
Abstract
Abstract
Recent results have shown that bone marrow mesenchymal stem cells (BMSCs) from human first-trimester abortus (hfBMSCs) are closer to embryonic stem cells and perform greater telomerase activity and faster propagation than mid- and late-prophase fetal and adult BMSCs. However, no research has been done on the plasticity of hfBMSCs into neuronal cells using single-cell cloned strains without cell contamination. In this study, we isolated five single cells from hfBMSCs and obtained five single-cell cloned strains, and investigated their biological property and neuronal differentiation potential. We found that four of the five strains showed similar expression profile of surface antigen markers to hfBMSCs, and most of them differentiated into neuron-like cells expressing Nestin, Pax6, Sox1, β-III Tubulin, NF-L, and NSE under induction. One strain showed different expression profile of surface antigen markers from the four strains and hfBMSCs, and did not differentiate toward neuronal cells. We demonstrated for the first time that some of single-cell cloned strains from hfBMSCs can differentiate into nerve tissue-like cell clusters under induction in vitro, and that the plasticity of each single-cell cloned strain into neuronal cells is different.
Introduction
Materials and Methods
Preparation of hfBMSCs
In our previous study, five human abortuses at the age of 10 to 12 weeks, with permission from the patients, hospital, and the Ethics Committee of Northwest A&F University, were obtained from a local hospital and used to isolate hfBMSCs (Zhang et al., 2008). Briefly, the limb long bones of the abortuses were taken out on a germ-free bench and washed with phosphate-buffered saline (PBS) containing 100 IU/mL penicillin and 100 IU/mL streptomycin in one 9-cm Petri dish. Their two cartilage ends were cut and medullary cavity opened with scissors lengthwise followed by calm swill with Minimum Essential Medium alpha (α-MEM; Gibco, Billings, MT, USA) containing 100 IU/mL penicillin and 100 mg/mL streptomycin. Bone bits were taken off and the swilled medium was filtered through 200-mesh nylon net. The filtrate was centrifuged for 5 min at 1500 rpm and the supernatant was discarded. The deposited marrow cells were cultured in plastic culture dishes on α-MEM containing 10% fetal calf serum (FCS; Stemcell Technologies Inc., Vancouver, Canada), 100 IU/mL penicillin, 100 mg/mL streptomycin, and 0.1 mmol/L β-mercaptoethanol (Sigma, Loveland, CO, USA). The culture medium was changed every 2 days, and blood cells were gradually removed by the medium changes. When they grew to monolayer and 80% confluency, fusiform cells were digested with 2.5 g/L trypsin (Gibco) containing 0.4 g/L edetic acid (Invitrogen, Carlsbad, CA, USA) and passaged with the same medium. Isolated cells of passage three were identified with flow cytometer (Beckman Coulter Inc., Fullerton, CA, USA) for antigen markers of the putative MSCs, CD29, CD44, CD71, CD105, and CD166, and those of hemopoietic stem cells, CD11a, CD14, CD34, and CD45 (all from Beckman Coulter Inc.) (Zhang et al., 2008). Then the isolated hfBMSCs were cryopreserved in liquid nitrogen.
Isolation, culture, and identification of single-cell cloned strains
In the successive double dilution, α-MEM + 20% FCS + 0.1 mmol/L β-mercaptoethanol was added in all wells of a 96-well culture plate (100 μL/well). And 100 μL of 1 × 104/mL cell suspension of hfBMSCs from passage 3 was added in each well in the first column of the plate and equably mixed. Then 100-μL mixed cell suspension was transferred from the first to the second well in each row, and 100-μL mixed cell suspension was transferred from the second to the third well in the same row. Other wells of each row were successively diluted to the 12th well in the same way. And 100-μL mixed cell suspension was removed from the last well of each row. The 96-well culture plate was put in a culture box and incubated for 24 h, then the wells with only one adhered cell was marked under inverted microscopy and cultured using fresh medium. To expand single-cell cloned strains, the monolayer at 80% confluenty from adhered single cell was digested with 2.5 g/L trypsin containing 0.4 g/L edetic acid and subcultured in 48-well, 24-well, 12-well, 6-well culture plates and 6-cm culture dishes, respectively. During expansion of each single-cell cloned strain, the growth pattern and shape of the cells were observed in detail, and growth curves were drawn according to MTT colorimetry and the population doubling time (PDT) in logarithmic growth phase was calculated according to the formula PDT = (T − T0) lg2/(lgNt − lgN0), and their antigen markers for CD29, CD44, CD71, CD105, CD166, CD11a, CD14, CD34, and CD45 (Beckman Coulter Inc.) were detected using a flow cytometry (Beckman Coulter Inc.) as described previously (Zhang et al., 2008).
Induction of single-cell cloned strains toward neuronal cells
In this experiment, five single-cell cloned strains were obtained from hfBMSCs and called strain A, B, C, D, and E. To investigate the trans-differentiation ability of the single-cell cloned strains toward neuronal cells, the cell suspensions (1 × 104/mL) of hfBMSCs as control, and of each strain A, B, C, D, and E at passage 6 as tests were all prepared using α-MEM + 20% FCS + 0.1 mmol/L β-mercaptoethanol, and added in 24-well culture plates (500 μL/well), each sample with 24 replication wells. All the culture plates were incubated in a cell culture box and cells in test group at 50% confluenty were induced using α-MEM + 0.5% bovine serum albumin (BSA) + 2% B27 (Stemcell Technologies Inc.) + 20 ng/mL epidermal growth factor (EGF; Chemicon, Temecula, CA, USA) + 20 ng/mL basic fibroblast growth factor (bFGF; Invitrogen) + 20 ng/mL nerve growth factor-β (NGF-β; Chemicon) + 0.1 mmol/L β-mercaptoethanol to differentiate toward neuronal cells but cells in control group were cultured always using α-MEM + 20% FCS + 0.1 mmol/L β-mercaptoethanol. During induction, cells were observed and fresh induction medium was changed every other day, and RT-PCR and immunocytochemical analysis were done on the fourth and eighth induction day, respectively.
RT-PCR analysis
Total RNA was extracted with Trizol Reagent (Invitrogen) from induced single-cell cloned strains and uninduced hfBMSCs. Transcriptional expression of Nestin, Pax6, Sox1, β-III Tubulin, neurofilament light chain (NF-L), and neuron-specific enolase (NSE) genes was determined by RT-PCR using the RevertAidTM First-Strand cDNA Synthesis Kit (Fermentas, Hanover, MD, USA) and Ex Taq® (TaKaRa, Shiga, Japan) according to the manufacturer's instructions. Brain tissue from human abortus (age of 10–12 weeks, derived from a local hospital with permission from the patient, hospital, and the Ethics Committee of Northwest A&F University) was used as positive control. The forward and reverse primers for each PCR set were designed to be located by the primer premier 5.0 [Nestin: For: CTCTTGACATACGGTTTCTG, Rev: TTAGCCTATGAGATGGAGC (150 bp); Pax6: For: AACATACCAAGCGTGTCATCAA, Rev: CTTTCTCCAGGGCCTCAATT (337 bp); Sox1: For: GGAATGGGAGGACAGGATTTT, Rev: CGTTTTGGTTCAGCGATTGT (391 bp); β-III Tubulin: For: AAGGTGCGTGAGGAGTATC, Rev: TGCAGTAGGTCTCATCCGT (145 bp); NF-L: For: GTGAAATGGCACGATACC, Rev: GTGAAACTGAGTCGGGTCT (121 bp); NSE: For: GGGCACTCTACCAGGACTTT, Rev: CCTACATTGGCTGTGAACT (106 bp); β-actin: For: TAGTTGCGTTACACCCTTTC, Rev: CCTTCACCGTTCCAGTTT (148 bp)]. The thermal profile for PCR was 95°C for 5 min, followed by 30 cycles of 30 sec at 95°C, and with 1 min annealing intervals (52°C for Nestin, Pax6, Sox1, β-III Tubulin, NF-L, NSE, and β-actin), followed by 1-min extension at 72°C. Additionally, 10-min incubation was included after completion of the last cycle. Following the PCR, 5 μL of the PCR product was electrophoresed on a 1% agarose gel and imaged using a gel imaging instrument (Syngene, Cambridge, UK).
Immunocytochemical analysis
After removal of the induction medium and washing with 0.01 mol/L phosphate-buffered saline (PBS), induced and uninduced cells were fixed with 4% paraformaldehyde in PBS for 30 min and washed triply with PBS. Nonspecific binding sites were blocked by a 1-h incubation in PBS containing 3% normal goat serum (Sigma), 1% BSA (fraction 5; Serva, Heidelberg, Germany), and 1% Triton X-100 (Sigma). The mouse monoclonal primary antibodies against human Nestin (Invitrogen), β-III Tubulin (Sigma), NF-L (ProSci Inc., San Diego, CA, USA) and NSE (Chemicon), the goat polyclonal primary antibodies against human Pax6 (LifeSpan, Bloomington, IN, USA), and the rabbit polyclonal primary antibodies against human Sox1 (Abcam, Cambridge, MA, USA) were each diluted in PBS containing 1% BSA and incubated at 4°C overnight. HistostainTM-Plus Kit and DAB substrates (Beijing Zhongshan Biotechnology Inc., Beijing, China) for peroxidase were used according to the manufacturer's instructions. Cells were observed using an inverted microscope (Leica, Wetzlar, Germany) with 10–40 × objective. Omission of primary antibodies served as negative control and resulted in no detectable staining. Evenly distributed nonoverlapping 10 fields were randomly chosen from each strain and control sample under microscopic and photographed to provide representative images of the stained and nonstained cells. The proportion of cells immunoreactive to particular antigens was quantified by counting 100 cells per field (cell body was only counted).
Results
Growth characteristics of five single-cell cloned strains
In this experiment, five single-cell cloned strains of hfBMSCs, called strains A, B, C, D, and E, were cloned. During the clone, it was observed that the shapes of five adhered single cells initially isolated were different from each other. They presented clubbed, spindle, cyprinoid-like, spindle-like, and triangle-like shape, respectively (Fig. 1a–e). The cells within each strain were consistent in shape during primary culture but gradually showed diversified cellular shapes with increasing passages, such as short spindle shape, long spindle shape, triangle-like shape, and anomalistic shape. After passage 5, the cells of all five strains approached consistent shape (Fig. 1h), similar to hfBMSCs (Fig. 1i). In primary culture, cells of each single-cell cloned strain grew slower, but more rapidly during 2–10 passages, and cell growth speeds were different among five strains (Fig. 2). PDTs of five strains at passage 6 were 36 h (strain A), 37 h (strain B), 45 h (strain C), 49 h (strain D), and 52 h (strain E), respectively.
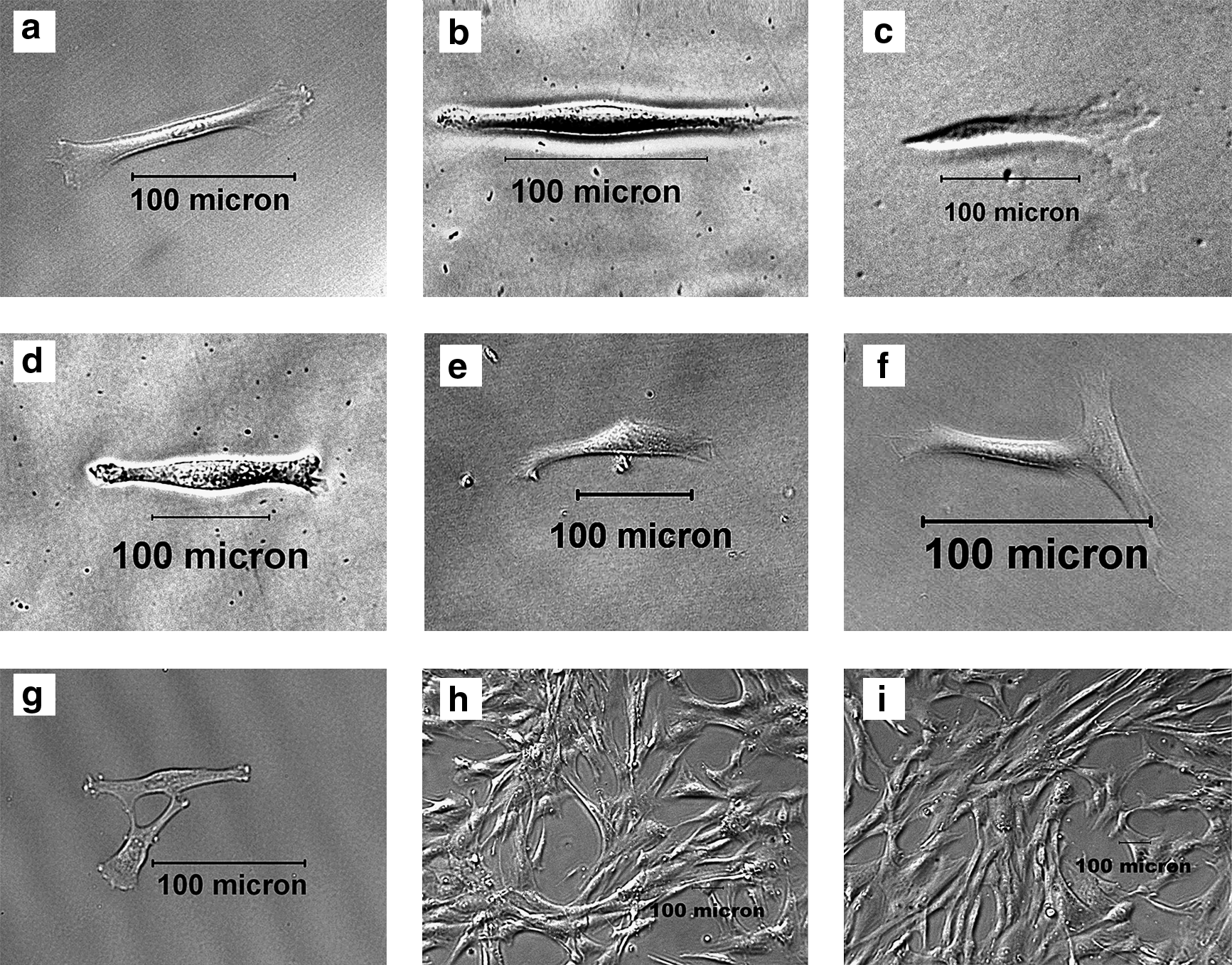
Establishment of single-cell cloned strains from hfBMSCs.
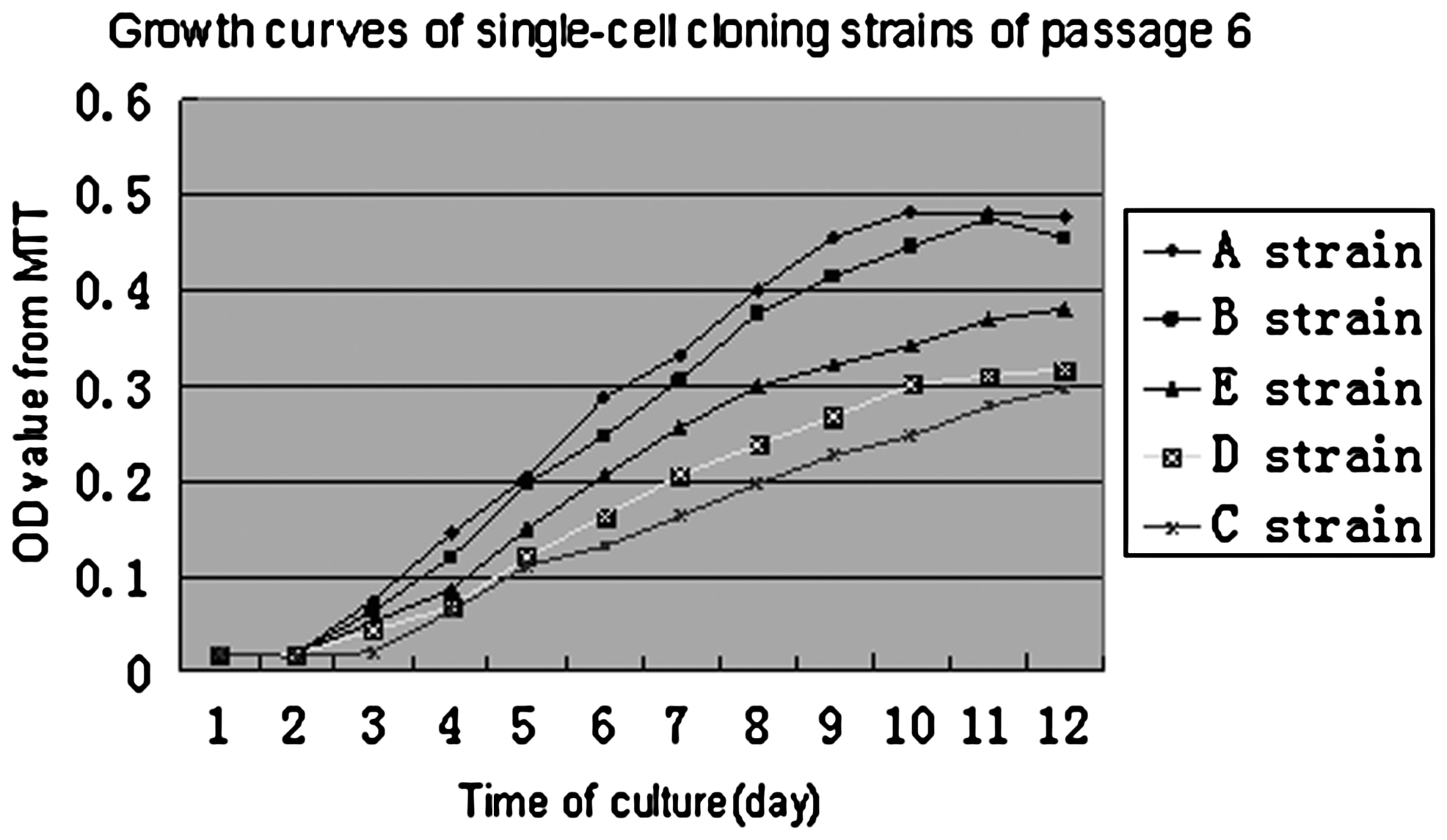
Growth curves of five single-cell cloned strains at passage 6.
Surface antigen markers of single-cell cloned strains
The surface antigen markers of give single-cell cloned strains and hfBMSCs were detected using flow cytometry. Except strain D, CD29, CD44, CD71, CD105, and CD166 of the other strains were positive, CD11a, CD14, CD34, and CD45 of the other strains were negative (Table 1 and Fig. 3a), and they were similar to those of hfBMSCs, according with characteristics of BMSCs. In strain D, CD29, CD44, CD105, and CD166 of the other strains were positive, CD11a and CD14 were negative, but CD71 was negative, CD34 dominantly expressed, and CD45 feebly expressed (Table 1 and Fig. 3b).
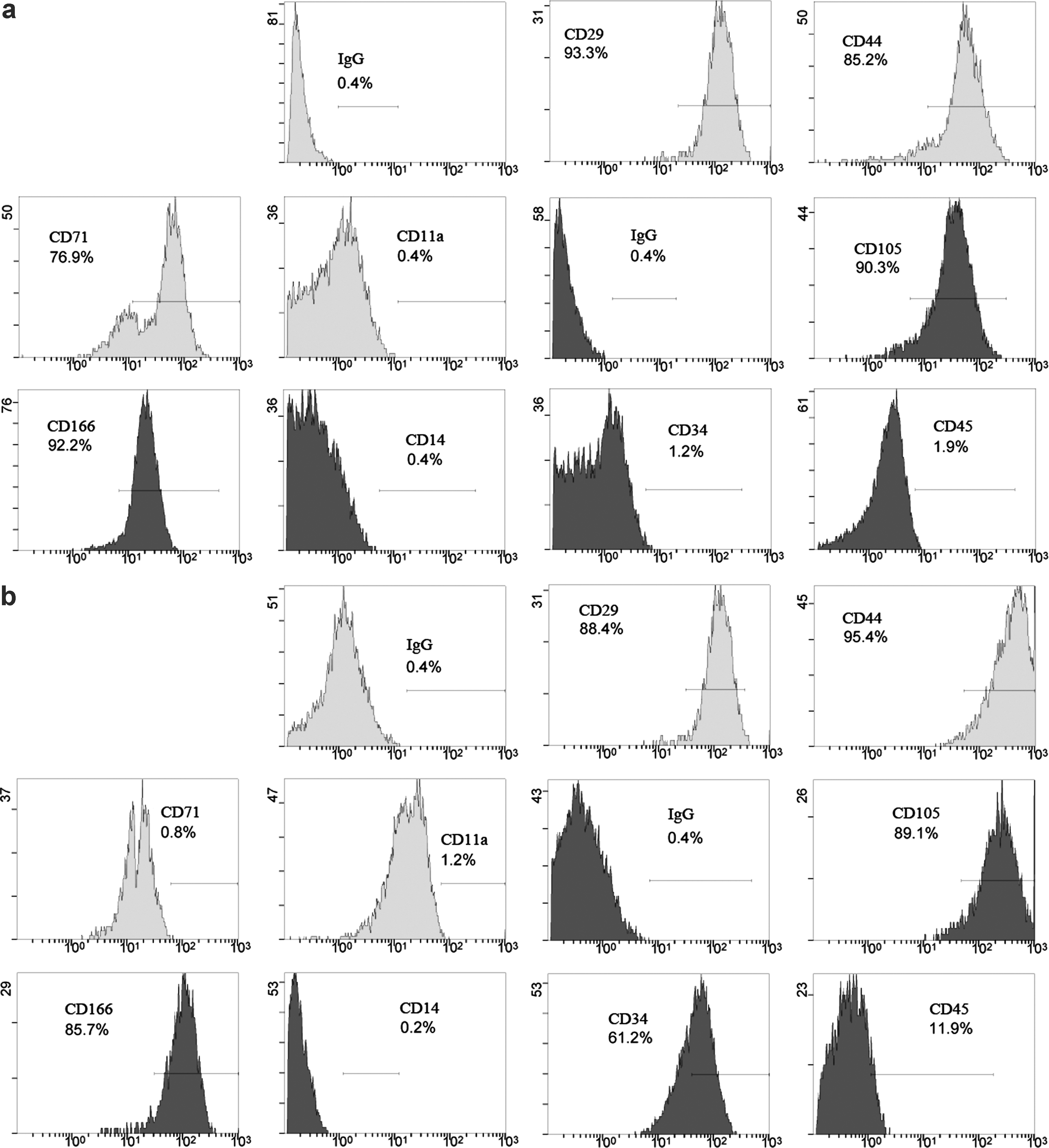
Flow cytometry analysis of surface antigen markers. Strain A cells are positive to CD29, CD44, CD71, CD105, and CD166, but negative to CD11a, CD14, CD34, and CD45
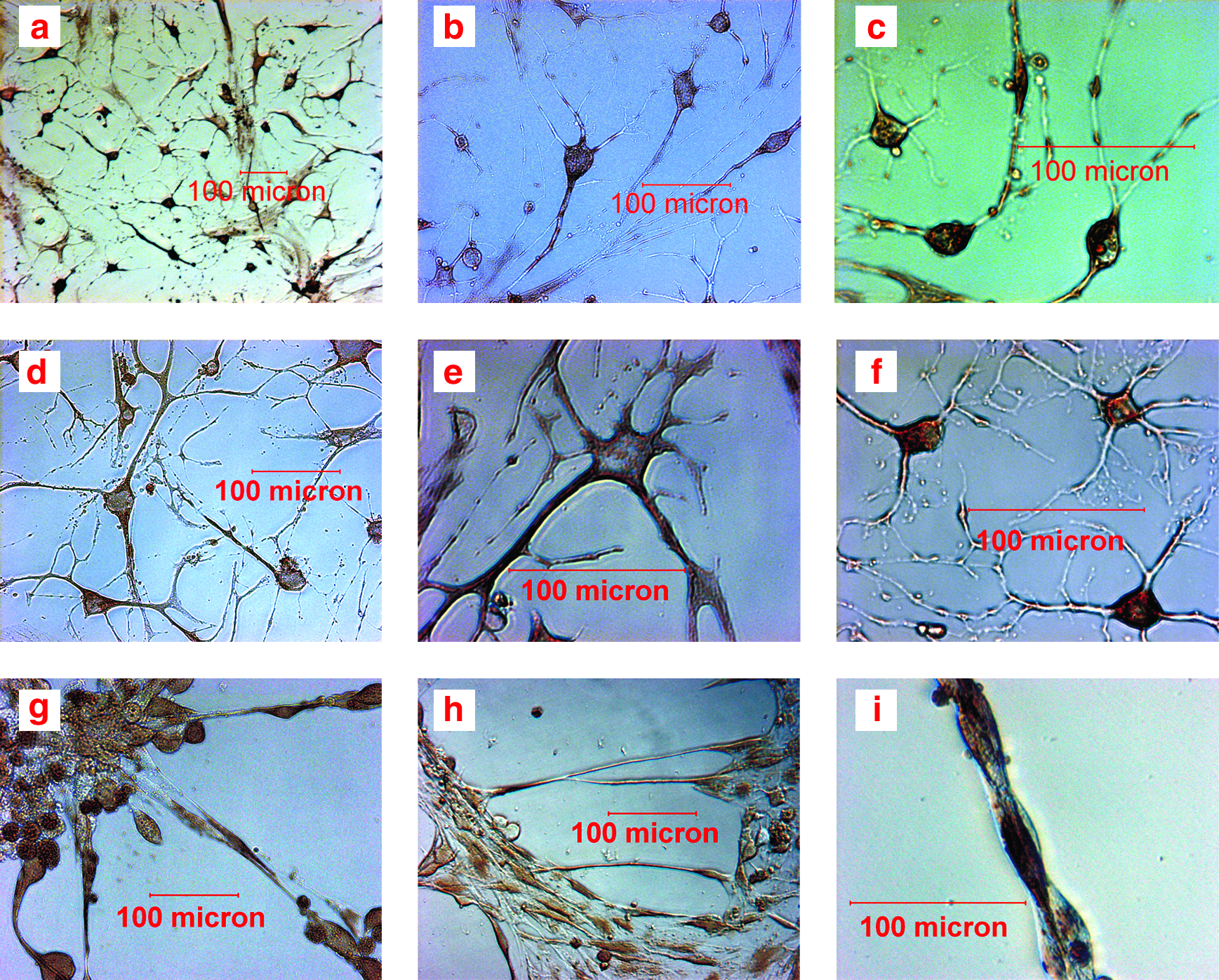
Trans-differentiation of single-cell cloned strains toward neuronal cells
Except strain D, the most cells of the other strains obtained obvious neuron-like phenotype after induction for 4 days. Especially in strains A and B, some cells turned out nerve tissue-like cell clusters after induction for 8 days and there were nerve fiber links among nerve tissue-like cell clusters (Fig. 4). Cellular expression of Nestin, Pax6, Sox1, β-III Tubulin, NF-L, and NSE mRNA after induction for 4 days and 8 days was analyzed with RT-PCR, respectively. Strain D and uninduced hfBMSCs expressed none mRNA of the six genes, whereas the other four single-cell cloned strains and brain tissue derived from abortus expressed mRNA of Nestin, Pax6, Sox1, β-III Tubulin, NF-L, and NSE (Fig. 5). The expression of Nestin, Pax6, and Sox1 mRNA was stronger after induction for 4 days, and the expression of β-III Tubulin, NF-L, and NSE mRNA was stronger after induction for 8 days. Cells after induction for 4 days and 8 days were analyzed with immunocytochemistry. Except strain D, there were Nestin, Pax6, Sox1, β-III Tubulin, NF-L, and NSE-positive cells in all other strains. Like the result of RT-PCR analysis, The expression of nestin, Pax6, and Sox1 was stronger after induction for 4 days and that of β-III Tubulin, NF-L, and NSE was each stronger after induction for 8 days in the other strains. Nerve tissue-like cell clusters formed in strains A and B also expressed Nestin, Pax6, Sox1, β-III Tubulin, NF-L, and NSE (Fig. 6). The percentages of positive cells for Nestin, Pax6, and Sox1 after induction for 4 days, and for β-III Tubulin, NF-L, and NSE after induction for 8 days, are listed in Table 2. Weighted means derived from the percentages of six neuronal antigen markers in Table 2 were compared using one-way analysis of variance (ANOVA); the percentages of neuron-like cells in strain A and B were significantly higher than those in strains C and E (p < 0.01), and that of strain C was significatively higher than that in strain E (p < 0.01). Uninduced hfBMSCs were negative for Nestin, Pax6, Sox1, β-III Tubulin, NF-L, and NSE, and immunocytochemical stain of induced and uninduced cells without primary antibody added was all negative.

Neuronal differentiation of single-cell cloned strains from hfBMSCs.
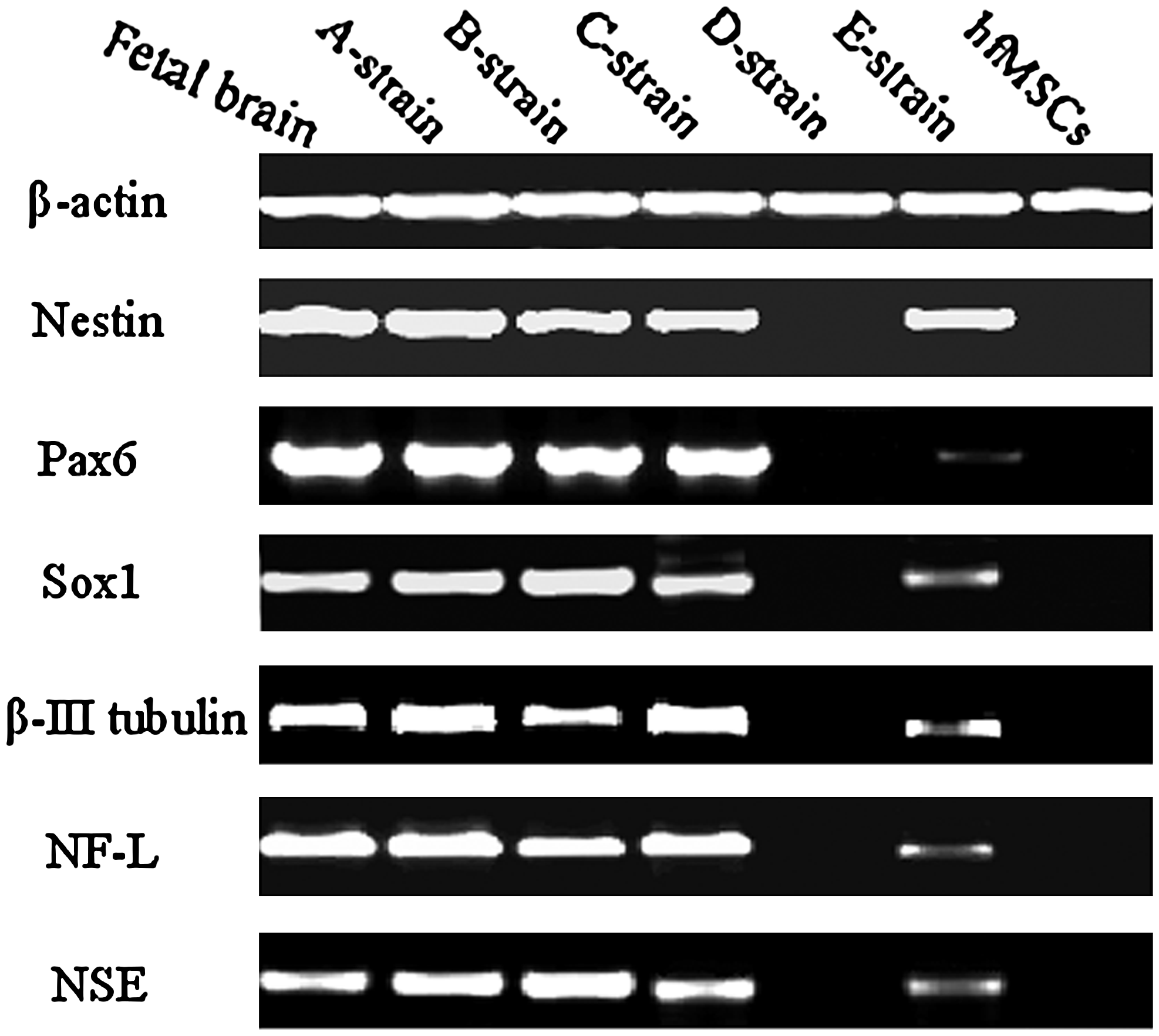
RT-PCR analysis. Human fetal brain tissue and uninduced hfBMSCs were used as the positive and the negative control, respectively, and beta-actin as a reference gene. Except strain D and uninduced hfBMSCs, which expressed none of the six genes, other four strains and fetal brain tissue each expressed all six genes. Nestin, Pax6, and Sox1 were the expression level after 4-day induction, and β-III Tubulin, NF-L, and NSE were the expression level after 8-day induction in the figure.
Immunocytochemical analysis. Strain A cells were positive for Nestin
Note: Same lowercase letter in the means represents no statistical difference (p > 0.05) in between two strains but different lowercase letters represent significant differences (p < 0.01) between two strains. Data were treated with Spss.13 and expressed as means ± SDM.
Discussion
Single-cell clones of hfBMSCs and their biological properties
A single-cell cloned strain of BMSCs plays an important role in research of biological property, especially of the plasticity of BMSCs, because they derive from a single cell without heterogeneity. However, it is very difficult to isolate and expand a single-cell cloned strain from adult BMSCs because of their limited proliferative ability. Tremain et al. (2001) seeded BMSCs in 100-mm dishes at 10 cells/cm2, cultured for 10 to 14 days, and then isolated visible colonies using cloning cylinders and trypsin–EDTA in order to microserial analysis of gene expression. Recently, Boker et al. (2008) extended the life span of adult BMSCs by ectopic expression of hTERT using lentiviral gene transfer and then selected a single-cell clone. In this study, five single-cells were isolated from hfBMSCs and expanded to five single-cell cloned strains, indicating that BMSCs derived from bone marrow of human first-trimester abortus have strong proliferation ability.
In this study, it was observed that the shape of the five single cells initially isolated was different each other, and cell shape in each strain was correspondingly consistent during primary culture and gradually become diversification with increasing passage, such as short spindle shape, long spindle shape, triangle-like shape, and anomalistic shape. After passage 5, cell shape of the five strains approached consistent, similar to hfBMSCs (Fig. 1). Cell growth rate (Fig. 2) differed among the five strains. Although surface antigen marker profile was similar among strains A, B, C, E, and control hfBMSCs, the expression rate of CD71 was much lower and those of CD34 and CD45 relatively higher in strain D (Table 1). All these indicated that biological properties of each cell in hfBMSCs were different and gradually changed during proliferation in vitro.
Trans-differentiation of single-cell cloned strains toward neuronal cells
Initially, Woodbury et al. (2000) rapidly induced BMSCs to differentiate into neurons using simple chemical agents. But this procedure was soon questioned by other authors, suggesting that chemical neuronal induction results in cellular stress, leading to a physical contraction of cells into a neuron-like morphology, and demonstrating that if simple chemical agents in induction medium are removed, the cells quickly reverse to typical BMSCs morphology, losing the expression of neuronal markers (Liu and Rao, 2003; Lu et al., 2004). Later experiments demonstrated that soluble neurotrophic factors can induce BMSCs to achieve permanent trans-differentiation (Vaquero and Zurita, 2009; Zurita et al., 2007). EGF and bFGF can promote the proliferation and differentiation of neural stem cells (Santa-Olalla and Covarrubias, 1999; Ostenfeld and Clive, 2004) and increase the multipotentiality of adipose-derived MSCs and BMSCs (Rider et al., 2008; Tamama et al., 2006). Therefore, in this study, induction medium containing neurotrophic factor NGF-β and cell growth factor EGF and bFGF was used to induce five single-cell cloned strains from hfBMSCs to differentiate toward neuronal cells. Except strain D, the most cells of the other strains obtained neuron-like phenotype after 4-day induction, especially in strains A and B; the cells not only had morphologically become to neuron-like cells, but also formed nerve tissue-like cell clusters after 8-day induction (Fig. 4), and they expressed Nestin, Pax6, Sox1, β-III Tubulin, NF-L, and NSE genes through RT-PCR and immunocytochemical analysis (Fig. 5 and Fig. 6), showing prominent neural differentiation ability. Their trans-differentiation rates were 80.5 ± 5.08% (strain A), 78.1 ± 5.59% (strain B), 68.1 ± 6.58% (strain C), 0 (strain D), and 49.8 ± 7.72% (strain E) in turn (Table 2). These results indicate the induction medium containing NGF-β, EGF, and bFGF was very effective and hfBMSCs possessed very strong plasticity into neuronal cells but the plasticity of each cell in them, even in descendants of a cell, was different. In addition, the plasticity of the single-cell cloned strains into neuronal cells showed a coincident tendency with the growth speeds and PDTs of those cells before being induced except strain D. Strain D, however, did not obtain neuron-like shape and did not express Nestin, Pax6, Sox1, β-III Tubulin, NF-L, and NSE genes after induction (Fig. 5). It needs to further research that whether no differentiation of strain D toward neuronal cells is relative to their nonexpression for CD71, dominant expression for CD34, and feeble expression for CD45.
We are not advocating carrying out abortions for the single purpose of obtaining human fetal BMSCs, but in some countries with “family planning policy, one-child policy or birthcontrol policy,” a human abortus, first-trimester abortus in particular, can be used as a source of obtaining a variety of fetal stem cells, especially BMSCs. Certainly, such an approach may be suitable for certain societies, but may be controversial in some others.
Footnotes
Acknowledgments
W acknowledge the grants from the Key Program of National Ministry of Education(03160), the National Natural Science Foundation of China(30671067) and the Key Program of Shaanxi Province(2006Kz05-G1). In addition, we acknowledge Doctor Xingrong Yan and Yun Wang, Master Ting Zhao and Hai Qiao in our lab for help during experiments.
Author Disclosure Statement
The authors declare that no conflicting financial interests exist.