
Abstract
Objectives:
The aim of this study was to investigate the role of cannabinoid (CB1) receptors on airway inflammation and hypersensitivity in allergic asthma and the potential interactions with TRPV1 channels.
Materials and Methods:
BALB/c mice were sensitized and provoked with ovalbumin to create a model of allergic asthma. CB1 selective agonist arachidonoyl 2′-chloroethylamide (ACEA) was administered intraperitoneally at doses of 0.5, 3, and 5 mg/kg. Receptor antagonism studies were performed utilizing selective CB1 antagonists AM251 at a dose of 3 mg/kg. TRPV1 channel was selectively blocked by capsazepine at a dose of 2.5 mg/kg. Penh values were recorded in vivo by a whole-body plethysmograph under methacholine challenge. Inflammatory cell count was performed in bronchoalveolar lavage fluid (BALF). Serum levels of proinflammatory cytokines were measured by Enzyme-Linked ImmunoSorbent Assay (ELISA). Inflammation in the lung tissue was scored histopathologically. Statistical significance was determined using one-way analysis of variance or Kruskal–Wallis test and expressed as p<0.05.
Results:
In sensitized animals, provocation with inhaled ovalbumin increased Penh values, serum interleukin (IL)-4, IL-5, IL-13 levels, eosinophil, neutrophil, lymphocyte, macrophage counts in BALF, and inflammation in the lung tissue. ACEA applications did not significantly alter Penh values, BALF inflammatory cell levels, and histological changes related to inflammation in the lung tissue according to the disease group; however, only at a dose of 5 mg/kg, it reduced the levels of the inflammatory cytokine IL-4. AM251 decreased Penh values, eosinophil and neutrophil migration in BALF, and inflammation score of lung tissue compared with the disease group. Although BALF inflammatory cell levels and Penh values were higher in the AM251+ACEA group than in the AM251 group, the differences were insignificant. In the CPZ+ACEA group, Penh values were significantly higher, and serum IL-4 and IL-13 levels and BALF eosinophil counts were lower than that in the CPZ group.
Conclusions:
This study demonstrated an important role of the CB1 receptors in allergic asthma. CB1 antagonism reduced airway hyperresponsiveness and inflammation and showed immunomodulatory effects. The effect of the CB1 agonist ACEA on asthma does not appear to be related to TRPV1 channels.
Introduction
Asthma is characterized by hyperreactivity, inflammation, and physiological and structural changes in the airways, and its pathogenesis is quite complex. Many factors including cytokines especially interleukins, prostaglandins, cysteinyl leukotrienes, chemokines, growth factors and immune cells such as mast cells, eosinophils, neutrophils, lymphocytes, and macrophages play a role in the pathogenesis.1,2 Structural changes involve epithelial damage, increased basement membrane thickness, smooth muscle hyperplasia, goblet cell metaplasia, and angiogenesis occur in the airways of asthmatic patients.1,3
The cannabinoidergic system is demonstrated to contribute to the pathophysiology of asthma, and its potential as an effective asthma treatment is an active and attractive area of research. The effects of natural and synthetic, nonselective cannabinoids on airway inflammation, smooth muscle contraction, and remodeling have been investigated in many studies, and hopeful results have been obtained.4–10 Recently, more focus has been put on selective cannabinoid receptor agonists and antagonists to better understand how cannabinoid-1 and -2 (CB1 and CB2) receptors impact respiratory diseases. It has been reported that CB1 agonists reduce tracheal hyperreactivity in vitro and in vivo, 11 inhibit nerve-mediated airway hyperreactivity in nerve growth factor (NGF)-induced airway inflammation in vitro, 12 and suppress reactive oxygen species-mediated vagal lung C fiber hypersensitivity in vivo. 13 However, CB1 antagonists have been shown to exert anti-inflammatory effects in animal models of high-fat diet-induced obesity, type 2 diabetes, and nonalcoholic fatty liver disease.14–16
Some effects of cannabinoids are mediated by noncannabinoid receptors such as transient receptor potential (TRP) channels. 17 TRPV1, TRPV2, TRPV3, TRPV4, TRPA1, and TRPM8 are TRP channel subtypes that can be activated by cannabinoids and are defined as ionotropic cannabinoid receptors. 18 Mounting evidence suggests a significant contribution of TRPV1 in inflammatory lung diseases. The expression of TRPV1 in the airway epithelium increases in asthmatic patients and is more pronounced in severe uncontrolled disease. 19 However, the roles of TRPV1 and the effects of TRPV1 antagonism in asthma animal models are controversial. Delescluse et al. demonstrated that SB-705498 and PF-04065463, two antagonists of the TRPV1 channel, abolished histamine-induced airway hypersensitivity (AHR) in ovalbumin-sensitized guinea pigs. 20
In addition, Choi et al. showed that in BALB/c mice with asthma induced by ovalbumin, capsaicin reduced airway inflammation and remodeling by decreasing the levels of Th2 cytokines in bronchoalveolar lavage fluid (BALF) and epithelial cell-derived cytokines in lung tissue. 21 However, a study by Mori et al. indicated that the absence of these TRPV1 receptors increased Th2-biased immune response by enhancing serum IgE and BALF IL-4 and eosinophil levels following intranasal ovalbumin or house dust mite sensitization. 22 The outcomes of TRPV1 blockade vary under different experimental conditions.
In this study, we aimed to demonstrate the effects of selective CB1 agonist and antagonist on AHR and inflammation under the same experimental conditions using an ovalbumin-induced allergic asthma model in mice. For this purpose, the impact of the CB1 agonist arachidonoyl 2′-chloroethylamide (ACEA) at three different doses and the CB1 antagonist AM251 on allergic asthma was investigated using a distinct experimental timeline for modeling. In addition, nonreceptor mechanisms of ACEA were investigated by focusing on TRPV1 channels using the TRPV1 antagonist capsazepine.
Materials and Methods
Chemicals and reagents
Ovalbumin and aluminum hydroxide (Al(OH)3) were purchased from Sigma (Sigma–Aldrich Chemie GmbH, Germany). Methacholine, ACEA, AM251, and capsazepine were obtained from Cayman (Cayman). The ACEA and AM251 were initially dissolved in ethanol, and then, the solution was diluted to the required volume. The final solution had an ethanol concentration of 5%. Capsazepine was dissolved in saline.
Animals
One week before the experiment, female BALB/c mice (10 weeks, 25–35 g) were quarantined and acclimated. The study was carried out after the ethics committee approval of Adıyaman University (protocol no: 2020/015).
Mice were divided into 10 groups (n=7): Control, OVA, Vehicle (saline containing 5% ethanol), ACEA-0.5 (0.5 mg/kg ACEA), 23 ACEA-3 (3 mg/kg ACEA), 24 ACEA-5 (5 mg/kg ACEA), 25 AM251(3 mg/kg AM251), 26 AM251+ACEA (3 mg/kg AM251+3 mg/kg ACEA), CPZ (2.5 mg/kg capsazepine), 27 and CPZ+ACEA (2.5 mg/kg capsazepine+3 mg/kg ACEA).
Allergic asthma was induced in mice as described previously with modification. 28 Briefly, mice were administered 1 mg/kg ovalbumin adsorbed on 100 mg/kg Al(OH)3 intraperitoneally on days 0, 14, and 21; Al(OH)3 acts as an adjuvant to stimulate the production of immunoglobulin (Ig)E. On days 28, 29, and 30, mice were challenged by inhaling ovalbumin solution (1%, w/v in saline) for 20 min using a nebulizer (Omron NE-C28P, Japan/nebulization rate: 0.5 mL/min and particle size: 3.0 μm) (Fig. 1).
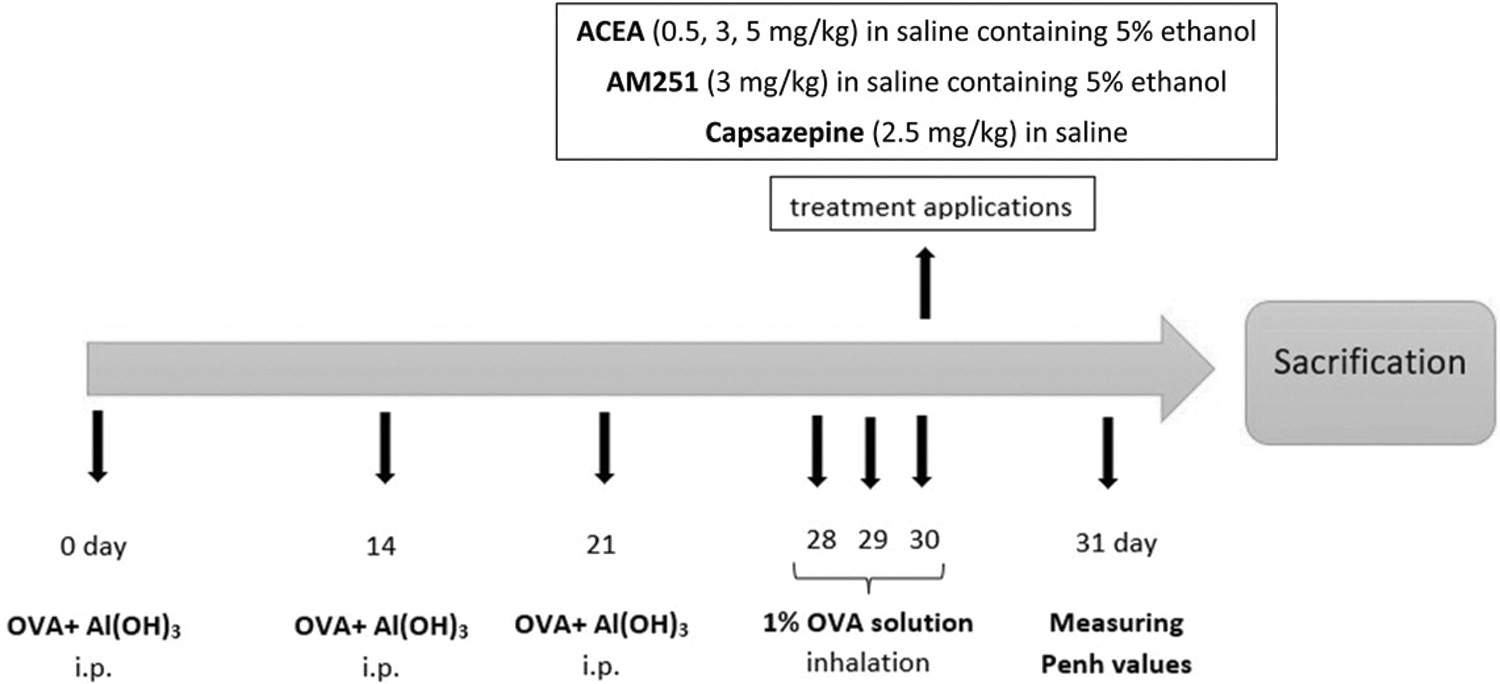
Experimental protocol.
Mice in the control group did not receive any treatment. In OVA group, mice sensitized and challenged with ovalbumin. Animals in the vehicle group were administered saline containing 5% ethanol after ovalbumin sensitization and provocation. For the treatment groups, on 30th day, before the third ovalbumin challenge, mice were injected intraperitoneally with ACEA (0.5, 3, and 5 mg/kg), AM251 (3 mg/kg AM251), and CPZ (2.5 mg/kg). Twenty-four hours after the last ovalbumin challenge, enhanced pause (Penh) values were measured by whole-body plethysmograph (EMKA Technologies, Paris, France) in mice. After a day, the mice were sacrificed, and samples were collected for subsequent experiments.
Measurement of lung function
Twenty-four hours after the last challenge, the mice were placed in whole-body plethysmograph and acclimatized for 10 min. Penh values were recorded for 3 min for each concentration by nebulizing increasing concentrations of methacholine (3.125, 6.25, 12.5, 25, 50, and 100 mg/mL). Penh values were calculated automatically using the iox2 software (iox2.10.0.40; EMKA Technologies, USA) according to the formula [(Te/Tr −1) (PEF/PIF)], where Te is the expiratory time, Ti is the inspiratory time, PEF is the peak expiratory flow, and PIF is the peak inspiratory time.
Serum cytokine levels
Mice were anesthetized with ketamine/xylazine (100/10 mg/kg), and after the cardiac puncture, blood samples were collected. Serum interleukin (IL)-4, IL-5, and IL-13 levels were measured using mouse Enzyme-Linked ImmunoSorbent Assay (ELISA) kits (Cloud-Clone Corp, Katy, TX), following the manufacturer's instructions. The amount of cytokine in the samples was calculated as ng/L using the standard curve.
BALF collection
To collect BALF, a cannula was placed into the trachea through tracheostomy and the left bronchus was ligated. The right lungs received 0.5 mL of saline and were gently massaged, and the BALF was removed via the injector. This process was repeated three times. The collected BALF was centrifuged at 4000 rpm at 4°C for 10 min, and the cell pellet was used for inflammatory cell count. The cell pellet was spread on slides and fixed with methanol. Preparations stained with Giemsa were examined under a light microscope at 40×objective magnification. Eosinophils, neutrophils, lymphocytes, and macrophages in 200 cells were counted and expressed as a percentage. 29
Histopathological analysis
Left lungs were isolated and fixed in 10% formaldehyde solution for histopathological analysis. Thin sections of 7 μm were cut and stained with Masson's trichrome and evaluated histopathologically with a light microscope and digital camera. Semi-quantitative evaluation according to the inflammation in the lung tissue was blindly performed by evaluating inflammatory cell infiltration, epithelial hyperplasia, alveolar septa thickening, and collapse regions. Scoring is done as follows: 0, no inflammation; 1, mild inflammation; 2, moderate inflammation; 3, severe inflammation.
Statistical analysis
Study data are demonstrated as mean±SEM. Statistical analysis was performed by using the SPSS software. Differences between groups were compared using the one-way analysis of variance test if the data were parametric and the Kruskal–Wallis test if they were not parametric. These tests followed by Bonferroni post hoc test for pairwise comparisons. Graphs were created using GraphPad Prism 9. Statistically, p<0.05 was considered to indicate a significant difference.
Results
Penh values
Sensitization and provocation with ovalbumin caused a significant increase in Penh values compared with the control group at all methacholine concentrations (p<0.05) (Fig. 2).
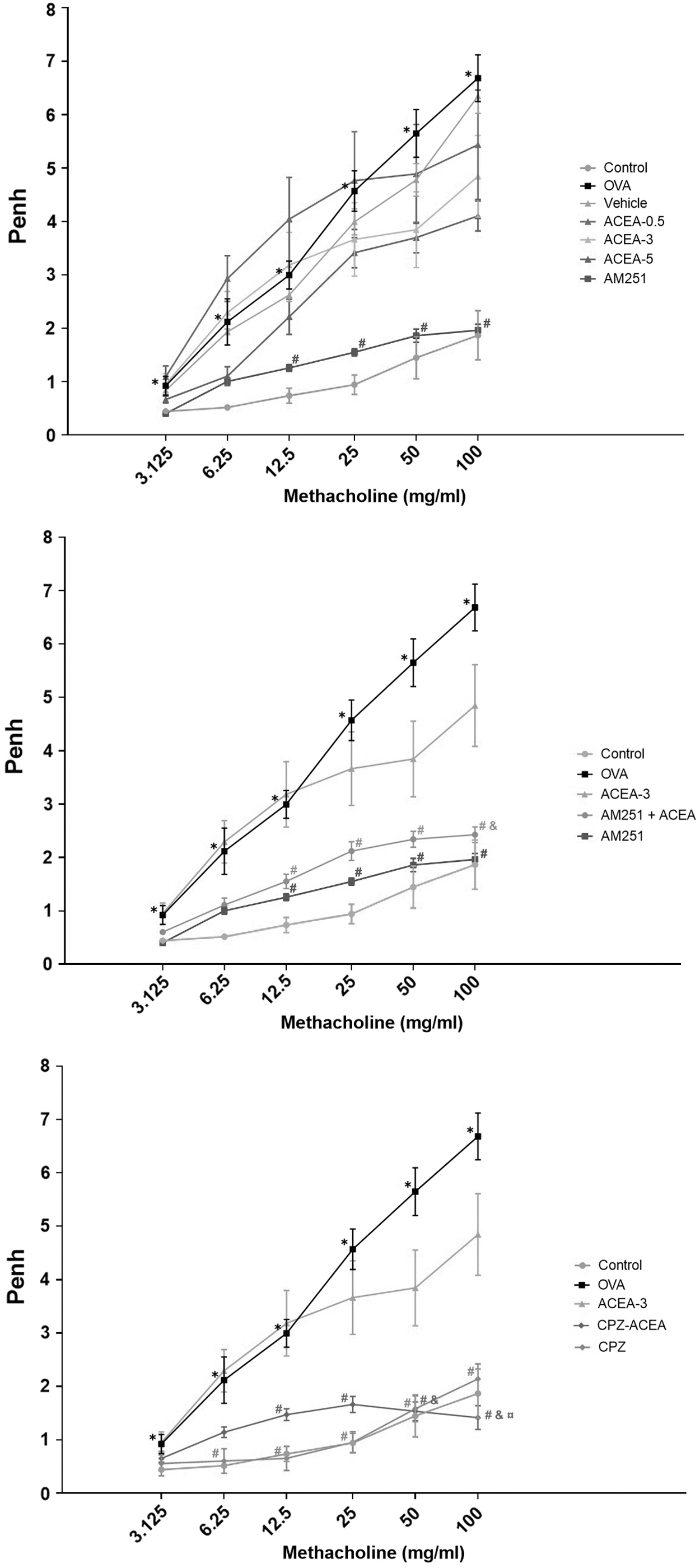
Enhanced pause (Penh) values. Control, normal control mice; OVA, ovalbumin-sensitized/challenged mice; Vehicle, saline containing 5% ethanol+ovalbumin-sensitized/challenged mice. ACEA-0.5, ACEA-3 and ACEA-5, ACEA (0.5, 3, or 5 mg/kg, respectively)+ovalbumin-sensitized/challenged mice; AM251, AM251 3 mg/kg+ovalbumin-sensitized/challenged mice; AM251+ACEA, AM251 (3 mg/kg), and ACEA (3 mg/kg)+ovalbumin-sensitized/challenged mice; TRPV1+ACEA, TRPV1 (2.5 mg/kg), and ACEA (3 mg/kg)+ovalbumin-sensitized/challenged mice. Values are expressed as mean±SEM. *p<0.05 compared with control. #p<0.05 compared with the OVA. &p<0.05 compared with ACEA-3. ¤p<0.05 compared with CPZ. ACEA, arachidonoyl 2′-chloroethylamide.
Penh values in the group administered the CB1 antagonist AM251 were significantly lower than the OVA group at 12.5, 25, 50, and 100 mg/mL methacholine (p<0.05). In the groups that were administered ACEA 0.5, 3, and 5 mg/kg, there was no significant difference in Penh values compared with the OVA group.
Compared with the OVA group, Penh values were significantly lower in the AM251+ACEA group at methacholine concentrations of 12.5, 25, 50, and 100 mg/mL (p<0.05). Although Penh values were higher in the AM251+ACEA group than in the AM251 group, there was no significant difference between the two groups.
Penh values of the CPZ and CPZ+ACEA groups were significantly lower than the OVA group at 12.5, 25, 50, and 100 mg/mL methacholine concentrations (p<0.05). Penh values of the CPZ+ACEA group were lower than those in the CPZ group at 100 mg/mL methacholine concentrations (p<0.05).
Serum IL-4, IL-5, IL-13 levels
The serum IL-4, IL-5, and IL-13 levels were higher in mice with allergic asthma model compared with the control group (p < 0.05) (Fig. 3).
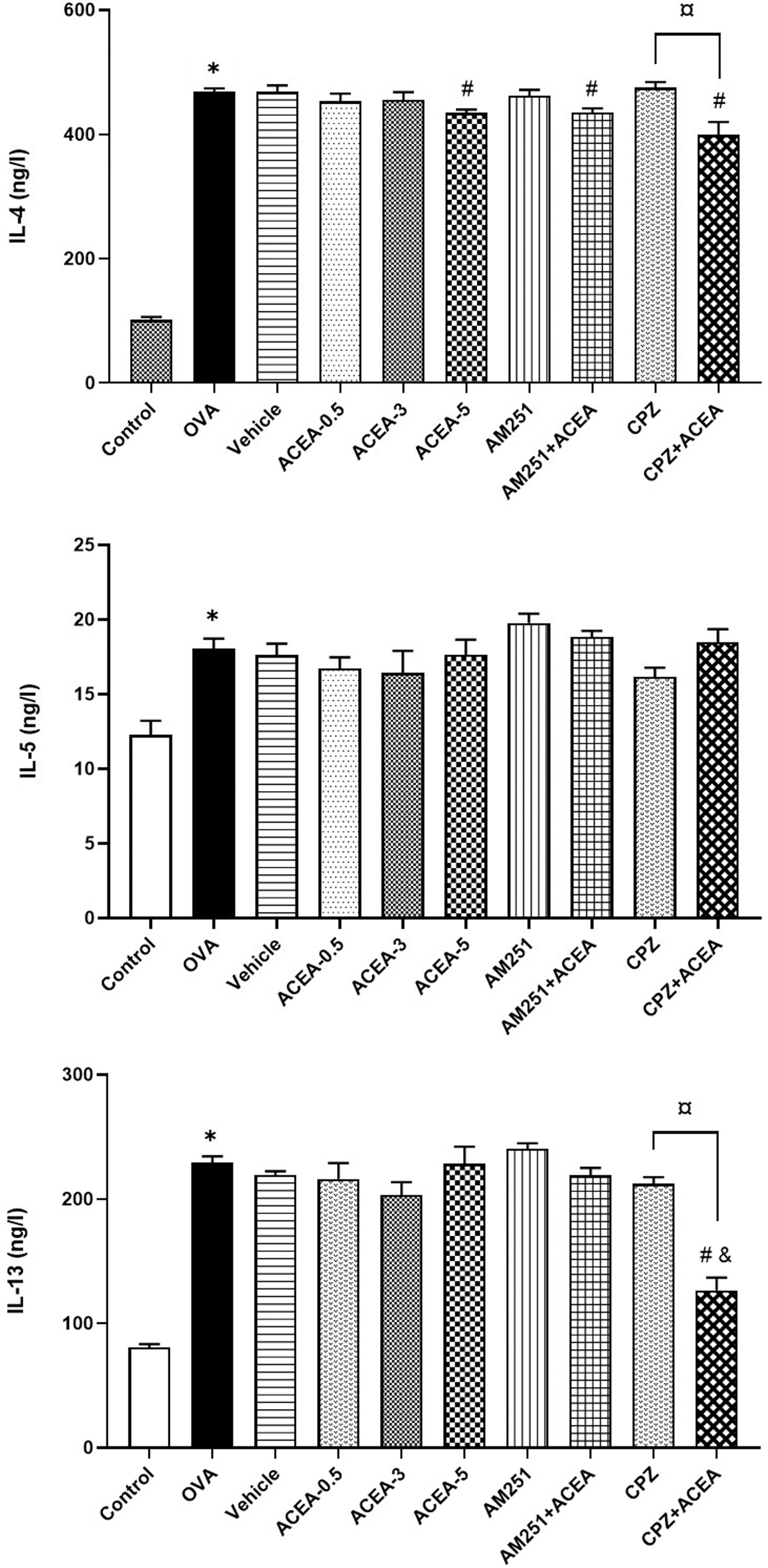
Serum IL-4, IL-5, and IL-13 levels. Control, normal control mice; OVA, ovalbumin-sensitized/challenged mice; Vehicle, saline containing 5% ethanol+ovalbumin-sensitized/challenged mice. ACEA-0.5, ACEA-3 and ACEA-5, ACEA (0.5, 3, or 5 mg/kg, respectively)+ovalbumin-sensitized/challenged mice; AM251, AM251 3 mg/kg+ovalbumin-sensitized/challenged mice; AM251+ACEA, AM251 (3 mg/kg), and ACEA (3 mg/kg)+ovalbumin-sensitized/challenged mice; TRPV1+ACEA, TRPV1 (2.5 mg/kg), and ACEA (3 mg/kg)+ovalbumin-sensitized/challenged mice. Values are expressed as mean±SEM. *p<0.05 compared with Control. #p<0.05 compared with the OVA. &p<0.05 compared with ACEA-3. ¤p<0.05 compared with CPZ. IL, interleukin.
When the CB1 agonist ACEA-treated groups were compared with the OVA group in terms of serum proinflammatory cytokines, only IL-4 levels were found to be significantly lower at high dose of ACEA, ACEA-5 (p<0.05). There was no statistical difference between the OVA and AM251 groups regarding serum IL-4, IL-5, and IL-13 levels.
Serum IL-4 levels of the AM251+ACEA group were lower than those of the OVA group (p<0.05); however, IL-5 and IL-13 levels were not statistically different. When IL-4, IL-5, and IL-13 levels were compared between the AM251 and AM251+ACEA groups, the differences for the three cytokines were not significant.
Serum IL-4 and IL-13 levels in the CPZ+ACEA group were significantly lower than those in the OVA (p<0.05) and CPZ (p<0.05) groups. There was no significant difference in terms of IL-5 levels (p>0.05).
Inflammatory cell levels in BALF
Sensitization and challenge with ovalbumin significantly increased eosinophil, neutrophil, lymphocyte, and macrophage levels in BALB/c mice (p<0.05) (Fig. 4).
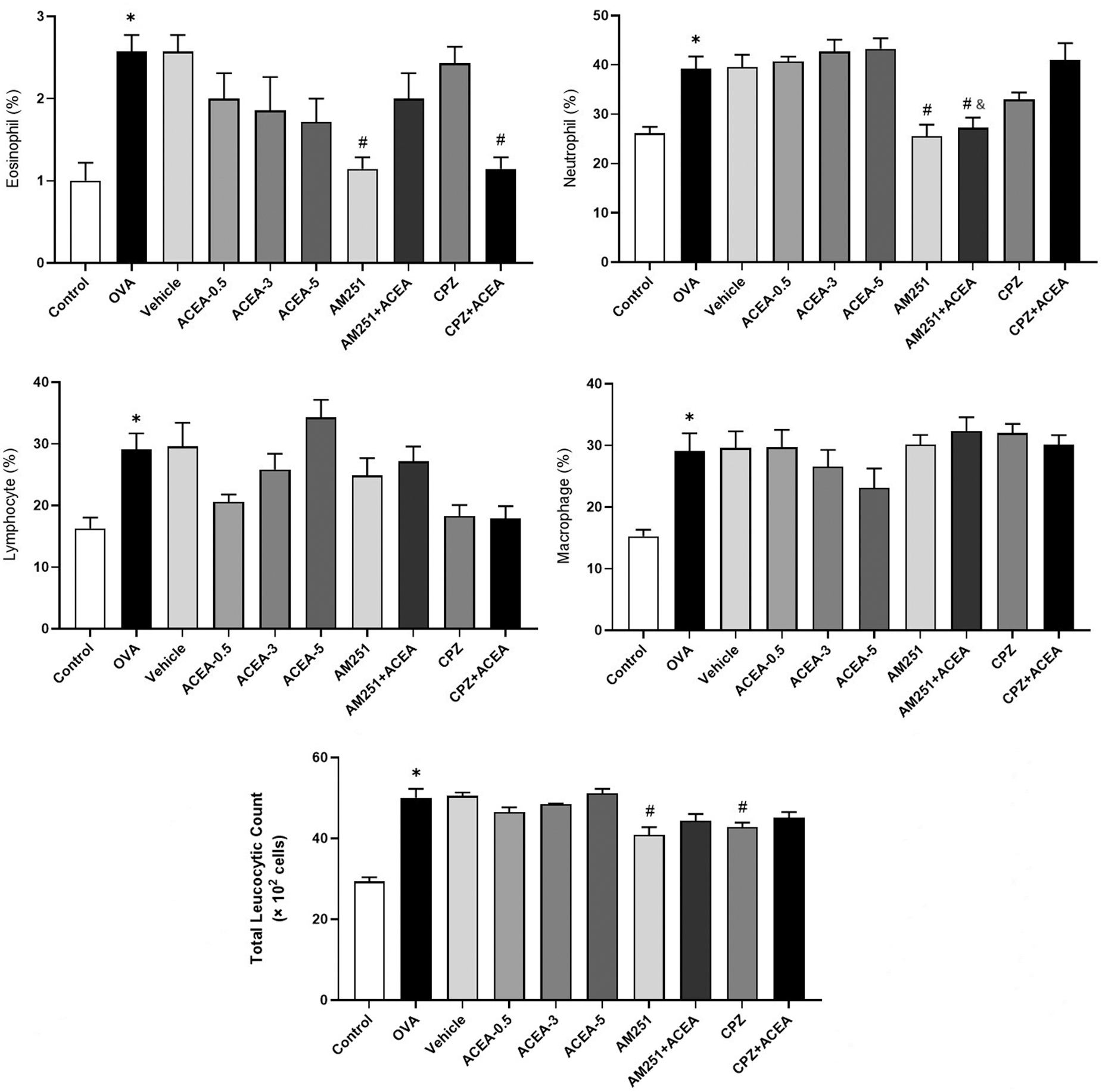
Eosinophil, neutrophil, lymphocyte, and macrophage levels, and total leukocyte count in BALF. Control, normal control mice; OVA, ovalbumin-sensitized/challenged mice; Vehicle, saline containing 5% ethanol+ovalbumin-sensitized/challenged mice. ACEA-0.5, ACEA-3 and ACEA-5, ACEA (0.5, 3, or 5 mg/kg, respectively)+ovalbumin-sensitized/challenged mice; AM251, AM251 3 mg/kg+ovalbumin-sensitized/challenged mice; AM251+ACEA, AM251 (3 mg/kg), and ACEA (3 mg/kg)+ovalbumin-sensitized/challenged mice; TRPV1+ACEA, TRPV1 (2.5 mg/kg), and ACEA (3 mg/kg)+ovalbumin-sensitized/challenged mice. Values are expressed as mean±SEM. *p<0.05 compared with Control. #p<0.05 compared with the OVA. &p<0.05 compared with ACEA-3. ¤p<0.05 compared with CPZ. BALF, bronchoalveolar lavage fluid.
The CB1 agonist ACEA demonstrated a dose-dependent decrease in eosinophil and macrophage counts. As the ACEA dose increased, neutrophil and lymphocyte counts increased. However, the changes were insignificant compared with the OVA group. The CB1 antagonist AM251 decreased eosinophil and neutrophil levels (p<0.05) in BALB/c mice sensitized/challenged with ovalbumin.
Compared with the OVA group, neutrophil levels were decreased in the AM251+ACEA group (p<0.05). BALF inflammatory cell levels were higher in the AM251+ACEA group compared with the group administered AM251 alone, but the difference was not statistically significant.
BALF eosinophil levels in the CPZ+ACEA group were lower than those in the OVA group (p<0.05). The CPZ and CPZ+ACEA groups were not significantly different when compared in terms of inflammatory cell levels in BALF.
Histopathological evaluation of lung tissue
Lung tissue inflammation was significantly increased in ovalbumin-sensitized/challenged BALB/c mice (p<0.05) (Fig. 5).
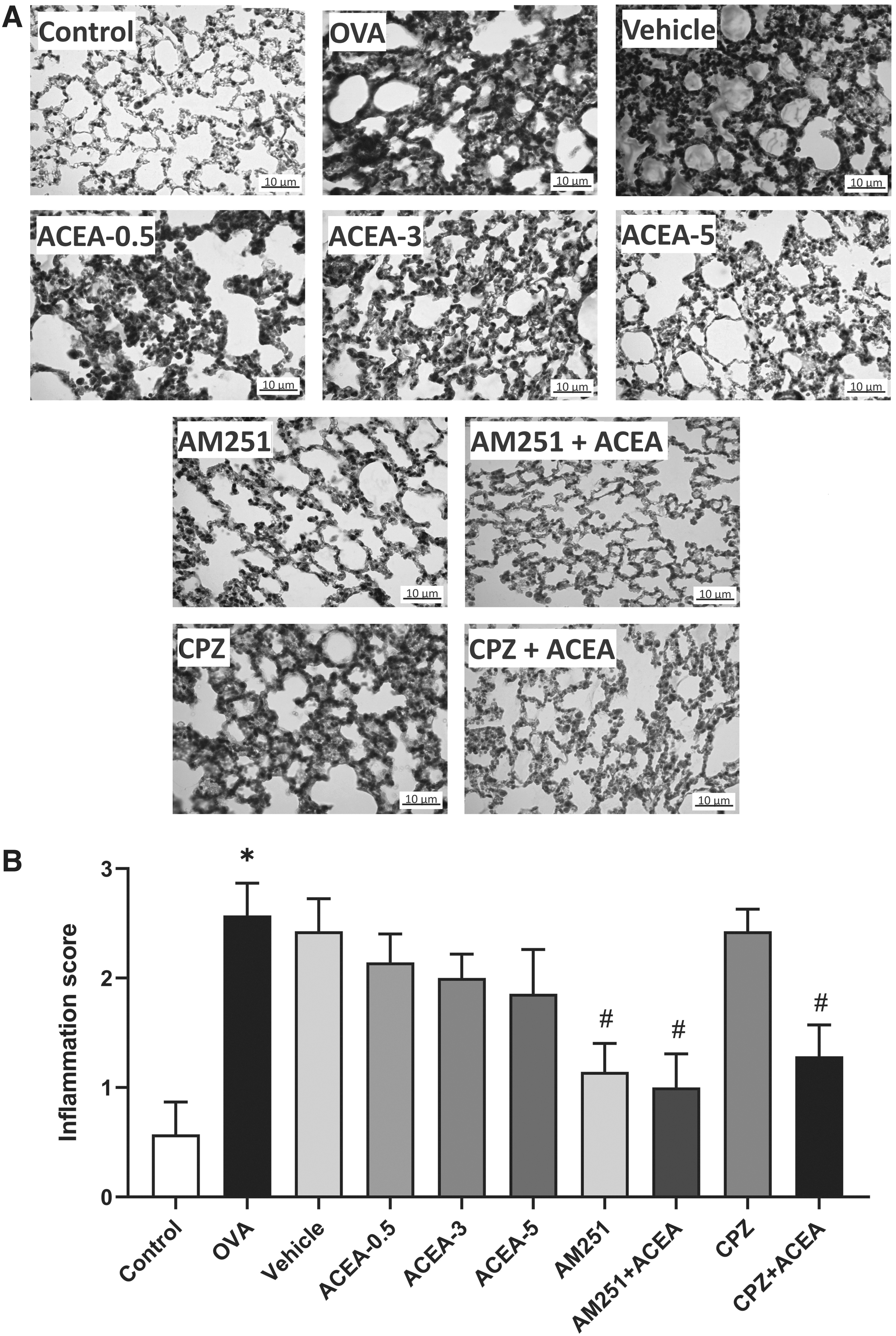
Histopathological evaluation of lung tissue. Alveolar septa stained with Masson's trichrome
Although the CB1 agonist ACEA treatment decreased the inflammation in the lung tissue depending on the dose, it did not create a significant difference with the OVA group. The administration of CB1 antagonist AM251 significantly reduced inflammation in the lung tissue compared with the OVA group (p<0.05).
Compared with the OVA group, the inflammation in the lung tissue was found to be significantly lower in the AM251+ACEA group (p<0.05). There was no statistically significant difference between the AM251 and AM251+ACEA groups.
In the CPZ+ACEA group, the lung tissue inflammation score was significantly lower than that in the OVA group (p<0.05). When the CPZ and CPZ+ACEA groups were compared in terms of lung tissue inflammation score, there was no significant difference.
Discussion
We aimed to elucidate the role of CB1 receptors in asthma pathology by exploring the effects of CB1 receptor agonist ACEA and antagonist AM251 in the identical conditions. We also inspected whether ACEA possess nonreceptor mechanisms, with a focus on TRPV1 channels. In this study, the effects of CB1 ligands in an animal model of allergic asthma were investigated by evaluating Penh values, serum proinflammatory cytokine levels, and inflammatory cell migration to the lung. The association with TRPV1 channels was examined by using a TRPV1 antagonist capsazepine. Our results show that the CB1 antagonist AM251 has the antiasthmatic effect by reducing AHR, and eosinophil and neutrophil migration to the lung. The CB1 agonist ACEA also tends to reduce AHR and inflammation, but not as much as AM251. ACEA may have CB1 nonreceptor mechanisms, but its effects on serum IL-4 and IL-13 appear to be independent of TRPV1 channels.
Pulmonary function tests are invaluable when investigating respiratory disease models. 30 Whole-body plethysmography gained popularity because it is noninvasive and can be used in unrestricted animals.31–33 In our study, Penh values, which are related but not a direct measure of airway resistance, were used to evaluate the effects of cannabinoid receptor ligands on AHR in ovalbumin-induced allergic asthma in BALB/c mice. 32
In asthma, excessive mediator release from inflammatory cells, increased contractility in airway smooth muscle, hypersensitivity of airway sensory nerves, and existing airway narrowing due to structural reasons are possible causes of AHR, but its mechanisms have not been fully understood. 2 In previous studies with nonselective cannabinoid receptor agonists, it was reported that the therapeutic effects on AHR were abolished by the administration of CB1 antagonists. Vuolo et al. displayed that cannabidiol, the main nonpsychoactive component of cannabis, reduced the airway resistance increased by methacholine challenge in ovalbumin-sensitized and provoked mice, and CB1 antagonist treatment has eliminated this effect. 7
In several studies using selective agonists, it was observed that specific stimulation of CB1 receptors may benefit asthma by reducing bronchoconstriction. In a study on experimental mice with dinitrofluorobenzene-induced nonatopic asthma, Bozkurt et al. stated that in vivo ACEA treatment as well as in vitro ACEA incubation prevented the increase in 5-hydroxytryptamine-caused contractions in the isolated trachea. 11 In our study, the CB1 agonist ACEA showed a slight decrease in Penh values, but this was not significant, probably because we used a lower dose than in the aforementioned study.
The nervous system is rich in CB1 receptors, which are highly localized in the presynaptic membrane. Thus, stimulation of these receptors may affect the regulation of synaptic release.34,35 It has been shown that ACEA diminishes the neuronal activity and intensity increase in NGF-induced airway inflammation in isolated mouse trachea and causes a direct inhibition of cholinergic contractions in the airways with a presynaptic mechanism. 12 We suggest that ACEA may reduce airway reactivity through a direct effect on presynaptic receptors. The effects of CB1 receptor antagonists on allergic asthma models have not been investigated. In our experiment, we administered AM251 to find out its effects on ovalbumin-sensitized and challenged BALB/c mice.
Evidence from previous studies showed a possible interaction of AM251 with postsynaptic beta receptors. Weresa et al. demonstrated that 1 μM AM251 exacerbated the increase in atrial rate caused by isoprenaline in isolated rat atria. At the same concentration, AM251 potentiated the positive inotropic effect caused by isoprenaline in the human atrium and the positive chronotropic and inotropic effects mediated by the B adrenergic receptor in hypertensive rats.36,37 In our study, the decrease in Penh values with AM251 was higher than ACEA, and it was significant. This pulmonary function-improving effect of AM251 may be due to its effect on Beta-2 (β2) receptors in the respiratory system.
Although the types of inflammation in asthma vary considerably, CD4+ T helper 2 (Th2)-mediated eosinophilic inflammation is the predominant inflammatory profile. In allergic asthma, Th2 cells secrete cytokines IL-4, IL-5, IL-9, and IL-13. These cytokines play a crucial role in the modulation of eosinophilic inflammation, and their levels are elevated in allergic asthma.1–3 IL-4 takes part in the activation of IgE and its production by B lymphocytes and contributes to the differentiation of naive Th cells into the Th2 phenotype. IL-5 is responsible for maturation, activation, and migration of eosinophils from the systemic circulation to the lung parenchyma. 38 IL-13 is a cytokine associated with the synthesis of IgE, chemoattraction of eosinophils, as well as mucus secretion, fibroblasts activation, and regulation of airway smooth muscle activity. 39
Vuolo et al. reported that cannabidiol decreased the levels of IL-4, IL-5, IL-13, and increased the level of IL-10, an anti-inflammatory Th2 cytokine, in serum of rats with ovalbumin-induced asthma. 6 In another research, it was shown that cannabidiol reduced the levels of IL-4, IL-5, IL-13, and eotaxin in lung homogenate and BALF obtained from BALB/c mice in which an experimental allergic asthma model was created. 7
However, in these studies, CB1 and CB2 were not targeted specifically to allocate their contribution in alleviating asthma features. Moreover, cannabidiol is a molecule that has pleiotropic effects and interacts with different receptors such as peroxisome-proliferator activated receptor γ, 33 which is also demonstrated to mediate anti-inflammatory activity in asthma. 34 We separately administered a CB1 agonist and antagonist to specifically define the role of activation and blockade of these receptors in terms of airway inflammation. Our results demonstrated that while AM251 did not change serum proinflammatory cytokine levels, ACEA reduced only serum IL-4 levels at the 5 mg/kg dose.
In an in situ study, it was stated that both the endocannabinoid anandamide and ACEA effectively prevented excessive human mucosal-type mast cells (hMMC) degranulation triggered by a secretagogue, and blockade of CB1 signaling with AM241 increased degranulation in hMMC without affecting proliferation. 40 The researchers concluded that endocannabinoids may control human hMMC activation and maturation via stimulation of CB1 receptors. It was found that IL-4 induced mast cell proliferation. 36 Therefore, we suggest that the activation of CB1 receptors also indirectly suppresses mast cells through reduction of IL-4.
In asthma, locally released chemotactic factors attract many inflammatory cells such as eosinophils, neutrophils, macrophages, mast cells, natural killer cells, dendritic cells, and T and B lymphocytes from the blood to the lungs. In addition, structural alterations including submucosal gland hyperplasia and an increase in goblet cells are observed in asthma. Type III and V collagen deposit beneath the true basement membrane causes subepithelial fibrosis and marked thickening of basement membrane and is related to eosinophil infiltration in patients with asthma. 2 In the study by Makwana et al., it was shown that Δ9-tetrahydrocannabinol (THC) inhibits lipopolysaccharide (LPS)-induced airway neutrophil recruitment. 41 In another study with THC, Braun et al. reported that the increase in the eosinophil count and the decrease in macrophage count observed in mice challenged and sensitized with ovalbumin were reversed by THC treatment. 42
The ability of CB1 antagonists to alleviate inflammation has also been demonstrated. In a model of type 2 diabetes, CB1 antagonist rimonabant has reduced the number of circulating neutrophils and monocytes. 16 Also, in a murine nonalcoholic steatohepatitis model, AM251 reversed the increase in macrophages and T helper cells in liver tissues. 43 In rats with LPS-induced inflammation, the CB1 antagonist-mediated anti-inflammatory effect has been shown to be inhibited using a selective β2-adrenoceptor antagonist. 44
In our study, AM251 decreased the amount of eosinophils and neutrophils in BALF and the lung's histological inflammation score. The mechanisms of antiasthmatic effect of AM251 may be mediated by β2 receptors in the respiratory system. Inflammatory cell counts in BALF and histological alterations in lung tissue were not significantly altered by ACEA. Penh values and BALF inflammatory cell levels tended to increase in the AM251+ACEA group compared with the AM251 group, implying that ACEA has non-CB1 receptor effects.
The sensory nerves of the respiratory system play a key role in the symptoms that are seen in asthma such as couch, dyspnea, and airway hyperreactivity. The stimulation of these nerves gives arise to cough, bronchoconstriction, chest tightness, and other vagal-mediated respiratory reflexes.20,45,46 TRPV1 channels that are highly expressed in sensory neurons and vagal afferent nerve endings a potential target for the treatment of airway hyperresponsiveness. 20 Enhanced expression of TRPV1 has been identified in the airway epithelium of individuals with asthma, particularly in cases characterized by severe and uncontrolled disease. 19 Choi et al. observed elevated levels of the Th2 cytokines IL-4, IL-5, and IL-13 in the BALF and the epithelial cell-derived cytokines thymic stromal lymphopoietin (TSLP), IL-33, and IL-25 in the lung tissue, as well as heightened expression of TRPV1 receptors in the lung tissue of ovalbumin-induced asthmatic BALB/c mice. 21
In addition, TRPV1 antagonists SB-705498 or PF-04065463 were also shown to significantly reduce histamine-provoked airway hyperresponsiveness in ovalbumin-sensitive guinea pigs.47–49 It is now known that TRPV1 channels are also expressed by T cells, mast cells, macrophages, dendritic cells, lung epithelial cells, and smooth muscle cells. They regulate inflammatory processes by the activation of CD4+ cells. They are activated by proinflammatory mediators such as lipooxygenase products, for example, leukotrienes. In a mouse model of ovalbumin-induced chronic asthma, Choi et al. have displayed that the administration of both a TRPV1 antagonist; capzasepine, and TRPV1 siRNA resulted in the attenuation of airway hyperresponsiveness and remodeling. They indicated that these effects were obtained by the reduction of the levels of Th2 cytokines; IL-4, IL-5 and IL-13, epithelial-derived cytokines; TSLP, IL-33 and IL-25, goblet cell hyperplasia, the increase in the α-smooth muscle actin, and collagen deposition. 21
Likewise, in our study, capsazepine administration significantly decreased Penh values compared with the disease group. Serum IL-5 and IL-13 levels and BALF neutrophil and lymphocyte counts also showed a tendency to decrease. In the CPZ+ACEA group, Penh values, serum IL-4 and IL-13, BALF eosinophil levels, and inflammation score in the lung tissue were significantly decreased compared with the disease group. When compared with the CPZ group, in the CPZ+ACEA group, Penh values were significantly increased and serum IL-4 and IL-13 levels were decreased. The rise in Penh values and the fall in IL-4 and IL-13 levels appear to be caused by ACEA's non-TRPV1-channel effects. These effects may be due to the effect of ACEA on TRPA1 channels, which are frequently expressed with TRPV1 channels. However, more research is mandated for this exclaim to be fully elucidated.
Findings of our experimental study demonstrate that the CB1 receptor blocking effect of AM251 significantly reduced asthmatic signs and symptoms, indicating that targeting CB1 receptors could be a promising therapeutic approach for asthma. The observation that the rise in serum cytokine levels caused by ovalbumin sensitivity and challenge did not exhibit statistically significant changes in the presence of AM251 may be associated with the temporal aspects of inflammatory cytokine release and its connection to systemic circulatory dynamics. Further exploration of the nuanced temporal dimensions of these processes will contribute to a more comprehensive comprehension of the observed effects. However, when the CB1 agonist ACEA was used, the effects on asthma parameters were mostly negative or ineffective. Since these negative effects were observed even under the blockage of CB1 receptors with AM251, it suggests that these unwanted effects might have occurred in a CB1 receptor-independent manner.
These effects do not seem to be mediated by TRPV1 receptors because blockade of TRPV1 receptors did not cause a significant change in most of the parameters evaluated according to CB1 agonist application. At this point, an important question arises: what is causing these effects? We speculated that TRPA1 channels may be involved in these pathophysiological processes, as they are typically coexpressed with TRPV1 and may have interactions with cannabinoids. To fully elucidate the involvement of CB1 receptors in the pathogenesis of asthma, further studies investigating their interactions with β-adrenergic 2 receptors and TRPA1 channels would be suggested.
In conclusion, the cannabinoidergic system seems to have a noticeable impact on allergic asthma. The blockade of CB1 receptors attenuated airway inflammation and hyperreactivity. Factors irrelative to CB1 receptors may have a role in the negative effects of ACEA on asthma. These effects do not appear to be associated with TRPV1 channels.
Footnotes
Author Disclosure Statement
No competing financial interests exist.
Funding Information
Support for this work was supplied by Ankara Yıldırım Beyazıt University research fund (project no: TDK-2020-2104).