
Abstract
Abstract
This study was designed to examine the effects of the presence of oocyte nuclei on the donor cell nuclear remodeling, including premature chromosome condensation (PCC) and DNA configuration, and subsequent embryo development. The results showed that: (1) the presence of oocyte MII spindles was more likely to induce donor cell PCC. (2) The positional relationship between the fused donor cell and the oocyte metaphase spindle had an effect on oocyte PB2 extrusion. When the fused donor cell was widely separated from the MII spindle, 94.4% of the reconstructed oocytes expelled a PB2. When the donor cell was fused adjacently to the MII spindle, almost all of the reconstructed oocytes did not expel the PB2; the majority (67.9%) formed a very large M-phase spindle in which the oocyte and the donor cell chromosomes merged. (3) After activation, the oocyte and donor nuclei exhibited a variety of pronuclear patterns and asynchronous development. (4) The embryos reconstituted with nonenucleated oocytes resulted in a similar cleavage rate as observed in the control embryos reconstituted with enucleated oocytes. Blastocyst developmental rates were no different between nonenucleated and enucleated cloned embryos; however, the development rates from early to hatching blastocysts significantly decreased in the nonenucleation group compared to enucleation controls (0 vs. 23.1%; 27.5 vs. 67.8%), regardless with either cumulus cells or fibroblasts as donor cells. (5) All nonenucleated oocyte-derived blastocysts contained mixed polyploidy with a variety of compositions that included 2n/4n, 2n/6n, 2n/8n, and 2n/4n/8n. (6) Nuclear transfer preceding the oocyte enucleation experiment indicated that prolonged presence of oocyte nuclei induced abnormal DNA configuration and reduced in vitro development of transferred somatic nuclei, but short time presence of oocyte nuclei did not affect the in vitro development of cloned embryos. We conclude that oocyte MII spindles induce donor cell PCC, the developmental capacity of cloned embryos reconstituted with nonenucleated oocytes is inferior to those with enucleated oocytes, and that all such derived blastocysts are polyploidy.
Introduction
Metaphase II (MII) oocytes have been shown the better choice as recipients for nuclear transfer cloning compared to metaphase I or germinal vesicle oocytes (Campbell and Alberio, 2003; Li et al., 2004a). MII oocytes contain high levels of active maturation-promoting factor (MPF) and mitogen-activated protein kinase (MAPK) that induce nuclear envelope breakdown (NEBD), premature chromosome condensation (PCC), and dispersion of nucleoli in the transferred nuclei; all of which may be essential for nuclear reprogramming (Collas et al., 1992; Lee and Campbell, 2006). Studies in a range of species have shown that the occurrence and extent of PCC in the donor nucleus are variable between different species, donor cell types, activation protocols, and the age of the oocyte (Loi et al., 1998; Shin et al., 2002; Simerly et al., 2003). In the mouse, PCC is essential for nuclear reprogramming of transferred nuclei (Wakayama et al., 1998). In the rat, fusing cells into intact oocytes caused PCC with various genetic content, which was influenced by the strain of the rat used and the age of the oocytes (Hirabayashi et al., 2003). In the ovine, nonenucleated oocytes caused significantly more transferred nuclei to undergo NEBD than enucleated oocytes (Lee and Campbell, 2006). In the bovine, PCC is not necessary for the development of an embryo and directly results from injected cell nuclei to cytoplasts (Tani et al., 2003). Our previous report has shown that a prolonged interval between fusion and parthenogenetic activation results in disfigured spindle morphology; embryos with these abnormal spindle structures may develop poorly (Aston et al., 2006). However, whether and how the presence of the oocyte nucleus affects the transferred cell nuclear PCC is unknown.
A major concern of incomplete enucleation during the cloning procedure is the remains of residual genetic materials that might lead to aneuploidy and result in subsequent detrimental effects on development and genetic interference of the recipient cytoplasm (Dominko et al., 2000). Haploidization of a somatic cell by transferring the somatic cell to nonenucleated/enucleated oocytes, referred to as “semicloning” (Tesarik, 2002), has been used to produce artificial gametes. Lacham-Kaplan and Trounson (2001) injected diploid somatic cells into nonenucleated mouse oocytes to create artificial zygotes, but very few blastocysts were created. The potential production of artificial oocytes or zygotes by semicloning have been investigated in the mouse (Chen et al., 2004; Heindryckx et al., 2004; Tateno et al., 2003), rat (Fujii and Funahashi, 2008), and Chinese hamster (Tateno et al., 2003). Unlike secondary spermatocyte nuclei injection into oocytes in which the chromosomes of both the oocyte and spermatocyte completed their second meiotic division with resulting zygotes developing into normal fertile offspring (Kimura and Yanagimachi, 1995), all the above reports on semicloning showed a low percentage (<15%) of normal chromosomal number, serious chromosomal, or spindle abnormalities, and poor in vitro developmental capacity of artificially produced oocytes or zygotes.
This study reports our investigation on the dynamic behavior of oocyte nuclei and the transfer of cell nuclei in the same cytoplast, and how the presence of the oocyte nucleus affects cloned embryo development and genetic composition.
Materials and Methods
Unless otherwise noted, all reagents used were obtained from Sigma Chemical Co. (St. Louis, MO, USA).
Oocyte in vitro maturation (IVM)
Maturation of bovine oocytes was as described previously (Li et al., 2004b, 2004c). Bovine cumulus oocyte complexes (COCs) were aspirated from 3–8-mm diameter follicles on ovaries obtained from a local abattoir. Only COCs with a compact and homogenous ooplasm were selected. The COCs were matured in TCM 199 with Earle's salts, L-glutamine, and sodium bicarbonate (Gibco Inc., Grand Island, NY, USA) supplemented with 10% FBS (HyClone, Logan, UT, USA), 25 μg/mL gentamycin, 0.01 U/mL FSH (NIH-FSH-S17), and 0.01 U/mL LH (USDA-bLH-6) in four-well plates at 39°C in humidified 5% CO2 in air. The COCs were cultured for 16 h, and the cumulus cells were removed by vortexing in 0.5% hyaluronidase medium. The denuded oocytes were then continually cultured in maturation medium to IVM 20 h. After the two-step maturation process, the expelled first twopolar body (PB1) remained close to the oocyte MII spindle and did not shift location. Therefore, PB1 was used to identify the oocyte chromosome location and resulted in an enucleation rate of 90% (Li et al., 2004a).
Donor cell culture
Bovine cumulus cells were separated from COC matured for 24 h with 0.1% hyaluronidase (Type-1), washed several times in DMEM/F12 (1:1) (Gibco Inc.), and cultured in DMEM/F12 supplemented with 10% FBS at 37°C under 5% CO2 in air.
The fibroblast cells were derived from an ear biopsy of a 3-year-old dairy cow. Tissues were washed thoroughly in Hank's balanced salt solution supplemented with 2% FBS and antibiotics. Following the wash, tissue was minced, suspended in DMEM/Ham's F12 (1:1) supplemented with 15% FBS and antibiotics, seeded in 25 cm2 tissue culture flasks, and cultured at 37°C in a humidified atmosphere of 5% CO2 in air for 7–10 days. Both cumulus cells and fibroblast cells were passaged when they achieved more than 80% confluence.
The cells were starved in 0.5% FCS-containing medium for 3 to 5 days before nuclear transfer. Immediately before nuclear transfer, cells were dissociated by trypsinization with 0.25% trypsin with EDTA solution (HyClone). Small-size (10–12-μm) cells were used as donors.
Nuclear transfer
Routine manipulations in the enucleation groups (Li et al., 2004b, 2004c) were performed in calcium- and magnesium-free phosphate-buffered saline (PBS) supplemented with 3 mg/mL fatty acid-free bovine serum albumin (BSA) and 5.0 μg/mL cytochalasin B. PB1 and oocyte genetic materials were removed in a small volume of cytoplast. Successful enucleation was confirmed by Hoechst 33342 staining and illumination with ultraviolet of the karyoplasts. Single donor cells were individually transferred to the perivitelline space of recipient cytoplasts.
In the nonenucleation group, oocytes were held to make PB1 at 3 o'clock. PB1s were removed and the donor cells were transferred to the perivitelline space either at about 4 o'clock (perinucleus) or at about 8:30 (apo-nucleus). The procedure could be performed easily with a piezo-actuated micropipette.
Fusion and activation
Fusions were induced by one direct current pulse of 1.2 kV/cm for 25 μsec with an Electro Cell Manipulator 2001 (BTX, San Diego, CA, USA) in 0.27 M mannitol supplemented with 0.1 mM CaCl2, 0.1 mM MgCl2, and 0.05% BSA. Fused embryos were activated at 24 to 25 h after the onset of maturation by exposure to 5 μM ionomycin for 5 min followed by 5 h incubation in 10 μg/mL cycloheximide.
Embryo culture
After activation, embryos were cultured in 30 μL CR1aa containing 3% BSA for 40 h. Then the cleaved embryos were transferred on a monolayer of bovine cumulus cells in 30 μL drops of CR1aa containing 3% FBS at 39°C in a humidified atmosphere of 5% CO2 in air for another 5 days. Medium was changed every 48 h. Cleavage rate and blastocyst development were determined at 48 h and 7 days after culture.
Evaluation of nuclear events during in vitro culture
After fusion the cell nucleus became swollen and then NEBD occurred. Then afterward, the cell chromatin materials condense to visible, discrete chromosome-like structures called PCC. The secondary polar body (PB2) became round and smooth after activation of the fused clones with intact oocytes. After parthenogenetic activation, the donor cell nucleus or oocyte nucleus formed pronuclei (PN).
Chromosomal analysis and cell number count
Day 7 blastocysts were incubated in 0.05 μg/mL of colcemid (Gibco) in culture medium for 5 h, then treated in 0.8% trisodium citrate for 10–15 min and transferred individually onto a glass slide and fixed and stained with Giemsa. Chromosomes were examined at 1000× under oil, and then chromosome composition was determined for each blastocyst. Images were captured by a digital camera with PIXERA Viewfinder Program (Pixera Corporation, Los Gatos, CA, USA) under a Zeiss microscope. The categories assigned based on chromosome composition were as follows: diploid (2n = 60), tetraploid (4n = 120), mixoploid (e.g., 2n/3n, 2n/4n, 2n/3n/4n, 2n/4n/6n, and 2n/4n/8n). Any embryo that did not show an interpretable metaphase spread due to gross overspreading or clumped chromosomes was not classified. Cell number was simultaneously counted.
Microtubule immunofluorescent staining
Microtubule immunofluorescent staining was carried out as described previously (Li et al., 2005) with some modification. In brief, oocytes were fixed with 3.7% (w/v) paraformaldehyde in PBS overnight at 4°C. Fixed oocytes were extracted in PBS containing 1% (w/v) Triton X-100 and 0.3% BSA for 1 h at 37°C. After washing twice with PBS containing 0.01% Triton X-100, oocytes were blocked in PBS containing 150 mM glycine and 1% Triton X-100 for 1 h at 37°C. The oocytes were then incubated in a mouse monoclonal antibody against α-tubulin (Sigma, T-5168) diluted by PBS (1:100) for 1 h at 37°C. Then they were washed with PBS and incubated in fluorescein isothiocyanate-labeled goat-antimouse Ig G (Southern Biotechnology Associate, Inc. Birmingham, AL, USA, Cat No. 1030-02) diluted with PBS (1:100) for 1 h at 37°C. DNA was stained with 10 μg/mL of propidium iodide. Oocytes were mounted on slides in 50% glycerol in PBS. The samples were examined under a Zeiss epifluenent microscope (Carl Zeiss Optical, Inc., Chester, VA, USA). Images were captured by digital camera with PIXERA Viewfinder Program (Pixera Corporation, Los Gatos, CA, USA).
Experimental design
Experiment 1 examined the donor cell nuclear behavior after transfer to intact or enucleated oocytes. The donor cells were transferred to intact oocytes far from the MII spindle. The fused reconstructed oocytes were cultured in CR1aa for 0.5, 1, 2, 3, 4, 5, 6, and 20 h, and then fixed and immunostained.
Experiment 2 tested the donor cell and oocyte nuclear behavior after transfer of donor cells to intact oocytes far from or close to the oocyte MII spindle. The fused reconstructed oocytes were cultured in CR1aa for 0.5, 1, 2, 3, 4, 5, 6, and 20 h, and then fixed and immunostained.
Experiment 3 investigated donor cell nuclear behavior in the intact oocyte cytoplasm after parthenogenetic activation. Reconstructed oocytes were activated with ionomycin and cycloheximide. The activated oocytes were fixed and immunostained after culture for 0.5, 1, 2, 3, 4, 5, 6, and 20 h.
Experiment 4 examined the development of reconstructed embryos by transferring donor cells into intact oocytes. The fused reconstructed embryos were cultured in CR1aa for 7 days. Cleavage rate, blastocyst development and chromosomal composition of the blastocysts were determined.
Experiment 5 investigated the effects of the presence of the oocyte nucleus on the development of cloned embryos by reverse nuclear transfer (RNT). In this experiment, the polar body was positioned at 6 or 12 o'clock, and then a donor cell was inserted at 3 o'clock through a 25–30 μm incision drilled by a piezo-actuated pipette. The incision could be observed under a stereomicroscope, which served as a marker for locating the donor cell when performing electrofusion. After cell insertion, the cells were induced to fuse with the oocytes by electrical stimulation. Enucleation was carried out 1 (RNT-1h) or 3 h (RNT-3h) postelectrofusion. Only the oocytes with a successfully fused donor cell were enucleated by aspirating PB1 and surrounding cytoplasm with a piezo-actuated pipette. Successful enucleation was confirmed by UV illumination of the Hoechst-stained karyoplasts as described above. The enucleated oocytes were allowed to recover for 30 min in an incubator and then activated with ionomycin and CHX treatments. The resulting embryos were cultured for further development; some embryos were fixed at 2, 4, and 6 h postonset of activation for microtubule and DNA staining. The karyotypes of RNT cloned blastocysts were characterized.
Statistical analysis
Data were pooled from four or more replicates per group for the in vitro development studies. Chi-square analysis was used to determine differences in cleavage rate and development to the blastocyst stage. A probability of p < 0.05 was considered statistically significant.
Results
Donor cell PCC occurred more readily in intact oocyte cytoplasts
In the enucleated group, the majority of donor cells occurred NEBD and entered premetaphase I stage with small tubule asters after fusion for 1.5 h (Fig. 1A and B); 2 h after fusion, the donor cell PCC occurred and formed typical M-phase spindles (Fig. 1C); at 3 or more hours after fusion the spindles became elongated or separated into several pieces. In the nonenucleated group, the donor cell PCC and M phase-like spindles formed as early as 30 min after fusion (Fig. 1D); 1 h after fusion, the oocyte MII spindle and the donor M-phase spindle coexisted in the same cytoplast (Fig. 1E and F); then 2 or more hours after fusion the donor cell spindles became disfigured (Fig. 1G and H) and separated into several pieces (Fig. 1I). Donor cell PCC occurred more readily in the intact oocyte compared to that in the enucleated oocyte.
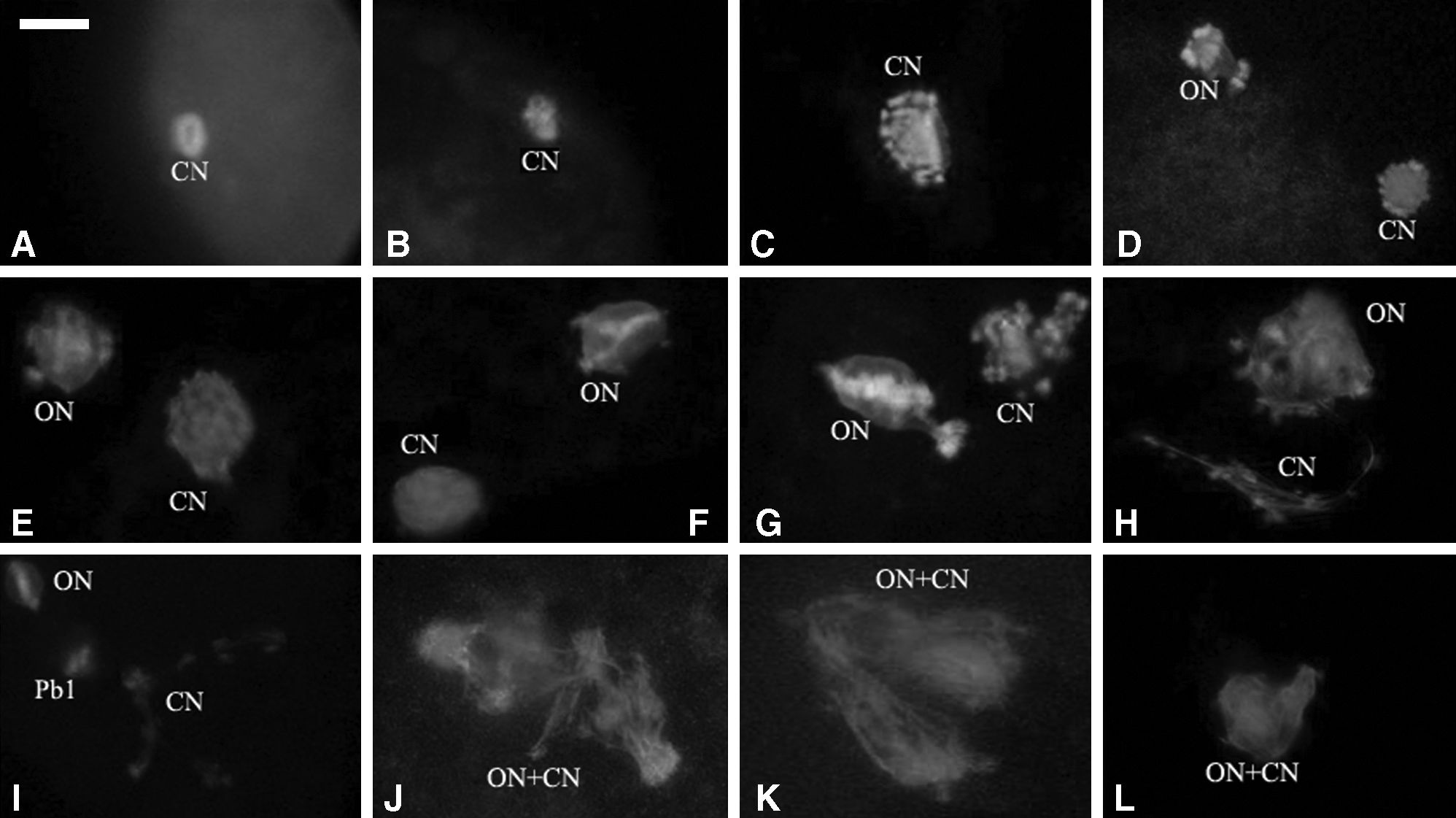
Nuclear state of the enucleated or unenucleated cloned embryos. (
Twenty hours after fusion, 74.4% (90/121) of oocyte nuclei in the nonenucleated group entered metaphase III (MIII). The donor cell nuclei showed a variety of structural patterns such as M-phase (50%), one to four long spindle pieces (18.2%), pronuclei (PN) (6.6%), and chromosomal spreads (8.3%) (Table 1). In 10.7% (13/121) of the reconstructed oocytes, M-phase and donor cell M-phase chromosomes connected and merged together (Fig. 1J–L). None of the oocyte nuclei formed PN and only 6.6% of the donor cells formed PN.
Fusion position of donor cell affected oocyte PB2 extrusion
When donor cells were fused far from the oocyte MII spindle (apo-nucleus), 94.4% (85/90) of the reconstructed oocytes expelled a PB2 (p < 0.01). When fused cells were adjacent to, or in close proximity to MII spindles (perinucleus), very few NT oocytes expelled a PB2. The oocyte MII spindle and donor cell M-phase spindle connected or merged together to form a large spindle. In the enucleated group, 5.2% (4/77) of the reconstructed oocytes expelled a PB2. In the parthenogenetic activation group, 93.8% (61/65) of MII oocytes expelled PB2 after activation (Fig. 2).
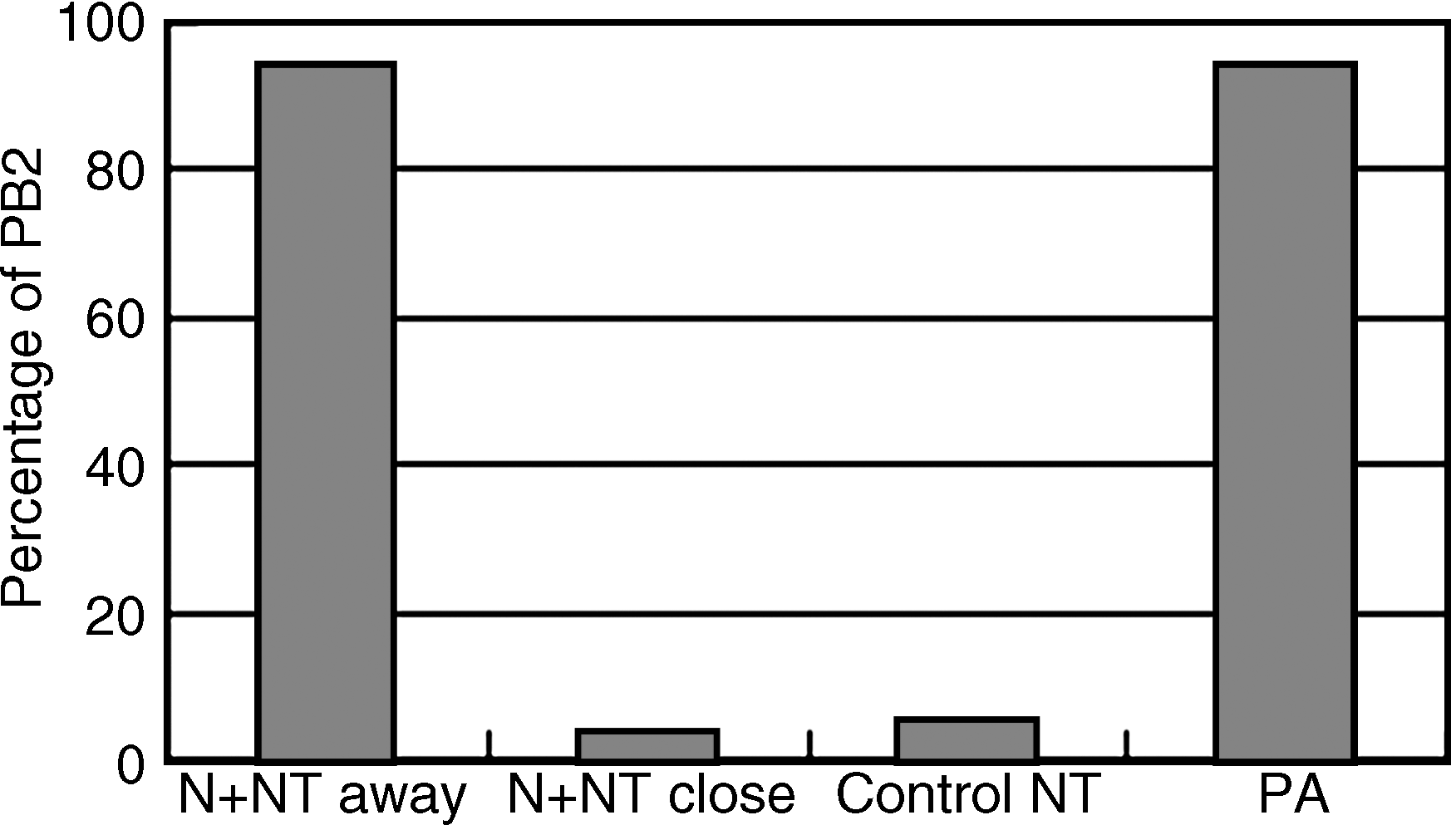
Fusion position of donor cell affected oocyte PB2 extrusion. NT + N away: unenucleated NT the donor cell far away from oocyte MII spindle; NT + N close: nonenucleated NT the donor cell fusing close to MII spindle; Control NT: enucleated nuclear transfer; PA: parthenogenetic activation.
Asynchronous pronuclear development of donor cell and oocyte nuclei after activation of fused clones
Fused clones were activated with ionomycin for 5 min and then CHX for 5 h. After CHX treatment, 81% of the transferred somatic cell nuclei formed a single PN. Simultaneously, 67% of the oocyte nuclei formed smaller PN when compared to those from donor cells (Fig. 3A), and 25% of the oocyte nuclei were at the telophase II stage. Twenty-hours after in vitro culture, 52.6% of the clones had PN from oocyte nuclei and the transferred donor nuclei (Fig. 3B). In 6.6% of the cloned embryos with nonenucleated oocytes, the oocyte PN and donor cell PN connected (Fig. 3C and D); and in 9.2% of the embryos, the oocyte nucleus and cell nucleus formed a large PN (Fig. 3E). Around 13% of the embryos had asynchronous nuclear development. The oocyte and cell nuclei were at either PN or in the metaphase stage. Irregular nuclear development such as several masses of chromatin (Fig. 3F and G), oocyte nuclear dividing (Fig. 3H), scattered chromosomes (Fig. 3I), and two sets of telophase spindles (Fig. 3J) occurred with 14.5% frequency (Table 2). The merged donor cell and oocyte nuclei entered the first mitosis, whereas the unmerged chromosomal masses formed PN-like structures (Fig. 3K).
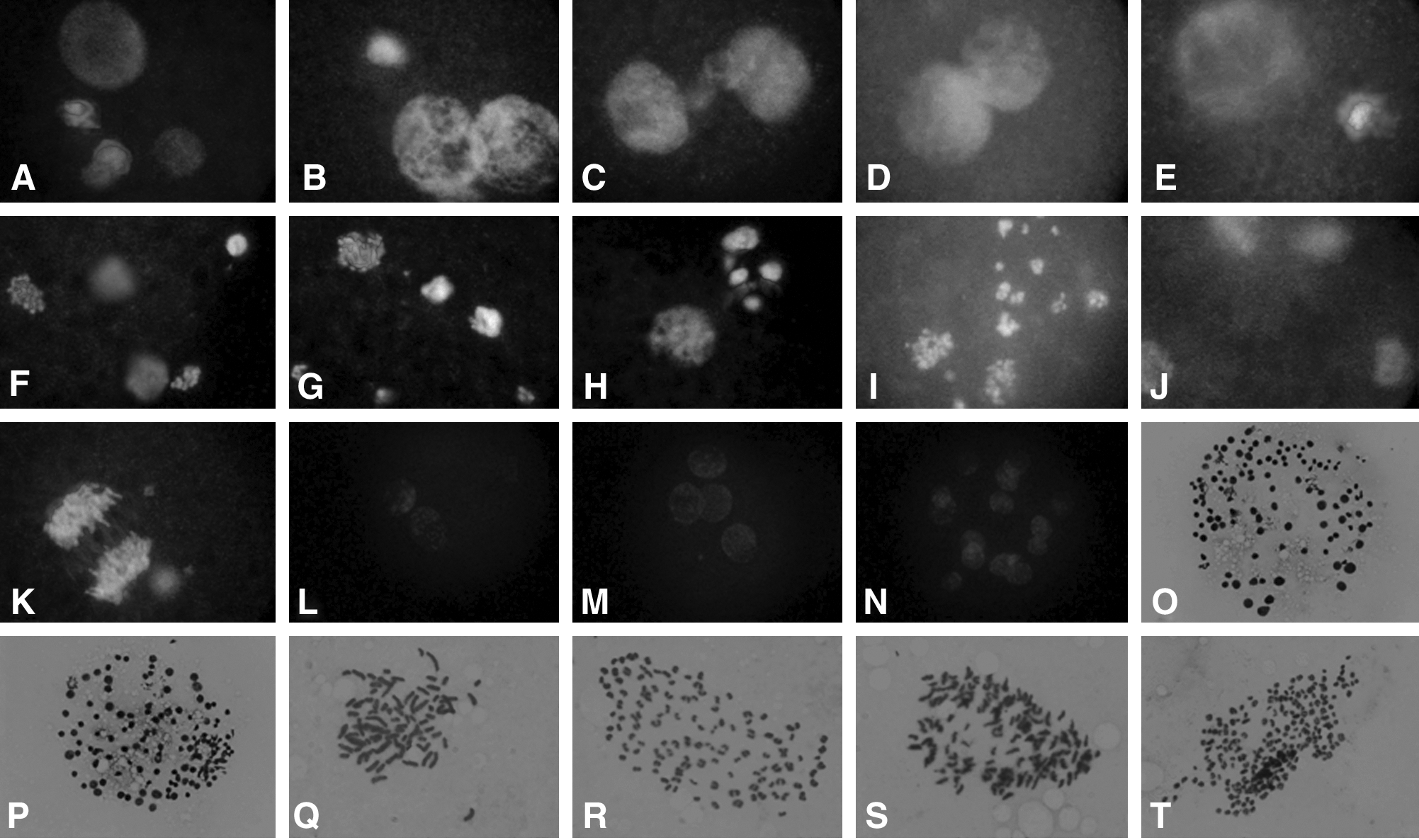
Development of unenucleated reconstructed oocytes after activation with ionomycin and cycloheximide and cultured in CR1aa. (
Development of the cloned embryos derived from nonenucleated oocytes
Of the nonenucleated cloned embryos at two-, three, four-, and six-cell stages, approximately 60% of blastomeres were mononuclear, 40% were dikaryon, and a few blastomeres had no DNA. In the enucleated groups, 94% of the blastomeres of embryos at two- to six-cell stage were mononuclear (Fig. 3L–N).
There were no differences in cleavage rates between nonenucleated and enucleated clones (82–90%) either with cumulus cells or fibroblasts as donor nuclei (Table 3). Both cumulus cells and fibroblasts resulted in about a 22% blastocyst development rate when transferred into nonenucleated oocytes. There was no statistical difference in the rate of development when compared to the enucleated controls (around 30%). Fibroblast cells would support development in nonenucleated NT blastocysts to the hatching and hatched stages of development (Table 3).
NT, normal enucleation nuclear transfer; N + NT, unenucleation nuclear transfer.
Values with different superscript in the column are significantly different.
Chromosomal composition of cloned blastocysts derived from nonenucleated oocytes
The average identifiable mitotic spreads in the control and nonenucleated NT blastocysts were around 15 and 10%, respectively. The results showed that 75% and 82% of the blastocysts were diploid when cumulus cells and fibroblasts were transferred to enucleated oocytes, respectively. The blastocyst cell number in the nonnucleated groups (about 60 cells) was significantly lower than the enucleated control groups (around 90 cells) when using cumulus or fibroblast donors cells (Fig. 4O–P). Chromosomal analysis of the resultant embryos showed that all the blastocysts derived from nonenucleated cloning exhibited a variety of mixed polyploidy such as 2n/3n, 2n/4n, 2n/6n, 2n/8n, 4n/6n, 4n/8n, 1n/2n/4n, 2n/3n/4n/, and 2n/4n/8n (Fig. 3Q–T). The mixtures of 2n/4n, 2n/8n, 4n/8n, 2n/4n/6n, and 2n/4n/8n occurred at a higher frequency (Table 4).
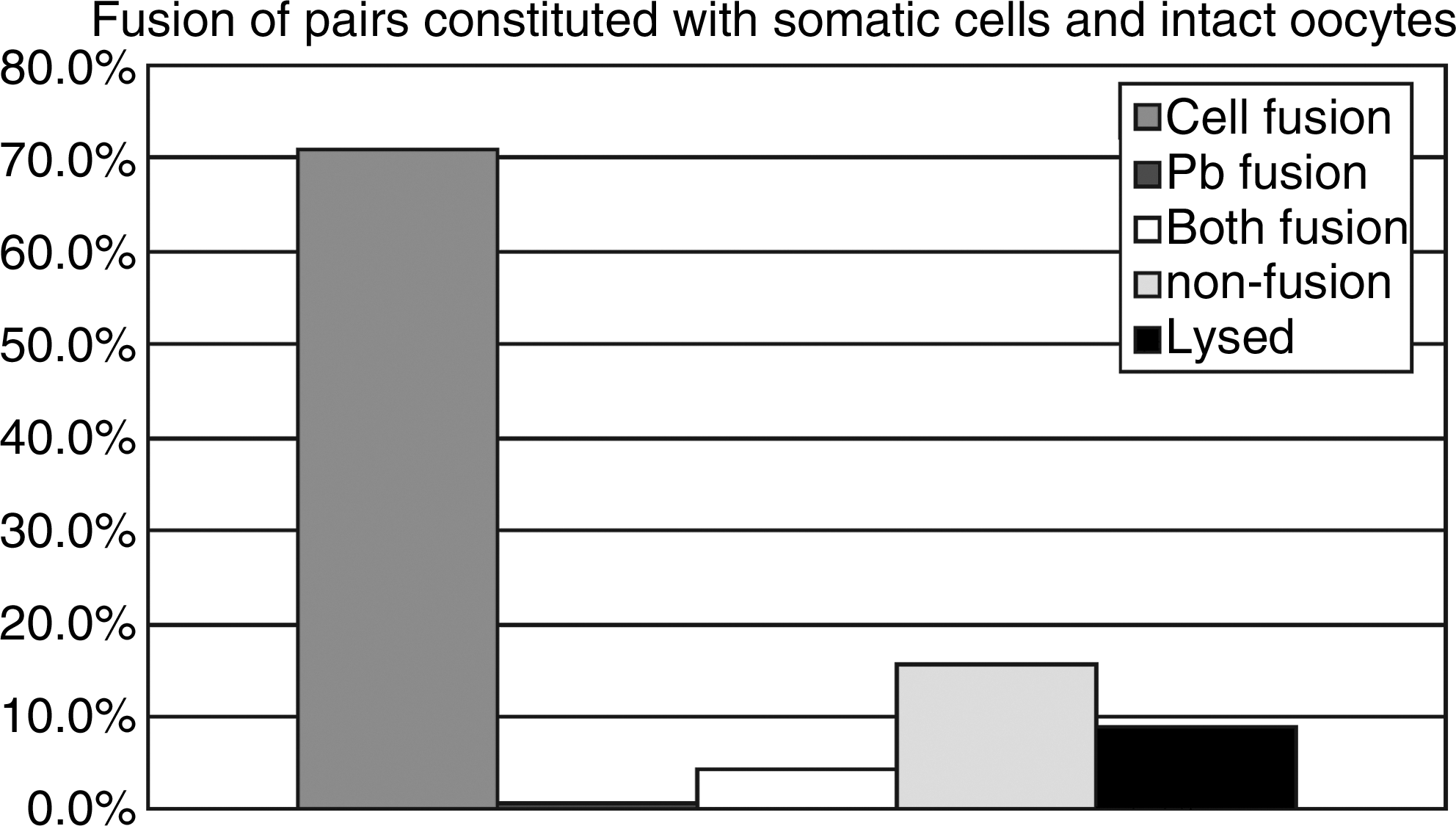
Fusion results of oocyte–cell pairs reconstituted by inserting somatic cells into perivitelline space of oocytes with polar bodies. The fusion rate of oocyte with cell only (cell fusion) was 70.9%; the fusion rate of oocyte with polar body only (Pb fusion), or both cell and polarbody (both fusion) was 0.6 and 4.2%, respectively; the rate of nonfusion, lysed oocytes was 15.7 and 8.7%, respectively (n = 357).
BCC, bovine cumulus cells; BFC, bovine fibroblast cells; NT, normal nuclear transfer; NT + N, nuclear transfer with presence of oocyte nuclei.
p < 0.05.
DNA configuration of somatic cell nuclei transfer by reverse nuclear transfer (RNT)
In the RNT experiment, a majority (70.9%) of the oocytes fused with transferred cells; only about 5% of oocytes fused with polar bodies, or with both polar bodies and cells (Fig. 4). In the short time interaction of the oocyte–somatic nuclei group in which oocyte enucleation was performed 1 h postcell fusion (RNT-1h), the nuclear DNA were not separated away in most oocytes (65%), and became swollen, deployed, and reorganized to form pronuclear-like structures within 6 h postonset of activation. In the longer time interaction of oocyte–somatic nuclei group, in which oocyte nuclei were removed 3 h postcell fusions (RNT-3h), nuclear DNA pieces remained separated from each other. They appeared more unorganized when compared with those in the RNT-1h group. Most of the somatic nuclei in this group failed to form pronuclear-like structures within 6 h postonset of activation (Fig. 5).
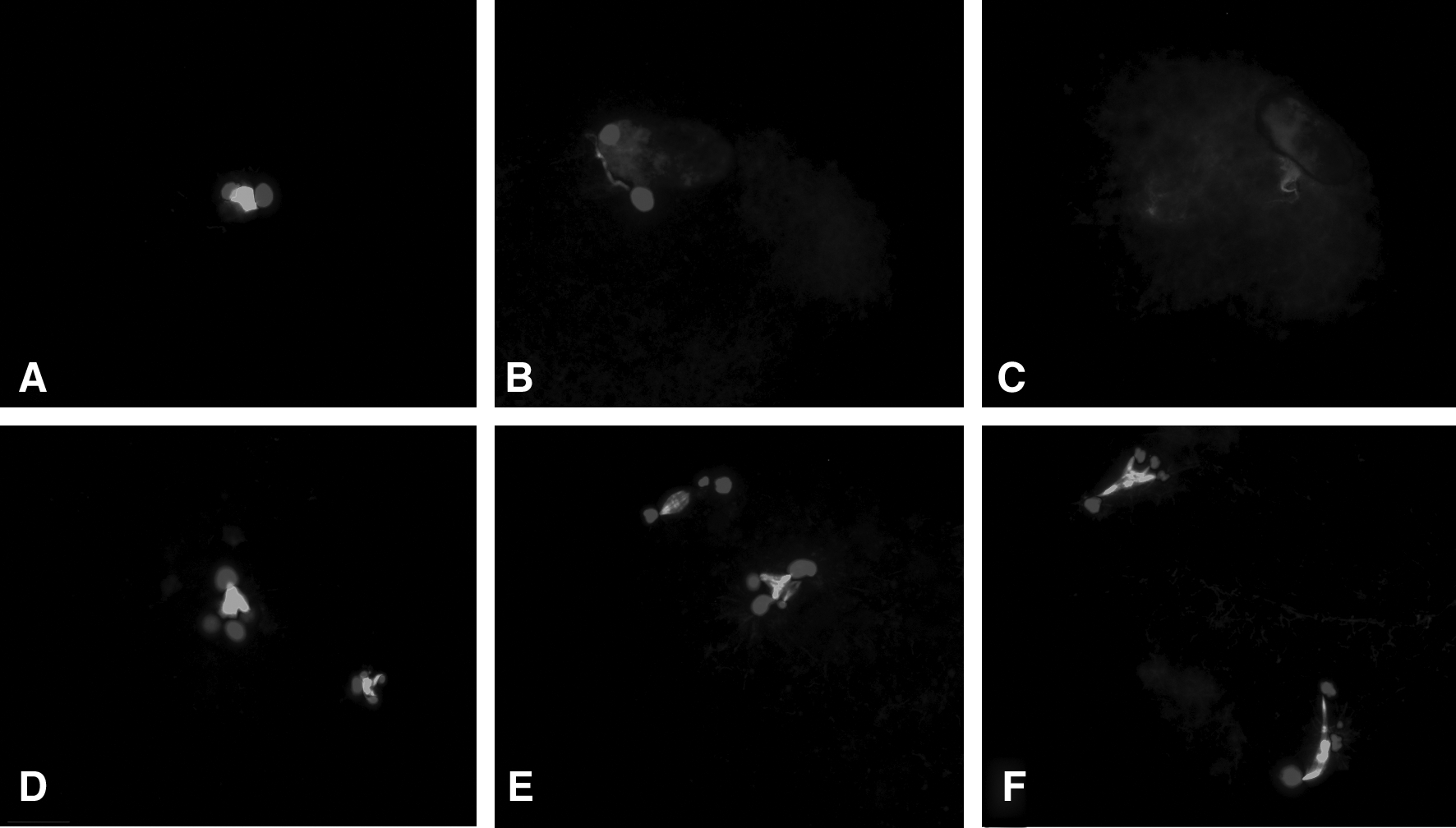
Somatic cell nucleus status of cloned embryos produced by RNT-1h (
Development of RNT cloned embryos
The in vitro developmental data of embryos produced by two reverse nuclear transfer protocols, RNT-1h versus RNT-3h, were summarized in Table 5. There was no difference in the cleavage rate between these two groups. The blastocyst development of the RNT-1h group, however, was much higher than the RNT-3h group (47.2 vs. 20%, p < 0.01). A similar proportion of the resulting blastocysts in the two groups hatched or expanded (92.9 vs. 83.8%, p > .05). There was no difference in the proportions of diploid cells in these two groups (81.2 vs. 80.6%, p > 0.05).
RNT-1h, enucleation was carried out 1 h postfusion of donor cell and oocyte.
RNT-3h, enucleation was carried out 3 h postfusion of donor cell and oocyte.
Values with different superscript in the column are significantly different.
Discussion
High MPF activity in vertebrate oocytes affects chromosome condensation and the maintenance of the meiotic spindle. Both mechanisms are involved in meiotic arrest at MII stage (Li et al., 2005). MPF resides near the chromatin structure at the M-phase of the cell cycle and spreads out over the cytoplasm at the G2 phase (Jones, 2004). When MPF activity was inactivated by the degradation of cyclin B (Ookata et al., 1992), oocytes were released from the MII stage. High levels of MPF and MAPK causes PCC in the transferred nucleus and have been implicated in nuclear reprogramming (Nurse, 1990). Donor nuclear PCC was shown to be essential for the success in mouse cloning. In the rat, the PCC-inducing potential was determined by the origin of the recipient cytoplasm, not by the origin of the donor nucleus (Hirabayashi et al., 2003). This is not the case in the bovine as PCC has not been shown to be essential for nuclear reprogramming (Abrieu et al., 2001; Aston et al., 2006). Our previous report (Aston et al., 2006) and the data in this study showed that a prolonged interval between fusion and activation of more than 3 h would result in the occurrence of two or multiple chromosomal complexes, which subsequently caused a diversity of single, double, and multiple nuclei structures in NT embryos. Tani et al. (2003) suggested that reprogramming of bovine somatic cell nuclei is not directly regulated by MPF or MAPK activity.
This study showed that the presence of oocyte MII chromosomes could easily induce donor cell nuclei to initiate PCC. PCC occurrence lagged behind at least 1 h in enucleated NT embryos when compared to nonenucleated clones. Because MPF is preferentially bound to chromosomes (Fulka et al., 1996; Sung et al., 2007), the presence of oocyte chromosomes would ensure the maintenance of high levels of MPF, which most likely contributed to the formation of donor cell PCC. Enucleation probably removes the majority of MPF and the functionality of high MPF levels in cytoplasts would therefore be lower than that in intact oocytes (Lee and Campbell, 2006). The reduced levels of MPF in enucleated cytoplasts could still be affective in inducing PCC (Fulka et al., 1996), but probably with a slower mode of action on the donor nuclei. A similar phenomenon was found in the ovine, when somatic cells were fused to nonenucleated oocytes. Most of the nuclei became PCC; while the majority of the donor nuclei remained intact when they were fused to enucleated oocytes (Lee and Campbell, 2006).
Usually a single mature oocyte contains only one set of haploid chromosomes. Two or more extra sets of chromosomes might result in polyploidy after fertilization or nuclear transfer and usually leads to early embryonic death. In standard nuclear transfer procedures, to maintain the diploid composition of NT embryos, diploid somatic cells are introduced into enucleated oocytes. The effects of transferring somatic cells to nonenucleated oocytes have been studied in the mouse. Injection of somatic cells into intact oocytes resulted in the extrusion of two PB2: one derived from the oocyte chromosomes and the other from the somatic cell chromosomes (Fulka et al., 1998). The authors suggested that a peripherally placed spindle could promote an extra polar body extrusion, although a centrally positioned donor spindle could lead to the formation of three blastomeres (Fulka et al., 1998). In our study using bovine nonenuleated oocytes, when the cell was fused far away from the oocyte MII spindle almost 100% of the oocytes expelled a PB2. When a cell was fused adjacent to an oocyte MII spindle, the two nuclei merged together and their chromosomes formed a large spindle, and the oocyte did not expel a PB2. Our data suggests that the locality of cell fusion at the surface of nonenucleated oocytes will affect embryo chromosomal configuration and developmental patterns as follows.
We examined whether cell nuclei injected into intact oocytes would support reconstructed embryo development. In mice, embryos reconstructed by injecting somatic cells into intact oocytes resulted in high (80–90%) cleavage and 50% morula rates; however, the developmental competence to blastocysts was very limited (Lacham-Kaplan and Trounson, 2001). In the bovine, we observed that the reconstituted embryos with nonenucleated MII oocytes resulted in similar cleavage rates (82–90%) compared to those with enucleated oocytes. The blastocyst development in nonenucleated clones, however, was lower than that in enucleated control.
All resultant blastocysts from nonenucleated oocytes had polyploidy chromosomal compositions. The analysis of two- to six-cell stage embryos reviled that approximately 40% of the embryos had blastomeres containing two nuclei and a few blastomeres contained no nucleus. Previous studies involving chromosomal analysis of cloned bovine embryos have shown that approximately 80% of blastocysts were diploidy (Mohamed and Takahashi, 2000; Li et al., 2004b, 2004c; Plusa et al., 2005). The presence of an oocyte nuclei and a donor cell produced a variety of diploidy in resultant NT blastocysts. The fusion of a diploid cell nucleus with a diploid (fused adjacent to MII spindle) or with a haploid (fused far from MII spindles) oocyte often resulted in tetraploidy or triploidy in reconstructed embryos. Such 3n or 4n embryos consisted of various combinations of ploidy. Embryo development was significantly retarded when the genetic complement of blastomeres was compromised. Embryos with polyploidy cells resulted in a significantly lower blastocyst cell number. Morphologically poor appearing blastocysts tended to have lower cell numbers and exhibited a higher frequency of chromosomal aberrations (Li et al., 2004c). Previous reports on nuclear transfer to nonenucleated oocytes in the mouse also indicated that the percentage of semicloned zygotes with diploid sets were very low (6 and 12%) (Chen et al., 2004; Heindryckx et al., 2004). Tatecno et al. (2003) demonstrated that homologous chromosomes separation in semicloned embryos happened randomly and the possibility of achieving correct chromosomal assortment was associated with the number of chromosomes. Thus, normal chromosomal number in semicloned zygotes may not assure the further normal cleavage or the normal chromosomal number in blastocysts, and the possibility of forming normal chromosomal number in the semiclones in bovine (2n = 60) is much lower than in the mouse (2n = 40). This could account for the lower diploidy in our data set compared to that reported in the mouse.
To investigate the presence of oocyte nuclei on the development of cloned embryos, we performed reverse nuclear transfer experiments by removing oocyte nuclei 1 (RNT-1h) or 3 h (RNT-3h) after the fusion of somatic cells and oocytes. In the RNT-1h group, reprogramming procedure, such as PCC, swollen nucleus, and DNA deploying, seemed to take place more readily and quicker than in regular nuclear transfer cloning. The formation of pronucleus-like structures could occur as early as 4 h postactivation. However, in the RNT-3h group somatic nuclear DNA in most oocytes was distributed into multiple pieces and failed to reorganize normally. This morphologic configuration remained unchanged at 6 h postonset of activation. Our in vitro developmental data also agreed with observations on nuclear DNA configurations of RNT cloned embryos. The blastocyst developmental rate in the RNT-1h group was significantly higher than the RNT-3h group, and comparable with regular nuclear transfer cloning. The results indicate that the short time presence of an oocyte nucleus could enhance nuclear reprogramming of the transferred somatic nucleus, whereas a prolonged presence could induce abnormal DNA configuration of a transferred nucleus and reduce further developmental. A previous report in mouse RNT also indicated that oocyte enucleation 30 min to 1 h after cell nuclear transfer (by injection) achieved compatible live birth of clones compared with regular nuclear transfer cloning, whereas oocyte enucleation 3 h after cell injection resulted in zero pregnancies after embryo transfer (Wakayama et al., 2003).
This study provides evidence that the presence of an oocyte nucleus resulting from unsuccessful enucleation will lead to the formation of defective reconstructed oocytes and subsequent genetically compromised embryos. This is why proper enucleation is so important for establishing a normal diploid chromosomal composition in NT reconstructed embryos.
In summary, our results show that (1) oocyte MII spindles can efficiently induce donor cell PCC; (2) the oocyte nucleus is involved in the chromosomal composition of the NT embryos reconstituted with intact oocytes; (3) intact bovine oocytes can support cloned embryo development to the blastocyst stage; and (4) all blastocysts derived from nonenucleated oocytes had mixed polyploidy.
Footnotes
Acknowledgments
This work was support by Hi-Tech Research and Development Program of China (863 Program, No. 2007AA100505, No. 2008AA10Z159) and grant from Inner Mongolia University for Chang Jiang Scholars Program to GPL.
Author Disclosure Statement
The authors declare that no conflicting financial interests exist.
The first two authors contributed equally to this study.