
Abstract
Abstract
Genetic manipulation of porcine genome to produce genetically modified pigs with high efficiency has been hampered by the unavailability of an ideal cell type. The cell type currently used for various genetic manipulations is fetal fibroblasts. These cells have very limited life span in culture, and efficiency of gene targeting is very low. In this study, we developed a simple but novel strategy to derive cell lines from adult porcine liver and adipose tissues with long life span. Small colonies with few cells became visible as early as 2 to 3 days on collagen-coated plates, and a full-grown colony took 10 to 14 days to form. These cells maintained a steady growth up to 80 population doublings with normal karyotype. Transfection of these cells with a plasmid containing a neomycin resistance gene and selected under G418 yielded clones with stable genetic modifications and extended expression of the transgene. Further, these cells were used as nuclear donors to produce somatic cell nuclear transfer (SCNT) embryos. The average fusion rates were 86.8, 80.5, and 90.4% for liver-derived cell lines (LDCs), fat-derived cell lines (FDCs), and fetal fibroblasts (FFs), respectively. We achieved a pregnancy rate of 50% with both LDCs and FDCs at day 30 and the efficiencies of generating fetuses from cloned embryos were 3.5, 2.1, and 4.0% for LDCs, FDCs and FFs, respectively.
Introduction
Materials and Methods
Isolation and culture of cells from porcine liver and fat
Porcine liver-derived cells (LDCs) were isolated as per the protocol described for derivation of a stem cell type from human liver (Herrera et al., 2006) with some critical modifications. Briefly, liver tissue was harvested from mature female crossbred domestic pigs (n = 3) between the ages of 8 and 10 months at a local surgical unit. Samples were transported on ice in Dulbecco's phosphate-buffered saline (DPBS; Invitrogen, Carlsbad, CA, USA). The tissue (1–2 g) was perfused with warm Liver Perfusion Medium (Invitrogen) and digested in Liver Digest Medium (Invitrogen) at 37°C for 10–12 min. The hepatocytes were released by mincing and pipeting. The cell suspension was filtered through a sterile 40 micrometer Nylon cell strainer and centrifuged at 400 × g for 5 min. The cell pellet was washed two times with cold Hepatocyte Wash Medium (Invitrogen), resuspended in Williams Medium E (Invitrogen) supplemented with glutamine and 5% FBS (Hyclone, Logan, UT, USA), and plated on 10-cm2 collagen I-coated plates (BD Biosciences). Unattached cells, that represented all hepatocytes, were poured off after 2 to 3 h and replaced with a combination media (α-MEM:EGM-MV 3:1) (Invitrogen/Lonza, Switzerland) supplemented with 10% FBS (Hyclone), 10% horse serum (Invitrogen, Cat # 16050-122), 12 mM HEPES, and 1% Pen/Strep (Invitrogen). The plates were maintained at 37°C with 5% CO2 and 10% O2. Individual attached cells and cell clusters ranging from two to six cells per cluster were identified on the culture dish the next day. When colonies were evident, cloning rings were used to transfer each colony to an individual well of a 24-well collagen I-oated culture plate and further expanded to a 10-cm2 dish.
Porcine FDCs was isolated as per the protocol described (Traktuev et al., 2009). Briefly, porcine subcutaneous abdominal adipose tissue samples were harvested from mature female crossbred domestic pigs (n = 3) between the ages of 8 and 10 months at a local surgical unit. Samples were transported on ice in DPBS (Invitrogen). The tissue was cut to fine pieces and digested in the mixture of Collagenase Type I (1 mg/mL) (Worthington Biochemical, Lakewood, NJ, USA) and dispase (50 caseinolytic units/mL) (BD Biosciences, Bedford, MA, USA) under agitation for 30–45 min at 37°C. Stromal cell fraction was separated from adipocytes by centrifugation at 300 × g for 10 min. The pellet was resuspended in DMEM/F12 medium supplemented with 10% FBS and filtered through 40 micrometer cell strainer (BD Falcon, Franklin Lakes, NJ, USA). Cells were pelleted by centrifugation at 300 × g for 10 min and treated with red blood cell lysis buffer (Qiagen, Chatsworth, CA, USA) for 5 min at 37°C. Cells were washed in the DMEM/F12 medium and cultured in a combination media (α-MEM:EGM-MV:: 3:1) (Invitrogen/Lonza) supplemented with 10%FBS (Hyclone), 10% horse serum (Invitrogen, Cat # 16050-122), 12 mM HEPES, and 1% Pen/Strep (Invitrogen) on collagen-coated plates. The plates were maintained at 37°C with 5% CO2 and 10% O2. After 10 to 14 days, single colonies were isolated by cloning rings, subcultured in 24-well collagen I-coated plates, and further expanded to a 10-cm2 dish.
Isolation and culture of fetal fibroblasts
Pig fetal fibroblasts were extracted from 30-day-old fetuses. Briefly, head, legs, and other internal organs were removed and discarded. The remaining fetus was minced and plated onto 100-mm plates in MEM-α media (GIBCO, Gaithersburg, MD, USA) supplemented with 10% FBS (Hyclone) plus 1% Antibiotic-Antimycotic (Invitrogen) and grown at 37°C with 5% CO2. After 2 to 3 days, cells were harvested by trypsinization, and single-cell suspension, without cell clumps, was plated on fresh plates. At 80–90% confluency, cells were harvested and further expanded for immediate use or stored in liquid nitrogen for future use.
Colony-forming unit (CFU) assay
Porcine hepatocytes and adipose stromal cell fraction were seeded at different concentrations, based on viable cell count as determined by the trypan blue (Invitrogen) under the culture conditions described above. The media was changed every 2 to 3 days and the colonies were counted on day 10. These numbers were used to calculate the number of colonies obtained per million seeded cells. CFU assay was done in triplicates and number of colonies obtained was plotted as average ± SD.
Growth curve
LDCs and FDCs colonies were cultured on collagen I-coated plates in the media described above. Cells were harvested at 80–90% confluency by trypsin digestion, counted by automatic cell counter (Invitrogen), and plated on fresh plates. This process was repeated 30 times for LDCs and 27 times for FDCs. The population-doubling level (PDL) was calculated as Log2 (D/D0), where D and D0 are defined as the number of cells at the harvesting and seeding, respectively. These numbers were used to plot the growth curve.
Live cell imaging
The images of LDCs and FDCs were taken by an Axio Observer Inverted Microscope (Carl Zeiss, Thornwood, NY, USA).
Karyotyping
Karyotyping of LDCs at the 30th passage (P30) and FDCs at the 27th passage (P27) was performed by Cell Line Genetics (Madison, WI, USA). Briefly, chromosome preparations were made following harvesting, hypotonic treatment, and fixation of cells. The chromosomes were G-banded and examined using an Olympus BX 41 microscope equipped with Genus image analysis software (Applied Imaging, San Jose, CA, USA). Twenty cells from each cell line were evaluated for numerical and structural chromosome abnormalities (Meisner and Johnson, 2008).
Transfection
FFs, LDCs, and FDCs were cultured to 60–70% confluency, washed with calcium and magnesium-free PBS (Invitrogen), and harvested with Trypsin. The cells were washed with PBS and transfected with Microporator (Invitrogen) as per the manufacturer instructions. Briefly, one million cells were suspended in 100 μL of transfection buffer and 3 μg of linearized DNA construct was added. Cells were electroporated at 1300 V, 1 pulse for a 30-msec duration. Electroporated cells were plated onto 10-cm2 plates. After 72 h normal media was replaced with selection media containing G418 (Invitrogen) at 100 μg/mL. G418-resistant colonies appeared after 10 to 14 days of selection.
Nuclear transfer
SCNT was performed using in vitro matured oocytes (Minitub of America, Mount Horeb, WI, USA, and/or APR80, Madison, WI, USA). Cumulus cells were removed from the oocytes by pippeting in 0.1% hyaluronidase. Only oocytes with normal morphology and a visible polar body were selected. Oocytes were incubated in manipulation media (Ca-free NCSU-23 with 5% FBS) containing 5 μg/mL bisbenzimide and 7.5 μg/mL cytochalasin B for 15 mins. Oocytes were enucleated by removing the first polar body plus metaphase II plate, and one cell was injected into each enucleated oocyte. Couples were fused by two DC pulses of 140 V for 50 μsec (BTX cell electroporator, Harvard Apparatus, Hollison, MA, USA) in 280 mM Mannitol, 0.001 mM CaCl2, and 0.05 mM MgCl2. One hour later, reconstructed oocytes were activated by two DC pulses of 120 V for 60 μsec in 280 mM Mannitol, 0.1 mM CaCl2, and 0.05 mM MgCl2. After activation, oocytes were placed back in NCSU-23 medium with 0.4% bovine serum albumin (BSA) and cultured at 38.5°C, 5% CO2 in a humidified atmosphere for less than 1 h, before being transferred into the recipient. Recipients were naturally cycling occidental pigs on their first day of estrus. In all cases recipients were first parity gilts.
Single nucleotide polymorphism (SNP) genotyping
DNA samples from nuclear donor cells, cloned fetuses, and surrogate recipients, were subjected to SNP genotyping using MassARRAY technology (Sequenom, Inc., San Diego, CA, USA) carried out at GeneSeek, Inc (Lincoln, NE, ISA). Three optimized multiplex SNP panels consisting of 96 highly informative swine SNPs were used (Fahrenkrug et al., 2002; Rohrer et al., 2002). Data were analyzed using MassARRAY Typer 3.4 software (Sequenom, Inc).
Results
Isolation and characterization of LDCs and FDCs from adult swine
To efficiently and rapidly isolate cell lines from adult tissues, hepatocytes and stromal cells from adipose tissues were directly plated on collagen I-coated plates in a combination media (α-MEM/EGM-MV supplement with 10% FBS and 10% HS) at different concentrations and maintained under reduced oxygen conditions. This media is neither optimal for hepatocytes nor stromal cells. Under these culture conditions, hepatocytes and stromal cells did not attach, and only a rare population of cells present among these cells attached and formed colonies. Colonies with few cells became visible as early as 2 to 3 days, and a fully formed colony took 10 to 14 days to form (Fig. 1A and E). At this stage, the cells had spindle-shaped morphology with distinct nucleus. However, after the first passage they changed to a small fibroblast-like morphology and maintained it thereafter (Fig. 1B and F). LDCs and FDCs were isolated from six different pigs; three liver samples came from three different pigs and three fat samples were obtained from three different pigs. Ten different clones from each cell type were isolated and expanded. Out of these three colonies (each coming from a different sample) from each cell type were selected to monitor their growth in culture. Both LDCs and FDCs proliferated robustly in the combination medium for more than 80 doublings without any change in morphology. All the colonies, irrespective of their origin, exhibited similar growth pattern and we did not observe senescence. Growth curves of one clone from LDCs and FDCs each are shown in Figure 1C and G. These clones were karyotyped at P30 (LDC) and P27 (FDC), and both had a normal chromosome number without any chromosomal abnormality (Fig. 1D and H). We extended the same procedure to other tissues—the adult kidney, heart, lungs, skin, and fetal liver—and used the same extraction protocol and culture conditions that we used for extraction of LDCs, and we got similar type of colonies from all the tissues. However, these colonies from adult tissues grew slowly, whereas fetal liver-derived colonies grew as efficiently as adult LDCs and the efficiency was 100 times more than the adult liver derived cell lines (data not shown).
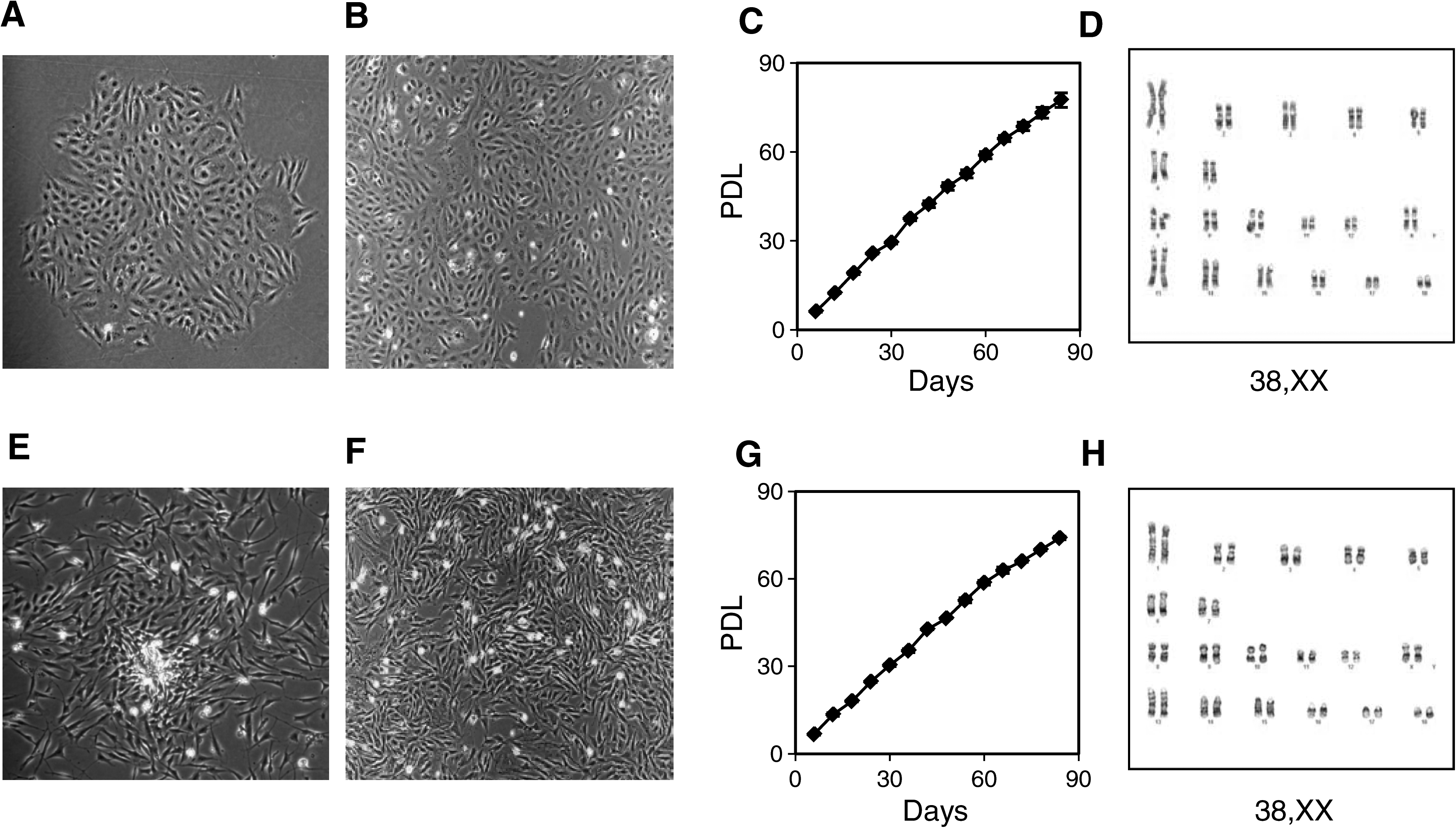
Characterization of cells derived from liver and fat. LDCs: (
To estimate the frequencies of LDCs and FDCs in liver and abdominal subcutaneous tissue of adult pigs, we used a CFU assay. We plated different numbers of viable cells per 10-cm2 plate and counted the number of colonies per plate on day 10 from the start of the culture. Only well-formed and distinct colonies were counted. The frequencies of these cell lines in adult liver and stromal cell fraction of abdominal fat were estimated as number of cells plated to get one colony. We obtained one LDC colony per 100,000 cells and one FDC colony per 962 cells (Table 1). The efficiency of FDCs colony generation was 104 times more than LDCs when the same number of liver cells and fat stromal cells were plated.
CFU, colony-forming unit; FDC, fat-derived cell lines; LDC, liver-derived cell lines.
Production of fetuses using porcine LDCs and FDCs as nuclear donors
LDCs and FDCs were used as nuclear donors in somatic cell nuclear transfer, and we were able to obtain healthy fetuses with both the cell types at day 30. A total of 582 cloned embryos (198 with LDCs, 186 with FDCs, and 198 with FFs) were transferred to six surrogate gilts on day 0 of the estrous cycle. Three of the animals became pregnant, one per cell type. Pregnancies were terminated at day 30 because of lack of resources to take them to term, and fetuses were obtained with all the three cell types. The fetuses were divided into three groups, that is, partially reabsorbed (small remnants of fetuses at the uterine attachment sites), dead (fetuses with severe discoloration), and normal (healthy-looking fetuses based on gross morphology) (Fig. 2). The efficiencies were 3.5, 2.1, and 4.0% for LDCs, FDCs, and FFs, respectively. The cells used as nuclear donors were at P12 (LDCs and FDCs) and P3 (FFs). SNP-based DNA fingerprinting was done with 96 Meat Research Center (MARC) standard SNP markers. SNPs that could be read across all samples of a subset were used for analysis. In the FFs set, out of 35 SNPs 13 are the same across all the samples, whereas 22 are shared between cells used as a nuclear donor and fetuses but are different from the recipient pig (Table 2). In the LDCs set, out of 43 SNPs 20 are the same across all the samples, whereas 23 are shared between cell used as a nuclear donor and fetuses but are different from the recipient pig (Table 3). In the FDCs set, out of 48 SNPs 24 are the same across all the samples, whereas 24 are shared between cell used as a nuclear donor and fetuses but are different from the recipient pig (Table 4). This analysis confirms that the fetuses are derived from respective cells by SCNT.

Fetuses obtained from SCNT. (
SNP, single nucleotide polymorphism; FF, fetal fibroblasts.
Ninety-six Meat Animal Research Center (MARC) standard SNP markers were tested. SNPs that could be read across all samples of a subset were used for analysis and shown in the tables. The SNPs that are same between the cells used as nuclear donors and fetuses but different from recipient pigs are color blocked.
Stable transfection of LDCs and FDCs
We tested whether these cells can be used for transgenesis. Both LDCs and FDCs were cultured to 60–70% confluency on collagen-coated plates, harvested, and electroporated with linearized DNA construct having neomycin acetyl transferase gene under a mammalian promoter. We achieved a transfection efficiency of 50% with all these cell types using microporation. Colonies with stable genetic modifications and extended expression of transgene were obtained when these cells were transfected with a plasmid containing neomycin resistance gene and selected under G418 (100 μg/mL for 10–14 days). The colonies obtained were healthy (Fig 3A and B) and grew very well without any change in cell morphology or growth characteristics. The DNA was extracted from these colonies, and a part of neomycin acetyl transferase gene was amplified by PCR to confirm that these colonies are transgenic (data not shown). As a comparison, we obtained a transfection efficiency of 50% with FFs (similar to LDCs and FDCs) using G418 at 100 μg/mL for 10 to 14 days as the selection agent (data not shown).
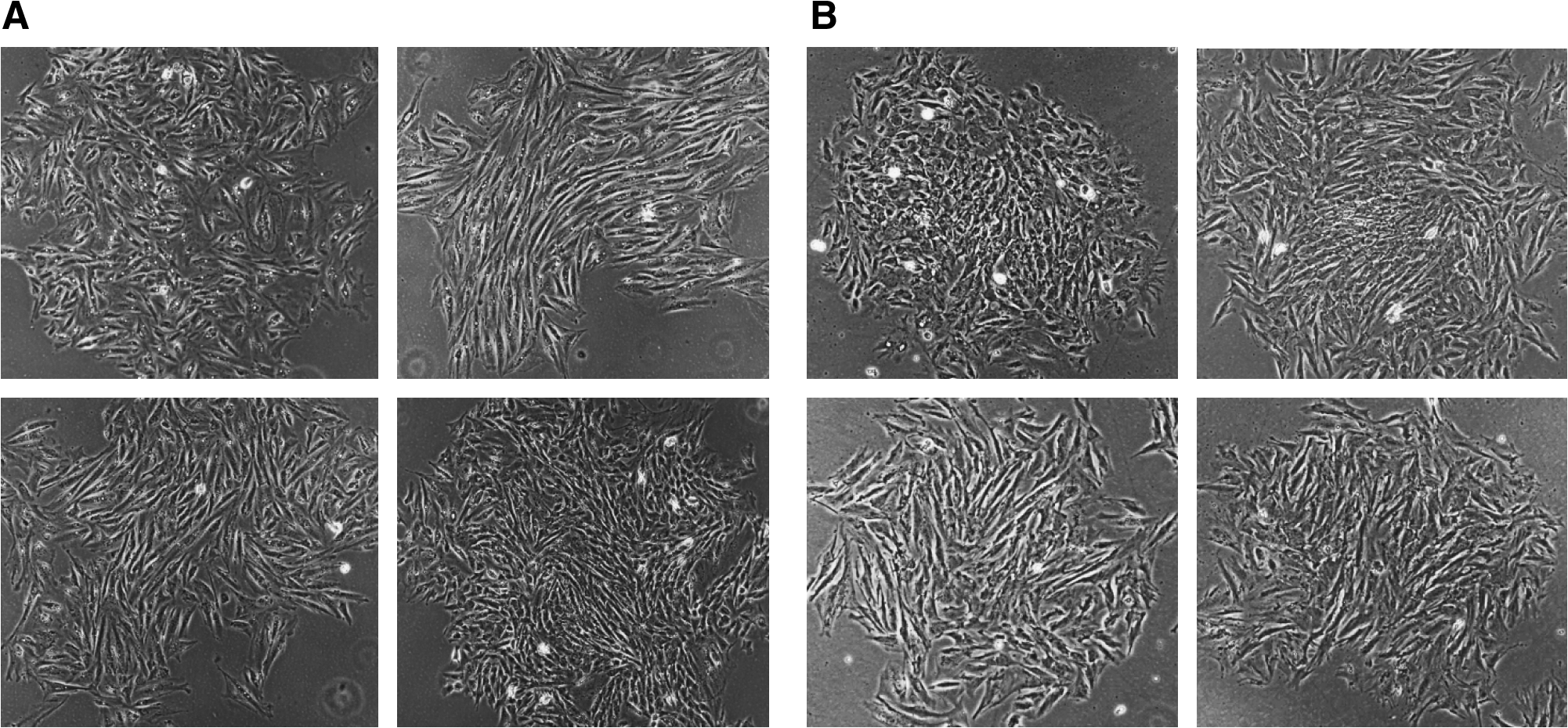
G418-resistant colonies obtained after transfection with linearized plasmid containing neomycin acetyl transferase gene and selection on G418 (100 μg/mL) for 10 days. (
Discussion
The scarcity of reagents to trace and characterize different cell types in a less molecularly characterized species like the pig makes it hard to identify different cells with the efficiency and speed as is possible in humans and mice. Therefore, the challenge is to devise simple and innovative strategies to identify different cell types for various applications. In the present study, we used a simple method to culture LDCs from an adult pig. We isolated hepatocytes as per the method described by Herrera et al. (2006), but manipulated the culture system in such a way that did not allow hepatocytes to attach. Under these conditions only a rare cell population present in the liver could attach and grow when maintained on a combination media (α-MEM/EGM-MV). The attached cells were visible the next day as single cells or clusters ranging from two to six cells/cluster. They were spindle-shaped, with a distinct nucleus and tapering thin bodies. It is apparent that these cells occur in clusters in the organ, and during extraction process some clusters remain intact while some break, and it is for this reason that we see cells as clusters ranging from one to six cells. These cells grow rapidly, the clusters better than individual cells, and form full-grown colonies in 10 to 14 days. After first passage, their morphology changed to small fibroblast-like, and they maintained this morphology throughout thereafter. We used same culture conditions for deriving cell lines from adipose tissue. We extracted stromal cell fraction from subcutaneous abdominal fat as per the standard method (Traktuev et al., 2009) and plated them under the same conditions as described for hepatocytes. Here again the media was not appropriate for stromal cells and we got the similar pattern as we got with liver cell lines; cell clusters ranging from oen to six cell per cluster and colony morphology similar to liver-derived cell lines. However, the efficiency of FDCs was 104 times more than LDCs (Table 1). This may be due to the fact that we extracted and plated all cell types from the liver and only stromal cell fraction from fat tissue. The stromal cell fraction may be enriched for long-living cells. In fact, we extended the same procedure to other tissues and we got similar type of colonies from all the tissues tested. It is evident that these culture conditions capture a very rare cell, may be a stem cell that is present in every organ, most likely originating from the endothelium that covers every organ.
Both these cell types proliferate rapidly and reached 80 population doublings in 91 days, that is, on an average of one cell division per day. We did not see any sign of replicative senescence or slowing of proliferation rate during this period. In fact, they are growing at present as efficiently as they were growing in the beginning. Other stem cells, both adult and fetal origin, reported so far undergo replicative senescence and exhibit a life span similar to or little more than fetal fibroblasts but significantly lower than these cells (Bosch et al., 2006; Colleoni et al., 2005; Dyce et al., 2004; Faast et al., 2006; Hao et al., 2009; Jin et al., 2007; Zhu et al., 2004). However, Brunetti et al. (2008) showed that they could extend the life span of pig fetal fibroblasts (PFF) and adult fibroblasts (PAF) to 110 and 87 population doublings, respectively, by adding bFGF (5 ng/mL) to the culture. We analyzed these cells for chromosomal abnormalities at PDL 70 (P30 for LDCs and P27 for FDCs) and all the cells checked (20 in each sample) had normal karyotype. These data indicate that these cells maintain normal karyotype after long-term in vitro culture. Further, we used these cells as nuclear donors in SCNT and compared their developmental potential with fetal fibroblasts. Based on the results we obtained, the efficiency of in vivo development at day 30 was comparable between LDCs and FFs, whereas FDCs were less efficient than FFs. However, it is hard to draw any conclusions based on the limited number of replications. Additional replicates will be necessary to establish any potential difference in the reprogramming ability between these cells and fetal fibroblasts. The cells used as nuclear donors were at P12 (LDCs and FDCs) and P3 (FFs). This indicates that even at higher passage level, these cells from adult origin have comparable nuclear transfer efficiency with early passage fetal fibroblasts. Further, we tested these cells for transgenesis and achieved a transfection efficiency of 50% with both these cell types using microporation. Cell lines with stable genetic modifications and extended expression of transgene were obtained when these cells were transfected with a plasmid containing the neomycin resistance gene and selected under G418. Therefore, these cells will be very useful for cloning desired adult animals and for producing transgenic pigs for various applications. It will be interesting to compare the full-term development of these cells via SCNT, and gene targeting efficiency to fetal fibroblasts; a better comparison may be between FFs and LDCs obtained from the same fetus. It will be informative to characterize these cell types at molecular level in detail to see whether they express various stem cell markers. It will also be interesting to see if similar type of cell can be obtained from other animal species (farm animals and endangered animals).
In conclusion, we report the isolation of cell lines with long life span from adult porcine liver and adipose tissue with a simple culture system. These cells will increase the repertoire of cell types available for cloning. Further, they can be maintained in culture for a long period of time without undergoing any chromosomal abnormality. That will provide an extended time for complex genetic manipulations.
Footnotes
Acknowledgments
This work was supported by Clarian Transplant Institute (CTI). We thank A. Joseph Tector, Chief of Transplant Division at Clarian Transplant Institute, for his support. We also thank Methodist Research Institute staff for help with the surgery, feeding, and taking care of the pigs. We thank Bess Ivary for help with the surgeries, embryo transfer, and fetus harvest. All the animals used and procedures performed on animals in this study were approved by Institutional Animal Care and Use Committee (IACUC).
Author Disclosure Statement
The authors declare no conflicting financial interests exist.