
Abstract
Objective:
Cannabis sativa is the most used recreational drug worldwide. In recent years, there has been a growing interest in the potential therapeutic benefits of medicinal cannabis to treat a variety of psychiatric and neurological conditions. In particular, cannabidiol (CBD), a nonpsychoactive cannabis constituent, has been investigated for its potential prosocial effects on behavior, although the molecular mechanisms underlying this effect are still largely unknown. The aim of this study was to investigate the effect of a C. sativa oil CBD rich (CS oil) on social interaction and ultrasonic communication in mice.
Study Design:
Twenty-seven adult male mice (B6; 129P F2) were treated daily with vehicle or CS oil for 2 weeks. At Day 14, mice were tested for behavior (social interaction test and ultrasonic communication). Forty minutes before the behavioral tests, mice were exposed to intranasal treatment with vehicle or the oxytocin receptor antagonist, L-371,257. After behavioral tests, VH- and CS oil-treated mice were sacrificed, RNA was extracted from the hypothalamus and used for quantitative Real Time-PCR experiments.
Results:
We found that a 2-week treatment with the CS oil on mice exerted a prosocial effect associated with an increase in ultrasonic vocalizations. These effects were inhibited by pretreating mice with an oxytocin receptor antagonist. In addition, at the molecular level, we found that CS oil treatment caused a significant increase in oxytocin and a decrease in oxytocin receptor expression levels in the brain hypothalamus.
Conclusion:
Our results suggest that CS oil promotes social behavior by acting on oxytocin pathway.
Introduction
Phytoextracts derived from cannabis (Cannabis sativa L.) have been used for millennia for recreational and pharmaceutical purposes. 1 C. sativa displays different chemical constituents, such as terpens, flavoinoids, carbohydrates, fatty acids, and specific neuroactive compounds called cannabinoids. 2 To date, more than 100 cannabinoids have been identified, 3 acting on a broad neuromodulatory system known as the endocannabinoid system (ECS), which is involved in central nervous system development, neuronal activity and modulation, synaptic plasticity, cognitive processes, and a variety of other functions. 4 The more relevant phytocannabinoids are Δ9-tetrahydrocannabinol (THC), cannabidiol (CBD), and their carboxylic acid derivatives.
THC is a potent cannabinoid with psychoactive properties used in oncologic patients to alleviate chemotherapy-induced nausea and vomiting and for relieving pain.5,6 From a pharmacological point of view, it is a partial agonist of cannabinoids 1 and 2 receptors (CB1R and CB2R).
CBD is the major nonpsychoactive cannabis compound and displays a very low affinity for CB1R and CB2R. 7 CBD has also been defined as an allosteric negative modulator of CB1R. 8 Recently, it has been demonstrated also an allosteric negative modulation activity of CBD on CB2R. 9 In addition, other non-cannabinoid receptors were found as targets of CBD. 10 The therapeutic benefits of the use of CBD have been extensively investigated by different studies, where CBD has been proven to display therapeutic properties for a variety of psychiatric disorders having antioxidant, antitumoral, anti-inflammatory, neuroprotective, and pain-relieving properties.11–16 Recently, there has been a growing interest in the use of CBD as a therapeutic agent or as a coadjuvant treatment for symptoms and comorbidities related to neurodevelopmental disorders and in particular autism spectrum disorders (ASD).17–20
These and other cannabinoids can have a synergic action with other molecules contained in C. sativa and exert therapeutic effects for different psychiatric and neurological diseases. In particular, CBD has been investigated for its potential prosocial effects on behavior,18,21,22 although the molecular mechanisms underlying this effect are still largely unknown. So, in this work, we want to investigate the effect of a C. sativa oil CBD rich (CS oil) on social behavior in mice. For this reason, a chronic in vivo treatment with CS oil was carried out, and social interaction tests with ultrasonic communication analysis were performed.
Considering the key role of oxytocin in the brain in modulating social behavior,23–25 oxytocin pathway was also studied. The oxytocin is a neurohormone primarily involved in female lactation and uterine contractions during labor. 24 Oxytocin is synthesized in the hypothalamus’ paraventricular and supraoptic nuclei by magnocellular neurosecretory neurons that project to a variety of brain structures, indicating a widespread oxytocin system across brain regions such as thalamus, hippocampus, amygdala, and cortex. 26 Oxytocin is able to act as a key modulator of social behavior primarily through the activation of oxytocin receptors, generally coupled to Gq/11α GTP-binding proteins, in a tissue-dependent manner. Oxytocin receptors are expressed in all brain areas, with a higher receptor density in brain regions such as the olfactory bulbs, limbic cortex, amygdala, hypothalamus, cingulate cortex, hippocampus and brainstem.27,28 In this context, to evaluate the involvement of the oxytocinergic system, we treated mice with oxytocin receptor antagonist 29 and we analyzed their social interaction and ultrasonic vocalizations (USVs). We found that CS oil promoted social behavior and ultrasonic communication, and these effects were inhibited by pretreating mice with an oxytocin receptor antagonist.
Materials and Methods
Animals
Twenty-seven adult male mice (B6; 129PF2) were used as testing mice for our experiments (Supplementary Data S1). In addition, 27 male and 27 female adult mice (B6; 129PF2) were used as stimulus mice during social interaction tests (Supplementary Data S1). Animals were maintained in a 12 h light/dark cycle (light phase: from 8:00 a.m. to 8:00 p.m.) with food and water ad libitum. All experiments were performed according to European Communities Council Directive guidelines (CEE N° 86/609) for the care and use of experimental animals and all protocols were approved by the Animal Welfare Organization (Organismo Preposto al Benessere Animale) of the University of Brescia and by the Italian Ministry of Health (project code: 211B5.38).
Drug treatment
The mice were treated daily with vehicle (VH, each animal received about 150 µL of medium-chain triglyceride oil, GARDAJOINT S.R.L.S) or CS oil (each animal received about 150 µL of CS oil whose CBD content corresponded to 120 µg, ∼3.5 mg/kg, GARDAJOINT S.R.L.S.) for 2 weeks (Days 1–14). Composition of CS oil was previously characterized by our research group. 21 Briefly, this oil contains CBD (24%), THC (5%), and acid precursors CBDA (62%) and THCA (6%). All drugs were prepared daily and administered orally by gavage. At Day 14, mice were tested for behavior. Forty minutes before the behavioral tests, mice were exposed to intranasal treatment with vehicle (0.9% saline solution) or the oxytocin receptor antagonist, L-371,257 (Tocris Bioscience, Bristol, UK) at a dose of 300 µg/kg (Supplementary Data S1) (Fig. 1).
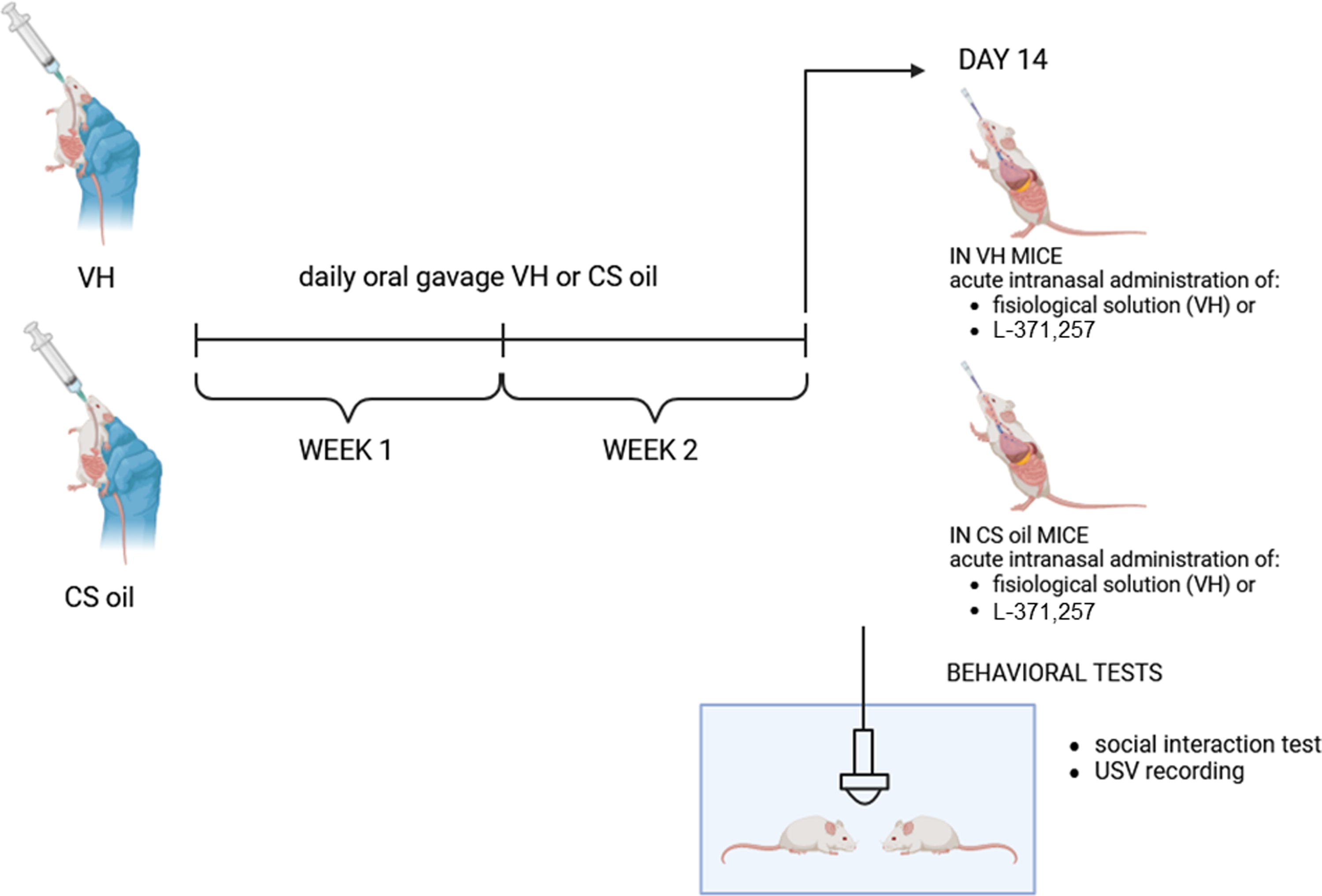
Schematic representation of the experimental design of the study. Daily oral gavage with vehicle (VH) or Cannabis sativa oil (CS oil) were given for 2 weeks (Days 1–14). On Day 14, mice were tested for behavior: social interaction tests and ultrasonic communication analysis. 40 min before the behavioral tests, mice received VH or CS oil with oral gavage and VH or L-371,257 (oxytocin receptor antagonist) with intranasal administration. After behavioral tests, mice were euthanized for molecular analysis.
So, there were four experimental groups: VH (n = 7), CS oil (n = 8), L-371,257 (n = 6), and CS oil + L-371,257 (n = 6) mice (Supplementary Data S1).
Behavioral analysis
Male–male social interaction
Male mice were isolated in their testing cage for 48 h as indicated in Silverman protocol 30 and tested for reciprocal social interaction. An unfamiliar stimulus male mouse, matched for age, was placed in the cage with the testing male mouse and left for 10 min as previously described. 31 Time spent doing social activities (sniffing of nose, sniffing of body or anogenital part of the partner, body contact and following) and nonsocial activities (cage exploring, self-grooming, and immobile) of testing mice was analyzed with Observer XT software (version 14.1, Noldus, Wageningen, The Netherlands) by an operator unaware of the experimental conditions of mice.
Male-female social interaction
Interaction of testing male mouse with an unfamiliar stimulus female mouse, matched for age, was video recorded for 5 min. Duration of social and nonsocial behaviors of testing mice was evaluated as described for male–male social interaction test. On the day of the test, the vaginal estrus phase of stimulus females was assessed as reported. 32 Females were half in diestrus and half in estrus phases, and they were distributed in a balanced way between experimental groups, as done previously. 33
Ultrasonic communication
During male–female social interaction test, ultrasonic communication was analyzed. An ultrasonic microphone UltraSoundGate Condenser Microphone CM 16 (Avisoft Bioacoustics, Berlin, Germany), suspended 20 cm above the cage of mice, recorded USVs emitted by male mouse during interaction with female mouse.34,35 These vocalizations are mostly produced by male mice to attract female mice and are the most extensively characterized in the literature.36–38
USVs were quantitatively analyzed in terms of number and duration of calls. Further, USVs were also classified into different types of calls according to Scattoni’s classification. 39 Two mice were excluded from USVs analysis because they did not vocalize (one VH- and one CS oil + L-371,257-treated mice).
Molecular analysis
RNA isolation from hypothalamus
After behavioral tests, VH- and CS oil-treated mice were sacrificed. The brains were removed and the hypothalamus isolated. Total RNA was extracted and quantified (Supplementary Data S1).
Quantitative Real-Time PCR
1 µg of total RNA from each hypothalamic tissue was transcribed into cDNA (Supplementary Data S1). Murine-specific primers, previously reported in the literature, 40 were used for hypothalamic tissues (Supplementary Data S1).
Statistical analysis
All the results are presented as the mean ± S.E.M. All statistical analyses were performed using GraphPad Prism 7 software (GraphPad Software, San Diego, CA). The data were analyzed by two-way analysis of variance (ANOVA) followed by Tukey’s multiple comparison test for social interaction tests and for analysis of USVs different types. One-way ANOVA with Dunnet’s multiple comparisons test was used for USVs quantitative analysis and one-way ANOVA with Tukey’s multiple comparison test for real-time PCR experiments. Statistical significance was indicated as *p < 0.05, compared with the control (VH) and §p < 0.05 compared with CS oil.
Results
Effect of CS oil on social behavior
To evaluate the effects of CS oil on sociability, male–male social interaction test was performed. As shown in Figure 2A, mice treated with CS oil spent significantly more time doing contact (mean ± SEM: 194.3 ± 9.3 s) compared with VH-treated mice (mean ± SEM: 132.8 ± 4.5 s). This social behavior was not associated with aggression, indeed no wrestling behavior was recorded in male mice tested. On the contrary, L-371,257 treatment reduced contact behavior (mean ± SEM: 73.8 ± 12 s) with respect to VH-treated mice. Interestingly, L-371,257 treatment not only induced a decrease in contact behavior but also reduced this social behavior in mice pretreated with CS oil (mean ± SEM: 104.1 ± 7 s) compared with CS oil-treated mice. In addition, acute L-371,257 treatment in CS oil-treated mice decreased also body sniffing (mean ± SEM: 25.8 ± 3.8 s) in comparison with CS oil-treated mice (mean ± SEM: 51 ± 6.5 s). Furthermore, the analysis of nonsocial behaviors, reported in Figure 2B, displayed that mice treated with CS oil spent less time doing exploring (mean ± SEM: 310.3 ± 15 s) than VH-treated mice (mean ± SEM: 386 ± 3.8 s). On the contrary, L-371,257 treatment caused a significant increase in exploring behavior both in CS oil pretreated mice (mean ± SEM: 442 ± 10.5 s) and in mice treated with L-371,257 alone (mean ± SEM: 471.2 ± 12.4 s).

Effect of Cannabis sativa oil (CS oil) treatment on social behavior in male–male social interaction test. Graphic representation of data collected during the male–male social interaction test by measuring the time in seconds
The results obtained support a prosocial effect of CS oil that was further confirmed by male–female social interaction test (Fig. 3). Indeed, male mice treated with CS oil spent more time doing social activities, such as ano-genital sniffing of females (mean ± SEM: 101.8 ± 7.6 s) than VH-treated mice (mean ± SEM: 67.7 ± 7 s). Instead, L-371,257 treatment reduced this social behavior both in mice treated only with oxytocin receptor antagonist and in CS oil pretreated mice (mean ± SEM: 46.6 ± 9.5 s and 51.2 ± 5.9 s, respectively) in comparison with VH- and CS oil-treated mice (Fig. 3A). As nonsocial activities, mice treated with CS oil spent less time performing exploring (mean ± SEM: 74.2 ± 23.6 s) than VH-treated mice (mean ± SEM: 148.9 ± 13.2 s) (Fig. 3B). Treatment with L-371,257 in CS oil pretreated mice caused a significant increase in time spent exploring (mean ± SEM: 175.9 ± 8 s) compared with CS oil-treated mice (Fig. 3B).
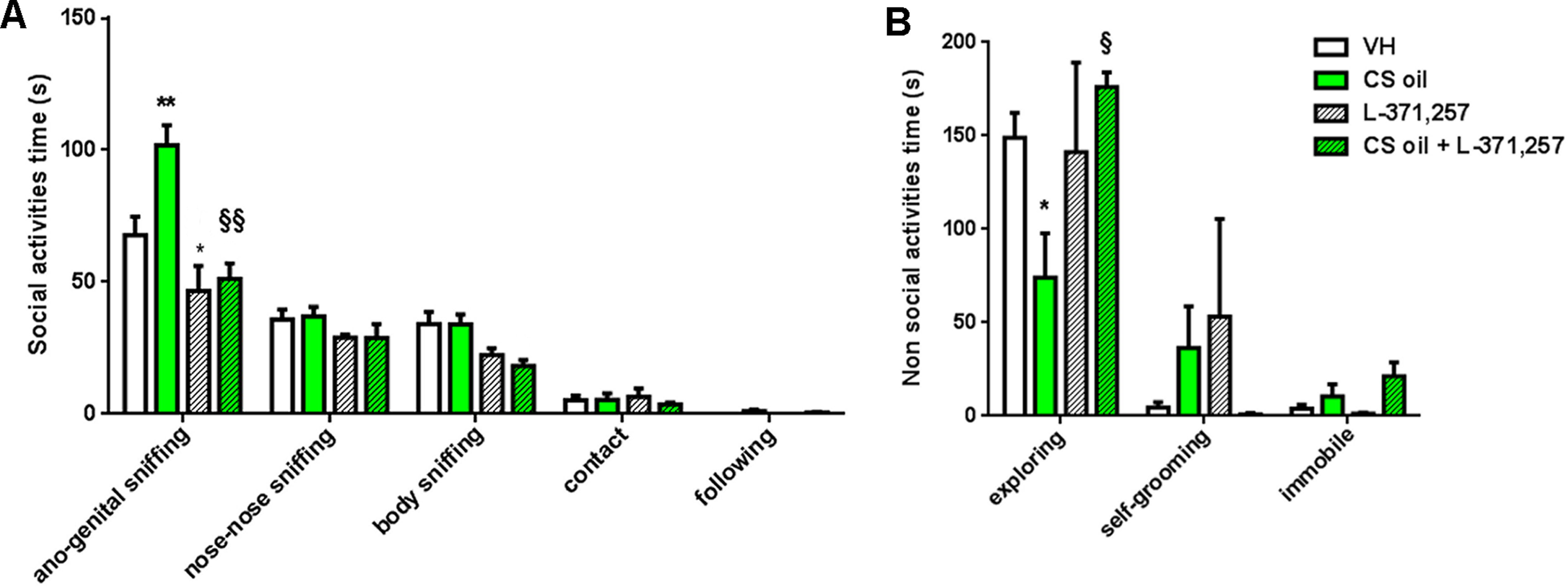
Effect of CS oil treatment on social behavior in male–female social interaction test. Graphic representation of data collected during the male–female social interaction test by measuring the time in seconds (s) spent by mice in doing social
Effect of CS oil on ultrasonic communication
During the male–female social interaction test, USVs emitted by male mice were recorded and analyzed. As reported in Figure 4A, CS oil treatment caused a significant increase in the number of USVs (mean ± SEM: 689.8 ± 177.9) compared with VH-treated mice (mean ± SEM: 218 ± 120.6). In addition, CS oil-treated mice displayed a tendency to emit calls with a longer duration (mean ± SEM: 47.6 ± 5.8 msec) than VH-treated mice (mean ± SEM: 23.3 ± 7.4 msec). Interestingly, L-371,257 treatment prevented the effect of CS oil in terms of increase in number of calls, indeed mice treated with L371,257 and CS oil emitted a lower number of USVs (mean ± SEM: 71.4 ± 65.9) than CS oil-treated mice.

Effects of CS oil treatment on mice communication.
Furthermore, a detailed qualitative analysis was performed in mice based on 1090 calls for VH-treated mice, 4139 calls for CS oil-treated mice, 439 calls for L-371,257-treated mice, and 357 calls for CS oil + L-371,257-treated mice (Fig. 4B). All these USVs were classified into 10 categories, as previously reported by Scattoni. 41 As shown in Figure 4B, CS oil-treated mice emitted a significantly higher number of “upward,” “harmonics,” and “frequency steps” (mean ± SEM: 133.8 ± 35.9, 97 ± 42.5, and 192.3 ± 61.5, respectively) compared with VH-treated mice (mean ± SEM: 57.8 ± 30.7, 10 ± 6.4, and 49.8 ± 32.6, respectively). Oxytocin receptor antagonist treatment prevented the increase of these typologies of calls emission induced by CS oil treatment; indeed, L-371,257 treatment in CS oil pretreated mice caused a reduction of “upward”, “harmonics” and “frequency steps” (mean ± SEM: 7.6 ± 6.1, 8.8 ± 8.3, and 6.4 ± 6.2, respectively) compared with CS oil-treated mice. In parallel, the duration of “harmonics” and “frequency steps” resulted increased in CS oil-treated mice (mean ± SEM: 6.8 ± 3 and 15.8 ± 5.2 msec) compared with VH-treated mice (mean ± SEM: 0.6 ± 0.4 and 3.7 ± 2.7 msec). L-371,257 treatment caused a decreased duration of these two types of calls in CS oil-pretreated mice (mean ± SEM: 0.7 ± 0.7 and 0.5 ± 0.5 msec) compared with CS oil-treated mice.
Gene expression in mice brain hypothalamus
As shown in Figure 5, mice treated with CS oil displayed a significant increase in hypothalamic levels of oxytocin mRNA vs VH (mean ± SEM: 8.5 ± 1.6 and 1.1 ± 0.2, respectively) and a decrease in levels of oxytocin receptor mRNA in comparison with VH-treated mice (mean ± SEM: 0.5 ± 0.1 and 1.0 ± 0.1, respectively). L-371,257 treatment did not cause any change in expression levels of oxytocin (mean ± SEM: 1.0 ± 0.1) and its receptor (mean ± SEM: 1.0 ± 0.1) compared with VH-treated mice. In addition, treatment with L-371,257 caused a significant reduction in oxytocin mRNA levels in CS oil-pretreated mice (mean ± SEM: 0.5 ± 0.1) in comparison with CS oil-treated mice. A significant increase of oxytocin receptor expression levels was observed in CS mice treated also with L-371,257 (mean ± SEM: 1.7 ± 0.1) compared with CS oil-treated mice.
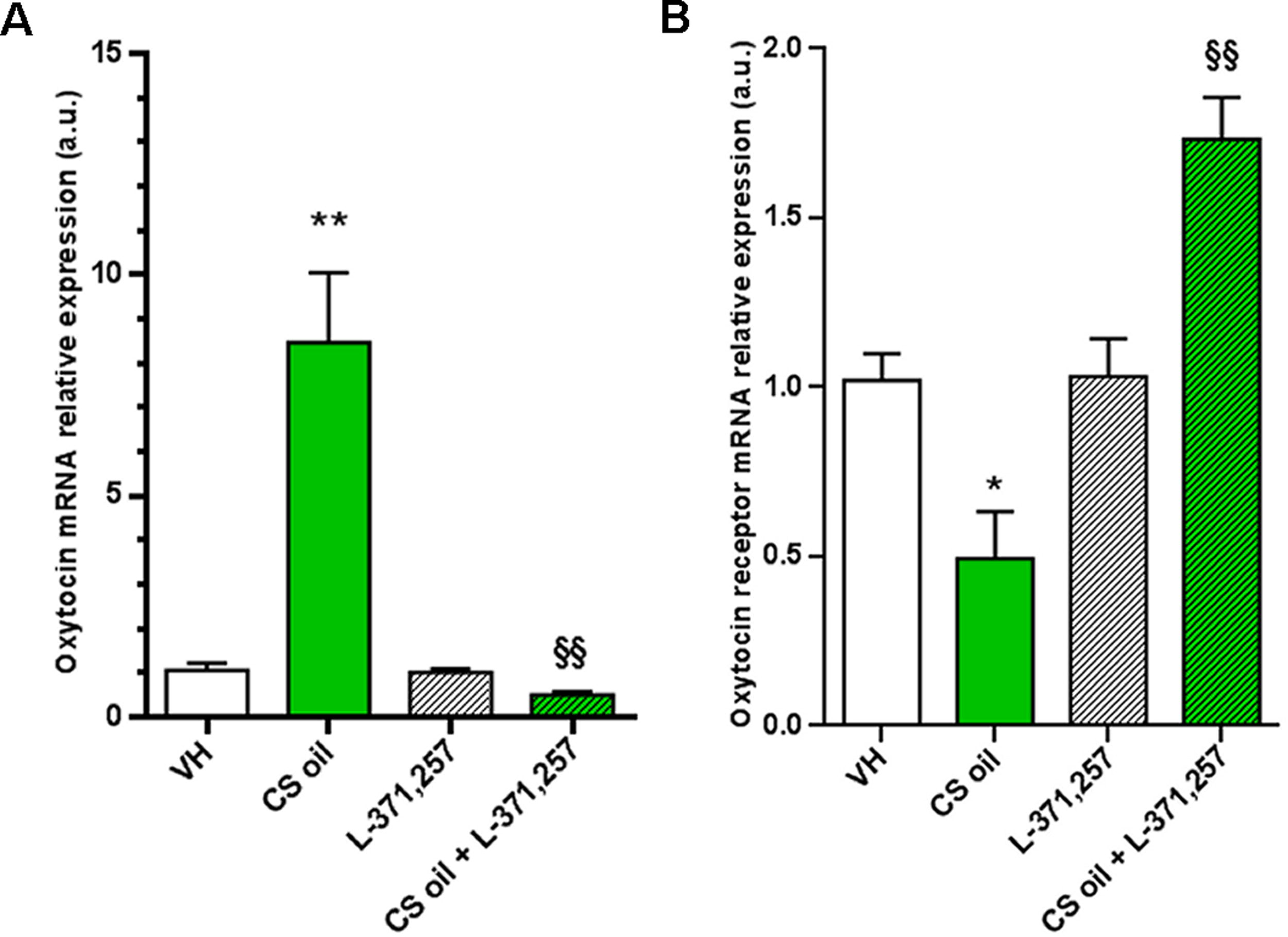
Gene expression in mice brain hypothalamus. Graphic representation of data collected on mRNA levels of oxytocin and oxytocin receptor quantified in mice hypothalamus at the end of the 2 weeks CS oil treatment by quantitative RT-PCR. Data are presented as mean ± SEM.
Discussion
The present study investigated the effects of C. sativa oil CBD rich on the social behavior of mice and the involvement of the oxytocinergic pathway in mediating CS oil effects.
On the behavioral level, we found that (1) CS oil had a significant effect in promoting social behavior in mice, identified with an increase in time spent doing social activities; (2) the oxytocin receptor antagonist L-371,257 alone was able to induce a negative effect on social behavior, supporting the key role of oxytocin signaling in modulating this behavior; (3) the L-371,257 treatment in CS oil pretreated mice prevented the CS oil-dependent prosocial effect; and (4) CS oil treatment stimulated mice communication through the oxytocin pathway, since CS oil treatment induced a significant increase in number of USVs compared with VH, and this effect was blocked by treatment with L-371,257.
Concerning social behavior, different studies have supported the key role of oxytocin in improving social behavior, and this effect seems to involve the melanoninergic 42 and endocannabinoid pathways.41,43 Indeed, as previously demonstrated by our research group, 42 treatment with THIQ, an agonist of the melanocortin 4 receptor, provoked an increase in social interaction in mice and this effect was prevented by an oxytocin receptor antagonist treatment. In addition, Wei and colleagues 43 reported an increment in social contact induced by anandamide-mediated signaling at CB1 receptors, and this effect was blocked by oxytocin antagonists.
Beyond the analysis of social behaviors, also USVs were studied. Increased sociability observed after CS oil treatment can be associated with the increased ultrasonic communication found in mice. Indeed, we reported that CS oil-treated mice vocalized more than VH-treated mice, and this effect was blocked by an oxytocin receptor antagonist. Detailed qualitative analysis of USVs displayed that CS oil-treated mice emitted higher number of “upward,” “harmonics,” and “frequency steps” with a longer duration of these types of calls in comparison with VH mice. So, the effect of CS oil on ultrasonic communication is not generalized but call-specific. Oxytocin receptor antagonist reduced the emission of these same typologies of calls compared with CS oil-treated mice. Interestingly, the number and duration of “frequency steps” calls have been found altered also in a murine model of neurodevelopmental disorders characterized by social deficit. 44 This can suggest a possible association between this specific typology of calls and social behavior and could have an essential role in the context of USVs studies.45,46
After behavioral studies, mRNA expression levels of key genes involved in oxytocin pathways were evaluated. Our results showed that chronic CS oil treatment induced an increase in oxytocin mRNA levels compared with VH-treated mice at the hypothalamic level, a crucial brain region of the social reward circuit,47,48 accompanied by a decrease in the levels of the oxytocin receptor, hence suggesting a downregulation mechanism induced by a robust oxytocin release and receptor stimulation. Oxytocin receptor antagonist alone did not cause modulation of expression levels of oxytocin and its receptor compared with VH. On the contrary, a significant increase of oxytocin receptor expression levels was observed in mice cotreated with CS oil and L-371,257 vs CS oil-treated mice. Since the treatment with L-371,257 was carried out acutely, it is difficult to think that this increase in mRNA levels is linked to the phenomena of upregulation of the receptor. However, it must be considered that oxytocin signaling is regulated very quickly, as it is a signal that is released on demand and which also inactivates very quickly. Therefore, we hypothesize that this regulatory effect could have occurred a short time after acute treatment (few hours).
Taken together, our results demonstrate that CS oil exerts a prosocial effect in mice and that these effects are mediated by the oxytocin pathway. However, our study reports some limitations that must be addressed. First of all, the use of a single mouse model to investigate sociability, can represent a limitation in the generalizability of the findings. In the future, we will test CS oil on other mouse models to verify the prosocial effect is not strain dependent. Another point is the omission of female subjects in the experiments. Indeed, we examined only male mice in our study, but we believe that the importance of testing the effects of CS oil on both genders is fundamental. So, in the future, we will include also females to enhance the robustness and applicability of the findings, allowing for a more comprehensive evaluation of the effects of CS oil on sociability.
Finally, since the CS oil used in the study is rich in CBD but contains also other different substances (e.g., terpenes, flavonoids, and other phytocannabinoids), it is plausible that these other substances played a role in the prosocial effect observed in vivo acting with an “entourage effect”. In this regard, Staben and colleagues 22 have found that inhalation of cannabis terpene blends increased social behavior in a mouse model of ASD. Interestingly, the prosocial effect of cannabis-inspired terpene blends was more robust when combined with CBD. This suggests that a combination of phytochemicals can improve therapeutic benefits than individual chemicals.22,49,50
In conclusion, the present study sheds light on the mechanism underlying the prosocial effects of CS oil involving the oxytocin pathway, providing cues regarding the possible molecular processes supporting the employment of both cannabis and oxytocin-based therapies in disorders characterized by social impairments such as ASD.
Footnotes
Data Availability Statement
Data are contained within the article or supplementary material.
Acknowledgments
Author Contributions
M.P.: methodology; validation; investigation; writing—original draft; writing—review and editing. M.C.: writing—original draft; writing—review and editing. A.M.: conceptualization; validation, investigation; writing—review and editing. G.M.: investigation; formal analysis. F.A.: investigation; formal analysis; data curation. E.M.S.: investigation; formal analysis. M.M.: conceptualization; writing—review and editing; supervision; funding acquisition. S.A.B.: conceptualization; validation; writing-original draft; writing—review and editing; project administration.
Author Disclosure Statement
The authors declare that there is no conflict of interest.
Funding Information
This study was supported by a grant from the
Abbreviations Used
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.