
Abstract
Background:
Cannabis and its primary psychoactive constituent delta-9-tetrahydrocannabinol (D9-THC) produce biphasic, dose-dependent effects on anxiety. In addition to D9-THC, cannabis contains other “minor” cannabinoids and terpenes with purported therapeutic potential for the treatment of anxiety. Empirical data on potential therapeutic effects of these compounds is limited. The current study evaluated the effects of selected minor cannabinoids and terpenes in a battery of tests sensitive to anxiolytic and anxiogenic drugs.
Methods:
In Experiment 1, adult male Sprague Dawley rats (N=7–8/group) were administered acute oral doses of one of five minor cannabinoids: delta-8-tetrahydrocannabinol (D8-THC; 10 mg/kg), tetrahydrocannabivarin (32 mg/kg), cannabidiolic acid (32 mg/kg), cannabidivarin (32 mg/kg), and cannabigerol (100 mg/kg), or one of five terpenes: D-limonene (17 mg/kg), ⍺-pinene (100 mg/kg), ⍺-terpineol (10 mg/kg), bisabolol (100 mg/kg), and β-caryophyllene (17 mg/kg), or vehicle (medium-chain triglycerides [MCT] oil). Ethyl alcohol was tested as an active comparator. Thirty minutes post-administration, the marble burying test, the three-chamber social interaction test, and the novelty-induced hypophagia test were completed; motor activity was assessed throughout testing. Experiment 2 examined the potential anxiolytic effects of minor cannabinoids when administered chronically; rats administered MCT oil or minor cannabinoids in Experiment 1 continued receiving once-daily doses for 21 days and were assessed using the same test battery after 7, 14, and 21 days of administration.
Results and Conclusions:
When compared to vehicle, acute administration of bisabolol and D-limonene increased the amount of food consumed and bisabolol-, D-limonene-, ⍺-pinene-, and β-caryophyllene decreased percent time spent in the outer zone in the novelty-induced hypophagia test, suggestive of an anxiolytic effect. Only ethanol increased social interaction. After acute administration, anxiogenic effects in the marble burying test were observed for D8-THC, but not for other minor cannabinoids and terpenes. Throughout chronic administration, only D8-THC displayed anxiogenic effects in the novelty-induced hypophagia test. The other cannabinoids did not show anxiolytic or anxiogenic effects in any of the tests at the doses or times tested. The minor cannabinoids and terpenes did not impair or stimulate general motor activity. These data provide a foundation for future studies investigating cannabinoid/terpene interactions.
Introduction
Regulatory changes in state laws legalizing medical and nonmedical use of cannabis and the federal deregulation of hemp have resulted in a growing retail industry of products varying in chemical “chemotype,” product formulation, and intended routes of administration. In addition to the most predominant cannabinoids in cannabis, delta-9-tetrahydrocannabinol (D9-THC) and cannabidiol (CBD), cannabis products also contain other, less abundant cannabinoids such as tetrahydrocannabivarin (THCV), cannabidiolic acid (CBDA), cannabidivarin (CBDV), cannabigerol (CBG), and terpenes.1,2 Terpenes in cannabis of particular interest included in the current studies are D-limonene, ⍺-pinene, ⍺-terpineol, bisabolol, and β-caryophyllene.
These cannabinoids and terpenes are produced in the glandular trichomes of the cannabis plant alongside D9-THC and CBD.1,2 Delta-8-tetrahydrocannabinol (D8-THC) is an isomer of D9-THC found in very small quantities in aged cannabis plant material. 3 D8-THC can also be synthesized in the laboratory via conversion from hemp-extracted CBD. 4 Cannabinoids and terpenes interact with cell membranes, neurotransmitter receptors, G-protein coupled receptors, second messenger systems, and enzymes, and are theorized to contribute to the therapeutic effects of cannabis and to mitigate some of the unwanted effects of D9-THC (i.e., the “entourage effect”).1,2 With selective breeding and refined extraction processes, products that have specific cannabinoid and terpene profiles are being developed and marketed for specific medical conditions and pharmacological effects. There is growing interest in cannabinoids and terpenes as novel therapeutics for treating a range of neuropsychiatric disorders, including anxiety.1,2,5
Cannabinoids other than D9-THC that are naturally present in cannabis are termed “minor” cannabinoids. Many of the minor cannabinoids display pharmacology that is similar to D9-THC, in that they act as a partial agonists at cannabinoid type 1 receptors (CB1R) and cannabinoid type 2 receptors (CB2R). 6 To date, studies examining the behavioral pharmacology of minor cannabinoids are limited. Existing preclinical evidence demonstrates that D8-THC has cannabimimetic effects, producing catalepsy and hypothermia, reducing thermal sensitivity, and altering motor behavior in a dose- and route-dependent manner.7–11 Several human studies indicate a weaker potency of D8-THC at CB1R compared with D9-THC.12,13 THCV, a propyl analog of D9-THC which exerts biphasic agonist/antagonist action at CB1R and partial agonist action at CB2R,1,2,5,6 reportedly rescues schizophrenia-like behavior in the phencyclidine rat model of schizophrenia without altering behavior in unmanipulated rats.14,15
CBDV is a propyl analog of CBD and has mild affinity for CB1R and CB2R as well as the transient receptor potential (TRP) superfamily.1,2,5 CBDV dose dependently attenuates autism-like behavior (e.g., hyperlocomotion, stereotypies and deficits in social behavior and working memory task performance) in adolescent rats exposed prenatally to valproic acid without altering behavior in saline-treated controls. 16 CBDA, which is the precursor to CBD and interacts with numerous signaling systems, including serotonergic neurotransmitter receptor subtypes and TRP channels,2,5 dose- and context-dependently decreases anxiety-like behavior and increases appetitive behavior in rats. 17
CBG, which is the precursor to both D9-THC and CBD biosynthetic pathways,1,2,5 also reportedly decreases anxiety-like behavior and increases appetitive behavior. 18 Taken together, preliminary evidence exists demonstrating that D8-THC, THCV, CBDV, CBDA, and CBG are physiologically and behaviorally disruptive with potential therapeutic applications for anxiety-related and other neuropsychiatric disorders.
Like the minor cannabinoids, studies examining the behavioral pharmacology of terpenes found in cannabis are limited. ⍺-Pinene is the most widely encountered naturally occurring terpene found in conifer (e.g., pine) and other plant oils.1,2 Existing preclinical evidence demonstrates that ⍺-pinene decreases anxiety-like behavior in mice. 19 Limonene is the second most widely encountered naturally occurring terpene found in lemons and other citrus. 1 β-Caryophyllene, found in a variety of plant oils, including from cloves, black pepper, and rosemary.1,20,21 Both D-limonene and β-caryophyllene decreased anxiety-like behavior in mice, and β-caryophyllene also reduced depressive-like behavior.22–24
Investigations of isolated ⍺-terpineol, commonly found in pine oil, and bisabolol, the primary constituent in chamomile essential oil, have focused largely on antinociceptive properties.25,26 One study has demonstrated that bisabolol decreases anxiety-like behavior in mice. 27 We are unaware of any studies in the published literature that tested possible anxiolytic or anxiogenic effects of isolated ⍺-terpineol.
Studies directly comparing the acute and chronic behavioral effects of orally administered minor cannabinoids and terpenes in the same rodent model are lacking. As the popularity and use of cannabis and cannabis/cannabinoid products increases, investigating the behavioral and psychoactive effects of isolated, biologically active minor cannabinoids and terpenes is necessary to educate users and physicians. In this study, we investigated the acute effects of orally administered minor cannabinoids and terpenes (Experiment 1) and the chronic effects of oral minor cannabinoids administered daily for 21 days (Experiment 2) on anxiety-like behavior using a battery of procedures sensitive to anxiolytic/anxiogenic, stimulant, and sedative drug effects. Procedures included the marble burying test, the three-chamber social interaction paradigm, and the novelty-induced hypophagia test; general motor activity was assessed throughout all tests.
Materials and Methods
Animals
Adult (∼8- to 9-week-old) male Sprague Dawley rats (Charles River Laboratories, Wilmington, MA) weighing 200–400 g at the start of experimentation were used (n=95). Animals were food restricted to 90% of their initial bodyweight before study initiation and maintained at this target weight throughout the study period. This ensured animals maintained healthy weight during the study and promote food seeking in the Novelty-induced hypophagia test (see below). Before dosing, rats were matched by weight to one of 12 treatment groups (n=7–8 per group). Body weights of individual rats were targeted to be within ±20% of the group mean.
This study complied with all applicable sections of the Final Rules of the Animal Welfare Act regulations (9 CFR Parts 1, 2, and 3), as well as the Guide for the Care and Use of Laboratory Animals (2011). All experimentation was performed at Lovelace Biomedical, an organization fully accredited by the Association for Assessment and Accreditation of Laboratory Animal Care.
Drugs
THCV, CBDV, CBDA, and CBG were obtained from Open Book Extracts (Roxboro, NC). D8-THC was obtained from ACS Laboratory (Sun City Center, FL). D-limonene, ⍺-pinene, ⍺-terpineol, bisabolol, and β-caryophyllene were obtained from Sigma-Aldrich (St. Louis, MO). All test compounds were mixed with medium-chain triglycerides (MCT) oil (100% Caprylic Capric Triglycerides; SpecializedRx Products, LLC, Fridley, MN) vehicle 24 h before treatment and stored overnight at 2–8°C. Rats were administered a single dose of one of the minor cannabinoids or terpenes (n=8 per group) or ethanol (n=7) via oral gavage at a volume of 1 mL/kg. The vehicle control group (n=8) was administered MCT oil at the same volume.
Doses for each minor cannabinoid were D8-THC (10 mg/kg), THCV (32 mg/kg), CBDV (32 mg/kg), CBDA (32 mg/kg), and CBG (100 mg/kg). Doses for each terpene were D-limonene (17 mg/kg), ⍺-pinene (100 mg/kg), ⍺-terpineol (10 mg/kg), ⍺-bisabolol (100 mg/kg), and β-caryophyllene (17 mg/kg). To facilitate similar pharmacodynamics, drugs were administered under fasted conditions (i.e., rats were not fed before drug administration on the day of testing).
Where possible, doses were selected based on existing literature; for example, D8-THC (10 mg/kg) is a behaviorally active dose in rats when vaporized and is a low effective dose intravenous (i.v.) while above 5 mg/kg is physiologically active in mice. CBG (100 mg/kg) is approximate to a dose of CBG that previously enhanced behavior in the novelty-induced hypophagia test.9–11,18 Ethyl alcohol (95%; Sigma-Aldrich) (3 g/kg) was administered by oral gavage as an active comparator based on the acute socially anxiolytic effects of ethanol administration reported in rats. 28
Behavioral testing: acute drug administration
To compare acute anxiolytic effects of minor cannabinoids and terpenes, rats underwent a test battery of three procedures beginning 30 min after their respective treatments, and the test order was the same for all animals: (1) marble burying test, (2) three-chamber social interaction test, and (3) novelty-induced hypophagia test. Each test was conducted consecutively as shown in Figure 1. Data were recorded by manual measurement or by using automated video tracking software (Ethovision XT 16; Noldus Information Tech, Inc., Leesburg, Virginia, USA) and overhead cameras (acA1300-60gm GigE; Basler AG, Ahrensburg, Germany). Videos were recorded at 30 frames per second and analyzed using the Multiple Arenas Module. Detection settings were set using the Automatic behavior recognition tool.

Graphical schematic representing the order and timing of behavioral tests during the acute and chronic timepoints. MBT, marble burying test; NIH, novelty-induced hypophagia test; SI, social interaction test.
Marble burying test
Testing occurred in a 12×12×19 inch acrylic cage (Chewy, Inc.; Item No. 248528) with 5 cm of fine sawdust bedding (Altromin; WH 3-4) and four rows of five glass marbles placed on the bedding surface, evenly spaced apart. Individual rats were placed into the cage and monitored for 15 min. Anxiolytic or anxiogenic drug effects were inferred from a decrease or increase, respectively, in the number of buried marbles (to 2/3 of their depth) and the latency (seconds) to bury the first marble. Latency data were recorded using automated video tracking software and the number of buried marbles was manually counted.
Three-chamber social interaction test
Testing occurred in a custom-built three-chamber apparatus, with an open chamber in the middle and chambers on the left and right side that were separated from the center chamber by a wire grid (chamber dimensions: 120×40×40 cm). A familiar rat (cage mate) was placed in one of the side chambers, and a novel rat was placed in the other side chamber. The test rat was placed in the center chamber and the amount of time spent interacting with the novel rat compared to the familiar rat was recorded for 15 min using automated video tracking software. A new novel rat was used for each repetition of social interaction testing during chronic drug administration (Experiment 2). Anxiolytic or anxiogenic drug effects were inferred from increased or decreased time spent interacting with the novel rat (i.e., in the novel rat zone), respectively.
Novelty-induced hypophagia test
Testing was completed in a novel environment (120-gallon Ace/Den Hartog Stock Tank; 48″H, 24″D) under a bright lighting (∼1200 lux) to enhance aversiveness. Using Ethovision software, the tank was divided into predefined areas as follows: (1) the outer zone was operationally defined as 10 cm from the tank wall. This length was chosen as being less than the length of a rat to ensure that their body was in contact with the outer walls of the apparatus (i.e., rats displayed thigmotaxis); (2) the food zone was defined as the diameter of the food bowl (12.7 cm), representative of interaction with food; and (3) the middle zone was defined as the remaining area excluding the outer and food zones.
Rats were placed along the edge of the tank in the outer zone at the start of the test and were allowed to explore the environment for 15 min. Anxiolytic or anxiogenic drug effects were inferred by decreased or increased latency, respectively, to enter the center food zone and consume a novel, highly palatable food reward. Data were recorded using automated video tracking software to track animals in the center food zone, the outer zone along the walls of the environment (i.e., tracking thigmotaxis), and by weighing the food before and after testing. Cocoa Puffs (General Mills, Inc., Minneapolis, Minnesota, USA) were chosen as the food reward, as through an initial pilot study, we determined that all rats exhibit a strong preference (all p<0.0001, data not shown) for this cereal over others.
Motor activity assessment
Drug effects on general motor activity were assessed based on total distance traveled (m) via automated video tracking software during the marble burying test, the three-chamber social interaction test, and the novelty-induced hypophagia test.
Behavioral testing: chronic drug administration
To assess chronic effects of repeatedly dosing minor cannabinoids, rats treated acutely with MCT oil vehicle or minor cannabinoids continued receiving daily administrations via oral gavage for a period of 21 days. Behavioral testing, exactly as described above, was repeated on days 7, 14, and 21, 30 min after treatments and in the same order for all animals. A new novel rat was used for each repetition of social interaction testing during chronic drug administration.
Statistical analysis
Outcome variables analyzed were number of marbles buried for the marble burying test; percent of time (sec) spent with a novel rat and number of entries into the novel zone for the three-chamber social interaction test; amount of food consumed (g), percent of time spent in the food zone, percent of time spent in the outer zone, and latency (sec) to enter the food zone in the novelty-induced hypophagia test, and distance traveled (m) for general motor activity. All data were analyzed with GraphPad Prism version 9.5.1.
For Experiment 1, data for each of the outcomes tested following acute administration of each of the five terpenes (bisabolol, D-limonene, ⍺-pinene, and β-caryophyllene), the five minor cannabinoids (D8-THC, THCV, CBDA, CBDV, and CBG), ethanol, and the MCT oil control were analyzed using one-way ANOVAs to compare effects across treatment conditions. When main effects of treatment were observed, Dunnett's post hoc tests comparing each active treatment condition to the MCT vehicle were used to isolate significant treatment effects.
For Experiment 2, the same animals that received the five minor cannabinoids or the MCT-control in Experiment 1 continued dosing to evaluate potential changes in effects after 21 days of chronic dosing (treatment×day). Therefore, for Experiment 2, separate two-way repeated-measures ANOVAs for each outcome were completed to compare treatment effects across days. Day 1 of chronic dosing was the first time that the animals received acute administration of each drug and thus is the same as the data from Experiment 1.
When main effects of treatment and day were observed, or interaction effects (treatment×day) were observed, Dunnett's post hoc tests were completed as follows: we compared each active treatment condition to the MCT oil vehicle to isolate significant effects of treatment for each day. In addition, we conducted Dunnett's post hoc tests to determine differences between day 1 (acute treatment) and 21 (last day of chronic treatment) within each treatment group. If there was only a main effect of the day observed, data were collapsed across treatment groups and Dunnett's post hoc tests were conducted to determine differences between day 1 (acute treatment) and 21 (last day of chronic treatment).
The Geisser and Greenhouse correction was used when sphericity of data could not be assumed. Data were examined for outliers and those meeting criteria (>3 standard deviations from the mean) were removed from analysis and instances were reported below. A p<0.05 for a two-tailed test was accepted as significant.
Results
Experiment 1: acute administration of minor cannabinoids and terpenes
Marble burying test
There was a main effect of treatment on the number of marbles buried after acute administration (F[11, 83]=3.127, p=0.001). Post hoc testing determined that D8-THC increased the number of marbles compared to vehicle (p=0.008; Fig. 2A); ethanol, the other minor cannabinoids, and terpenes tested did not differ from vehicle. There was no effect of treatment on distance traveled (Fig. 2B).
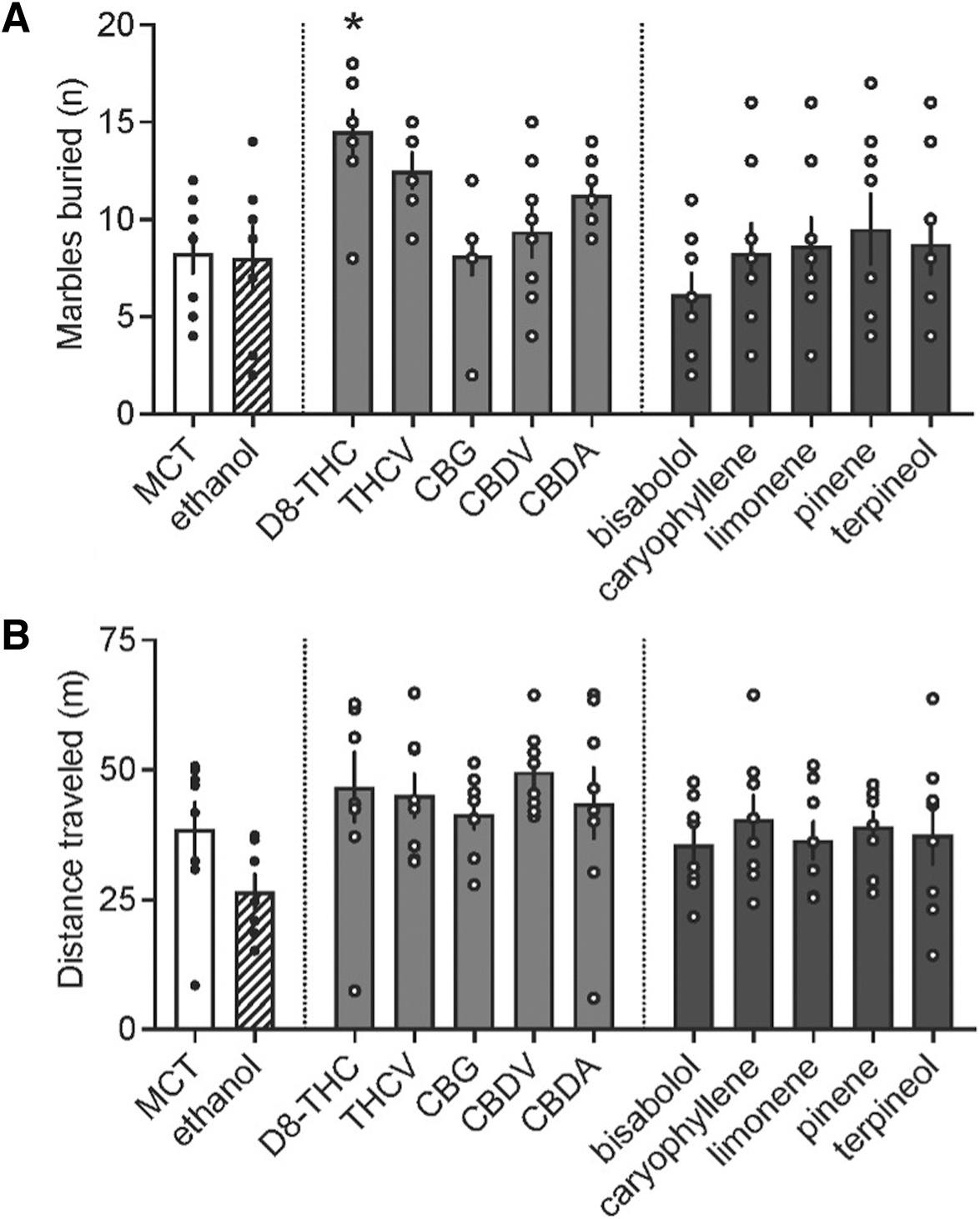
Marble burying test after acute oral administration of minor cannabinoids and terpenes.
Social interaction test
There was a main effect of treatment on percent time spent with a novel rat after acute drug administration (F[11, 83]=6.464, p<0.001). Post hoc testing revealed that only ethanol-treated rats spent more time interacting with a novel rat than vehicle control rats (p<0.001; Fig. 3A); minor cannabinoids and terpenes tested did not differ from vehicle. There was also a main effect of treatment on the number of entries into the novel rat zone (F[11, 83]=3.386, p<0.001) with post hoc testing determining that ethanol-treated rats entered the novel rat zone more than vehicle treated rats (p<0.001; Fig. 3B); minor cannabinoids and terpenes tested were no different from vehicle. Although significant main effects of treatment on distance traveled (F[11, 83]=2.923, p=0.003) were found, post hoc tests determined that treatment groups were not significantly different from vehicle controls (Fig. 3C).

Three-chamber social interaction test after acute oral administration of minor cannabinoids and terpenes.
Novelty-induced hypophagia test
One outlier was removed from the test dataset (vehicle control rat; food consumed was >3 SD from group mean). There was a main effect of treatment on amount of food consumed (F[11, 82]=4.130, p<0.001). Post hoc testing determined that compared to vehicle control rats, bisabolol- and D-limonene-treated rats consumed more food (p=0.004 for both; Fig. 4A). There was a main effect of treatment on latency to enter food zone (F[11, 82]=2.465, p=0.0102; Fig. 4B) and percent of time spent in the novel food zone (F[11, 82]=2.904, p=0.003; Fig. 4C), however, post hoc testing did not isolate any differences from minor cannabinoid, terpene, or ethanol compared to vehicle for these measures.

Novelty-induced hypophagia test after acute oral administration of minor cannabinoids and terpenes.
There was also a main effect of treatment on percent time spent in outer zone (F[11, 82]=7.649, p<0.001). Post hoc testing determined that compared to vehicle control rats, ethanol-, bisabolol-, D-limonene-, ⍺-pinene-, and β-caryophyllene-treated rats spent less time in the outer zone (ps<0.05; Fig. 4D). No differences in distance traveled were observed between treatment and control conditions (Fig. 4E).
Experiment 2: chronic administration of minor cannabinoids
Marble burying test
There was an interaction of treatment×day (F[15, 126]=2.592, p=0.002) on marbles buried in the marble burying test. Post hoc testing determined that D8-THC and THCV treatment increased the number of marbles buried compared to vehicle on day 1 (p=0.005 and p=0.03, respectively), but this effect was mitigated with continued treatment as shown by a decrease in the number of marbles buried from day 1 to 21 (ps<0.05). There were no differences in marbles buried across test days for vehicle controls or CBG-, CBDV-, and CBDA-treated rats (Fig. 5A).

Repeated marble burying tests throughout 21 days of daily oral administration of minor cannabinoids.
There was a main effect of day on distance traveled during the marble burying test (F[2.549, 107.1]=3.245, p=0.03). Post hoc testing did not isolate any differences between tests on days 1 and 21 of chronic administration (Fig. 5B).
Social interaction test
There were no main effects of treatment or day, nor an interaction of treatment×day, on percent time spent in, or entries to, the novel rat zone in the social interaction test (Fig. 6A, B). There was a main effect of day on distance traveled (F[1.734, 72.84]=8.580, p<0.001) in the social interaction test; post hoc tests indicated that distance traveled was lower on day 21 compared with day 1 when collapsed across treatment groups (ps<0.05; Fig. 6C).

Repeated social interaction tests throughout 21 days of daily oral administration of minor cannabinoids.
Novelty-induced hypophagia test
There were main effects of treatment (F[5, 41]=3.961, p<0.005) and day (F[2.903, 119.0]=107.6, p<0.0001) on the amount of food consumed in the novelty-induced hypophagia test. Post hoc testing determined that CBDV increased food intake (g) greater than vehicle (p=0.04) only on day 1 and D8-THC reduced food intake compared to vehicle on day 14 (p=0.004). Post hoc testing determined that food consumption increased across all treatment groups from day 1 to 21 (ps<0.05; Fig. 7A).

Repeated novelty-induced hypophagia tests throughout 21 days of daily oral administration of minor cannabinoids.
There was an interaction of treatment×day (F[15, 123]=2.067, p=0.0158) on latency to enter the food zone. Post hoc testing did not isolate any differences between any treatment and vehicle controls on any day. Latency to enter the food zone decreased from day 1 to 21 for rats treated with CBDA (p=0.03; Fig. 7B). Latency to enter food zone did not change for vehicle controls or any other minor cannabinoid tested.
There was a main effect of day on percent time spent in the food zone (F[2.622, 107.5]=111.1, p<0.001). Across days 1 and 21, post hoc testing determined that percent time in the food zone increased across all treatment groups (ps<0.05; Fig. 7C).
There was a main effect of day on percent time spent in the outer zone (F[2.562, 105.0]=156.9, p<0.001). Across days 1 and 21, post hoc testing determined that percent time spent in the outer zone decreased across all treatment groups (ps<0.05; Fig. 7D).
There was a main effect of day on distance traveled (F[2.060, 84.45]=3.317, p=0.04), but no treatment×day interaction. Post hoc testing determined that there were no differences between days 1 and 21 across treatment groups (Fig. 7E).
Discussion
The current study extends existing knowledge of the pharmacological effects of minor cannabinoids and terpenes. We observed some evidence of acute anxiolytic effects of some terpenes. Namely, bisabolol and D-limonene significantly increased food intake compared to vehicle in the novelty-induced hypophagia test. In addition, bisabolol, D-limonene, ⍺-pinene, and β-caryophyllene reduced time spent in the outer zone during the novelty-induced hypophagia test. The terpenes bisabolol, D-limonene, ⍺-pinene, ⍺-terpineol, and β-caryophyllene did not produce anxiolytic effects in marble burying tests at the doses tested. Acute oral administration of minor cannabinoids D8-THC, THCV, CBDV, CBDA, and CBG did not produce anxiolytic effects in any of tests at the doses tested. There was, however, mixed evidence of potential anxiogenic effects of acute D8-THC. D8-THC increased marble burying, although anxiogenic effects were not evident in the other behavioral tests.
When minor cannabinoids were administered chronically, there were few differences observed compared to vehicle. In the marble burying test, the anxiogenic effect of D8-THC on day 1 dissipated across repeated testing, suggestive of a tolerance to this effect. In the novelty-induced hypophagia test, rats treated with D8-THC consumed less food during the test on day 14. Notably, there were differences observed across days in all groups tested: primary and secondary outcomes either increased (i.e., food consumed and time in the food zone, distance traveled) or decreased (i.e., number of marbles buried, latency to enter the food zone and time spent in the outer zone), although some outcomes remained consistent across tests (i.e., percent time spent and entries in the novel rat zone, motor speed) for rats in the MCT vehicle control and minor cannabinoid treatment conditions. These observations are most likely attributable to habituation to the testing apparatuses and conditions over time.
Considering bisabolol and D-limonene increased food consumed and bisabolol, D-limonene, ⍺-pinene, and β-caryophyllene reduced time spent in the outer zone of the novelty-induced hypophagia test, bisabolol and D-limonene displayed an anxiolytic effect. Although ⍺-pinene and β-caryophyllene reduced time spent in the outer zone, they did not concomitantly increase the amount of food consumed or reduce the latency to enter the food zone (the primary measures of the novelty-induced hypophagia test). A prior study in adult female Wistar rats treated with D-Limonene (10 mg/kg, per os [p.o.]) demonstrated reduced stress-related behaviors compared to vehicle controls (corn oil), including reduced irritability caused by the approach or touch of a stylus and reduced auditory startle responses. 29 In this study, we did not observe anxiolytic effects of any terpenes in the marble burying or social interaction tests.
Thus, taken together, our findings on the potential anxiolytic effects of these terpenes are inconclusive. The lack of any discernable effect of oral D-limonene administration on behavioral outcomes in the marble burying test in our study contrasts a prior study which treated adult male Swiss mice with D-limonene (0.5 and 1 mg/kg, p.o.) and reported reductions in the number of marbles buried in the marble burying test and increases in the time spent in the light side of the light/dark box 30 ; species- and dose-related differences may partially explain this discrepancy. A prior study in ICR strain mice exposed to vaporized ⍺-pinene showed increases in distance traveled, as well as percent of total visits to and time spent in the open arms of the elevated plus maze (EPM). These data suggest some effects may be due to changes in motor activity, although changes in motor activity were not observed in our study. 19
As far as we are aware, this is the first demonstration of a behavior-modifying effect of bisabolol in the novelty-induced hypophagia test and complements existing evidence demonstrating an anxiolytic effect of bisabolol in the EPM at lower doses (0.5, 1, 2 mg/kg, intraperitoneal [i.p.]) and a confounding sedative effect at higher doses (5, 10 mg/kg). 27 In our study, oral administration of a higher dose of bisabolol did not produce a sedative effect. Investigations of possible therapeutic effects of bisabolol must consider differences due to administration route.
The demonstration that 10 mg/kg oral D8-THC increased the number of marbles buried at the acute timepoint is suggestive of an anxiogenic effect. It is well established that D9-THC can produce anxiolytic effects at low doses (e.g., ≤1 mg/kg) and anxiogenic effects at higher doses in rodents (≥2.5 mg/kg).31–34 Thus, our data are consistent with other studies suggesting D9-THC-like effects of D8-THC. For example, D8-THC (30–100 mg/kg, i.p.) produced classic cannabinoid effects (catalepsy, antinociception, hypothermia, and hypolocomotion) in adult male and female C57BL/6J mice. 8 D8-THC fully substituted for D9-THC in C57BL/6J mice trained to discriminate a moderate dose (3 mg/kg D9-THC), but was less potent than D9-THC. 8 Twice daily treatment with D8-THC (50 mg/kg, subcutaneous [s.c.]) for 6 days produced THC-like physical dependence, as evidenced by precipitated withdrawal following administration of CB1R antagonist rimonabant (3 mg/kg, i.p.). 8
Comparative studies of D9-THC and D8-THC behavioral pharmacology consistently demonstrate D9-THC-like effects of D8-THC on the cannabinoid behaviors tetrad assay and drug discrimination assays as recently reviewed by Tagen and Klumpers. 35 Interestingly, chronic administration of D8-THC resulted in less anxiogenic behavior in the marble burying test across days (i.e., no differences from vehicle controls on days 7, 14, and 21), indicating tolerance to this effect. In contrast, repeated testing of D8-THC in the novelty-induced hypophagia test revealed a potentially anxiogenic effect on day 14 of testing. While vehicle control animals increased their food consumed with repeated tests (likely due to habituation to the test chamber), D8-THC rats did so to a lesser extent. This may suggest an anxiogenic effect of D8-THC.
Novelty-induced inhibition of feeding (hypophagia) provides a measure of anxiety-related behavior in rodents that is sensitive to the effects of multiple classes of anxiolytic and anxiogenic drugs and has excellent predictive validity for translating drug effects from animals to humans. 36 The lack of a detectable anxiogenic effect of D8-THC or the other cannabinoids in the novelty-induced hypophagia test on day 1 may be attributable to a floor effect as the test was optimized to detect anxiolytic effects (i.e., fasted rats seeking a highly palatable food reward in a brightly-lit novel environment). 36 Alterations to this test (e.g., a longer assessment duration) may permit detection of anxiogenic effects. 36 It is also possible that the effects of D8-THC on reducing food intake in this test may be due to effects on appetite and/or hedonic eating; acute and chronic D9-THC administration (doses >2.5 mg/kg) is known to reduce food intake in rats. 37
Finally, D8-THC likely has activity outside of CB1R and CB2R (e.g., serotonin) that may contribute to some of these effects, or the differences observed here and in studies that examined D9-THC.
Oral administration of cannabinoids tends to have a longer onset of effects due to slower and more variable absorption rates as well as first-pass metabolism in the liver, leading to lower and delayed peak plasma concentrations compared to injected or inhaled administration routes (reviewed by Dr. Marilyn Huestis in 2005). 38 Assessing the behavioral outcomes of orally administered minor cannabinoids and terpenes beyond 30 min post-treatment may reveal additional patterns in the anxiolytic and social effects of these compounds. Given that some terpenoids appeared promising for anxiety relief, future work should establish whether the terpenoids show long-lasting efficacy for antianxiety behavior.
Furthermore, associating behavioral markers of cannabinoid and terpene exposure with levels in blood plasma and other biological matrices is required, although partially limited by the narrow detection windows for some minor cannabinoids, such as CBG or cannabinol (<30 min in plasma after smoke or vapor inhalation, not detectable after oral administration).39–41 It should be noted that pharmacokinetics and pharmacodynamics of cannabinoids can be altered depending upon form of administration, food (fed vs. non-fed), and/or diet composition (e.g., high vs. low fat). 42 As rats were weight maintained and not fed before dosing, it is possible that this may have altered treatment efficacy and/or masked behavioral disruptions. Indeed, diet is reported in the literature as a modifying factor for drug efficacy.43,44
Future studies would need to use behavioral assays in animals maintained at their standard weights. While we report that oral CBG (100 mg/kg, p.o.) did not affect behavioral outcomes, including food consumed, another study reported that oral CBG (120 mg/kg, p.o.) in presatiated adult male Lister Hooded rats increased total food intake; thus, the appetitive state of the animal may influence the behavioral outcomes of cannabinoid administration. It is also important to consider our selected doses: some of the prior studies have included higher doses, and it is possible that our doses were too low to precipitate additional behavioral disruptions, although the doses chosen for this study were still far above any translational exposure in humans.
In adult female and male CD1 mice administered higher doses of various terpenes, including β-pinene and β-caryophyllene (50–200 mg/kg, i.p.), and tested in the cannabis tetrad assay, enhanced catalepsy and antinociception on the tail-flick assay and reduced body temperature and locomotion were observed. Coadministration of these terpenes with CB1R agonist WIN55-212,2 (10 mg/kg, i.p.) produced an additive effect across all terpenes and for all mice tested, with linalool demonstrating sex-specific additive effects in males compared to females. 45
Conclusion
In conclusion, terpenes bisabolol or D-limonene demonstrated acute anxiolytic effects and D8-THC demonstrated acute anxiogenic effects in Sprague Dawley rats tested using the marble burying and novelty-induced hypophagia tests. Future investigations into the combined effects of minor cannabinoids and terpenes, as well as with major cannabinoids and existing pharmacotherapies, are required. In addition, given the lack of controlled dosing investigations in the relevant behavioral pharmacology literature, additional dose-exploration studies of minor cannabinoids and terpenes are needed. Finally, future investigations into sex-dependent differences in the behavioral outcomes of minor cannabinoid and terpene exposure are crucial for continued therapeutic development and the education of users and physicians.
In conclusion, the current study data add to the limited preclinical literature examining the extent to which isolated minor cannabinoids and terpenes modify behavior. Much like investigations into D9-THC and CBD, studies must be cautiously interpreted and consider differences related to the experimental conditions such as the species, strain, or sex of the animal used for the study and the administration route or dose.
Footnotes
Acknowledgments
The authors thank Lovelace technicians Xiuping Gao and JoAnna O'Neill for excellent technical assistance.
Authors' Contributions
B.W.J.: data curation, investigation, visualization, writing - original draft, and formal analysis; C.F.M.: data curation, investigation, visualization, writing - original draft, writing - review and editing, and formal analysis; D.C.: methodology, investigation, data curation, and writing - review and editing; J.D.M.: supervision, methodology, validation, investigation, and writing - review and editing; T.W.L.: conceptualization, methodology, project administration, and writing - review and editing; M.O.B.-M.: conceptualization, project administration, and writing - review and editing; E.M.W.: data curation, investigation, visualization, writing - original draft, writing - review and editing.
Author Disclosure Statement
Authors T.W.L., and M.O.B.-M. were full-time employees of Canopy Growth Corporation, a producer and retailer of cannabis, and hemp products, at the time of this project.
Authors E.M.W. and C.F.M. have received funds from MyMD Pharmaceuticals, Inc., and MIRA-1 Pharmaceuticals, Inc., for contract preclinical research. E.M.W. received support from Cultivate Biologics LLC, and Canopy Growth Corp for clinical research projects not related to this research. E.M.W., C.F.M., and B.W.J. did not receive funding or compensation from Canopy Growth Corp for this project. B.W.J. and D.C. have no competing financial interests.
Funding Information
This research was funded by Canopy Growth Corporation and conducted by Lovelace Biomedical under contract (J.D.M.).