
Abstract
Objective:
The most commonly reported primary brain tumor in adults is glioma. Choline kinase alpha (CHKA) has been proved to play important roles in glioma. However, the mechanism of CHKA involved remains unclear. Therefore, this study aims to explore the mechanism of CHKA in glioma development.
Methods:
Immunohistochemistry, qRT-PCR, and Western blot were used to detect the expression of CHKA. Flow cytometry, Cell Counting Kit-8 (CCK-8), transwell, and wound healing assays were performed to evaluate cell apoptosis, proliferation, invasion, and migration, respectively. RNA sequencing was used to explore the differentially expressed genes affected by CHKA. The enrichment analysis of gene ontology (GO) and Kyoto Encyclopedia of Genes and Genomes (KEGG) helped to detect the signaling pathways CHKA affected. Tumor-bearing mice were established and evaluated by TUNEL assay, Ki-67 immunohistochemistry. and hematoxylin and eosin staining.
Results:
CHKA increased in glioma tissues and promoted cell proliferation, invasion, and migration, while inhibiting the glioma cell apoptosis. It was also showed that CHKA promoted glioma development in vivo. GO and KEGG analysis indicated that PI3K/AKT was significantly enriched in CHKA knockdown U251 cells. And CHKA promoted glioma development by activating PI3K/AKT signaling pathway.
Conclusions:
The authors demonstrated that CHKA was significantly elevated in glioma tissues. Mechanism analysis indicated that CHKA could promote glioma development by activating PI3K/AKT signaling pathway, suggesting that CHKA is promising to be a biomarker and therapeutic strategy for prognostic prediction of patients with glioma.
Keywords
Introduction
The most commonly reported primary brain tumor in adults is glioma, accounting for 60%–70% of neuroepithelial tumors.1,2 In terms of the histological type of glioma, the glioma is divided into four subtypes, including grade I, grade II, grade III, and grade IV, representing pilocytic astrocytoma, diffuse astrocytoma, anaplastic astrocytoma, and glioblastoma, respectively.3,4 Glioma infiltrates extensively during the malignant process, and it is difficult to remove them completely by surgery. The postoperative recurrence rate is high, and the treatment effect is poor due to the existence of glioma stem cells and extensive infiltration. 5 The current treatment principle of glioma is still surgery, supplemented by radiotherapy and chemotherapy. However, combination therapy can only prolong the survival time of patients with high-grade glioma for 2–4 months.6,7 Therefore, a better understanding of the molecular mechanism of glioma is of great significance for developing new targeted treatment strategies and methods.
Choline kinase alpha (CHKA) belongs to the family of choline kinase proteins and is a key enzyme in the CDP-choline pathway. 8 The expression level of CHKA elevated in various tumors, such as colorectal cancer, breast cancer, prostate cancer, and lung cancer. 9 For instance, Trousil et al. depicted that the metabolism of choline phospholipid altered in endometrial carcinoma, which was induced by overexpressing CHKA and activating deacylation process, suggesting that choline biochemistry could serve as a vital noninvasive biomarker for endometrial cancer. 10 Hu et al. revealed that CHKA overexpression contributed to tumor progression and metastasis, and served as a poor prognosis predictor in colorectal carcinoma. 11 Rizzo et al. demonstrated that CHKA downregulation suppressed the aggressive ovarian cancer cellular behaviors significantly, and also affected cell sensitivity response to drugs, suggesting that CHKA could act as a new target in epithelial ovarian cancer therapy. 12 These results demonstrated that CHKA may exert vital effects in glioma progression due to its high expression and abnormal activation in tumor tissues.
In the previous study, the authors demonstrated that knockdown of CHKA inhibited the proliferation, invasion, and migration of human U87MG glioma cells. 13 However, the signaling pathway that CHKA was involved in remains unclear. Therefore, this study aimed to explore the mechanism of CHKA in regulating glioma development.
Materials and Methods
Patients and specimens
All glioma samples were collected from 75 patients who underwent routine surgery in the General Hospital of Ningxia Medical University from March 2019 to December 2020, including 37 grade II patients, 13 grade III patients, and 25 grade IV patients, without receiving chemotherapy before the surgery. The clinical data of patients were specified in Table 1. All the patients participating in this study were formally informed of the purpose of using the sample and signed an informed consent form. This study was approved by the Ethics Committee in the General Hospital of Ningxia Medical University. The approval number was (2019NX-YK007A).
Clinicopathological Features of 75 Patients with Glioma
Cell culture and treatment
The two glioma cell lines, including U251 and U87, were obtained from ATCC and they were cultured in DMEM supplemented with 10% FBS in 5% CO2 at 37℃C. For cell transfection, U251 and U87 were inoculated in six-well plates and incubated for 48 h till the cells reached at 70%–90% confluence. And then the overexpressing and suppressing CHKA plasmids were transfected into the glioma cell lines U87 and U251 with Lipofectamine 3000 Transfection Reagent (L3000001; Thermo Fisher). For RNA sequencing, the U251 cells were treated with silent CHKA lentivirus. Briefly, culture well-growing U251 cells (density 5 × 104 cells/mL) and inoculate them into six-well plates. Dilute the lentivirus into CHKA silencing lentivirus and its control lentivirus with a concentration of MOI = 10 with DMEM according to the instructions. Add 1 mL of the diluted lentivirus and dye assistant (50 μg/mL) to each well. Gently mix and put it in the incubator to continue culture for 12 h. Replace the complete medium, continue to culture for 12 h, observe the cell transfection with a fluorescence microscope, and ensure that the infection efficiency reaches more than 90%. The transfected U251 cells were sent to GENE company for sequencing.
Immunohistochemistry
The collected tumor tissues were fixed with formalin, and then embedded in paraffin. Next, the samples cut into slices (3 μm) before antigen retrieval with 10 mM citrate buffer for 20 min. After blocking endogenous peroxidase activity by 0.3% hydrogen peroxide for 10 min, the slices were incubated with primary antibody (1:200, ab88053; Abcam) at 4°C overnight. Then the secondary antibody (Dako) was incubated with the samples for 30 min after washing. The slides were washed with phosphate buffered saline (PBS), and reacted with DAB at RT. Next, the samples were counterstained with hematoxylin before being dehydrated by gradient ethanol solution. The images were photographed by a microscope (Olympus, Japan).
qRT-PCR
The total RNA was obtained from tissue cell lines with or without treatment or tissues by a TRIZOL kit (Ambion, Huntingdon, UK) following the product directions. Then the quantity of RNA was detected using NanoDrop one (Thermo Fisher Scientific, Inc.). Reverse transcription was performed with a SuperScript™ II Reverse Transcriptase Kit (Thermo, Bremen, Germany). qRT-PCR was conducted using SYBR Green qRT-PCR Kit (Roche) on an ABI7500 System (Applied Biosystems). The amplification reaction process is as follows: (1) 94°C for 10 min and (2) 94°C for 15 s and 59°C for 20 s (40 cycles). GAPDH was used as the internal reference. Gene expression was analyzed by 2−ΔΔCT method. The primer sequences used in qRT-PCR are shown as follows: CHKA forward: 5′-AGTTTTTGGCTTCCAGCAGA-3′, reverse: 5′-ACATTAGTCATGGTCACGCG-3′; and GAPDH forward: 5′-TGCACCACCAACTGCTTAGC-3′, reverse: 5′-GGCATGGACTGTGGTCATGAG-3′.
Western blot analysis
The total protein from cells and tumor tissues was obtained by Total Protein Extraction Kit (KeyGEN BioTECH Co.). Protein concentrations were quantified with the BCA Protein Assay Kit (KeyGEN BioTECH Co.). Twenty micrograms protein was loaded into the lane of SDS-PAGE (10%; Beyotime) and separated. Then they were transferred onto polyvinylidene difluoride (PVDF) membranes (Millipore), before blocking with 5% non-fat milk dissolved in PBS. The PVDF membranes were, respectively, incubated with anti-AKT (1:1000, ab8805; Abcam, Cambridge MA), PI3K (1:1000, ab32089; Abcam), p-AKT (1:1000, ab38449; Abcam), p-PI3K (1:500, ab278733; Abcam), and GAPDH (1:10000, ab8245; Abcam). Then the membranes were rinsed and mixed with secondary antibody by using HRP goat anti-rabbit/mouse IgG antibodies (Sigma-Aldrich). Enhanced chemiluminescence detection system (ECL, Millipore, Massachusetts) was used for photographing by X films. The integral optical density values of proteins were detected by Image Pro Plus software 6.0.
Flow cytometry analysis
Flow cytometric method was performed to measure cell apoptosis with an Apoptosis Detection Kit (Invitrogen, Waltham, MA) following the product manuals. Briefly, U87 or U251 cells that reached at 90% confluence were collected at 48-h transfection and washed with PBS. After that, cells were incubated with Annexin V-APC and propidium iodide in binding buffer. And then, the apoptosis rate was analyzed with a flow cytometer (FACSCalibur).
Cell proliferation assay
Cell Counting Kit-8 (CCK-8) assay was carried out for cell viability with a CCK-8 kit (Shanghai Genechem Co., Ltd.) following the product manuals. In brief, cells (1 × 103 cells/well) were seeded in 96-well plates and cultured with conditional culture medium for 24, 28, and 72 h. CCK-8 reagent (10 μL/well) was added into the medium before incubating at 37°C for 2 h. Then, the absorbance was determined at 450 nm by a microplate reader SpectraMax M5 (Molecular Devices).
Transwell assay
Cell invasion was determined in vitro by Transwell assay. Briefly, DMEM conditioned by U87 or U251 cells was added into the lower chamber, while the upper chamber contained DMEM with 1% serum and 0.1% BSA, and then incubated for 24 h at 37°C. Next, the filters (8 μm) were fixed with methanol (analytically pure) for 15 min. Then the invasive cells breached onto the lower side of the filters were conducted hematoxylin and eosin (H&E) staining. The cells were photographed and counted with a microscope (Olympus) (magnification 100 × ). Five fields were selected in each sample randomly, and each experiment was repeated thrice.
Wound healing assay
To evaluate the cell migration, U251 and U87 were seeded into six-well plate. When the confluence was 80%–90%, the wound was generated by a sterilized one-milliliter pipette tip. Then, at the time points of 0 and 24 h, the cells migrating into the area of wounds were observed and photographed under a microscope (Olympus) (magnification 100 × ). Five fields were selected in each sample randomly, and each experiment was repeated thrice.
Transcriptome sequencing and analysis
Total RNA in the samples was extracted with TRIzol reagent (Takara, Beijing, China). The quantity and integrity of extracted RNA were measured by Bioanalyzer 2100 system (Agilent). The mRNA library was constructed with rRNA depletion and then subjected to Illumina sequencing. After the clean reads were mapped to a reference genome with HISAT2 software, the results were transferred to StringTie, merged by Cuffmerge software, and quantified by StringTie software. Then, RPKM (Reads Per Kilobase of transcript per Million mapped reads) values were obtained. The p-values were adjusted through the Benjamini and Hochberg method. Genes with | log2 (fold change) | > 1 and adjusted p-value (padj) <0.05 acted as differentially expressed. The function among differentially expressed genes (DEG) was enrichment analyzed by gene ontology (GO) and Kyoto Encyclopedia of Genes and Genomes (KEGG) dataset.
Tumor-bearing mice establishment
Female BALB/C nude mice purchased from Charles River, weighing 18–22 g at 5 weeks, were randomly divided into different groups used for the function analysis of CHKA; each group has three mice. Tumor-bearing mice were established by injecting 1 × 106 U251 cells subcutaneously. The tumor volume was measured at 7, 14, and 21 d after simultaneous administration. The using of animals was conformed to the institution's guidelines of the General Hospital of Ningxia Medical University. The tumors collected were fixed with 10% formalin and embedded in paraffin before they were sliced into sections (5 μm), following H&E staining. The samples were observed and photographed under a microscope (Olympus).
Evaluation of apoptosis and proliferation using TUNEL assay and Ki-67 immunohistochemistry
The mouse tumor tissue apoptosis was examined by a TUNEL staining detection kit (C10245; Life Technologies). Briefly, mouse tumor tissues from different groups were obtained and fixed with 4% paraformaldehyde (PFA), embedded in paraffin, and sliced into 4 μm sections. For Ki-67 immunohistochemistry, the slices were incubated with Ki-67 antibodies (1:1000, ab15580; Abcam) at 4℃ overnight. Positive pixel intensity was determined as integrated optical density with ImagePro Plus v.6.2 software (Media Cybernetics). For TUNEL staining, the slices were mixed with 50 μL of TUNEL reaction mixture. The TUNEL intensity was examined under a fluorescence microscope (AxioCam MRc5, Carl Zeiss).
Immunofluorescence staining
Immunofluorescence staining was used to detect the AKT and PI3K level in tumor tissues. Briefly, the sections fixed with 4% PFA were blocked with 10% goat serum, and then mixed with p-AKT (1:1000, ab38449; Abcam) and p-PI3K (1:1000, ab278733; Abcam) antibody overnight at 4°C. Subsequently, these sections were incubated with secondary antibody TRITC-conjugated Goat Anti-Mouse IgG H&L (Alexa Fluor® 488) (1:1000, ab150113; Abcam) and then counterstained with DAPI (0.1 μg/mL; Sigma Aldrich). Images were taken under the OLYMPUS fluorescent microscope at 647 nm (IX71; OLYMPUS).
Statistical analysis
The data analysis (Student's t-test analysis or one-way analysis of variance followed by Tukey's post hoc test) was carried out by SPSS 20.0 (Chicago). *p < 0.05; **p < 0.01; ***p < 0.001; ##: no significance.
Results
CHKA was upregulated in glioma tissues
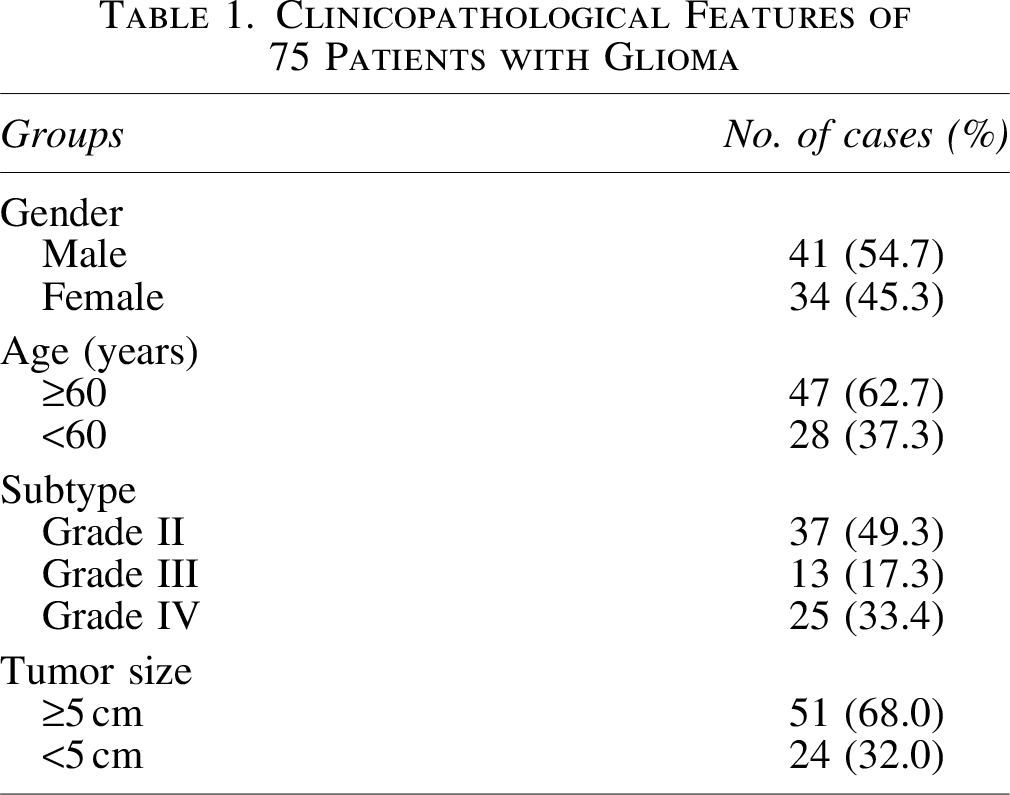
To know the function of CHKA in glioma, the expression of CHKA was detected in glioma tissues by qRT-PCR, Western blot, and immunohistochemistry staining. Immunohistochemistry staining indicated that the CHKA expression was significantly elevated in glioma compared with control group (Fig. 1A). qRT-PCR and Western blot analysis also confirmed these results (Fig. 1B, C). In addition, the authors found that CHKA level was higher in high-grade glioma tissues compared with low-grade glioma tissues (p < 0.05) (Fig. 1A–C). These results demonstrated that CHKA might involve in glioma progression.
CHKA was upregulated in glioma tissue.
CHKA promoted glioma cell behaviors in vitro
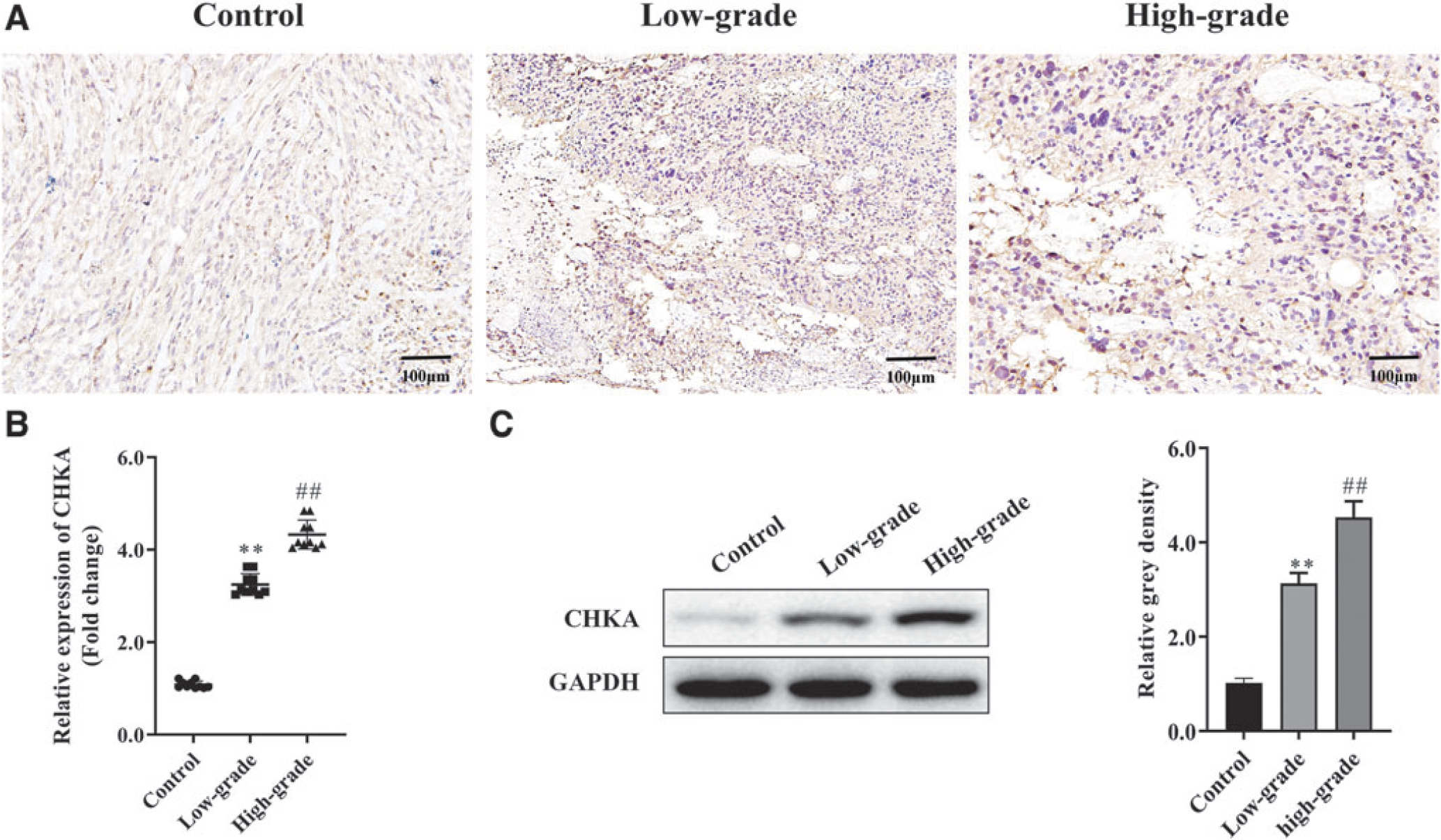
To know the function of CHKA on glioma, loss- and gain-of-function experiments were performed in U251 and U87 cells. As shown in Figure 2A, CHKA was successfully overexpressed or suppressed in U251 and U87 cells. CCK-8 analysis indicated that the cell proliferation of U251 and U87 cells was sharply elevated in over-CHKA group, while it declined in Sh-CHKA group compared with control group (Fig. 2B). Transwell and wound healing assays showed that overexpressing CHKA promoted cell invasion and migration, while suppressing CHKA inhibited the invasion and migration compared to control group (Fig. 2C, D). Flow cytometry analysis showed that the apoptosis was significantly inhibited in over-CHKA group, while it was promoted in Sh-CHKA group compared with control group (Fig. 2E). These data demonstrated that CHKA acted as an oncogene by promoting cell growth.
CHKA promoted glioma cell growth in vitro.
CHKA promoted glioma tumor growth in vivo
To confirm the results in vivo, tumor-bearing mice were injected subcutaneously with 1 × 106 U251 cells harboring with over-CHKA and Sh-CHKA vectors, respectively. Larger tumors were observed in overexpressing CHKA mice than those in the control group, while smaller tumors were found in mice when suppressing CHKA at 7, 14, and 21 d after injection (Fig. 3A). H&E staining of the mouse tissues indicated that overexpressing CHKA significantly increased inflammatory cell infiltration, while the inflammatory cell infiltration was suppressed in the CHKA-knockdown group compared with NC group (Fig. 3B). Ki-67 staining and TUNEL assay results showed that overexpressing CHKA promoted glioma cell proliferation and inhibited cell apoptosis, meanwhile, CHKA suppression inhibited glioma cell proliferation and promoted cell apoptosis (Fig. 3C, D). These results demonstrated that overexpression of CHKA promotes glioma development in vivo.

CHKA promoted glioma development in vivo.
PI3K/AKT was significantly enriched in CHKA-knockdown U251 cells
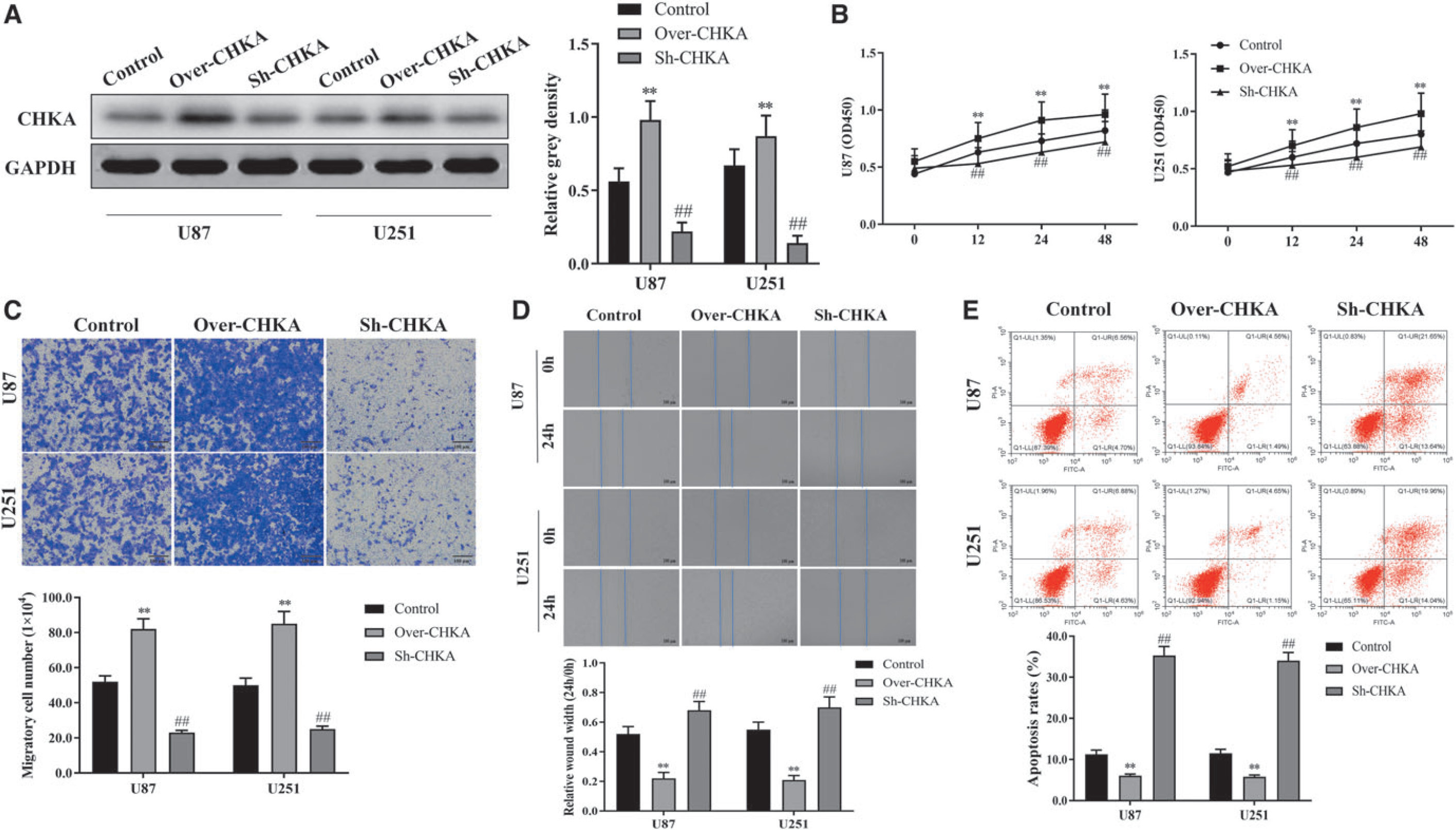
To know the signaling pathway affected by CHKA, the CHKA knockdown U251 cells were used to mRNA transcriptome sequencing. Enrichment analysis by GO suggested that most of the DEGs enriched in neutrophil activation involved in immune response, response to acid chemical and ribonucleoside monophosphate metabolic process (Fig. 4A). KEGG analysis indicated that most of these DEGs enriched in the PI3K-AKT, Salmonella infection, and human cytomegalovirus infection signaling pathways (Fig. 4B). And PI3K/AKT signaling showed the most significant enrichment among the signaling pathways (Fig. 4B). Therefore, the authors speculated that CHKA might involve in glioma development by affecting the PI3K/AKT signaling pathway.
PI3K/AKT was significantly enriched in CHKA-knockdown U251 cells.
CHKA promoted glioma development through PI3K/AKT signaling activation
To confirm whether CHKA promoted glioma development by activating PI3K/AKT signaling pathway, the expression of PI3K, AKT, p-AKT, and p-PI3K was detected under indicated transfection. The total protein abundance of AKT and PI3K was not changed. However, Western blot showed that the phosphorylated levels of p-PI3K and p-AKT were dramatically elevated in overexpressing CHKA U251 and U87 cells, while decreased in suppressing-CHKA U251 and U87 cells compared with the control group (Fig. 5A). Immunohistochemical staining revealed that the signaling of p-PI3K and p-AKT was promoted in the overexpressing CHKA tumor tissues, while decreased in suppressing-CHKA tumor tissues compared with control group (Fig. 5B). These results demonstrated that CHKA promoted glioma development by activating PI3K/AKT signaling pathway.
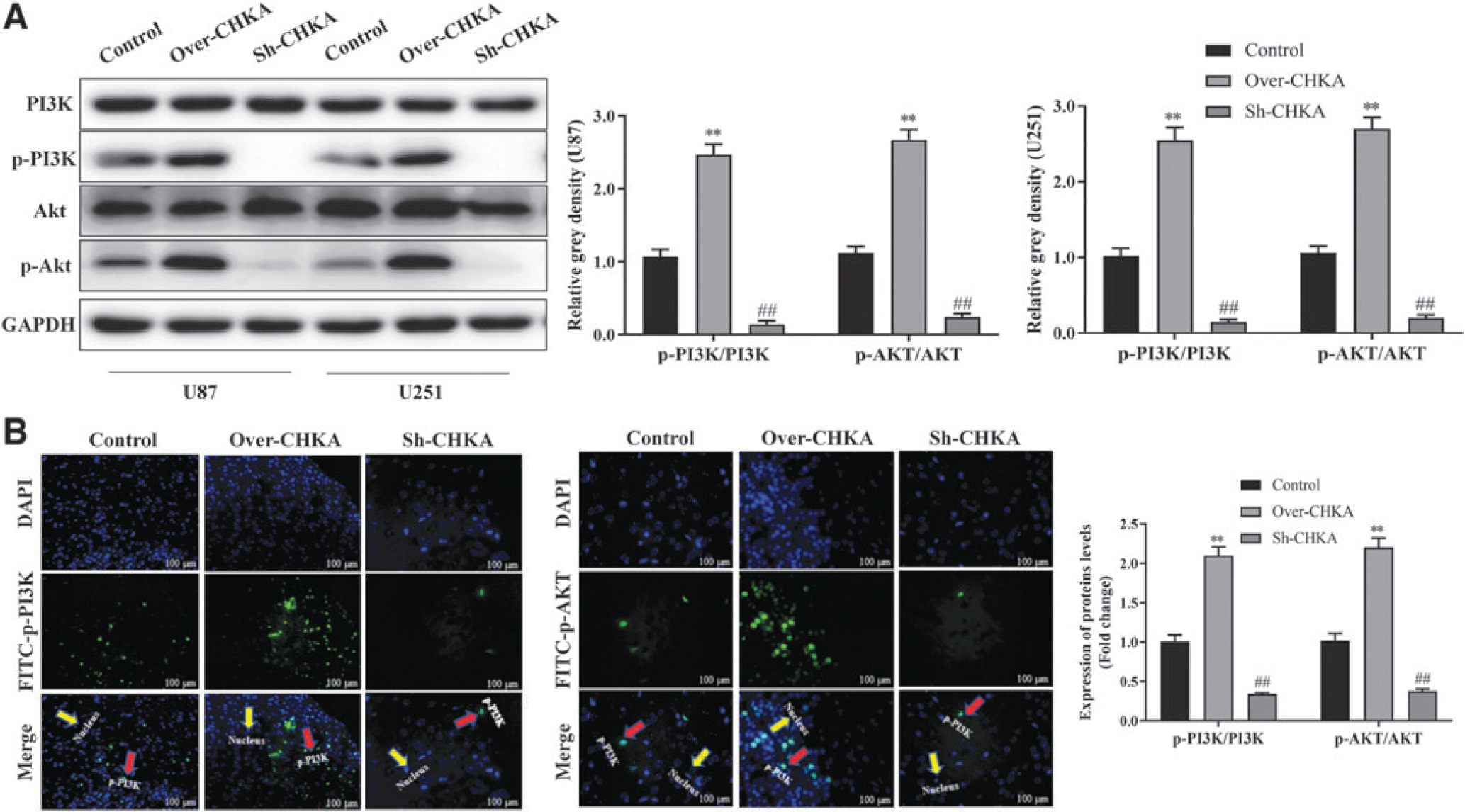
Overexpression of CHKA promotes glioma development by activating PI3K/AKT signaling pathway.
Discussion
Glioma is a malignant intracranial tumor. Although great advances have been achieved in surgery, radiotherapy, and chemotherapy, the patients' prognoses are still very poor owing to its strongly aggressive and progressive characteristics.2,14,15 Therefore, it is urgent to find new targets specifically targeting glioma. Herein, the authors confirmed that CHKA expression was significantly elevated in glioma tissues and is positively correlated with the grade of glioma. Mechanism analysis indicated that CHKA could promote glioma development by activating PI3K/AKT signaling pathway.
CHKA, the first reaction enzyme in the choline pathway, mainly catalyzes the conversion process of choline to phosphocholine. 9 In recent years, CHKA and its related metabolites have been reported to be significantly upregulated in a variety of malignant tumors and can be used as a molecular marker of poor prognosis for patients,16,17 suggesting that CHKA has the characteristics of oncogene. For example, Lin et al. showed that CHKA involved in the interactions between the epidermal growth factor receptor and the target of rapamycin complex 2 in hepatocellular cancer cells, which promoted xenograft tumor progression and/or drug resistance. 18 Gokhale et al. considered that elevated CHKA-mediated choline metabolism supported the prolonged survival of TRAF3-deficient B lymphocytes. 19 Recently, Rizzo et al. demonstrated that receptor-mediated cell death or drug induction could impair ChoKα/CHKA, which could be a potential therapy that was the integration into the combination strategy with immunomodulators or chemotherapeutics for improving ovarian cancer patients' outcomes. 12 Consistent with these results, the authors demonstrated that CHKA expression increased in glioma tissues. High CHKA expression was related to high glioma grade. Moreover, CHKA promoted the proliferation, invasion, and migration, while it decreased the apoptosis rate of the glioma cell lines in vitro and promoted glioma tumor growth in vivo. However, the mechanism of CHKA in regulating glioma development remains unclear.
To know the signaling pathway in which CHKA involved in glioma development, transcriptome sequencing was performed by CHKA-knockdown U251 cell line. The results of enrichment analysis by GO indicated that most of these DEGs enriched in neutrophil activation involved in immune response, response to acid chemical, and ribonucleoside monophosphate metabolic process. The results of KEGG analysis suggested that most of these DEGs enriched in PI3K-AKT, Salmonella infection, and human cytomegalovirus infection signaling pathways. Of them, PI3K/AKT signaling was the most significantly enriched signaling pathway, suggesting that CHKA contributed to glioma development by regulating PI3K/AKT signaling. In fact, the PI3K/Akt pathway is one of the most important pathways in cells, and PI3K/Akt regulated cell growth, proliferation, survival, metabolism, and angiogenesis, and participated in a variety of important biological processes.20,21 Current studies have confirmed that PI3K/Akt signaling is abnormally expressed in various tumors, and is closely related to tumor metastasis and prognosis,22,23 as well as in glioma. For example, celastrol suppressed glioma vasculogenic mimicry formation and angiogenesis by the inactivation of PI3K/Akt/mTOR pathway. 24 FoxD2-AS1 promoted glioma progression by regulating miR-185-5P/HMGA2 axis and PI3K/AKT signaling pathway. 25 In glioblastoma, TRIM31 enhanced chemoresistance by activating PI3K/Akt signatures. 26 Thus, the involvement of PI3K/Akt signaling in glioma affected by CHKA was further confirmed. The expression of PI3K and AKT was explored under CHKA elevation or depletion. The results demonstrated that the phosphorylated levels of p-AKT and p-PI3K were dramatically increased when overexpressing CHKA, while decreased when suppressing CHKA in cell lines. All these data demonstrated that CHKA promoted glioma development by activating PI3K/AKT signaling.
This study preliminarily verified the effect of CHKA on glioma cells and explored the underlying regulatory mechanism, which may provide a possible guiding direction for the treatment of glioma. Although there are currently targeted CHKA inhibitors and PI3K/Akt signaling pathway inhibitor drugs, they cannot be widely used clinically due to their instability, poor solubility, or high toxicity. Moreover, CHKA-related inhibitors, although inhibiting its activity, cannot effectively promote tumor cell apoptosis.18,27 Therefore, the combination of CHKA and PI3K/Akt signaling pathway inhibitors may provide a new combined application method for the glioma treatment.
Conclusions
In conclusion, the authors demonstrated that CHKA could promote glioma development by activating PI3K/AKT signaling pathway, suggesting that CHKA is an expected biomarker and therapeutic strategy for prognostic prediction of glioma patients.
Footnotes
Funding Information
This work was supported by the Natural Science Foundation of China (No. 81760449) and the Natural Science Foundation of Ningxia (No. NZ16127).
Disclosure Statement
No competing financial interests exist.