with 20 mg/kg ketamine and the pharmacological blockade of CB1R with 10 mg/kg rimonabant significantly reduced the immobility time of mice submitted to the FST when compared to the vehicle-treated group (male: significant effect of ketamine treatment [F(3,34)=8.133, p<0.05], and of rimonabant injection [F(3,34)=9.381, p<0.05], but no interaction [F(3,34)=1.048, p>0.05]; female: significant effect of ketamine treatment [F(3,27)=9.176, p<0.05] and of rimonabant injection [F(3,27)=12.51, p<0.05], but no interaction [F(3,27)=2.437, p>0.05]). The post-hoc analyses revealed that both ketamine and rimonabant induced an antidepressant-like effect in male and female mice (Sidak's post-test, p<0.05 vs. control).
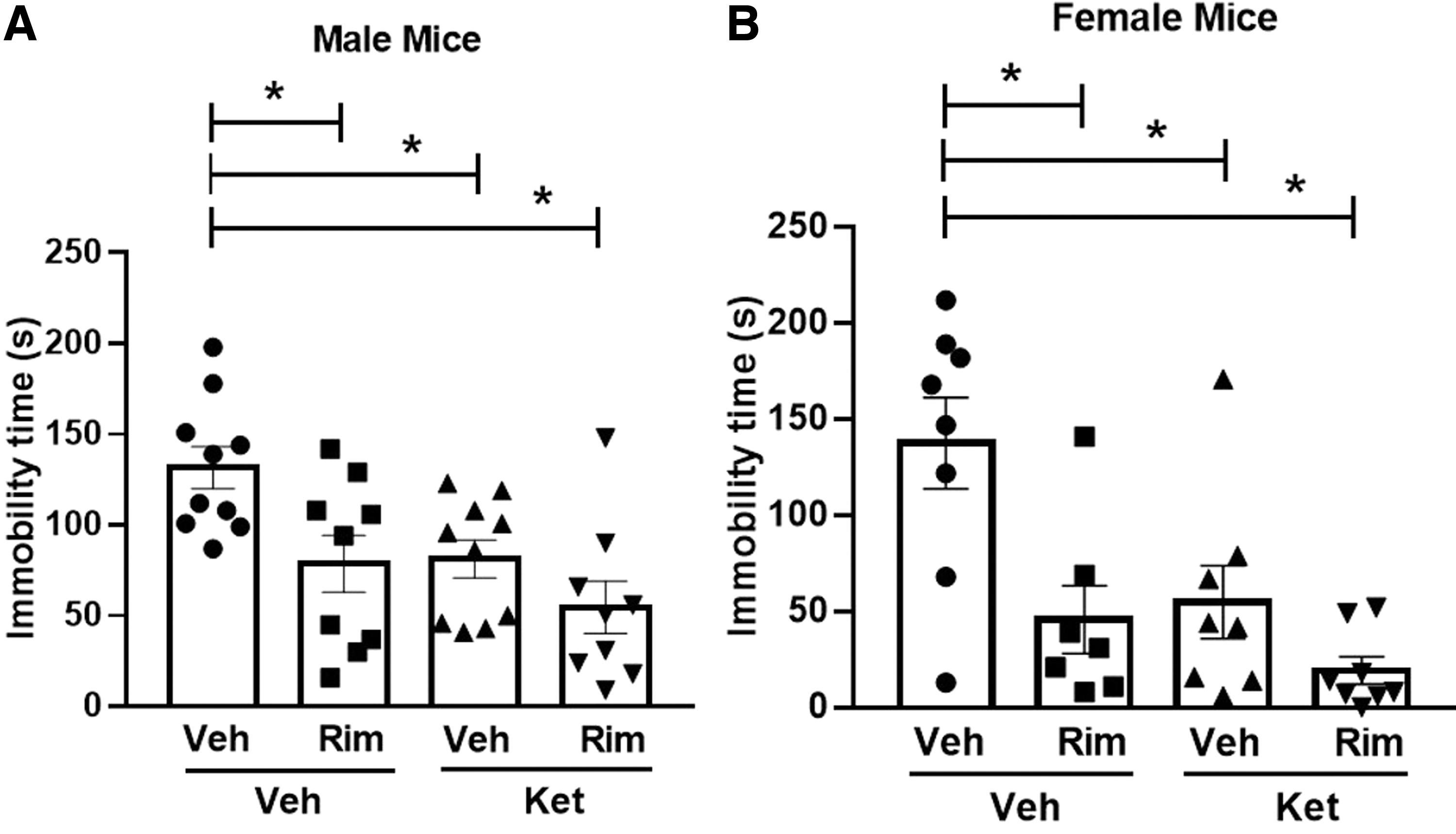
Effects of ketamine (20 mg/kg) and rimonabant (10 mg/kg) in male
Effects of combined administration of a lower dose of ketamine with a subeffective dose of rimonabant in mice submitted to the open-field test and FST
We tested if a lower dose of rimonabant (3 mg/kg) would facilitate the effects of a lower dose of ketamine (10 mg/kg). Figure 6A shows that this combination did not change the locomotor activity in male mice. The two-way ANOVA indicated no effect of ketamine injection (F(3,23)=0.93, p>0.05), rimonabant injection (F(3,23)=0.96, p>0.05), or interaction (F(3,23)=0.02747, p>0.05). Figure 6B shows that treatment with ketamine (10 mg/kg) significantly reduced the immobility time of male mice submitted to the FST compared to the vehicle-treated group.

Effects of combined administration of lower dose of ketamine (10 mg/kg) with a subeffective dose of rimonabant (3 mg/kg) in male mice submitted to open-field
The two-way ANOVA indicated an effect of interaction (F(3,23)=6.086, p<0.05) and of ketamine injection (F(3,23)=37.65, p<0.05), but not of rimonabant (F(3,23)=1.51, p>0.05). The post-hoc analyses revealed that ketamine induced an antidepressant-like effect (Sidak's post-test, p<0.05 vs. control), and this effect was not modified by previous treatment with a lower dose of rimonabant (Sidak's post-test, p>0.05 vs. ketamine group).
Discussion
This study provides evidence that the hyperlocomotion induced by ketamine was significantly reduced in CB1R KO animals and mice treated with rimonabant, a CB1R antagonist. Conversely, the antidepressant-like effect of ketamine was not blocked by the eliminating CB1Rs or by rimonabant. Considering that ketamine-induced hyperlocomotion has been associated with its psychostimulant properties in animal models, 44 our results suggest that CB1R are involved in the psychostimulant, but not in the antidepressant effects of ketamine.
The modulation of cannabinoid receptors has contradictory effects on motor behavior. While some studies have demonstrated that both CB1R activation and blockade biphasically45–47 modulate motor activity, our findings agree with those from other works that did not observe locomotor changes after blockade of CB1R.26,27 Moreover, corroborating our findings, CB1R signaling has been implicated in the modulation of psychostimulant and rewarding properties of drugs. For instance, locomotor responses to cocaine were also significantly reduced in CB1R KO mice compared with WT. 28 Moreover, a significant reduction in the acquisition of cocaine self-administration was observed after the KO of CB1R. 48 Similarly, pharmacological blockade of CB1R in WT mice attenuated the motor hyperactivity and rewarding effects of psychostimulant drugs.26,49
This study found that rimonabant treatment or absence of CB1R prevented hyperlocomotion induced by ketamine in both male and female mice. Although we did not directly assess other behaviors predictive of psychostimulant properties, previous publications have shown an association of hyperlocomotion in the open-field test with the psychostimulant properties of drugs, including ketamine. 50 Moreover, ketamine-induced hyperlocomotion is sensitive to antipsychotic drugs, 51 thus suggesting that hyperlocomotion might reflect aspects of ketamine psychostimulant properties.
Consistent with the interpretation of our results in the OFT, a recent study demonstrated that endocannabinoid signaling regulates the reinforcing and psychostimulant effects of ketamine. Repeated treatment with ketamine for 7 days and ketamine self-administration significantly increased 2-AG levels in the caudate-putamen. 36 The authors observed that repeated ketamine exposure significantly downregulated the expression of monoacylglycerol-lipase (MAGL), a 2-AG degrading enzyme.
They also showed that rimonabant administered either systemically or microinjected bilaterally into the dorsal lateral striatum (DLS) reduced ketamine-induced hyperactivity and self-administration. In accordance with the behavioral responses, intra-DLS infusion of rimonabant reversed the effects of repeated exposure to ketamine on phosphorylation of ERK and cAMP response element-binding protein, essential to the long-lasting effects of drugs of abuse. 36 These CB1R blockade studies suggest a critical role of cannabinoid signaling in the psychostimulant and reinforced behaviors driven by ketamine. Moreover, antagonism of CB1R also attenuated the reinstatement of ketamine-conditioned place preference in rats. 52 Collectively, these data indicate that CB1R plays a critical role in mediating the psychostimulant properties of ketamine in both male and female mice, which supports our findings observed in the open-field test.
Since ketamine induces rapid antidepressant effects, which are often accompanied by psychostimulant-like side effects, we investigated whether the absence of CB1R would also modulate the behavioral responses associated with the antidepressant effect in the FST. We observed that CB1R silencing did not modify the baseline immobility time in the FST in either male or female mice, but produced a discrete increase in the immobility time of males on day 2. Similar changes have previously been described upon re-exposure to the FST in CB1R KO male mice. However, the authors also described increased baseline immobility in both male and female CB1R KO mice. 53
Some of the contradictory findings can be associated with the mice strain used in different studies (CD1 vs. C57BL6/J) as the mice strain is one of the factors known to affect stress and drug-induced effects in the FST. 54 The effects induced by the absence of CB1R in depressive-like behavior are further complicated by the baseline condition of the animal (stressed vs. nonstressed) and the cell types from which CB1R is deleted.19,53,55 For instance, selective ablation of CB1R from forebrain GABAergic neurons or cells expressing dopamine D1 receptors did not change immobility time in the FST.13,56
However, an antidepressant-like effect was observed in mice lacking the CB1R in glutamatergic cortical neurons. 13 In another study, global CB1R KO mice showed an increased immobility time in the tail suspension test and higher sensitivity to develop depressive-like responses in the chronic unpredictable mild stress procedure.14,57 Our study observed no baseline difference in the immobility time in male and female CB1R KO mice on day 1 (pretest), but increased immobility time in CB1R KO male mice on the test day (24 h after the first swim session). Therefore, CB1R might have a more pronounced effect in regulating behavioral changes associated with depression under repeated stressful situations rather than from an acute challenge.
Similar to what is observed with the genetic deletion of CB1R, pharmacological CB1R blockade also induces heterogeneous response profiles in behavioral tests. Treatment with the CB1R antagonists/inverse agonists, such as rimonabant or its close structural analog, AM251, reduced immobility in both the tail suspension test and the FST.17,58,59 Moreover, in the chronic mild stress model, repeated administration of a CB1R antagonist for 5 weeks attenuated the deleterious effects produced by stress. 16 In accordance with these studies, we observed a reduction in the immobility time in male and female mice following treatment with rimonabant. Modulation of cortical levels of monoamines could be involved in such effects since acute blockade of CB1R increases the efflux of noradrenaline and 5-HT in the medial pre-frontal cortex, hypothalamus, and hippocampus. 60
Surprisingly, we found that, despite blocking its psychostimulant effects, silencing or pharmacologically blocking CB1R did not interfere with ketamine antidepressant properties. To further explore the possible interaction of ketamine with CB1R, we investigated the effects of an ineffective dose of the CB1R antagonist with a smaller dose of ketamine. Different from Khakpai et al’ observations, 39 we found that rimonabant pre-treatment did not potentiate ketamine effects in the FST. The discrepant results may come from experimental differences, such as the species used and the doses of rimonabant and ketamine administered.
Even though CB1R blockade elicits antidepressant-like effects per se, contradictory results in which the pharmacological blockade of these receptors did not promote an antidepressant-like effect have also been described.61,62 Furthermore, evidence indicates that CB1R blockade/silencing can attenuate or facilitate the behavioral effects of monoaminergic antidepressant drugs.63,64 Such discrepancies can reflect the lack of selectivity of the drugs used, the diversity of animal models, and the species and strain in which drugs have been tested. In this study, our data from pharmacological CB1R blockade and KO mice consistently suggest that CB1R plays a minimal role, if any, in the antidepressant effects of ketamine.
Our results were consistent in both male and female mice, with minimal differences between sexes, except for the increased immobility time in the KO mice on day 2. Corroborating our findings, other studies failed to detect changes in female mice treated with ketamine. 65 However, previous work demonstrated that female mice are more sensitive to ketamine than males, especially during the proestrus phase.66,67 Some of the differences between male and female mice are not detected if mice have previously been stressed, 67 which was the case in our study. Moreover, we did not investigate the influence of the estrous cycle on ketamine effects in female mice since this was not a primary outcome of our investigation. Thus, we cannot exclude that the different estrous cycle phases of the female mice might have interfered with our findings.
The results of this investigation have to be interpreted in light of its experimental limitations, for example, the lack of additional animal models to investigate ketamine-induced psychostimulant and antidepressant effects. Nevertheless, the FST remains one of the most investigated animal models to test the antidepressant effects of drugs and has excellent predictive validity. 68 Similarly, the locomotion in the open-field test has been a valuable experimental tool to describe the potentially psychostimulant properties of drugs. 50 A detailed evaluation of ketamine effects, including different treatment times and other animal models and species, is required to better understand the involvement of CB1R in the antidepressant and psychostimulant properties of ketamine.
In conclusion, our results support the hypothesis that CB1R mediates the hyperlocomotion effects induced by ketamine since the hyperactivity promoted by this drug in the open-field test was impaired by pharmacological and genetic inhibition of CB1R. Moreover, genetic CB1R deletion did not modify the antidepressant effect of ketamine in male mice. We also demonstrated that pharmacological blockade of CB1R induced an antidepressant-like effect per se in male and female mice, but it did not facilitate ketamine's antidepressant properties. These results advance our understanding of mechanisms underlying ketamine's side effects and might contribute to a safer clinical application of this drug in the future under concomitant administration of rimonabant or another CB1R antagonist to prevent its psychostimulant side effects. Furthermore, our results shed light on an essential aspect behind ketamine antidepressant effects by demonstrating that it can be dissociated from its psychostimulant properties.
Footnotes
Author Disclosure Statement
No competing financial interests exist.
Funding Information
S.J. received a productivity fellowship from Conselho Nacional de Desenvolvimento Científico e Tecnológico (CNPq; no. 304780/2018-9). This work was funded by Coordenação de Aperfeiçoamento de Pessoal de Nível Superior (CAPES; Finance Code 001), Sao Paulo State Research Foundation (FAPESP; no. 2017/24304-0), Aarhus University Research Foundation (AUFF-E-2020-7-1 9), Independent Research Fund Denmark (grant 8020-00310B), and Gill endowment.
Abbreviations Used
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.