
Abstract
Introduction:
Cannabis is the most used illicit drug in the United States. With many states passing legislation to permit its recreational use, there is concern that cannabis use among adolescents could increase dramatically in the coming years. Historically, it has been difficult to model real-world cannabis use to investigate the causal relationship between cannabis use in adolescence and behavioral and neurobiological effects in adulthood.
Materials and Methods:
We used a response-contingent vapor administration model to investigate long-term effects of cannabis use during adolescence on the medial prefrontal cortex (mPFC) and mPFC-dependent behaviors in male and female rats.
Results:
Adolescent (35- to 55-day-old) female rats had significantly higher rates of responding for vaporized Δ9-tetrahydrocannabinol (THC)-dominant cannabis extract (CANTHC) compared with adolescent males. In adulthood (70–110 days old), female, but not male, CANTHC rats also took more trials to reach criterion and made more regressive errors in an automated attentional set-shifting task compared with vehicle rats, thereby indicating sex differences in behavioral flexibility impairments. Notably, sex–treatment interactions were not observed when rats of each sex were exposed to a noncontingent CANTHC vapor dosing regimen that approximated CANTHC vapor deliveries earned by females. No differences were observed in effort-based decision making in either sex. In the mPFC, female (but not male) CANTHC rats displayed more reactive microglia with no changes in myelin basic protein expression or dendritic spine density.
Conclusion:
Altogether, these data reveal important sex differences in rates of responding for CANTHC vapor in adolescence that may confer enduring alterations to mPFC structure and function and suggest that there may be subtle differences in the effects of response-contingent versus noncontingent cannabis exposure that should be systematically examined in future studies.
Introduction
Cannabis is the most widely used illicit substance among adolescents and the prevalence of heavy cannabis use during adolescence has seen a three-fold increase over the past 25 years. In the United States, vaping of high-potency cannabis concentrates during adolescence has increased dramatically, from 24.2% in 2017 to 52.1% in 2019 among past 30-day cannabis users.1,2 This is concerning because adolescence is a critical period of cortical development, 3 for which the endogenous cannabinoid system plays a prominent role 4 —thus, cannabis could interfere with these important neurodevelopmental processes, potentially increasing the risk for cognitive dysfunction later in life. 5
Adolescent cannabis use in humans is associated with deficits in prefrontal cortex (PFC)-dependent functions, such as attention, visuospatial functioning, and behavioral flexibility, which are all necessary for optimizing goal-directed behavior.6–9 For instance, adolescent-onset cannabis users show impairments in their ability to shift strategies in the Wisconsin Card Sorting task, which is a test commonly used to assess behavioral flexibility. 8 Moreover, the earlier participants began using cannabis, the more difficulty they had learning new strategies. 8
Poor behavioral flexibility is associated with neuropsychiatric disorders that are common to cannabis users, such as schizophrenia and major depression.10–12 Thus, a better understanding of the impact of adolescent cannabis use on behavioral flexibility could provide valuable insight into this link and help to identify mechanisms underlying increased vulnerability among cannabis users. These deficits could be related to willingness to expend effort, as both acute and chronic cannabis use have been associated with apathy, amotivation, and other reward processing deficits.13,14
Adolescent cannabis use also has been associated with changes in cortical structure, including decreased white matter, which may contribute to long-term cognitive deficits.15,16 These data suggest that there are relationships between adolescent cannabis use and cognitive- and brain-related endpoints. However, human studies are not well suited to interrogate cause–effect relationships and as such, the long-term consequences of adolescent cannabis use remain elusive.
Preclinical studies using rodent models mirror data from human studies, demonstrating long-term cognitive impairments following injections of synthetic cannabinoid CB1 receptor agonists17–19 or isolated cannabis constituents such as Δ 9 -tetrahydrocannabinol (THC).20–25 Of note, behavioral flexibility is impaired in adult male rats with a history of synthetic cannabinoid agonist injections during adolescence, 17 while THC injections in adolescent female rats produce long-lasting cognitive dysfunction in adulthood, which is associated with increased microglial activation in the PFC. 24
Microglia play an important role in the pruning of synapses in the PFC during adolescence, 26 which critically contributes to maturation of PFC-mediated cognitive functions. 27 Thus, cognitive deficits observed following adolescent cannabis exposure may be caused by cannabis-induced activation of microglia in the PFC. However, it remains unknown whether these findings using injection models generalize to human cannabis users where cannabis is typically consumed through intrapulmonary routes of administration. 28
Given the established pharmacokinetic and behavioral differences observed using different routes of cannabinoid administration, 29 it remains possible that data generated from bolus injections of isolated cannabis constituents or synthetic CB1 receptor agonists may not directly translate to human populations. 30 Accordingly, comparisons of injected versus vaporized THC have revealed markedly different pharmacokinetic profiles despite producing equal peak concentrations of circulating THC. 29 Specifically, higher initial concentration of THC is observed in the brain following vaporized THC exposure, whereas THC injection results in dramatically higher 11-OH-THC concentration, a potent THC metabolite, in both blood and brain that continues to increase over time. 29 Moreover, whole-plant cannabis extracts contain multiple phytocannabinoids that may interact to produce different behavioral effects.31,32 To address this translational gap, we recently developed a more translational model of cannabis use in rats.
This approach more accurately mimics the human experience of cannabis use by allowing response-contingent administration of vaporized whole-plant cannabis extracts using commercially available e-cigarette technology. 33 This model builds on seminal work conducted by other groups that have employed noncontingent isolated THC vapor delivery,34–40 or volitional intake of vaporized drugs like fentanyl and sufentanil,41–43 heroin, 44 nicotine,45–49 and nicotine with flavor additives50,51—however, we are the first to use response-contingent delivery of whole-plant cannabis extracts.33,52 We leverage this model in the current study to compare whole-plant cannabis extracts dominant in THC or cannabidiol (CBD), which are becoming increasingly common in today's market.
Specifically, we used this approach to determine whether response-contingent cannabis vapor delivery during adolescence impairs behavioral flexibility and recapitulates the long-lasting effects of injected cannabinoids on medial PFC (mPFC) microglia activation in adulthood. Moreover, we tested whether adolescent cannabis vapor administration impacts motivation for reinforcement using an effort-based decision-making test. Finally, we tested whether the phytocannabinoid content of whole-plant cannabis extract influenced the long-lasting effects on executive functioning and prefrontal cortical structure by comparing THC-dominant and CBD-dominant vaporized cannabis extracts to vehicle (VEH) vapor. Importantly, and in contrast to much of the extant literature, we used rats of both sexes to allow us to detect potential sex differences in adolescent cannabis use.
Methods
Animals
Male and female Sprague-Dawley rats obtained from Simonsen Laboratories (Gilroy, CA; Exp. 1) or Envigo (Indianapolis, IN; Exp. 2) on postnatal day (PND) 25 were housed in our climate-controlled vivarium on a 12:12-h reverse light cycle (lights off at 7:00) with food and water available ad libitum. After habituation and handling (Supplementary Data), rats underwent experimental procedures as shown in Figure 1. All experimental procedures adhered to the National Institutes of Health Guide for the Care and Use of Laboratory animals and were approved by the Washington State University Institutional Animal Care and Use Committee.
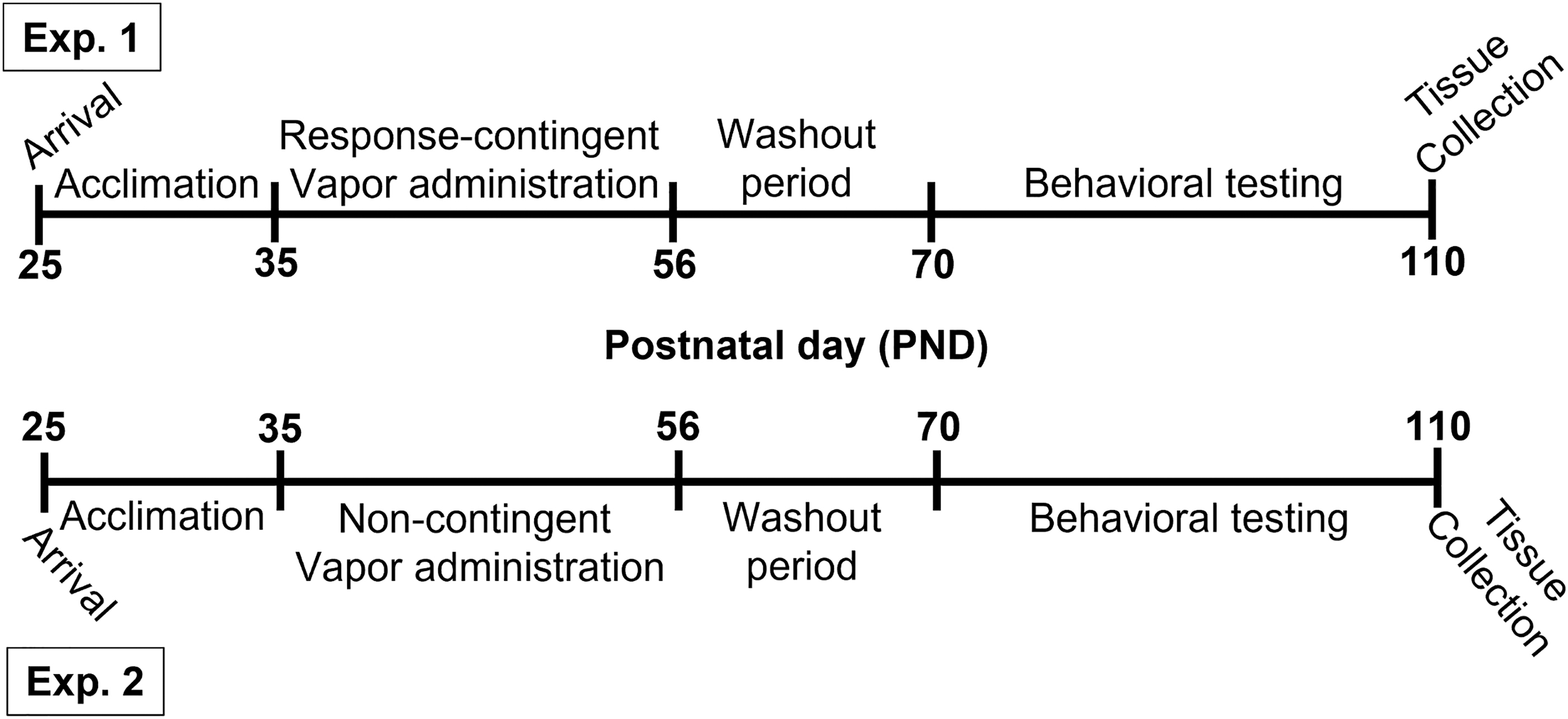
Experimental timeline. Sprague-Dawley rats of both sexes arrived at our facility on PND 25. After acclimation and handling, rats underwent daily sessions from PND 35 to PND 55 during which they received response-contingent (Exp. 1) or noncontingent (Exp. 2) vapor from cannabis extracts with high concentrations of THC (CANTHC), CBD (CANCBD), or vehicle (VEH). Rats were given a 2-week washout period (PND 56–69) before tests of behavioral flexibility (Exp. 1 and 2) and effort discounting (Exp. 1) in adulthood (PND 70–110). Rats were maintained at 90% free-feeding weight during behavioral testing. Brains were collected ∼24 h after completion of behavioral testing to assess changes in the mPFC. CBD, cannabidiol; mPFC, medial prefrontal cortex; PND, postnatal day; THC, Δ 9 -tetrahydrocannabinol.
Drugs
Natural cannabis extracts that contained high concentrations of THC (CANTHC) or CBD (CANCBD) were acquired from the National Institute on Drug Abuse (NIDA) Drug Supply Program. The certificates of analysis provided by NIDA indicated that the CANTHC extract contained 28.4% THC, 1.38% CBD, and 1.8% cannabinol, whereas the CANCBD extract contained 1.16% THC, 59.34% CBD, 2.1% cannabichromene, 1.1% cannabigerol, and <0.01% tetrahydrocannabivarin and cannabinol. Based on previous research, 26 CANTHC and CANCBD solutions were prepared at a 400 mg/mL concentration by dissolving raw cannabis extract into a VEH of 80% propylene glycol/20% vegetable glycerol under continuous stirring at 60°C as described previously. 33
Vapor administration
Experiment 1: response-contingent vapor administration
Response-contingent cannabis vapor administration was conducted using second-generation vapor chambers from La Jolla Alcohol Research, Inc. ([LJARI] La Jolla, CA) equipped with commercial e-cigarette tanks (SMOK Tank TFV8 with M2 atomizers), two nosepoke ports, and corresponding cue lights controlled by MED Associates IV software (Fairfax, VT) as previously described. 33 Rats were trained to respond for deliveries of CANTHC, CANCBD, or VEH vapor preparations on a fixed ratio 1 schedule from PND 35 to PND 55. During these 1 h sessions, a response made on the active port resulted in 3 s delivery of vapor and simultaneous illumination of a cue light located above the active port. The cue light remained illuminated for a 60 s time-out period during which active responses were recorded but had no consequence. Responses made on the inactive port were recorded but had no consequences.
A discrimination index for the active port was calculated as
Experiment 2: noncontingent vapor exposure
A separate cohort of adolescent male (n=32) and female (n=32) Sprague-Dawley rats received 21 daily sessions of noncontingent vehicle or CANTHC vapor deliveries from P35 to P55. One 3-s puff of vapor was given once every 2 min, and the total number of vapor deliveries each session was equal to the mean vehicle or CANTHC vapor deliveries earned by the adolescent females during the response-contingent vapor session (Exp. 1) as shown in Figure 2A. Thus, vehicle-exposed rats received the following number of deliveries across the 21 sessions: 13, 10, 10, 11, 9, 10, 11, 11, 13, 13, 13, 12, 12, 14, 15, 16, 16, 13, 15, 15, and 15; while CANTHC-exposed rats received 11, 8, 9, 9, 8, 11, 12, 12, 11, 13, 14,16, 16, 15, 15, 18, 18, 15, 14, 13, and 16.
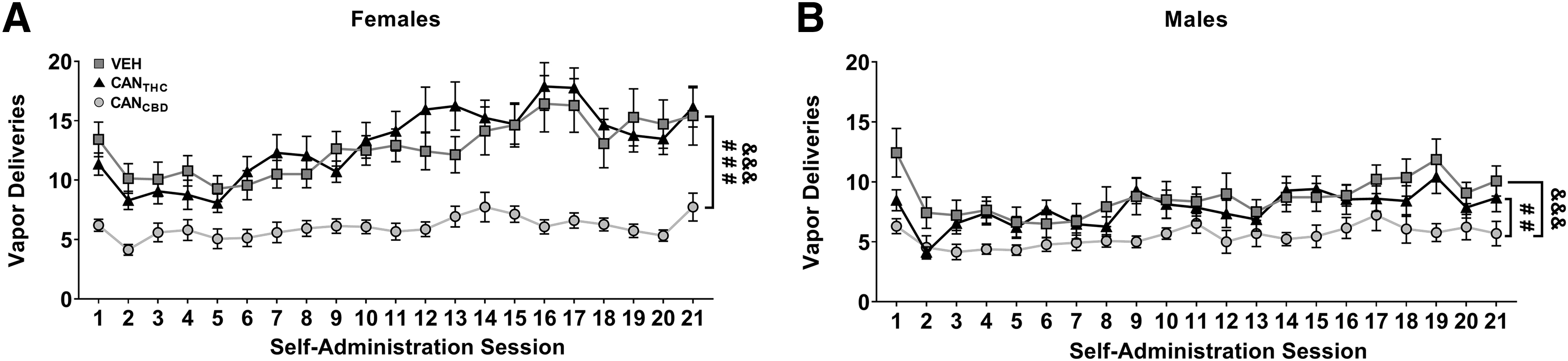
Adolescent rats of both sexes earn vapor deliveries of cannabis extracts with high concentrations of THC (CANTHC), CBD (CANCBD), or vehicle (VEH). Females
Behavioral testing
Beginning on PND 70, tests of attentional set-shifting (Exp. 1 and 2) and effort-based decision making (Exp. 1) were conducted in 10″×12″×12″ (L×W×H) Coulbourn Habitest chambers (Coulbourn Instruments, Holliston, MA) using Graphic State 4 software.54,55 Briefly, rats underwent lever shaping and retractable lever training until ≤5 omissions were made for 2 consecutive sessions, followed by visual cue discrimination, set-shifting, and reversal learning test phases to assess domains of behavioral flexibility as described previously. 56 In Exp. 1, rats were then retrained on retractable lever training and effort discounting was assessed. 57 See Supplementary Data for a complete description of behavioral testing procedures.
Immunofluorescence for mPFC myelination and microglial morphology
Approximately 24 h after completion of behavioral testing (Exp. 1 and 2), brains were collected following transcardial perfusion, postfixed in 4% paraformaldehyde overnight, sunk in 30% sucrose solution, and stored in 1×phosphate-buffered saline + sodium azide until needed. Brains from a subset of animals in Exp. 1 (n=6/group such that mean vapor deliveries were roughly equal between groups) were stained for myelin basic protein (MBP) and ionized calcium-binding adaptor molecule 1 (Iba1) to assess myelination and microglial morphology, respectively. For MBP analysis, mean gray value from four different slices containing the mPFC for each animal was quantified using ImageJ software (NIH, Bethesda, MD). For Iba1 staining of microglia, four parameters were analyzed as described previously 58 : cell surface area, cell perimeter, soma area, and roundness of the soma.
Since microglia are known to mediate synaptic pruning, another subset of rats was used to assess effects of response-contingent vapor administration during adolescence on dendritic spines in the mPFC (Supplementary Data). Brains from a subset of males in Exp. 2 (the second cohort, n=7–8/group) were collected and Iba1 fluorescence assessed.
Statistical analyses
The alpha level was set to p<0.05, and dependent measures were assessed using ANOVAs and linear mixed models (where appropriate). For response-contingent vapor analyses (Exp. 1) and all analyses conducted following noncontingent vapor exposure (Exp. 2), the between-subjects factors were sex and treatment. For endpoints tested after response-contingent vapor session, analyses were stratified by sex and treatment was the between-subjects factor. Significant main effects of treatment were further examined using Dunnett's post hoc tests with vehicle vapor as the control. All other significant main effects or interactions were followed up with Tukey's post hoc tests. Effect size estimates were calculated using Cohen's d. All statistical analyses were conducted in SAS (version 9.4; SAS Institute, Inc., Cary, NC) using PROC GLIMMIX.
Results
Experiment 1: response-contingent vapor administration
Adolescent female rats earn more THC- (but not CBD-) dominant cannabis vapor than male rats
Adolescent rats of both sexes earned a similar number of CANTHC and vehicle vapor deliveries (treatment by sex: F2,82=10.24, p=0.0001, Fig. 2A, B), and earned more vehicle than CANCBD (females: p<0.0001, d=1.33; males: p=0.0003, d=0.77). Females earned more vehicle (p<0.0001, d=0.68) and CANTHC (p<0.0001, d=0.95) vapor deliveries than their male counterparts. Sex differences were also seen in active responses, inactive responses, and discrimination indices (Supplementary Fig. S1). Given the significant effect of sex on all vapor administration measures, data were stratified by sex for all subsequent statistical analyses.
Response-contingent cannabis administration in adolescence impairs behavioral flexibility in female rats in adulthood
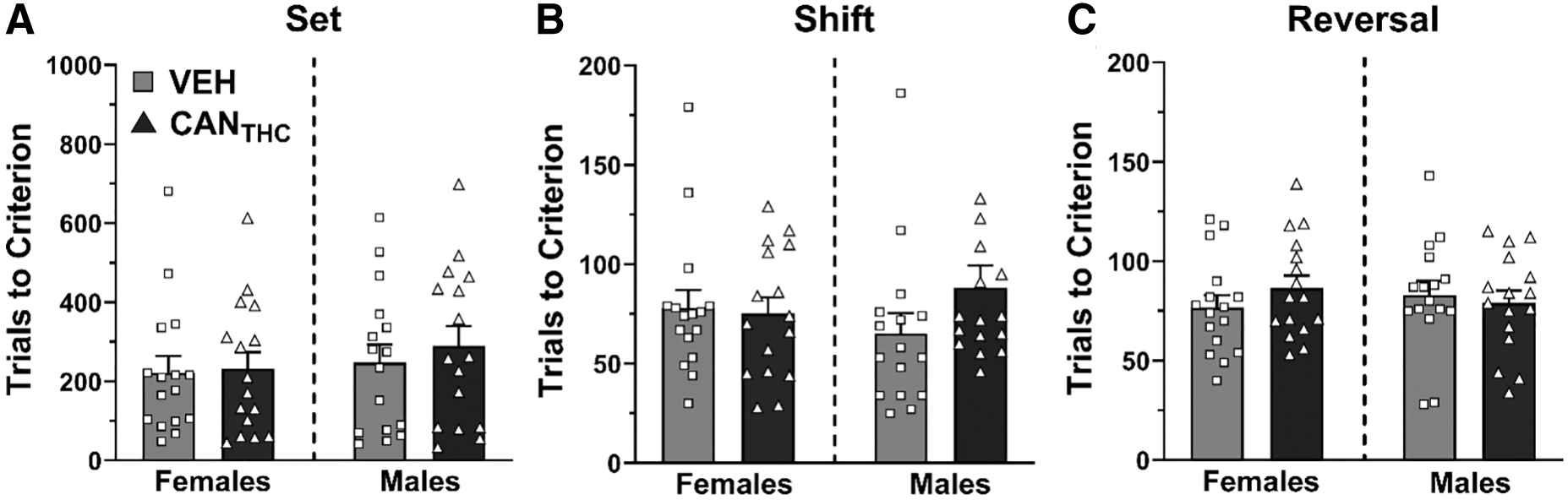
After a 2-week washout period, behavioral flexibility was assessed in visual cue discrimination (set), set shifting, and reversal tasks. For females, treatment did not significantly impact trials to reach criterion during the initial visual discrimination (Fig. 3A) or the reversal tasks (Fig. 3C). However, during the set-shift (Fig. 3B), females that earned CANTHC vapor during adolescence took significantly more trials to reach criterion compared with vehicle females (p=0.025, d=1.21) (treatment: F2,37=3.39, p=0.0445). This increase in trials to criterion was driven by an increase in regressive errors made by CANTHC females compared with vehicle females (p=0.015, d=1.14) (treatment by error type interaction: F4,38=2.80, p=0.0394, Fig. 3D).

CANTHC vapor during adolescence impairs behavioral flexibility in female rats when tested in adulthood. Trials to reach criterion in the visual cue discrimination
For males, there were no significant effects of treatment on trials to reach criterion for the initial visual cue discrimination, the set-shift, or the reversal. There were no significant effects of response-contingent vapor administration on effort discounting for either sex (Supplementary Fig. S2).
Response-contingent cannabis vapor administration in adolescence produces sex-dependent effects on microglial morphology (but not myelination) in adulthood
Microglia
Iba1 fluorescence was conducted in mPFC tissue (n=6/group) to determine if microglia are altered in adulthood following response-contingent vapor administration during adolescence (Fig. 4A). In females, there were significant effects of treatment on all four measures—cell surface area (F2,15=89.68, p<0.0001, Fig. 4B), cell perimeter (F2,15=42.42, p<0.0001, Fig. 4C), soma area (F2,15=38.61, p<0.0001, Fig. 4D), and soma roundness (F2,15=14.05, p=0.0004, Fig. 4E). Female CANTHC rats had significantly reduced microglia surface area (p<0.0001, d=6.48), reduced microglia perimeter (p<0.0001, d=4.29), larger soma area (p<0.0001, d=4.50), and rounder soma (p=0.0012, d=3.41) compared with vehicle females. Female CANCBD rats had significantly increased soma size relative to vehicle females (p=0.026, d=1.86), but no other significant microglial changes. In contrast to females, males exhibited no significant effects of treatment on microglial morphology (Fig. 4).
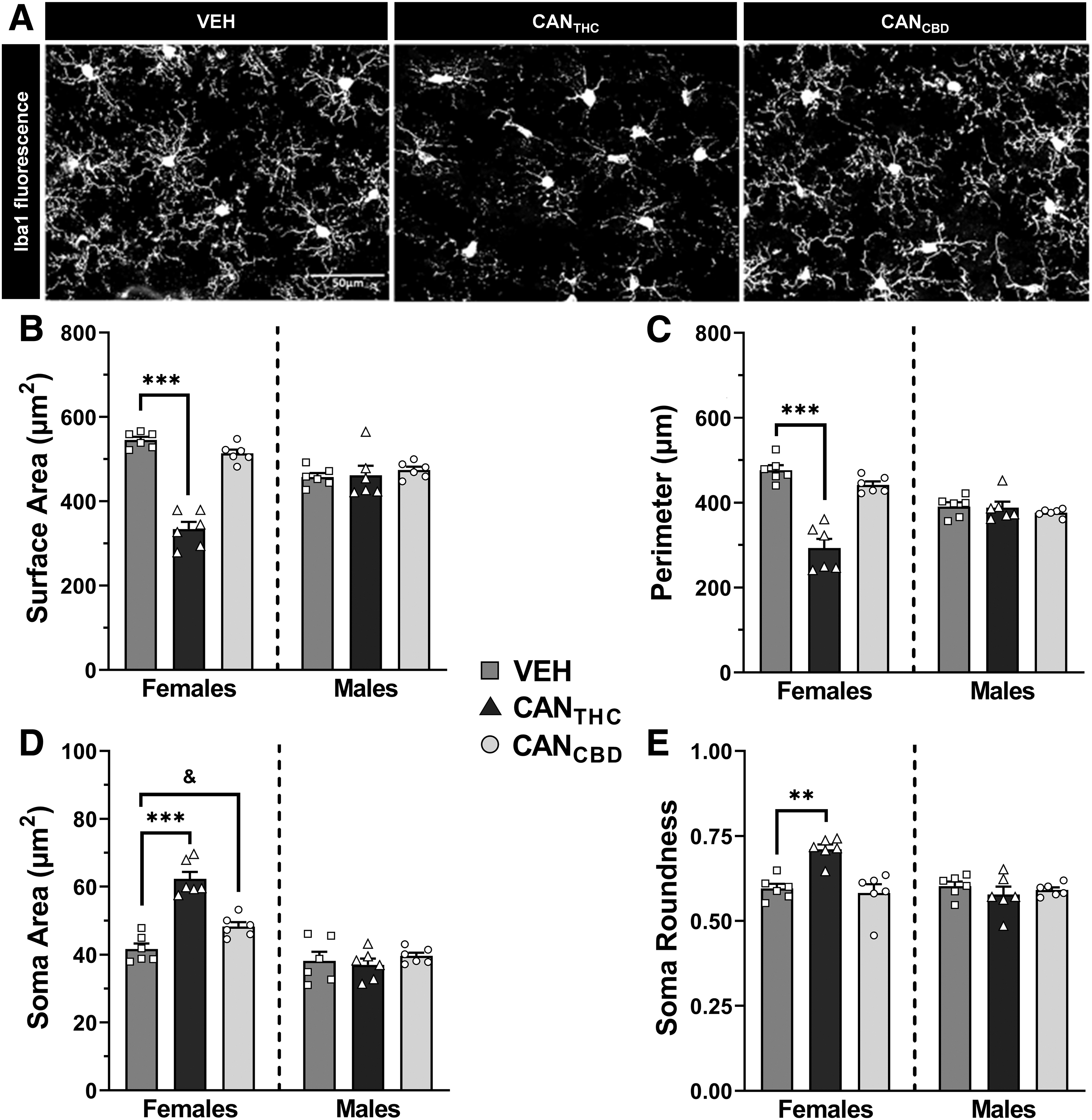
Response-contingent cannabis vapor administration affects mPFC microglial morphology in adulthood in a sex- and cannabis vapor-specific manner. In females, response-contingent CANTHC vapor administration during adolescence leads to more reactive microglia in the mPFC in adulthood. In males, microglia morphological changes in the mPFC occur following CANCBD vapor administration in adolescence.
Since microglia are known to mediate synaptic pruning, we also examined the effects of response-contingent vapor administration during adolescence on dendritic spines in the mPFC in adulthood in females, but no significant effects were detected (Supplementary Fig. S3).
Myelination
MBP fluorescence was measured to assess the effect of response-contingent vapor administration on myelination in the mPFC (Fig. 5A). For both males and females, there were no significant effects of treatment on MBP Gray Value (Fig. 5B).

Myelination in the mPFC in adulthood is not significantly altered by response-contingent cannabis vapor administration during adolescence.
Experiment 2: noncontingent vapor administration
Noncontingent vapor delivery during adolescence fails to recapitulate effects of response-contingent cannabis administration on behavioral flexibility and microglial morphology
Sex differences in responding for CANTHC vapor during adolescence compromised our ability to determine whether the effects on behavioral flexibility and microglial morphology were driven by sex or quantity of CANTHC exposure. Thus, behavioral flexibility was also assessed in a separate cohort of adult male and female rats that were exposed during adolescence to noncontingent CANTHC or vehicle vapor at quantities comparable to that earned by adolescent females (Fig. 6).
Noncontingent CANTHC vapor exposure during adolescence did not impair behavioral flexibility in rats, regardless of sex, when tested in adulthood. Trials to reach criterion in the visual cue discrimination
Behavioral flexibility measures did not significantly differ between noncontingent CANTHC vapor and vehicle vapor groups. However, it is worth noting that the number of trials needed to reach criterion in the set-shifting task in VEH-exposed females was significantly higher than the number of trials required by females following response-contingent VEH administration (

Effects of noncontingent vapor exposure during adolescence on mPFC microglial morphology in adulthood in males. Microglial (
Notably, we similarly observed that microglia parameters (cell perimeter and soma area) in noncontingent VEH males were on par with female rats that underwent response-contingent CANTHC administration. Thus, there may be subtle differences in the effects of response-contingent and noncontingent CANTHC administration that should be considered when implementing vapor delivery approaches in rats.
Discussion
In the present study, male and female rats earned CANTHC, CANCBD, or vehicle vapor during adolescence with behavioral and biological endpoints assessed in adulthood, following a 2-week washout period. Adolescent female rats earned more CANTHC and vehicle (but not more CANCBD) vapor deliveries than adolescent males. Notably, adolescent rats earned CANTHC vapor more readily than CANCBD. Since cannabis use in humans has been associated with executive dysfunction and amotivation, we used an attentional set-shifting task 56 and an effort discounting 57 task to assess behavioral flexibility and amotivation, respectively. Our findings partially supported our hypothesis, as females (but not males) that obtained CANTHC vapor committed more regressive errors leading to impaired shifting between strategies, while effort-based decision making was not altered in rats of either sex.
Our finding that adolescent females earn more CANTHC vapor than adolescent males is consistent with our previous report of sex differences in adult rats, 52 suggesting that sex differences in response-contingent vapor administration are present as early as adolescence and persist into adulthood. Adolescent males appear to earn fewer vapor deliveries compared with adult males in our previous reports,33,52 suggesting that there may be age differences in the reinforcing properties of cannabis vapor. Future work is needed to directly assess age differences using this vapor paradigm.
Interestingly, adolescent rats earn less CANCBD than CANTHC vapor, consistent with our prior work in adult males, 33 suggesting that the phytocannabinoid concentrations present in the cannabis extract influence volitional vapor administration. The complex interactions between phytocannabinoids in the whole-plant cannabis extract likely contributes to the CANCBD-induced suppression of volitional responding for vehicle vapor, perhaps due to changes to cannabinoid pharmacokinetics31,32 or potential olfactory differences. This CANCBD-induced suppression of responding for vehicle vapor was also present in adult male rats in the first 15 min bin in our previous report. 33 Thus, age differences in pharmacokinetics or sensitivity to olfactory stimuli may contribute to the lack of continued suppression in the adult males in our previous study.
It is important to note that both adolescent males and females readily respond for the vehicle vapor to a similar extent as the CANTHC vapor. This finding is in line with our previous report in adult rats of both sexes, where differences in responding for cannabis vapor versus vehicle vapor only emerged after increasing the demand of the reinforcement schedule. 52 Notably, while CANTHC and vehicle vapor were earned at similar levels during adolescence, only CANTHC vapor administration induced behavioral deficits in adulthood in female rats.
Deficits in behavioral flexibility, but not effort-based decision making, were observed in adulthood following adolescent cannabis vapor administration. Specifically, CANTHC females were impaired when shifting strategies in an attentional set-shifting task compared with vehicle females. This deficit was primarily due to an increase in regressive errors made during the shift, indicating that CANTHC females were impaired in maintaining the new strategy. When we noncontingently exposed rats of both sexes (Exp. 2) to the same quantity of CANTHC vapor that adolescent female rats earned in Exp. 1, we failed to find the same treatment effects in the shift component.
Our inability to recapitulate effects from the response-contingent administration study may be due to the stressful nature of noncontingent vapor exposure. While we did not measure corticosterone levels in the current study, our laboratory has previously reported lower birthweights of offspring from dams noncontingently exposed to vehicle vapor compared with air-exposed controls, 54 an effect that was not present when dams underwent response-contingent vehicle vapor administration, 59 suggesting that the noncontingent vapor exposure is stressful.
In support of this possibility, mild stress during adolescence has been shown to specifically increase trials to criterion during the extradimensional set-shifting phase of an attentional set-shifting task in adulthood,
60
and both controllable and uncontrollable stress during adolescence have been shown to impair behavioral flexibility in adulthood.
61
Although our experimental design precludes direct statistical comparison of rats from Exp. 1 and Exp. 2, it is worth noting that females exposed to vehicle vapor noncontingently during adolescence took more trials to reach criterion in the shift component of the behavioral flexibility task than females that responded for vehicle vapor during adolescence (
Another potential explanation for the lack of treatment effects in the noncontingent vapor exposure study could be that these rats were obtained from different vendors (Exp. 1: Simonsen Laboratories vs. Exp 2: Envigo). Unfortunately, Simonsen Laboratories' rat colony closed during the COVID-19 pandemic, and we had to obtain rats from a different vendor for follow-up studies. Prenatal/early postnatal environment and shipping stress are important mediators of cannabinoid receptors with opposing effects in each sex, 62 which could uniquely impact their response to exogenous cannabinoid exposure.
Another aim was to test whether response-contingent cannabis vapor administration during adolescence disrupts ongoing development of the mPFC. During adolescence, the brain undergoes extensive development and reorganization of cortical regions that are critical for executive functioning.3,63,64 These developmental processes include myelination 65 and synaptic pruning, 66 which is partly mediated by microglia.26,27 Supporting the endocannabinoid system's vital role in mPFC development, injections of THC during adolescence critically disrupt these processes.24,67–69
Interestingly, observed behavioral deficits following CANTHC vapor in adolescence were specific to the shift component of the behavioral flexibility task, which has been shown to be mPFC dependent. 56 We predicted that adolescent use of cannabis vapor would decrease myelination but increase microglial activation and synaptic pruning, thereby producing a net reduction in dendritic spine density. While we found no effects of response-contingent vapor administration on myelination, we did find that mPFC microglia of female rats that earned CANTHC were more amoeboid-like—characteristic of the reactive state.
This finding is consistent with previous reports in females using THC injections.24,64 Interestingly, Zamberletti et al. 24 found that preventing the altered microglia phenotype through coadministration of THC and ibudilast (a compound that blocks microglia activation) prevented THC-induced behavioral deficits in adulthood, suggesting that more reactive microglia may play a causal role in adult behavioral dysfunction from adolescent exposure. Future work should determine whether more reactive microglia also contributes to the behavioral flexibility deficits we observed.
Notably, when we noncontingently exposed male rats to vapor during adolescence, we did not find the same effects on microglial morphology. Notably, microglial cell perimeter and soma size of vehicle-exposed males were significantly different than males who earned vehicle vapor, such that the vehicle-exposed males had more microglial activation comparable to females who earned CANTHC vapor. These findings of increased microglial activation after noncontingent vehicle vapor exposure suggest that the stress of forced vehicle vapor exposure may alter microglial activation, thereby occluding potential vapor effects. Altogether, our behavioral flexibility and microglia data following noncontingent vehicle vapor exposure indicate that factors associated with forced exposure to vehicle vapor may have long-lasting impacts on behavior and brain-related endpoints.
More reactive microglia may lead to premature and/or greater synaptic pruning during adolescence, potentially resulting in fewer dendritic spines in the PFC. However, analyses of dendritic morphology did not reveal a decrease in spine density in our CANTHC group. This is consistent with a recent study in male rats that reported premature synaptic pruning immediately following adolescent THC exposure that did not manifest as a difference in spine number after a washout period. 65 Premature synaptic pruning may lead to aberrant synaptic connections, which could then influence cognitive function in adulthood without a change in the total number of spines. However, it is important to note that we observed more reactive microglia only in females, and our spine analyses were not powered to detect sex differences due to issues with cell filling in brain slices taken from male rats. Thus, further investigation is needed to determine if the more reactive microglia seen in females is associated with premature and/or greater synaptic pruning.
Conclusion
The current study revealed sex differences in response-contingent cannabis vapor administration in adolescence that produced cognitive dysfunction and alterations in microglial morphology later in life. Importantly, the effects observed herein are in line with effects that have been observed following injections of low doses of THC. Overall, these data support the generalizability of preclinical data generated using low-dose injection approaches and contribute to a growing body of literature indicating long-term effects on behavior and cortical development from repeated cannabis use during adolescence. Moreover, these findings highlight the subtle, but important, distinctions between response-contingent and noncontingent vapor exposure that should be systematically investigated in future studies.
Footnotes
Acknowledgments
The authors would like to thank Maury Cole and LJARI, Inc. for their continued support with vapor chamber troubleshooting and optimization. The authors would also like to thank Abigail Rossi, Alyssa Hampton, and Manuel Rojas for their assistance with behavioral testing and tissue analysis.
Authors' Contributions
T.G.F.: data curation (equal), investigation (lead), project administration (lead), and software (equal); S.R.W.: data curation (equal), formal analysis (lead), writing—original draft (lead), and writing—review and editing (equal); E.Z.: formal analysis (supporting), investigation (supporting), and visualization (supporting); J.R.K.: investigation (supporting), project administration (supporting), and writing—review and editing (supporting); H.R.W.: investigation (supporting), project administration (supporting), software (equal), and writing—review and editing (supporting); A.N.M.: investigation (supporting); M.W.M.: investigation (supporting); A.M.B.: investigation (supporting); N.C.G.: investigation (supporting); D.E.G.: investigation (supporting); C.M.K.: formal analysis (supporting), visualization (supporting); K.M.D.: resources (equal), software (equal), supervision (supporting), and writing—review and editing (supporting); T.R.: conceptualization (supporting), resources (equal), software (equal), and supervision (supporting); R.J.M.: conceptualization (lead), funding acquisition (lead), methodology (lead), resources (lead), supervision (lead), visualization (supporting), writing—original draft (supporting), and writing—review and editing (equal).
Author Disclosure Statement
No competing financial interests exist.
Funding Information
This study was supported by an NIH R21 grant from the National Institute on Drug Abuse (R21DA043722-01) awarded to Ryan J. McLaughlin, as well as funds provided for medical and biological research by the State of Washington Initiative Measure Number 171 (Ryan J. McLaughlin). Erica Zamberletti and Tiziana Rubino are supported by funds from the Zardi-Gori Foundation (zamb001zardigori).
Abbreviations Used
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.