
Abstract
Introduction:
Empathy is a fundamental prosocial behavior. It has been defined as perception, awareness, and understanding of others' emotional states, including painful processes. Mice living in pairs with conspecific chronic suffering from constriction injury exhibit pain hypersensitivity mediated by the amygdaloid complex. Nevertheless, the underlying mechanisms in the amygdala responsible for this response remain to be determined. This study investigated if the anxiolytic benzodiazepine midazolam (MDZ) and cannabidiol (CBD), a phytocannabinoid with multiple molecular targets, would attenuate this behavioral change. We also investigated if serotonergic and γ-aminobutyric acid (GABA)ergic mechanisms in the amygdala are involved in this effect.
Materials and Methods:
Male Swiss mice were housed in pairs for 28 days. The pairs were divided into two groups on the 14th day: cagemate nerve constriction and cagemate sham. On the 24th day, cagemates underwent a stereotaxic surgery and, on the 28th day, were evaluated on the writhing test.
Results:
The results showed that living with chronic pain leads to hypernociception in the cagemate and increases the expression of 5-HT3 receptor (5-HT3R) and glutamic acid decarboxylase 67 within the amygdala. MDZ (3.0 and 30 nmol) and CBD (30 and 60 nmol) attenuated the hypernociceptive behavior. The 5-HT3R antagonist ondansetron (0.3 nmol) prevented the antinociceptive effects of MDZ and CBD.
Conclusion:
These findings indicate that 5-HT3R and GABAergic mechanisms within the amygdala are involved in the pain hypersensitivity induced by the empathy for pain model. They also suggest that MDZ and CBD could be a new potential therapy to alleviate emotional pain disorders.
Introduction
The painful phenomenon is a subjective and multidimensional experience, which can be associated with a strong affective component. 1 Diverse social and environmental factors can influence the progress from acute to chronic pain. However, the underlying molecular mechanisms of this process are still poorly known.2,3
Emotional responses involved in painful situations in human beings are triggered not only in those who feel the pain but also in those experiencing potentially painful conditions. 4 When behavior signs of an unpleasant situation are identified, the observer can try to avoid or escape the aversive source (defensive behaviour) or help (prosocial behavior) those who are suffering. 5 The property of perception, awareness, and understanding of others' sensory and emotional states—including painful processes—is described as empathy. 6
Previous studies have shown that cohousing with a conspecific undergoing chronic pain or stressful situation induces hypernociception and increases anxiety-related responses.7,8 Although the neural basis of these responses is not completely clear, the amygdala stands out as an important candidate. The serotonergic system modulates this limbic area 9 and integrates the nociceptive pathways that may act in empathic responses. 10 Serotonin (5-HT) is involved in pain disorders 11 and empathic behaviors. 12 Recently, our group described an increase in the amygdala 5-HT turnover in the empathy for pain model. 13
Among the serotonergic receptor subtypes, the 5-HT3 receptor (5-HT3R) has been investigated for acting on sensory pain transmission. 14 5-HT3R, a member of the ion channel receptor family, is present in γ-aminobutyric acid (GABA) interneurons in the amygdala. 15 It modulates the release of several neurotransmitters. 16 It has been suggested that 5-HT3R could modulate the behavioral effects of cannabinoids 17 and drugs that act on the GABAA receptor (GABAAR) complex. 18 Natural and synthetic cannabinoid compounds could inhibit 5-HT3R through an allosteric mechanism. 19
In this study, we further investigated the association of 5-HT3R and GABA mechanisms with behavioral changes related to empathy to a chronic painful condition. We hypothesized that mice living with a conspecific suffering from chronic pain would present changes in amygdala expression of 5-HT3R and glutamic acid decarboxylase 67 (GAD67), the enzyme responsible for synthesizing GABA. 20 We also investigated whether the intra-amygdala administration of the anxiolytic benzodiazepine midazolam (MDZ) or the phytocannabinoid cannabidiol (CBD) would attenuate the hyperalgesic responses. Finally, we tested if the effects of these two drugs depend on 5-HT3R in the amygdala.
Materials and Methods
Subjects
For this study, 604 male Swiss mice at 21 days of age from the animal breeding facility of the Federal University of São Carlos were used. Animals were housed in pairs (cage size: 19 cm×30 cm×14 cm) with food and water ad libitum, except during the brief test periods, on a 12-h light/12-h dark cycle (7:00 am lights on) in a temperature-controlled (23°C±1°C) room. The experiments were carried out between 9:00 am and 4:00 pm. Different batches of experimentally naive mice were used for each experiment. All procedures were also approved by the Ethics Committee on the Use of Animals of the Federal University of São Carlos (Processes CEUA/UFSCar, approval numbers 8863220217 and 3305221018). All efforts were made to reduce animal suffering and to minimize the number of animals used.
Drugs
The following drugs were used for intra-amygdala drug injection procedure: MDZ (8-chloro-6-[2-fluorophenyl]-1-methyl-4H-imidazo[1,5a][1,4]hydrochloric benzodiazepine), Roche, Brazil, dissolved in physiological sterile saline (0.9% NaCl), at the doses of 3.0 and 30 nmol/0.1 μL; CBD 2-[(1R, 6R)-6-isopropenyl-3-methylcyclohex-2-en-1-yl]-5-pentylbenzene-1,3-diol, THC Pharm, Germany, dissolved in vehicle (sterile saline with 2% Tween 80), at the doses of 30 and 60 nmol/0.1 μL; and the 5-HT3R antagonist ondansetron {1,2,3,9-tetrahydro-9-methyl-3[(2-methyl-1H-midazol-1-yl)]-methyl-4H–carbazol-4-um}, Sigma-Aldrich, Brazil, dissolved in vehicle (physiological sterile saline with 2% Tween 80), at the dose 0.3 nmol/0.1 μL. All doses were based on previous studies.13,21,22 The total volume of injection within the amygdala was 0.1 μL per side.
Chronic constriction injury
The Bennett and Xie 23 method was used as a model of chronic constriction injury. After ketamine and xylazine anesthesia (100 and 10 mg.kg−1, respectively, intraperitoneally [i.p.]), the fascia between the gluteus and biceps femoral was sectioned, and the right sciatic nerve was exposed close to its trifurcation. The tissue around the nerve was carefully cut at ∼8 mm from this point, and the nerve was compressed with three ligatures using sterile noninflammatory mononylon 6.0 threads. The sham surgery was performed by exposing the sciatic nerve as described above without ligation. After surgery, the animals received ceftriaxone (ceftriaxone sodium hemipentahydrate, 4 mg.kg−1) 24 and were transferred to their home cages and left to recover.
Pain-related behaviors increase over the days after this procedure, and the peak is reached on the 14th day after constriction. 25 The hot plate test described below (see Experimental Design section) was used as a parameter to measure chronic nociception.23,26 Animals subjected to chronic sciatic nerve constriction show behavioral signs interpreted as spontaneous pain, including licking and limping the paw, besides avoiding putting weight on the injured side.
Surgery and microinjection
Bilateral stainless-steel guide cannula (25-gauge×7 mm; Insight Instruments, Brazil) were then implanted under ketamine and xylazine anesthesia (100 and 10 mg.kg−1, i.p.) in a stereotaxic frame (Insight Instruments). The guide cannula was fixed to the skull with dental acrylic and jeweller's screws. Stereotaxic coordinates for the target site in the amygdaloid complex were −1.3 mm posterior to bregma, ±3.3 mm lateral to the midline, and −2.8 mm ventral to the skull surface. 27 To prevent occlusion, each guide cannula was sealed with a dummy cannula (33-gauge stainless steel wire; Fishtex®, Brazil) to protect it from blockage at the time of surgery. During the stereotaxic surgery, animals received ketoprofen (benzene acetic acid, 5 mg.kg−1) and ceftriaxone (ceftriaxone sodium hemipentahydrate, 4 mg.kg−1). 24 Mice were allowed to recover for 4 days after the surgery before behavioral tests.
Solutions were injected into the amygdala by a microinjection needle (33-gauge stainless steel cannula; Insight Instruments) that extended 2 mm beyond the tip of the bilateral guide cannula. The microinjection needles were connected to a 10 μL Hamilton microsyringe by a polyethylene tube (PE-10). The flow rate was controlled by an infusion pump (BI 2000; Insight Instruments), programmed to deliver 0.1 μL of each solution throughout 60 sec. The microinjection procedure consisted of gently restraining the mice, inserting the injection needle, and infusing the solution for 60 sec with an additional 90 sec to maximize diffusion from the needle tip. The movement of a small air bubble in the PE-10 tube confirmed the solution's delivery.
Nociception test
Previously described by Vander Wende and Margolin, 28 the writhing test applies irritant substances in rodents' peritoneal cavity, allowing pain measurement by recording the number of writhes induced by this chemical stimulus. This stimulus causes an acute visceral tonic and diffuses painful sensation mediated by spinal and supraspinal sites, evoking different emotional responses. 29 In this study, writhes were induced by injecting 0.1 mL.10 g−1 body weight of 0.6% acetic acid, i.p., 5 min after the intra-amygdala drug injections. Five minutes later, the number of writhes was recorded.
Experimental design
In all experiments, the term “cagemates” refers to mice that cohabited in the same cage with animals subjected to sciatic nerve constriction or sham surgery. Twenty-one days after birth (weaning), the mice were housed in pairs to establish familiarity.7,30 Fourteen days later, the animals were divided into two groups, in which one animal of each pair was subjected to the sham surgery or the sciatic nerve constriction procedure. The other animal of the couple, which was left undisturbed in the cage, comprised the cagemate nerve constriction (CNC) and cagemate sham (CS) groups. For the experiments using intra-amygdala microinjections, the mice were submitted to a stereotaxic surgery on day 24.
The peak of the chronic constriction injury is reached on the 14th day after constriction. 25 Therefore, on the 28th day, CNC and CS groups received the treatments (systemic or intra-amygdala) and were subjected to the abdominal writhe test in the pair's presence (sham or constriction injury animal) for assessing the nociceptive behavior for 5 min. All sessions were recorded, and data were subsequently evaluated using the program X-Plo-Rat 2005 1.1.0. 31
The effectiveness of the nerve constriction was evaluated in the hot plate test. Animals were individually placed on a heated metal plate (52°C±0.2°C) until they reacted to the thermal stimulus (i.e., the latency to lift or lick their paws, a typical nociceptive response). The latency of removing the right hind paw after applying the heat was measured 26 using a hand-controlled timer. The cutting-off time in which the animal remained on the hot plate was 30 sec to avoid tissue damage. 32
In total, 302 mice were subjected to the chronic constriction injury/sham surgery, and 302 mice were subjected to cohabitation with a pair in the chronic pain/sham condition.
Histology
To control the microinjection sites, mice received an intra-amygdala 0.1 μL infusion of 1% Evans blue according to the previously described microinjection procedure. The animals were euthanized in a carbon dioxide chamber. Their brains were removed, stored in a formaldehyde solution (10%), and later coronally cut with a cryostat microtome (Leica CM 1850). Injection sites were microscopically (Olympus BX41) confirmed by the presence of Evans blue staining in the amygdala region according to the Paxinos and Franklin 27 mouse brain atlas. Data from animals with injection sites outside the amygdala were excluded from the study.
Brain dissection
Mice were transcardially perfused with 30 mL of 1×phosphate-buffered saline (PBS) at pH 7.4, followed by 50 mL of 4% fresh paraformaldehyde. The brain was dissected and transferred to a 30% sucrose solution in PBS for 48 h at 4°C. The brains were then frozen, and three series of 35 μm thickness sections of the amygdala were cut in a cryostat at −20°C (Leica CM 1850) with the help of the Paxinos and Franklin atlas. 27 Section triplicates were placed in serial order in a 12-well plate containing 0.1 M phosphate buffer (PB) with 0.01% sodium azide and processed for immunohistochemistry and immunofluorescence as described below.
Immunohistochemistry
Slices were washed thrice (10 min each) in PBS at room temperature on a shaker and placed into a blocking buffer (normal goat serum 3% and Triton X-100 0.25% in PBS) for 1 h at room temperature. After blocking, slices were incubated overnight with an anti-5-HT3 antibody raised in rabbit (Sigma-Aldrich, St. Louis, MO) dilution of 1:1000. Slices were again washed thrice and incubated in a solution of biotinylated anti-rabbit secondary antibody (Vector Laboratories, Burlingame, CA) at a dilution of 1:600 for 2 h and then placed in the mixed avidin-biotin horseradish peroxidase complex solution (ABC Elite Kit; Vector Laboratories) for 1 h. Immunoreactivity for 5-HT3 was revealed by adding the chromogen diaminobenzidine (0.02%; Sigma-Aldrich, Brazil) and hydrogen peroxide (0.04%), and was visualized as a brown insoluble reaction product inside the neuronal nucleus after 8 min.
The reaction was stopped by washing in PBS. Sections were mounted on gelatin-coated slides and then dehydrated and coverslipped with Permount (Sigma-Aldrich, Brazil). The 5-HT3 immunoreactive neurons in the amygdala were counted using an optic microscopic (Zeiss Axioskop 2) coupled with a camera and the Axio Vision Rel. Four images were captured from each mouse, and the ImageJ software (NHI) was used to quantify 5-HT3R-expression. 33
Immunofluorescence
Sections were washed thrice in 0.1 M PB and then incubated in a blocking solution containing 10% normal goat serum and 0.3% Triton-X 100 in 0.1 M PB for 1 h at room temperature with gentle rocking. Sections were incubated overnight with the primary antibody previously diluted in a blocking solution. After blocking, slices were incubated overnight with an anti-GAD67 antibody [K-87] (100 μg; primary antibodies, mouse; Abcam, Ref. no. ab26116). Sections were washed five times in 0.1 M PB and then incubated for 2 h at room temperature with secondary antibody Goat Anti-Mouse IgG H&L (Alexa Fluor® 568; ab175473). Following secondary incubation, sections were washed five times in 0.1 M PB, mounted onto glass slides, coverslipped using Fluoroshield Mounting Medium, and sealed with nail polish, once cured.
The images from each slide were acquired using a fluorescence microscope (Axio Imager.D2; Carl Zeiss Microscopy, LLC, Thornwood-NY) connected Zen Pro 2.0 software (Carl Zeiss Microscopy, LLC), and were analyzed using ImageJ software (NIH). Background staining was subtracted from each image before image analysis. The corrected total cellular fluorescence (corrected total cellular fluorescence=integrated density − [area of selected tissue area×mean fluorescence of background readings]) of the amygdala was measured by subtracting the background fluorescence from the integrated intensity and performed as described previously.34,35
Data and statistical analysis
The analyses were performed using Statistica 8 (TIBCO Software, Inc.). All data were initially checked for homogeneity of variance (Levene's test). For Experiment 1 (5-HT3R and GAD67 expression) and hot plate, data were analyzed by the Student's t-test. In Experiments 2 and 3, data were analyzed by two-way analysis of variance (ANOVA; cohabitating type×treatment). Data from Experiments 4 and 5 were subjected to a three-factor ANOVA (cohabitating type×pretreatment×treatment). In case of significant ANOVA effects, Duncan's multiple comparison test was employed for post hoc comparisons. Values of p≤0.05 were regarded as significant.
Results
Hot plate test: assessing the effectiveness of nerve constriction surgery
Student's t-test indicated that the chronic constriction injury procedure increased pain response in all experiments: Experiment 1 [t(13)=9.21, p<0.05], Experiment 2 [t(48)=11.19, p<0.05], Experiment 3 [t(63)=10.40, p<0.05], Experiment 4 [t(34)=10.11, p<0.05], and Experiment 5 [t(54)=10.90, p<0.05]. Table 1 represents the effect on spontaneous pain after 14 days of sciatic nerve constriction. For this procedure, 222 mice were used (Table 2).
Evaluation of Spontaneous Pain Assessed 14 Days After Sciatic Nerve Constriction
Latencies (in sec) of pain response in the hot plate test are presented as mean±SEM.
p<0.05 versus respective Sham control group.
SEM, standard error of mean.
Sample Sizes for each Experimental Group
The numbers represent the sample size used per group. Drug doses are represented in nmol/0.1 μL. Vehicle or saline groups received a volume of 0.1 μL per hemisphere. The sham and constriction injury animals did not receive pharmacological treatment.
5-HT3R, 5-HT3 receptor; CBD, cannabidiol; CNC, cagemate nerve constriction; CS, cagemate sham; GAD67, glutamic acid decarboxylase; MDZ, midazolam; OND, ondansetron.
Histology
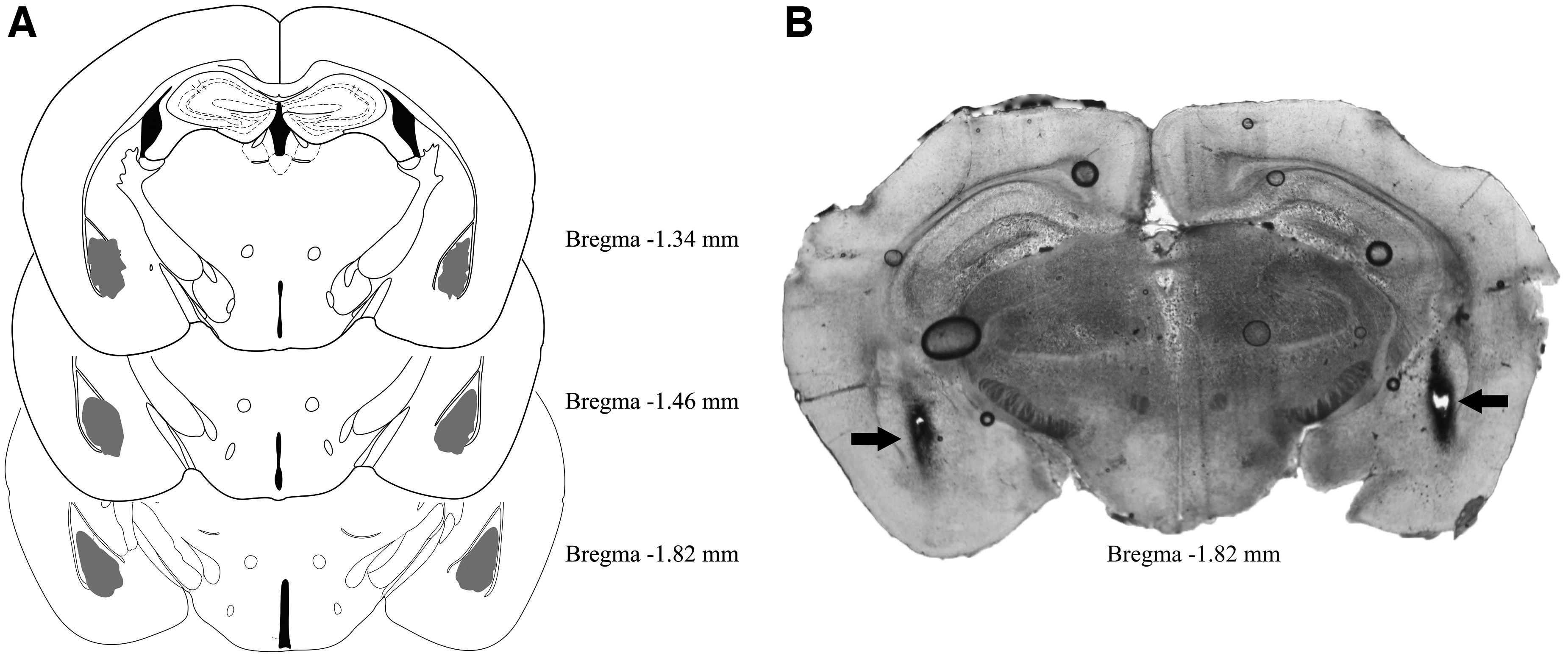
Diagrammatic representations showing the bilateral injection sites in the amygdala and an illustrative photomicrograph of these sites are presented in Figure 1. The number of animals included in the study after the histological analysis of the injection sites was described in Table 2.
Experiment 1: cohabitation with mice subjected to chronic constriction injury increases 5-HT3R and GAD67 expression in the amygdala
To investigate 5-HT3R expression in the amygdala (see Fig. 2A, for experimental procedure), 15 animals were used (seven CS and eight CNC). Figure 2B shows coronal brain slices of two representative subjects that cohabited in pairs with CS or CNC animals. Student's t-test indicated that 5-HT3R expression increased in CNC compared to CS mice [t(13)=12.92, p<0.05] (Fig. 2C).

Ten mice were used to investigate the GAD67 expression in the amygdala (see Fig. 2A) (five CS and five CNC). Figure 2D shows coronal brain slices of two representative mice that cohabited in pairs with CS or CNC animals. Student's t-test indicated that GAD67 expression increased in the amygdaloid complex of CNC compared to CS animals [t(8)=−2.44, p<0.05] (Fig. 2E).
Experiment 2: intra-amygdala MDZ attenuates pain hypersensitivity in animals that cohabited in pairs with mice subjected to chronic constriction injury
In Experiment 2 (see Fig. 3A, for experimental procedure), 50 animals (25 CS and 25 CNC) were used to investigate the effects of MDZ injected into the amygdala on hypernociception induced by cohabitation with a partner suffering chronic pain. Two-way ANOVA indicated significant differences for cohabitation [CS and CNC: F(1,44)=4.35, p<0.05] and treatment [F(2,44)=14.11, p<0.05] factors, and for cohabitation versus treatment interaction [F(2,44)=9.46, p<0.05]. The Duncan test revealed an increase in the number of writhes in the CNC-saline compared to CS-saline (p=0.0001). MDZ (3.0 and 30.0 nmol) decreased the number of writhes in CNC compared to the CNC-saline animals. None of the MDZ doses changed the number of writhes in CS mice (Fig. 3B).
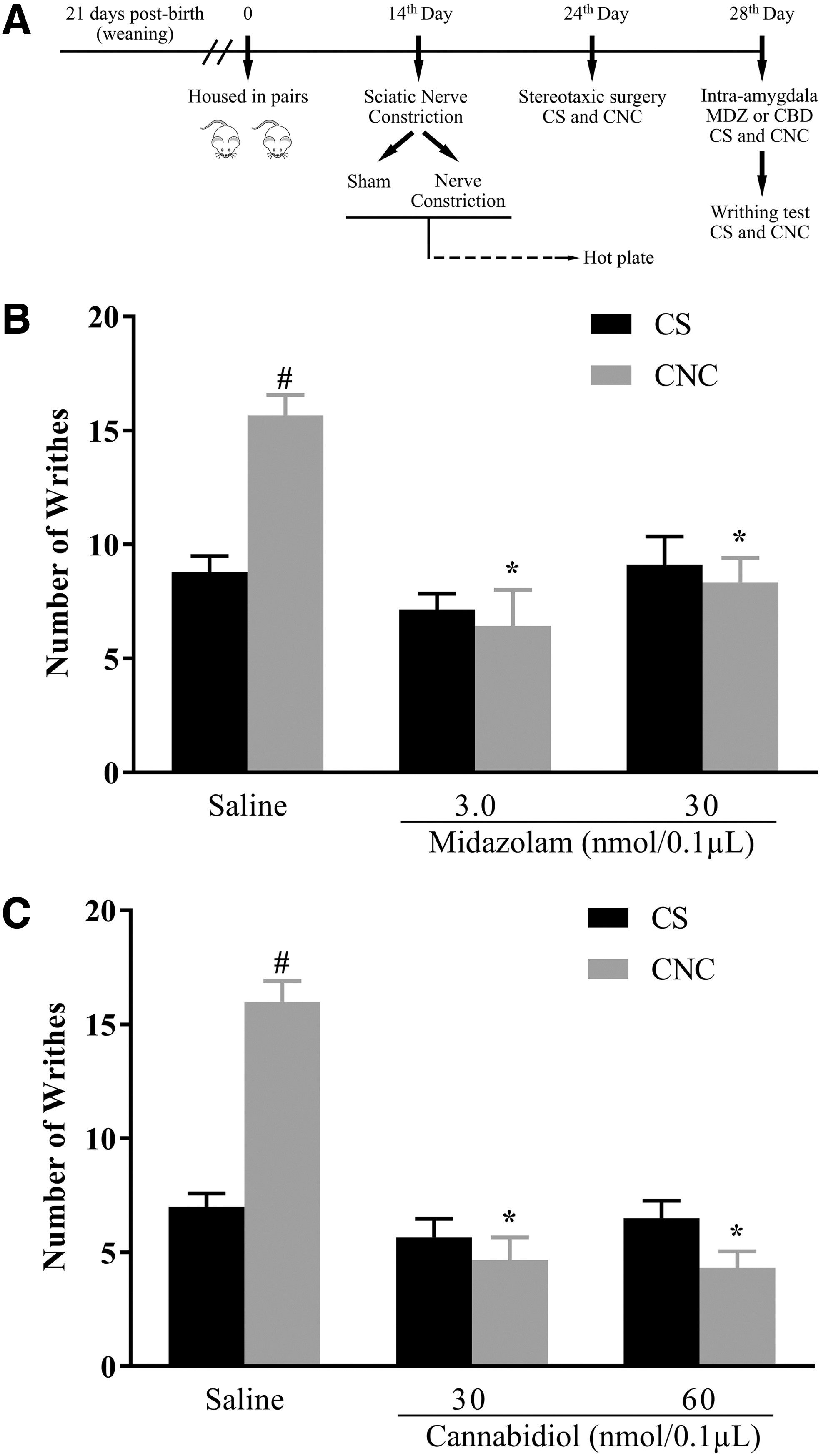
Experiment 3: intra-amygdala CBD attenuates pain hypersensitivity in animals that cohabited in pairs with mice subjected to chronic constriction injury
In Experiment 3 (see Fig. 3A, for experimental procedure), 36 animals (18 CS and 18 CNC) were used to investigate the effects of CBD injected into the amygdala on hypernociception induced by cohabitation with a pair suffering chronic pain. Two-way ANOVA indicated significant differences for cohabitation [CS and CNC: F(1,30)=8.83, p<0.05] and treatment [F(2,30)=40.09, p<0.05] factors, and for cohabitation versus treatment interaction [F(2,30)=29.35, p<0.05]. Post hoc test revealed an increase in the number of writhes in vehicle-treated CNC mice compared to vehicle-treated CS animals (p=0.0001). Moreover, both doses of CBD (30 or 60 nmol) reduced the number of writhes in CNC mice compared to the respective CNC mice that received vehicle (p=0.00003 and p=0.00002, respectively). None of the CBD doses interfered with the number of writhes in CS animals (Fig. 3C).
Experiment 4: ondansetron injected into the amygdala impairs the antihypernociceptive effect induced by local injection of MDZ in animals that cohabited with a pair subjected to chronic constriction injury
In Experiment 4 (see Fig. 4A, for experimental procedure), 65 mice (33 CS and 32 CNC) were used to investigate the effects of ondansetron pretreatment followed by MDZ (treatment) within the amygdala on the hypernociception induced by cohabitation with a pair suffering from chronic pain. Three-way ANOVA indicated significant differences for cohabitation [CS and CNC: F(1,57)=21.56, p<0.05], and treatment [F(1,57)=12.77, p<0.05] factors, and for cohabitation×pretreatment×treatment interactions [F(1,57)=4.09, p<0.05], without revealing significant differences for pretreatment factor [F(1,57)=1.89, p>0.05]. Duncan's post hoc test showed an increase in the number of writhes in the vehicle/saline CNC mice compared to vehicle/saline CS mice (p=0.0003).

Similar increases were revealed in the ondansetron/saline CNC and ondansetron/MDZ CNC mice compared to ondansetron/saline CS (p=0.0057) and ondansetron/MDZ CS mice (p=0.015), respectively. MDZ (3.0 nmol) decreased the number of writhes in the vehicle/MDZ CNC compared to the vehicle/saline CNC group (p=0.00002). Moreover, the number of writhes of the ondansetron/MDZ CNC group was higher than that exhibited by the vehicle/MDZ CNC mice (p=0.0023), indicating that ondansetron pretreatment blocked the MDZ effect. None of the treatments significantly changed the number of writhes in the CS animals (Fig. 4B).
Experiment 5: ondansetron injected into the amygdala impairs the antihypernociceptive effect induced by local injection of CBD in animals that cohabited with a pair subjected to chronic constriction injury
In Experiment 5 (see Fig. 4A, for experimental procedure), 56 animals (27 CS and 29 CNC) were used to investigate the effects of ondansetron pretreatment followed by CBD (treatment) within the amygdala on the hypernociception induced by cohabitation with a pair suffering from chronic pain. Three-way ANOVA indicated significant differences for cohabitation [CS and CNC: F(1,48)=12.25, p<0.05] and treatment [F(1,48)=16.50, p<0.05] factors, and for cohabitation×pretreatment×treatment interactions [F(1,48)=10.96, p<0.05], without revealing significant differences for pretreatment factor [F(1,48)=3.61, p>0.05]. Duncan's post hoc test revealed more writhes in the vehicle/vehicle CNC mice than vehicle/vehicle CS mice (p=0.0003).
Similar increases were shown in the ondansetron/vehicle CNC and ondansetron/CBD CNC mice compared to ondansetron/vehicle CS (p=0.015) and ondansetron/CBD CS mice (p=0.019), respectively. Post hoc analysis also revealed that CBD (30 nmol) decreased the number of writhes in the vehicle/CBD CNC mice compared to the vehicle/vehicle CNC animals (p=0.00001). Moreover, the number of writhes recorded for the ondansetron/CBD CNC group was higher than for the vehicle/CBD CNC mice (p=0.00004), indicating that ondansetron pretreatment abolished the CBD effect. None of the treatments interfered with the number of writhes in the CS animals (Fig. 4C).
Discussion
This study investigated neural mechanisms in the amygdaloid complex involved in pain hypersensitivity induced by living with a conspecific subjected to chronic nerve constriction injury. In this study, we demonstrated that cohousing with a pair suffering from chronic pain enhanced 5-HT3R and GAD67 expression in the amygdala. Furthermore, we demonstrated that MDZ and CBD administered directly into the amygdala attenuated the hypernociceptive behavior in cagemates after cohabitation with conspecific suffering pain. Moreover, intra-amygdala injection of ondansetron before local injection of MDZ or CBD impaired the antihypernociceptive effect of benzodiazepine and cannabinoid compounds.
Our results corroborate previous findings, demonstrating that living with a mouse subjected to nerve constriction injury induces pain hypersensitivity in cagemates.7,36 Different research fronts have been developed to clarify the underlying physiological and neurochemical mechanisms involved in the behavioral responses related to empathy. Serotonin dysfunction joint to empathy deficits were demonstrated through intra-ACC serotonin microinjection led to reductions in observational fear learning. 37 In that wise, 5-HT is related to persistent pain mainly by acting through 5-HT3R, a receptor with a relevant role in acute and neuropathic pain. 38
Corroborating this proposal, in this work, we showed that 5-HT3R expression increases in the amygdala of mice that cohabit with animals subjected to nerve constriction injury. Sustained nociceptive stimulus increases the expression of 5-HT3R, leading to neuronal death of inhibitory interneurons, which contribute to the development of pain hypersensitivity. 39 Considering that the pain empathy model also induces pain hypersensitivity, it is possible that a similar mechanism could be activated in mice that cohabit with a partner suffering from chronic pain. The emotional responses related to pain could promote overexpression of 5-HT3R in the amygdala, facilitating the hypernociceptive responses.
GABA synthesis involves two GAD isoforms, GAD65, present in axon terminals and microvesicles, and GAD67, preferentially located in the cytoplasm, engaged in nonvesicular GABA release. 40 This enzyme may be involved in the GABA release from glial cells by reversing transporters or other nonvesicular mechanisms.20,41 Previous findings have demonstrated that GAD67 expression increases in the central and medial amygdaloid nuclei, the bed nucleus of the stria terminalis, and the hippocampus after exposure to chronic stress.42,43 In this context, we suggest that the 5-HT3R, which are overexpressed in the amygdala of CNC mice, might be colocalized within GABAergic interneurons and when activated, would increase GABA inhibitory neurotransmission.44,45
A positive modulator of the GABAergic neurotransmission, MDZ, decreased pain hypersensitivity in CNC animals when microinjected into the amygdala. This result agrees with previous reports from our group showing that systemically injected MDZ modulates the pain response in mice exposed to aversive situations, 21 including the empathy for pain model. This treatment impaired the pain hypersensitivity and anxiety induced by this model.36,46 Furthermore, recent evidence has shown that GABAergic neurons in the medial amygdala have important roles in social behavior. 47 Together, these results suggest that the GABAergic neurotransmission in the amygdaloid complex modulates the emotional consequences of cohabitating with a conspecific suffering from chronic pain. Moreover, altogether our molecular and pharmacological approaches indicate that the overexpression of GAD67 in the amygdaloid complex could be interpreted as a compensatory mechanism to attenuate pain hypersensitivity.
5-HT3R and GABAAR belong to the same family of ion-channel receptors.15,44 Pretreatment with ondansetron, a selective and 5-HT3R competitive antagonist, 48 at a dose that did not affect nociception, 13 prevented the antinociceptive effect produced by MDZ (Experiment 4). The activation and blockade of 5-HT3R in GABA interneurons present in the amygdala increase and reduce, respectively, GABAergic neurotransmission.44,45 In addition, 5-HT3R antagonists block the antinociceptive effect of GABAergic agents. 38 Corroborating these results, our data support the proposal that GABA and 5-HT mechanisms interact in the amygdala to modulate empathy for pain.
Similar to MDZ, intra-amygdala CBD attenuated pain hypersensitivity in animals that have been cohoused with mice suffering from constriction injury. CBD effects on emotional responses have been shown after systemic and intra-amygdala administration of this cannabinoid.49,50 Moreover, Genaro et al 51 showed that CBD injected into the anterior cingulate cortex reduced mechanical allodynia, suggesting that it could affect sensory and affective dimensions of pain. The mechanisms responsible for these effects are still poorly understood.
The endocannabinoid (eCB) signaling is a promising target for pain relief. 52 The several stages of the eCB, including its synthesis pathways, interaction with receptors, and degradation by enzymes, have been studied as a way of elucidating the potential for providing analgesia.52,53 CBD has a low affinity for CB1 receptors (Ki=4350 μM), 54 but can increase anandamide levels by inhibiting the fatty acid amide hydrolase enzyme. 55 However, several other mechanisms have been associated with CBD and anandamide actions, 56 including modulation of 5-HT3R.56,57 Adding to the complex pharmacology of CBD, our results show that it could also involve mechanisms that result in interfering with the serotoninergic neurotransmission within the amygdala.
The antihypernociceptive effect provoked by the cannabinoid compound in this study was blocked by the intra-amygdala injection of ondansetron. Previous studies have shown that CBD is a negative allosteric modulator of 5-HT3R. 56 Furthermore, the inhibitory allosteric interaction leading to conformational changes in 5-HT3R was proposed to be the primary mechanism of action of cannabinoid compounds.19,56,57 In this study, ondansetron—a selective and competitive antagonist of 5-HT3R 48 —impaired the CBD's antinociceptive response, probably by blocking the orthosteric site and thus remodeling the receptor's conformation, preventing CBD to bind to its allosteric site. Again, although attractive, this hypothesis needs to be clarified by further experiments.
Conclusion
In conclusion, these results indicate that 5-HT3R- and GABA-mediated neurotransmissions within the amygdala are involved in the pain hypersensitivity induced by the empathy for pain model. They also suggest that CBD and MDZ could be a new potential therapy to alleviate emotionally associated pain disorders. Summarizing it is relevant to highlight the adversity to find effective pain treatments due to the complexity of pain perception within chronic pain conditions. 58 In this sense, these findings sought to clarify the interaction of distinct neurotransmission systems located in the amygdaloid complex in the modulation of pain processing in mice under the influence of strong emotional conditions.
Footnotes
Authors' Contributions
L.R.R.T. and A.C.S. conceived the project. L.R.R.T., D.B.S., and L.C.S. carried out the experiments, data analysis, and article drafting. C.S.P. and F.S.G. edited and revised the article. R.L.N.S. and A.C.S. contributed to the formulation and supervision of the experiments, and approved the article's final version.
Author Disclosure Statement
F.S.G. is a co-inventor of the patent “Fluorinated CBD compounds, compositions, and uses thereof. Pub. No.: WO/2014/108899. International Application No.: PCT/IL2014/050023” Def. US no. Reg. 62193296; 29/07/2015; INPI on 19/08/2015 (BR1120150164927). The University of São Paulo has licensed the patent to Phytecs Pharm (USP Resolution No. 15.1.130002.1.1). The University of São Paulo has an agreement with Prati-Donaduzzi (Toledo, Brazil) to “develop a pharmaceutical product containing synthetic CBD and prove its safety and therapeutic efficacy in the treatment of epilepsy, schizophrenia, Parkinson's disease, and anxiety disorders.” The other authors declare no conflict of interests.
Funding Information
The experiments described in this article were supported by CNPq (National Council for Scientific and Technical Development, 482356/2013-8), Brazil. L.R.R.T. received a scholarship from CNPq, Brazil (153163/2016-0). CAPES Brazil financing code 001. D.B.S. received a scholarship from FAPESP, São Paulo, Brazil (2015/0004-6). L.C.S. received a scholarship from FAPESP, São Paulo, Brazil (2016/08665-0). C.S.P. (306557/2015-0), F.S.G. (300940/2018-1), A.C.S., Brazil (309201/2015-2), and R.L.N.S. (306556/2015-4) were the recipients of CNPq research fellowships.