
Abstract
Abstract
As the phenotype of a given single-gene mutation in mice is modulated by the genetic background of the inbred strain, embryonic stem (ES) cells derived from various inbred mouse strains are required to produce gene-targeted mice without the need for backcrossing and for detailed analysis of gene function in vivo. Here, we performed a comparative investigation of the effects of three culture conditions, LIF + KSR/ES medium described previously, High LIF + KSR/ES medium and iSTEM + LIF medium containing three inhibitors of glycogen synthase kinase 3, mitogen-activated protein kinase kinase, and fibroblast growth factor receptor signaling (3i), on the establishment of germline-competent ES cells derived from strains BALB/c and NZB mice. The results indicated that LIF + KSR/ES medium was permissive for the derivation of ES cells from NZB mice, which contribute to the somatic lineage in vivo, but not to the germline lineage. In contrast, ES cells that contribute to the makeup of chimeric mice were not propagated from blastocysts of BALB/c mice. Both germline and somatic competency were improved by increased LIF concentration in cultures of BALB/c ES cells, although we failed to establish germline-competent NZB ES cells using the same concentration of LIF. Unexpectedly, iSTEM + LIF medium containing 3i showed a negative effect on the derivation of NZB ES cells with normal chromosome numbers, but not on the maintenance of previously established ES cells. Our findings suggest that the stability of pluripotency in the inner cell mass isolated from blastocyst embryos may differ according to the genetic background of inbred mouse strains, and that although the concentration of LIF is a determinant for authentic pluripotency, including germline and somatic competency in BALB/c ES cells, additional factor(s) are required for commitment to germline lineage independent of somatic lineage in NZB ES cells.
Introduction
In addition to C57BL/6 strains, a wide range of inbred mouse strains have been characterized in detail and are widely used in biological and medical research according to phenotypic requirements. A variety of ES cell lines derived from inbred mouse strains will provide novel insight into gene function that cannot be obtained from conventional ES cells. However, germline-competent ES cells have only been established from a few mouse strains. Therefore, the present study was performed to establish authentic ES cells derived from various inbred mouse strains for use in the generation of gene-targeted mice without the requirement of backcrossing.
Previously, we showed that supplementation of the medium with 20% knockout serum replacement (KSR) and 1000 IU/mL leukemia inhibitory factor (LIF) and eight-cell blastomeres as recipient embryos allowed establishment of ES cells derived from C57BL/6J and C57BL/6N mice and those successful production of germline chimeras, respectively (Shimizukawa et al., 2005; Tanimoto et al., 2008). In contrast to C57BL/6, preliminary findings in our laboratory indicated that BALB/c and NZB mice are not permissive for the development of germline-competent ES cell lines under the culture conditions described above. As BALB/c mice are useful for research into both cancer and immunology and as NZB mice are a model for autoimmune disease, further progress in medical research will require the development of ES cells derived from both of these strains. Recent reports have indicated that combinations of chemical agents with suppressive effects against glycogen synthase kinase 3 (GSK3), mitogen-activated protein kinase kinase (MAPK/ERK kinase or MEK) and fibroblast growth factor (FGF) receptor signaling (hereafter called 3i) are useful for isolating pluripotent stem cells, and allow derivation of the ICM of blastocysts from nonpermissive species, such as the rat (Buehr et al., 2008; Li et al., 2008; Ying et al., 2008). Although these chemical approaches appear to be effective for the establishment of ES cells derived from intractable mouse strains, insufficient data are available from which to make definitive conclusions.
To address these issues, we performed a comparative investigation of the effects of three culture media, LIF + KSR/ES medium, High LIF + KSR/ES medium, and iSTEM + LIF medium containing 3i, on the establishment of germline-competent ES cells from strains BALB/c and NZB. Our findings suggest that the stability of pluripotency in the ICM isolated from blastocyst embryos differ according to the genetic background of inbred mouse strains and that although the concentration of LIF is a determinant of authentic pluripotency, including germline and somatic competency in BALB/c ES cells, additional factor(s) are required for commitment to germline lineage independent of somatic lineage in NZB ES cells.
Materials and Methods
Animals
BALB/cAJ, ICR (CD1) and C57BL/6J mice were purchased from CLEA Japan. NZB/BINJ mice were obtained from the Jackson Laboratory. Animals were kept in plastic cages (four to five per cage) under pathogen-free conditions (sentinel mice were examined periodically throughout the study) in a room at 23.5 ± 2.5°C and 52.5 ± 12.5% relative humidity under a 14:10-h light:dark cycle. Mice had free access to commercial chow (CE2, CLEA Japan) and filtered water throughout the study. Animal experiments were carried out humanely in accordance with the Regulations for Animal Experiments of the University of Tsukuba and Fundamental Guidelines for Proper Conduct of Animal Experiment and Related Activities in Academic Research Institutions under the jurisdiction of the Ministry of Education, Culture, Sports, Science, and Technology of Japan and with approval from the Institutional Animal Experiment Committee of our university.
Embryos
Naturally fertilized embryos were collected at the eight-cell stage from female mice treated with pregnant mare serum gonadotropin (PMSG) and human chorionic gonadotropin (hCG). Eight-cell embryos were cultured in KSOM medium (Millipore, Bedford, MA, USA) until the blastocyst stage, at 37°C under an atmosphere of 5% CO2 and 95% air.
Media
For culture of ES cells, Dulbecco's modified Eagle's medium (Invitrogen, Carlsbad, CA, USA) were supplemented with 20% KSR (Invitrogen), 0.1 mM 2-mercaptoethanol, nonessential amino acids, sodium pyruvate, and 1000 or 5000 IU/mL LIF (Millipore). iSTEM medium (Stem Cell Sciences, Palo Alto, CA, USA) was composed of N2B27 with three signal inhibitors (3i; final concentrations: 0.8 μM PD184352, 2 μM SU5402, and 3 μM CHIR99021).
Establishment and culture of ES cells
Blastocysts from which the zonae pellucidae had been removed with acidic Tyrode's solution (Irvine Scientific, Santa Ana, CA, USA) were placed on a feeder layer of mitomycin C-inactivated confluent embryonic fibroblasts prepared from BALB/cAJcl fetuses at 14 days post coitus in four-well dishes. A vigorous ICM appeared within the attached blastocysts by 5 days of cultivation in the medium described above. To expand the next generation, the ICM was picked up with a micropipette, dissociated into single cells with 0.25% trypsin/EDTA, and seeded onto a new feeder layer. Medium was changed every day, and cells were split with 0.25% trypsin/EDTA every 2–4 days. The putative ES cells were frozen at a concentration of 2 × 106 cells per vial after passage 5. Cultures were maintained in a humidified incubator at 37°C under an atmosphere of 5% CO2 and 95% air.
Alkaline phosphatase activity
The putative ES cells were stained for cytoplasmic alkaline phosphatase activity using a commercial alkaline phosphatase detection kit (Millipore). Fixation and staining were performed according to the manufacturer's protocol.
Immunofluorescence staining
Cells were fixed in 4% paraformaldehyde for 10 min at room temperature and incubated overnight at 4°C with primary antibody against Oct3/4 (c-10, Santa Cruz, Santa Cruz, CA, USA; 1:250) or Nanog (ab21603, ReproCELL, Livermore, CA, USA; 1:400). Alexa Fluor fluorescent secondary antibodies (Invitrogen) were used at 1:500 dilution. Nuclei were visualized with DAPI staining.
Chromosome number analysis
ES cells were arrested in metaphase with colcemid at a final concentration of 0.02 μg/mL. The ES cells were trypsinized, swelled with 75 mM KCl, fixed with a solution of methanol and acetic acid (3:1), and put on slides. After desiccation, chromosomes were stained with 5% Giemsa solution. The chromosome numbers of 50 well-spread metaphases for each ES cell lines were evaluated by microscopy and counted on the micrographs.
Production of chimeric mice by coculture aggregation method
Frozen–thawed ES cells were placed on a feeder layer 2 days before embryo manipulation. The propagated ES cells were dissociated with 0.25% trypsin/EDTA immediately before coculture aggregation. Embryos at the two-cell stage were collected from female mice that had been superovulated and mated naturally, and were then cultured to the eight-cell stage in KSOM medium. From 15 to 20 ES cells derived from BALB/c and NZB mice were aggregated with eight-cell stage embryos from C57BL/6J and ICR, respectively, in KSOM. Manipulated chimeric embryos were transferred at the blastocyst stage into the uterine horns of ICR mice at 2.5 days post coitus. The somatic competency of ES cells was determined only by the coat color chimerism. The germline competency of ES cells was examined by crossing chimeric mice with ICR mice and then was evaluated by coat color pigmentation in the offspring.
Results
Establishment of BALB/c and NZB ES cells using ES medium containing 20% KSR and 1000 IU/mL LIF (LIF + KSR/ES medium)
We demonstrated previously that supplementation of medium with 20% KSR and 1000 IU/ml LIF is extremely effective for derivation of C57BL/6J and C57BL/6N ES cells with high capacity for germline differentiation as well as somatic differentiation (Tanimoto et al., 2008). KSR is a serum-free formulation designed to directly replace fetal bovine serum (FBS) in ES cell culture and the cytokine LIF is known to be essential for maintenance of mouse ES cells in an undifferentiated state (Smith et al., 1988; Williams et al., 1988). LIF signaling pathways, including Jak-STAT3, PI(3)K-Akt, and MAP kinase pathways, are linked to the pluripotency-associated transcription factors Oct3/4, Sox2 and Nanog (Niwa et al., 2009).
First, we investigated the establishment of ES cells from the blastocysts of BALB/c and NZB mice using LIF + KSR/ES medium, which is effective for derivation of C57BL/6 ES cells. Blastocysts developed in vitro from eight-cell stage embryos obtained from BALB/c and NZB mice via natural mating and superovulation with PMSG–hCG, were cultivated on a feeder cell layer in LIF + KSR/ES medium. Four ES cell lines were isolated from 29 BALB/c blastocysts (13.7%), which formed a smooth outline and piled up on the feeder layer. Six ES cell lines were established from 20 NZB blastocysts (30.0%). The chromosome numbers of 50 cells in each established ES cell line were counted to test for chromosomal abnormalities. The majority of cells in these populations had the normal complement of 40 chromosomes (two of three and three of five cell lines from BALB/c and NZB mice, respectively) (Table 1). Based on their karyotypes and the results of proliferation assay (data not shown), we selected two ES cell lines for further study: BALB-1000-26 derived from BALB/c (Fig. 1a) and NZB-1000-39 derived from NZB (Fig. 1b). These cell lines were morphologically indistinguishable from B6J-25, a C57BL/6 ES cell line with adequate germline competency (Fig. 1c). Furthermore, these cell lines exhibited positive staining for alkaline phosphatase (data not shown) and expressed pluripotent markers Oct3/4 and Nanog (Fig. 1d).
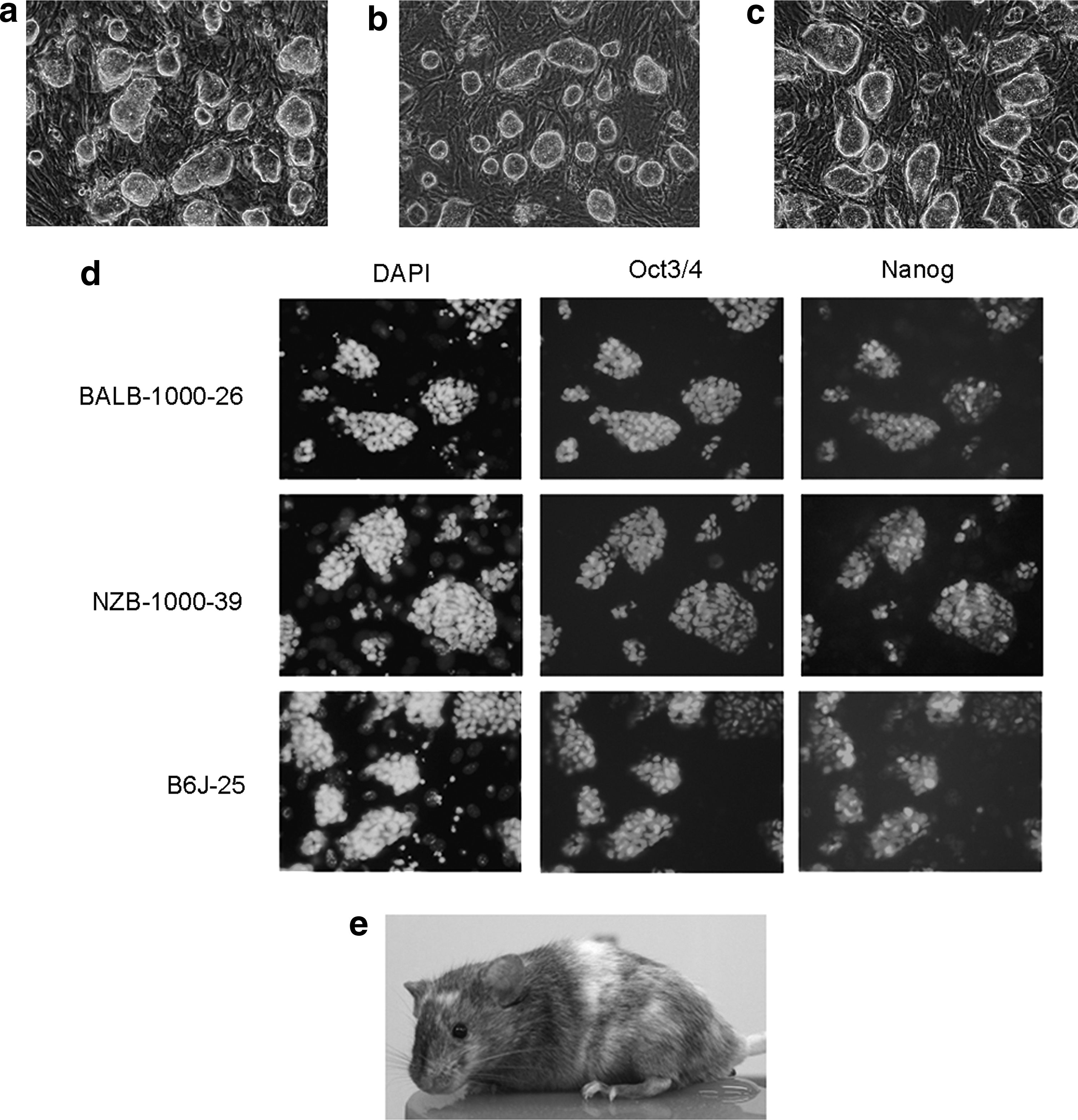
Established ES cells using LIF + KSR/ES medium. (
For each cell line, 50 cells were used.
To investigate the pluripotency in vivo, the ES cell lines were aggregated with eight-cell stage embryos (Table 2). Forty-nine chimeric embryos derived from BALB-1000-26 cells were surgically transferred into the uteri of psudopregnant female mice, and coat color chimeras were not observed in 13 pups. In those from NZB-1000-39 cells, 100 chimeric embryos yielded five chimeric mice; the rates of chimerism assessed by coat color were relatively low (Fig. 1e) and there were no germline transmission of the ES cells. These results indicated that in KSR/ES medium, which was supplemented with 1000 IU/mL LIF, the strains BALB/c and NZB are nonpermissive to the establishment of germline-competent ES cells.
No. of mice born/no. of chimeric embryos transferred.
No. of chimeric mice/no. of chimeric embryos transferred.
Degree of chimerism is determined by coat color.
No. of 100% chimeric mice/no. of chimeric embryos transferred.
NE, not examined.
Establishment of BALB/c and NZB ES cells using KSR/ES medium containing 5000 IU/mL LIF (High LIF + KSR/ES medium)
It was reported previously that ES medium containing of 5000 IU/mL LIF facilitated derivation of BALB/c ES cells (Baharvand and Matthaei, 2004). To examine whether the addition of 5000 IU/ml LIF is effective for derivation of germline-competent ES cells, we established ES cells from the blastocysts of BALB/c and NZB mice using ES medium containing 20% KSR and 5000 IU/ml LIF.
Nine ES cell lines were derived from 106 BALB/c blastocysts (8.5%) and three ES cell lines were derived from 20 NZB blastocysts (15.0%). All of the cell lines derived in High LIF + KSR/ES medium showed typical morphology of ES cell colonies. We analyzed the chromosome numbers to examine the properties of ES cells; 40 chromosomes were seen in the majority of populations (six of six and two of two cell lines of BALB/c and NZB, respectively) (Table 3).
For each cell line, 50 cells were used.
To further examine the features of pluripotency, we used two ES cell lines, BALB-5000-5 derived from BALB/c (Fig. 2a) and NZB-5000-6 derived from NZB (Fig. 2b). Both cell lines exhibited ES cell markers including alkaline phosphatase (data not shown), Oct3/4 and Nanog (Fig. 2c), and there were no differences in the expression patterns of pluripotent markers between the two cell lines.
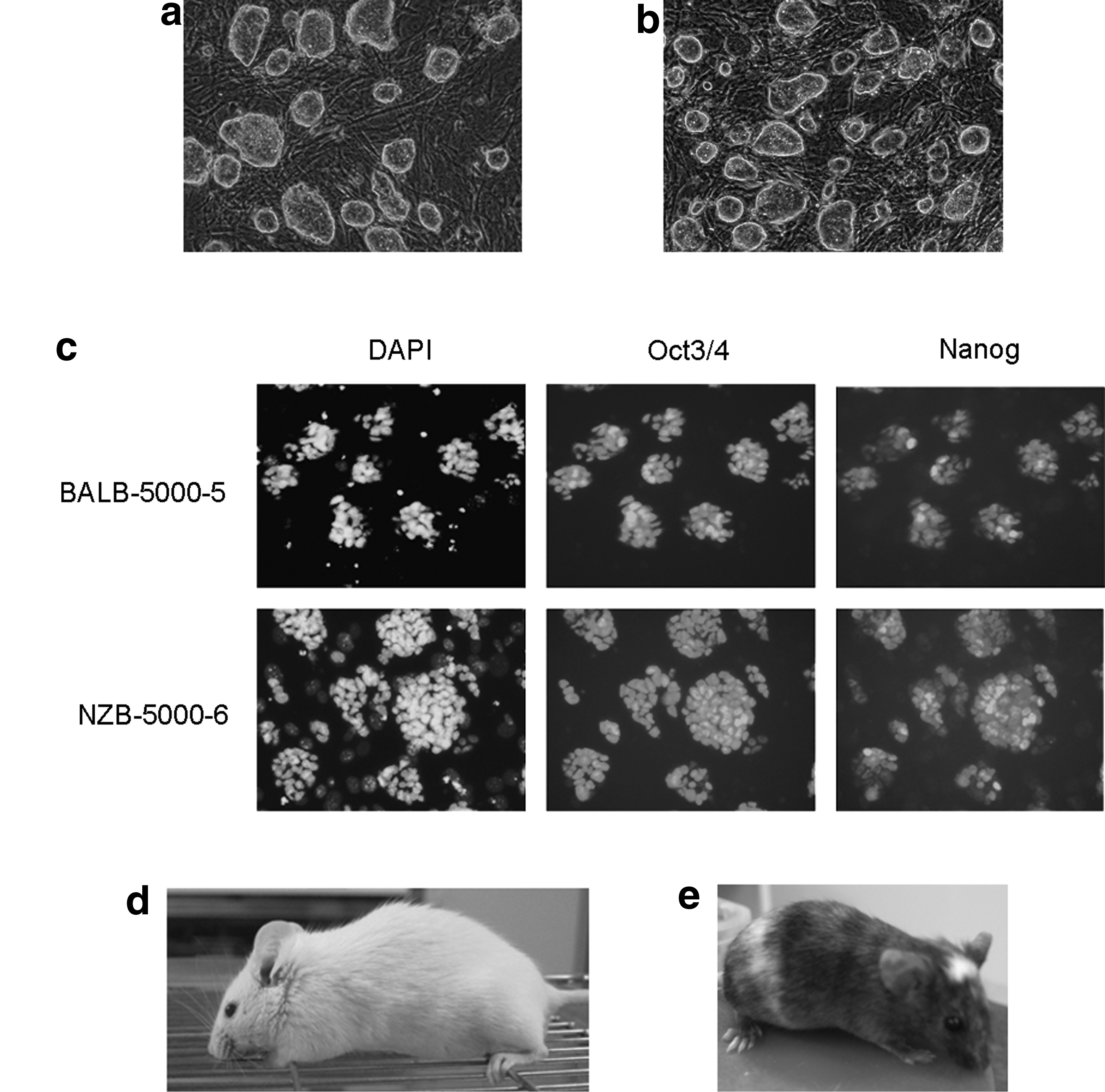
Established ES cells using High LIF + KSR/ES medium. (
As shown in Table 4, BALB-5000-5 cells were aggregated with 100 eight-cell stage embryos, yielding six chimeric mice; five of these offspring were chimeric mice with a completely ES cell-derived coat color (Fig. 2d). Moreover, all three completely white chimeras examined were germline chimeras, suggesting that LIF plays an important role in the establishment of germline-competent ES cells. In contrast, we obtained no germline chimeras among four chimeric mice derived from 80 chimeric embryos composed of NZB-5000-6 cells and eight-cell stage embryos (Fig. 2e). These results suggested that increasing the concentration of LIF in KSR/ES medium had no effect on the establishment of germline-competent NZB ES cells and that there are differences between mouse strains in the response to LIF.
No. of mice born/no. of chimeric embryos transferred.
No. of chimeric mice/no. of chimeric embryos transferred.
A degree of chimerism is determined by coat color.
No. of 100% chimerism mice/no. of chimeric embryos transferred.
Establishment of NZB ES cells using ES medium containing 3i and 1000 IU/mL LIF (iSTEM + LIF medium)
We investigated whether addition of not only LIF but also other factors allow the establishment of germline-competent ES cells from NZB mice. We used iSTEM (SCF-SF-ES-01; Stem Cell Sciences) consisting of N2B27 as the basal medium and three inhibitors: CHIR99021, PD184352, and SU5402; CHIR99021 is the most selective inhibitor of GSK3 reported to date (Ring et al., 2003), while PD184352 (Bain et al., 2007; Davies et al., 2000) and SU5402 (Mohammad et al., 1997) are selective pharmacological inhibitors of MEK and FGF receptor tyrosine kinase, respectively. It has been reported that 3i can support the efficient establishment and maintenance of germline-competent ES cells from mice (Ohta et al., 2009; Ying et al., 2008) and rats (Buehr et al., 2008; Li et al., 2008).
We established eight ES cell lines from 27 plated NZB blastocysts (30.0%). The ES cells derived in 3i formed smaller clumps than typical ES cell colonies (Fig. 3a), although these cells were also positive for alkaline phosphatase assay (data not shown) and Oct3/4 and Nanog expression (Fig. 3b). Surprisingly, the majority of cell lines showed an abnormal karyotype (Table 5). To examine the capacity to contribute to chimeric mice, NZB-(i)-4 cells were aggregated with ICR eight-cell stage embryos. Transfer of 100 chimeric embryos yielded 9 chimeric mice, among which the highest degree of chimerism was markedly lower than that in chimeric mice derived from NZB-5000-6 ES cells (Fig. 3c). They failed to achieve germline transmission of NZB-(i)-4 cells.
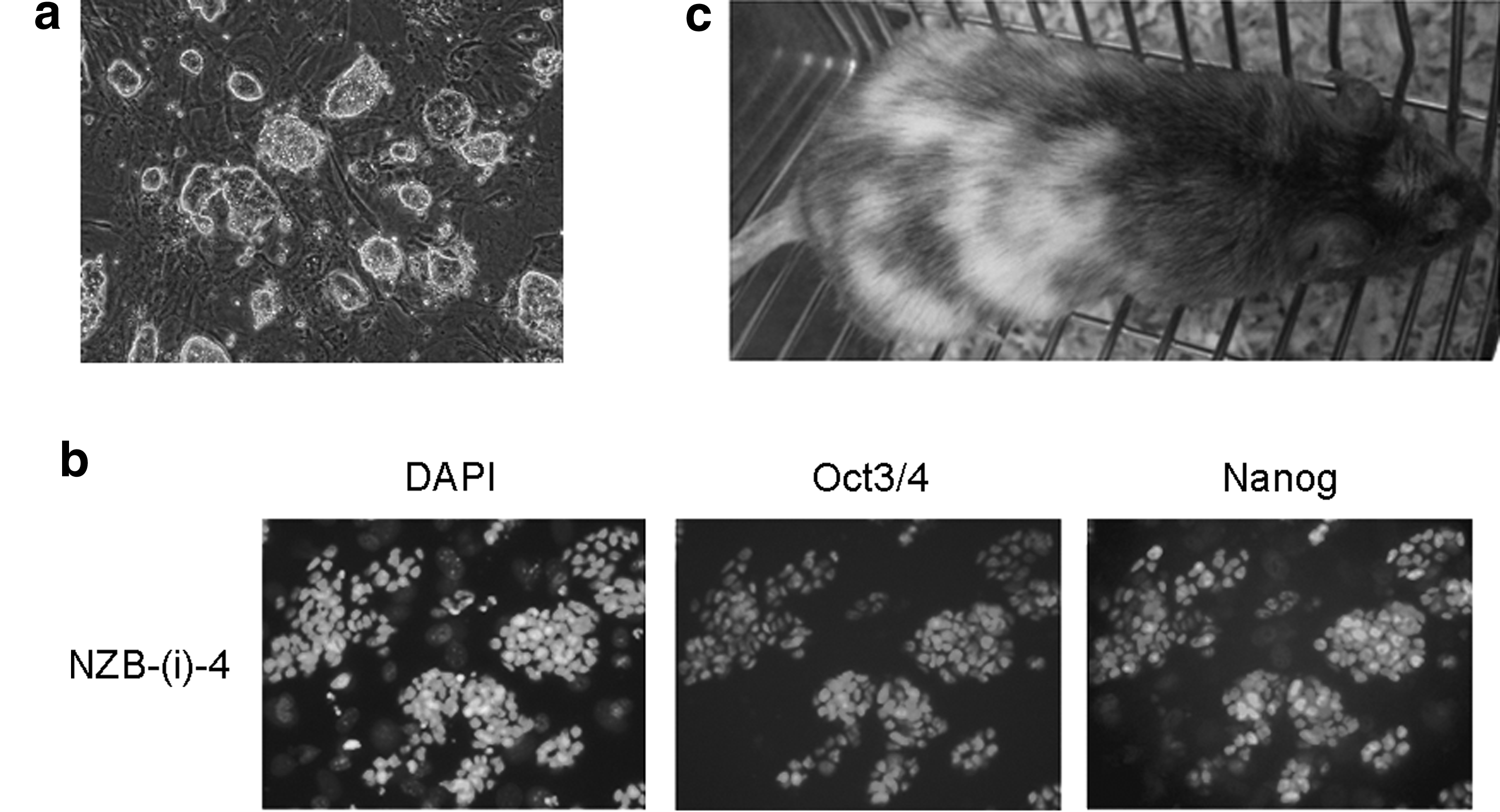
Established NZB ES cells using LIF + iSTEM medium. (
For each cell line, more than 50 cells were used.
Effects of 3i on maintenance of chromosome number
To investigate the improvements in culture condition for NZB ES cell derivation, we examined whether the combination of three signal inhibitors induces abnormalities in ES cell chromosome number. Germline-competent B6J-25 ES cells, which were previously established from blastocysts of C57BL/6J and NZB-5000-6 ES cells with normal chromosome numbers but that did not have the germline competency were propagated in LIF + KSR/ES medium containing 3i and chromosome number was counted at passage 7. The morphology of B6J-25 and NZB-5000-6 cells observed before passage (passage 0) were maintained to passage 7 in 3i conditions as in control conditions (Fig. 4a). Further, in B6J-25 and NZB-5000-6 cells at passage 7, there was no difference in the percentage of ES cells with 40 chromosomes between control and 3i conditions (Fig. 4b). Moreover, immunofluorescence staining indicated that NZB ES cells cultured in 3i expressed Oct3/4 and Nanog as under control conditions (Fig. 4c). These results suggested that 3i did not have a negative effect on the maintenance of morphology, chromosome number, or expression of pluripotent markers in established ES cells. Instead, culture in 3i showed a negative effect on the achievement of complete pluripotency of cells derived from the blastocysts of NZB mice in the early stages of establishment.
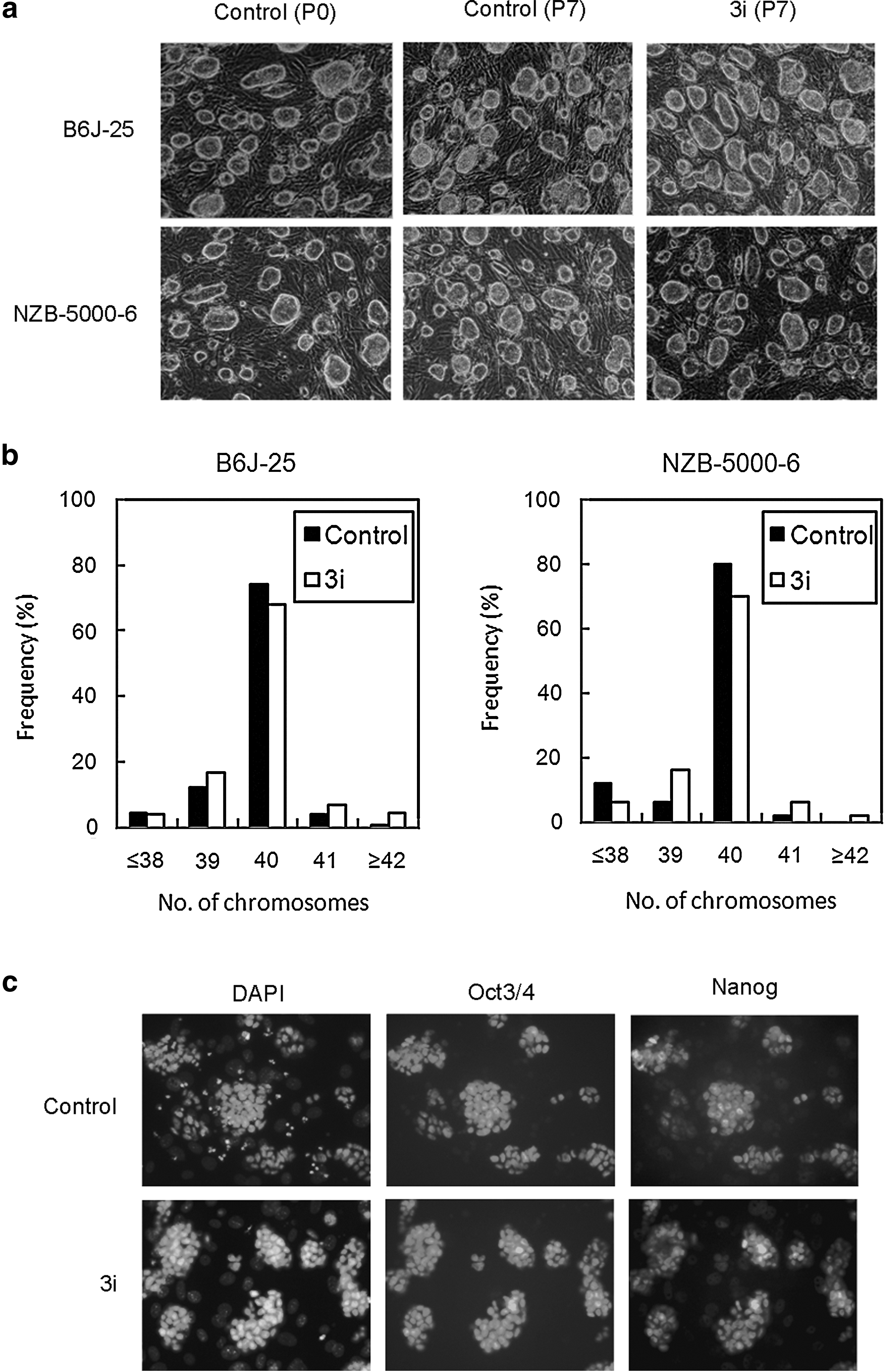
Effects of signal inhibitors on maintenance of chromosome number in established ES cells. (
Discussion
The present study was performed to establish ES cells from BALB/c and NZB blastocysts. Three different culture conditions were investigated for successful establishment of germline-competent ES cells.
The results of the present study indicated that although addition of 1000 IU/mL LIF was effective in derivation of C57BL/6 ES cells (Tanimoto et al., 2008), 5000 IU/mL LIF supported the effective establishment of germline-competent ES cells from BALB/c mice. However, increasing the concentration of LIF in medium was not effective in NZB mice. These observations suggested that the response to LIF in ES cell derivation is dependent on genetic background. Recent studies have indicated that the establishment of ES cells derived from C57BL/6 and CBA mice, both of which are nonpermissive for ES cell derivation, requires reduction of Erk activation, whereas this is not necessary in the permissive strain 129 (Battle-Morera et al., 2008). Similarly, Hanna et al. (2009) reported that the establishment of ES cells derived from the nonpermissive NOD strain, but not the permissive strain 129, required constitutive expression of Klf4, c-Myc, or chemical agents that can replace these factors. Therefore, ongoing research has begun to elucidate the differences between strains permissive and nonpermissive for ES cell derivation. Differences in response to LIF between strains may reflect the efficiency of ES cell derivation.
Chambers et al. (2007) reported that ES cells with conditional loss-of-function mutation of the Nanog gene exhibited multilineage differentiation in both fetal and adult chimeras, but did not contribute to the germline. As this defect was rescued by repair of the mutant allele, they suggested that Nanog is dispensable for somatic pluripotency but is required for the production of germ cells. However, although NZB-1000-39, NZB-5000-6, and BALB-1000-26 cell lines expressed Nanog, they were not transmitted through the germline. These results suggest that NZB-1000-39, NZB-5000-6, and BALB-1000-26 cell lines may have lost or may not have gained as yet unidentified factors that are necessary to contribute to the germ lineage during establishment of ES cells. Epiblast stem cells (EpiSCs) are derived from postimplantation mouse epiblasts in culture conditions without LIF but including bFGF and Activin (Brons et al., 2007; Tesar et al., 2007). These cells display characteristic hallmarks of pluripotent stem cells, such as the expression of Oct3/4, Sox2, and Nanog, but differ from ES cells in expression of several other transcripts. EpiSCs are also able to differentiate into various cell types in vitro, but do not contribute to chimeric mice. Recently, bFGF, Activin, and BIO-derived stem cells, FAB-SCs, that share a number of features with EpiSCs, were derived from mouse blastocysts under EpiSCs culture conditions (Chou et al., 2008). FAB-SCs also express Oct3/4, Sox2, and Nanog, but these cells do not show hallmarks of pluripotent differentiation, such as in vitro embryo body formation, teratoma formation, and contribution to chimeras. Therefore, BALB-1000-26 cells that are not incorporated to developing embryos may be in an EpiSCs-like pluripotent state. Furthermore, NZB-1000-39 and NZB-5000-6 cells may be in a pluripotent state intermediate between those of authentic ES cells and EpiSCs because these cell lines show somatic pluripotency but are incapable of contributing to the germ lineage. To understand these pluripotent states, further studies are required to examine the expression of not only genes expressed in ES cells but not in EpiSCs, such as Rex1, Nr0b1, and Klf4, but also genes expressed in EpiSCs but not in ES cells, such as Fgf5, T (brachyury) and Lefty.
It is also possible that NZB-1000-39 and NZB-5000-6 cells are competent to colonize germ cells. Generally, blastocyst injection is performed to produce germline chimeric mice. However, efficient colonization of germ cells by TT2 ES cells generated from (C57BL/6 × CBA)F1 embryos occurred when the cells were introduced into eight-cell stage embryos but not when introduced into blastocysts (Yagi et al., 1993). Further, simple aggregation of R1 ES cells derived from (129/Sv × 129/SvJ)F1 mice with morula-stage embryos resulted in viable chimeras at a similar frequency to that observed after blastocyst injection (Wood et al., 1993). We also showed that eight-cell embryos are efficient recipients of C57BL/6 ES cells for generation of germline chimeras (Tanimoto et al., 2008). These results indicated that the stage of the recipient embryos is important for generation of germline chimeras. Moreover, the genetic background of the recipient embryos is also an important determinant of germline transmission of the resulting chimeric mice (Araki et al., 2009; Dinkel et al., 1999; Lemckert et al., 1997). However, we also observed the failure of ES cells to yield germ cells in chimeras when NZB-5000-6 cells were aggregated with eight-cell stage embryos of BALB/c mice and injected into blastocysts of ICR mice (data not shown). Therefore, pluripotent states appear to be more important in the in vivo germline differentiation of ES cells derived from NZB mice than developmental stages or strains of the recipient embryos.
Anuploid cells were observed in relatively large numbers in NZB ES cell lines established under 3i conditions. We predict that 3i may influence chromosome number in the first step of the transition from ICM to stable ES cell line as 3i conditions did not induce the instability of chromosome context in previously established ES cells. Although in rat ES cell derivation using 3i, trisomy for chromosome 9 was observed (Buehr et al., 2008), no such abnormalities in chromosome number have yet been reported in mice. In the additional experiments using iSTEM + LIF medium, we revealed that most of established BALB/c ES cell clones showed ES-like cell colony formation (Supplementary Fig. S1; see online supplementary material at www.liebertonline.com), and that half of the established BALB/c ES cell clones had a normal karyotype (Supplementary Table S1). Therefore, the stability of pluripotency in the ICM isolated from blastocyst embryos may differ according to the genetic background of inbred mouse strains, and although the concentration of LIF is a determinant for authentic pluripotency including germline and somatic competency in BALB/c ES cells, additional factor(s) are required for commitment to germline transmission independent of somatic differentiation in ES cells of NZB mice.
In the present study, we were successful in establishing BALB/c ES cells with germline competency, suggesting that the BALB/c ES cells are a useful tool for the generation of BALB/c knockout mice. On the other hand, NZB ES cells might be useful for differentiation studies. In the supplemental experiments using NZB-5000-6 ES cells, which express enhanced green fluorescent protein (EGFP) under the beta-actin promoter, we produced a number of chimeric fetuses that emitted green fluorescence in a variety of tissues in the head, back, lower abdomen, and extremities (Supplementary Fig. S2). Therefore, NZB-5000-6 ES cells have potential not only to contribute to the development of coat color, but also to the differentiation into cells belonging to a wide variety of somatic cell lineages, suggesting that the NZB ES cells are a useful tool for basic research in the field of regenerative medicine.
As ES medium containing the two inhibitors, CHIR99021 and PD0325901 (Bain et al., 2007), which are more potent but equally MEK inhibitor compared with PD184352, allowed the establishment of germline-competent ES cells from NOD mice (Hanna et al., 2009; Nichols et al., 2009), which are known as a nonpermissive strain for ES cell derivation, and resolved chromosomal abnormalities seen in rat ES cells (Buehr et al., 2008), the 2i condition might sustain the complete pluripotency of isolated ICM derived from NZB mice. The development of an efficient method to establish germline-competent NZB ES cells would be of value for derivation of ES cells in various inbred mouse strains.
Footnotes
Acknowledgments
This work was supported by grants from the Ministry of Education, Science, Sports, and Culture of Japan (to F.S. and K.Y.) and RIKEN (to F.S.).
Author Disclosure Statement
The authors declare that no conflicting financial interests exist.
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.