
Abstract
Abstract
UV-irradiation of oocytes during enucleation and serum starvation of donor cells during cell cycle synchronization may compromise the development competence of cloned embryos through excessive generation of reactive oxygen species (ROS). Here, we show that 3-hyroxyflavone (a flavonoid having hydroxyl group at 3 carbon position) inhibits UV- and serum starvation-induced ROS production in oocytes and donor cells, respectively, and thereby improves the in vitro development of cloned porcine embryos (p<0.05). In a parthenogenetic model, UV-irradiation for 5 sec or more was found to reduce the in vitro development and quality of the embryo, which could be rescued by their culture in the presence of 3-hydroxyflavone. The rescuing effect of 3-hydroxyflavone was associated with significant reduction in ROS level (14.4±1.0 vs. 47.1±6.7), increase in ERK signaling molecules by 2.1-fold, and decrease in Caspase3 expression by 3.2-fold. Culture of donor cells (18.5±1.4 vs. 13.0±1.7%) or cloned embryos (20.6±1.1 vs. 12.2±1.1%) in the presence of 3-hydroxflavone also increased (p<0.05) the rates of blastocyst formation in cloned embryos produced by the nuclear transfer of serum-starved donor cells into recipient cytoplasts exposed to UV-irradiation during the enucleation step. Importantly, both parthenotes and cloned embryos cultured in the presence of 3-hydroxyflavone had significantly increased ability to expand, and contained a higher number of cells than those of the control group (p<0.05). These results suggest that 3-hydroxyflavone may be useful for improving the in vitro developmental potential of cloned embryos through inhibition of ROS production induced by the UV-irradiation of oocyte and/or the serum starvation of donor cells.
Introduction
UV irradiation in combination with the bisbenzimide stain, Hoechst 33342, is routinely used in the SCNT procedure to visualize the chromosomes during enucleation of oocytes to be used as recipient cytoplast. Although UV-induced nuclear damages to oocytes are of less concern in the SCNT procedure, exposure of oocytes to UV irradiation can affect oolemma integrity, damage mitochondrial DNA and cytoplasmic organelles, alter protein synthesis pattern, and interfere with the maturation-promoting factor (MPF) kinase activity, all of which can compromise the viability of recipient cytoplasts, and thereby can potentially reduce their ability to support early embryonic development (Bradshaw et al., 1995; Dominko et al., 2000; Leal et al., 1999; Smith 1993; Villa et al., 1992; Yang et al., 1990). UV irradiation can also result in massive accumulation of reactive oxygen species (ROS) as a UV response, which is initiated at or near the plasma membrane (Devary et al., 1992; Misawa and Takahashi 2011; Yun et al., 2011). The ROS are highly reactive with plasma membrane, intracellular proteins, lipids, nucleic acids, and cell signaling molecules to cause their dysfunction and can result in aberrant cell signaling, abnormal gene transcription, cell cycle arrest, DNA fragmentation, mitochondrial abnormalities, cellular senescence, apoptosis, and cell death in both somatic cells (Devary et al., 1992; Kang et al., 2011; Terada et al., 1995; Zhang et al., 1997) and embryos (Dennery 2007; Favetta et al., 2007; Guerin et al., 2001; Gupta et al., 2010; Uhm et al., 2011; Wells et al., 2009). Increase in ROS level is also seen during serum starvation of somatic cells—a common procedure for the cell cycle synchronization of donor cells to G0/G1 stage (Chen et al., 2009; Kuznetsov et al., 2011; Lee and Piedrahita 2002). Thus, serum starvation of donor cells may induce massive DNA fragmentation, apoptosis, or cell death, and therefore, may also contribute to the low efficiency of SCNT (Boquest et al., 1999; Kues et al., 2000, 2002; Lee and Piedrahita 2002).
Flavonoids are naturally occurring plant pigments enriched in vegetables and soy-containing foods and are widely consumed by humans for putative health benefits (Bisht et al., 2010; Havsteen, 2002). We recently showed that 3-hydroxyflavone (a flavonoid having hydroxyl group at 3-carbon position) significantly improves the in vitro development and quality of porcine embryos when used at an optimal concentration of 10 μM (Uhm et al., 2011). Furthermore, culture of embryos in the presence of 3-hydroxyflavone rescued the poor development competence of in vitro aged oocytes by inhibiting the ROS level and modulating the expression of proteins related to extracellular signal-regulated kinase (ERK) signaling pathway (Uhm et al., 2011). The 3-hydroxyflavone was also found to consistently inhibit ROS production in various somatic and cancer cell lines (Kim et al., 2008; Lee et al., 2008).
In this study, therefore, we extended our earlier study (Uhm et al., 2011) to investigate the effect of 3-hydroxyflavone on cloned porcine embryos produced by SCNT of serum-starved donor cells into recipient cytoplasts exposed to UV-irradiation during enucleation.
Materials and Methods
All chemicals were obtained from Sigma-Aldrich Co. (St. Louis, MO) unless specifically indicated. At least four replicates were performed for each experiment, and in each of the replication, oocytes retrieved from same batch of abattoir-derived ovaries were randomly allotted to different groups.
Oocyte retrieval and in vitro maturation (IVM)
Retrieval of porcine oocytes from abattoir-derived ovaries and their IVM was performed essentially as we described earlier (Gupta et al., 2007b). Briefly, cumulus–oocyte complexes were aspirated from medium-sized follicles (3–6 mm diameter) and matured in groups of 50 in 500 μL of Tissue Culture Medium 199 with Earle's salts (TCM-199; Gibco BRL, Grand Island, NY) supplemented with 25 mM NaHCO3, 10% (v/v) porcine follicular fluid, 0.57 mM cysteine, 0.22 μg/mL sodium pyruvate, 25 μg/mL gentamicin sulfate, 0.5 μg/mL FSH (Folltropin V; Vetrepharm, Canada), 1 μg/mL estradiol-17β, and 10 ng/mL epidermal growth factor under mineral oil at 39°C in a humidified atmosphere of 5% CO2 in air for 40–42 h.
SCNT
Cloned embryos were produced by SCNT as we described earlier (Gupta et al., 2008) with partial modifications. Briefly, in vitro matured oocytes were denuded of cumulus cells in TL-HEPES medium supplemented with 0.1% hyaluronidase and incubated with cytochalasin B (7.5 μg/mL) and Hoechst 33342 (10 μg/mL) for 15 min. The oocytes were then enucleated, using a beveled borosilicate pipette (25 μm internal diameter), by direct visualization of fluorescent metaphase plate under UV-illumination provided through×40 objective (LUCPlanFl, Olympus, Japan) of an inverted epifluorescence microscope (Olympus IX71, Japan) fitted with blue filter cassette (excitation filter: 330–385 nm; emission filter: 420 nm; dichromatic beam splitter: 400 nm) and connected to a 100 W mercury lamp (USH-102D, Ushio Inc, Tokyo, Japan). The total duration of UV exposure to the oocyte was kept to a minimum (∼5 sec/oocyte). It was not possible to accurately measure the precise dose of UV to the oocytes. However, the settings used for the UV-illumination was same as that generally used for UV-assisted physical enucleation in most SCNT protocol (Das et al., 2010b; Gupta et al., 2008; Li et al., 2004; Uhm et al., 2007a). Enucleated oocytes were subsequently reconstructed by inserting a small sized (∼15 μm in diameter), smooth bordered porcine fetal fibroblast (pFF) cell into the perivitelline space of each enucleated oocyte using the same pipette used for the enucleation. The donor cells for the SCNT were prepared and cultured in Dulbecco's modified Eagle's medium (DMEM, high glucose formulation; Gibco BRL) supplemented with 10% (v/v) fetal bovine serum (FBS; Hyclone, Logan, UT), 1% (v/v) MEM nonessential amino acids (Gibco BRL), and 1% penicillin–streptomycin (Gibco BRL) at 39°C in a humidified atmosphere of 5% CO2 in air essentially the same as we described earlier (Uhm et al., 2007a). Cells were routinely maintained on 50 mL tissue culture flasks (Falcon BD, Lincoln Park, NJ) up to two to seven passages and used as donor cells for nuclear transfer after synchronizing their cell cycle stage to G0/G1 either by confluence culture or by serum starvation (Boquest et al., 1999). In experiments that evaluated the effect 3-hydroxyflavone on serum-starved donor cells, the culture medium was further supplemented with or without 3-hyrodyflavone (10 μM). Membrane fusion of donor cell with recipient cytoplast was induced with single DC pulse of 2.1 kV/cm for 30 μsec delivered by a BTX Electro Cell Manipulator 2001 (BTX, San Diego, CA).
Parthenogenetic activation (PA) of oocytes
Presumptive diploid parthenotes were produced by electroactivation essentially as we described earlier (Gupta et al., 2007a). Briefly, in vitro matured oocytes were denuded of cumulus cells in TL-HEPES medium supplemented with 0.1% hyaluronidase and were subsequently electroactivated in activation medium (0.3 M mannitol, 0.1 mM MgSO4, and 1.0 mM CaCl2) by a single DC pulse of 1.0 kV/cm for 30 μsec delivered by a BTX Electro Cell Manipulator 2001 (BTX). Activated oocytes were then cultured in North Carolina State University 23 (NCSU23) medium supplemented with 7.5 μg/mL cytochalasin B for 4 h. In experiments that evaluated the effect of UV irradiation on porcine parthenotes, activated oocytes were exposed to different duration (0, 5, 10, 20, 30, 60 sec) of UV irradiation essentially as described above.
In vitro culture (IVC) of embryos
Reconstructed and activated embryos were cultured in groups of 12–15 embryos per 50 μL microdroplets of NCSU23 medium supplemented with 0.4% essentially fatty acid free bovine serum albumin under mineral oil at 39°C in a humidified atmosphere of 5% CO2 in air for 7 days essentially as we described earlier (Uhm et al., 2007b). The IVC medium was further supplemented with or without 3-hydroxyflavone (10 μM) to evaluate its effect on the embryos. The concentration of 3-hydroxyflavone was chosen based on our previous study (Uhm et al., 2011). The rates of cleavage (percentage of embryos that cleaved) and blastocyst formation and expansion were recorded on day 2 and day 7 of IVC, respectively. Total nuclei number of blastocysts were counted on day 7 of IVC by fluorescent Hoechst 33342 staining essentially as we described earlier (Gupta et al., 2009).
Measurement of intracellular ROS level
Intracellular ROS level in embryos was measured by 2′,7′-dichlorofluorescene (DCF) fluorescence assay essentially as we described earlier (Gupta et al., 2010; Nasr-Esfahani et al., 1990). Briefly, two- to four-cell stage embryos were incubated with 10 mM 2′7′-dichlorodihydrofluorescein diacetate (DCHFDA) for 20 min at 39°C, washed three times in NCSU23 medium to remove the traces of dye, and immediately analyzed for intracellular fluorescent DCF under epifluorescent microscope (Olympus, Tokyo, Japan) using excitation of 450–490 nm and emission of 515–565 nm. A digital camera (Nikon, Tokyo, Japan) attached to the microscope acquired the images and mean gray value of fluorescent oocytes were measured using ImageJ software (NIH, Bethesda, MD). Background fluorescent values were subtracted from the final values before analyzing for the statistical difference among the groups. The experiment was replicated three times with 20–25 oocytes in each replicate.
Western blot analysis
Western blot analysis was performed essentially as we described earlier (Uhm et al., 2011). Briefly, total proteins were extracted from two- to four-cell stage embryos (50 per sample) in extraction buffer (1% Triton X-100, 100 mM Tris-HCl, pH 7.5, 10 mM NaCl, 10% glycerol, 1 mM sodium orthovanadate, 50 mM sodium fluoride, 1 mM p-nitrophenyl phosphate, 1 mM phenylmethylsulfonyl fluoride), separated on 10% SDS-PAGE, transferred electrophoretically onto enhanced chemiluminescence (ECL) Hybond nitrocellulose membranes (Amersham, Piscataway, NJ) and subsequently probed with primary antibody against ERK1/2 and Caspase 3 (Abcam Inc., Cambridge, MA) in Tris-buffered saline, which contained 5% nonfat dry milk and 0.1% Tween 20. The antibody–antigen complexes were detected using antimouse IgG or antirabbit IgG peroxidase conjugates, followed by the use of an ECL detection kit (Amersham Bioscience, Piscataway, NJ). Experiments were done in triplicates. Intensity of the bands on the blots was measured by a densitometric analysis system (Bio-Rad, Hercules, CA). Intensity of β-actin bands were used for normalization.
Cell cycle analysis
Distribution of cells into different stages of cell cycle was analyzed by flow cytometry essentially as we described earlier (Jung et al., 2010). Briefly, control, confluence-cultured and serum-starved cells were trypsinized into single cell suspension and fixed in 70% ice-cold ethanol. The fixed cells were then incubated with staining solution containing 50 μg/mL propidium iodide, 100 50 μg/mL ribonuclease A, and 1% Triton X-100 for 20 min at room temperature prior to flow cytometry (FacsCalibur, Becton Dickinson, Mountain View, CA). Ten thousand events were recorded for each sample and all analyzed events were gated to remove ebris and aggregates. Cell Quest software (Becton Dickinson, Bedford, MA) was used for the data analysis.
Statistical analyses
Statistical analyses were performed using SPSS software (SPSS Inc, Chicago, IL) for chi-square test or analysis of variance (ANOVA) as appropriate. Percentage data were subjected to arc sine transformation before statistical analysis. Data are presented as mean±SEM. Differences at p<0.05 were considered significant.
Results
Effect of 3-hydroflavone on UV-irradiated oocytes
In the first set of experiments we evaluated the effect of UV irradiation on in vitro development of parthenogenetically activated porcine oocytes. The parthenotes were chosen for the study because they are relatively easier to produce in large numbers for the analysis, and their in vitro development characteristics are similar to those of SCNT embryos (Das et al., 2010a; Gupta et al., 2007c). Presumptive diploid parthenotes were exposed to the UV irradiation for different duration (0, 5, 10, 20, 30, or 60 sec) and were subsequently cultured in vitro for 7 days. Results showed that, UV irradiation for 10 sec or more significantly reduced (p<0.05) the rates of cleavage and blastocyst formation in a time-dependent manner (Table 1). The blastocysts generated from UV-irradiated parthenotes also had lower ability to expand and contained significantly reduced number of cells than those of control (p<0.05).
Experiments were replicated six times.
Values in parenthesis indicate the number of embryos.
Values with different superscripts (a–e) within column denote a statistical difference (p<0.05).
We next evaluated the ability of 3-hydroxyflavone to rescue the in vitro development of UV-irradiated parthenotes. The diploid parthenotes were irradiated for either 10 or 30 sec and were subsequently cultured in the absence (control) or presence of 3-hydroxyflavone (10 μM) for 7 days. Results showed that culture in the presence of 3-hydroxyflavone improved (p<0.05) the rates of blastocyst formation and expansion in porcine parthenotes that were UV irradiated for either 10 or 30 sec, respectively (Table 2). The 3-hydroxyflavone-treated blastocysts also contained a higher (p<0.05) number of cells than those of control (Table 2). However, the presence of 3-hydroxyflavone in the IVC medium did not affect the rate of cleavage (p>0.05). Nonetheless, two- to four-cell stage embryos in the 3-hyroxyflavone group had significantly lower level of ROS (14.4±1.0 vs. 47.1±6.7) than those of control group (Fig. 1). Furthermore, Western blotting revealed that two- to four-4 cell stage embryos in the 3-hyroxyflavone group had 2.1-fold higher and 3.2-fold lower expressions of ERK1/2 and Caspase 3 proteins, respectively, than that of control group (Fig. 2).

ROS level in two- to four-cell stage porcine parthenotes exposed to UV-irradiation for 30 sec and cultured in the absence (control) or presence of 3-hydroxyflavone (10 μM). Left panel: brightfield; right panel: green fluorescence indicating ROS level. Numerical values on figures indicate the average fluorescence intensity (mean±SEM). Values with different superscript (a, b) denote statistical difference (p<0.05).
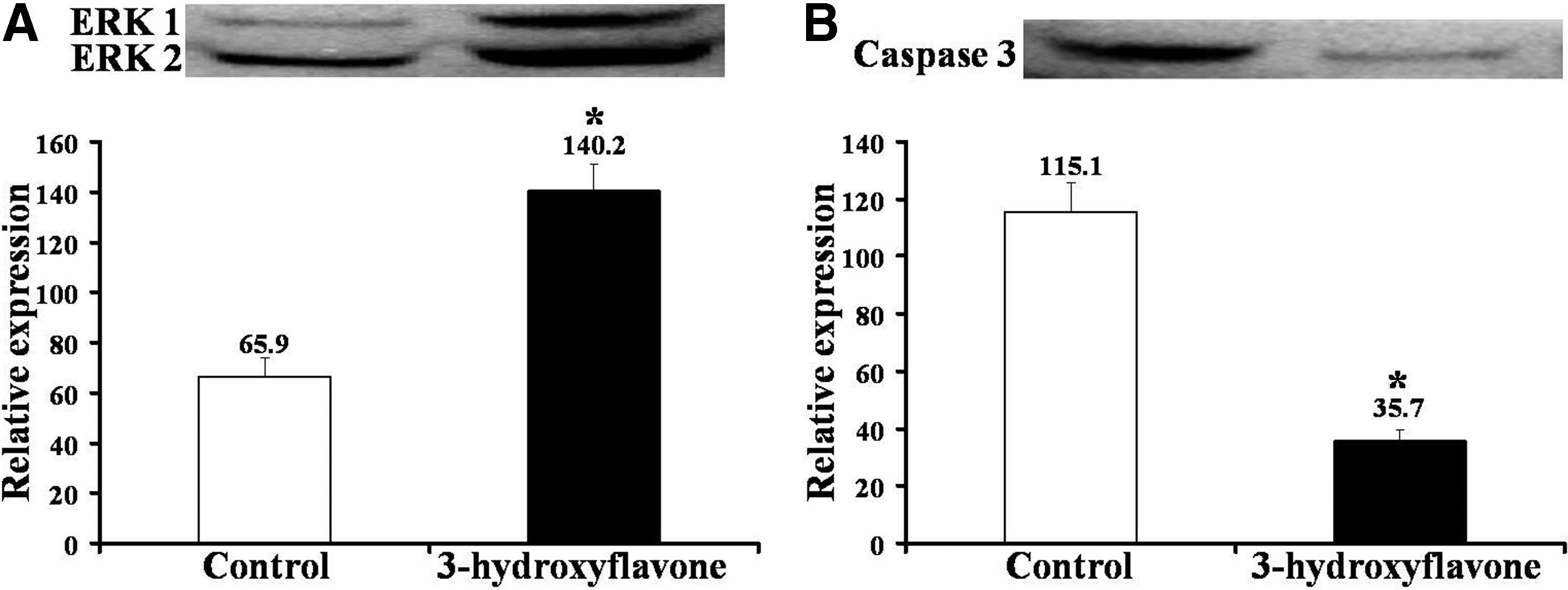
Effect of 3-hydroxyflavone on the expressions of ERK 1/2 (
Experiments were replicated four times.
Values in parenthesis indicate the number of embryos.
Values with different superscripts (a, b) within column denote a statistical difference within each group (p<0.05).
Finally, we evaluated the effect of 3-hydroxyflaone on in vitro development and quality of cloned embryos produced by SCNT of confluence cultured donor cells into recipient cytoplasts that were exposed to UV irradiation during the enucleation step. Results showed that, similar to PA embryos, culture of cloned embryos in the presence of 3-hydroxyflavone significantly improved the rates of blastocyst formation and expansion and the total cell number per blastocyst but did not affect the rate of cleavage (Table 3).
Cloned embryos were produced by SCNT of donor cells that were confluence cultured to synchronize their cell cycle to G0/G1 phase.
Experiments were replicated five times.
Values in parenthesis indicate the number of embryos.
Values with different superscripts (a, b) within column denote a statistical difference (p<0.05).
Effect of 3-hydroflavone on serum starved donor cells
In this set of experiments, we evaluated the effect of 3-hydroxyflavone on serum starvation-induced ROS production and subsequent embryonic development after SCNT. The donor cells were serum starved for 24 h in the absence (control) or presence of 3-hydroxyflavone and were subsequently used for SCNT into recipient cytoplasts that were exposed to UV irradiation during the enucleation step. As expected, serum starvation resulted in better cell cycle synchronization than confluence cultured or control cells (Table 4) but increased the ROS level (Fig. 3). Culture of serum-starved somatic cells in the presence of 3-hydroxyflavone inhibited the accumulation of ROS (Fig. 4). Furthermore, SCNT of these serum-starved, 3-hydroxyflavone-treated donor cells resulted in improved (p<0.05) rates of blastocyst formation and expansion than that of nontreated serum-starved cells (Table 5). The blastocyst generated from 3-hydroxyflavone-treated donor cells also contained a significantly higher number of cells than those generated from nontreated donor cells.

Effect of serum starvation on ROS production in porcine fetal fibroblasts. Top panel: green fluorescence indicating ROS level; bottom panel: bright field.
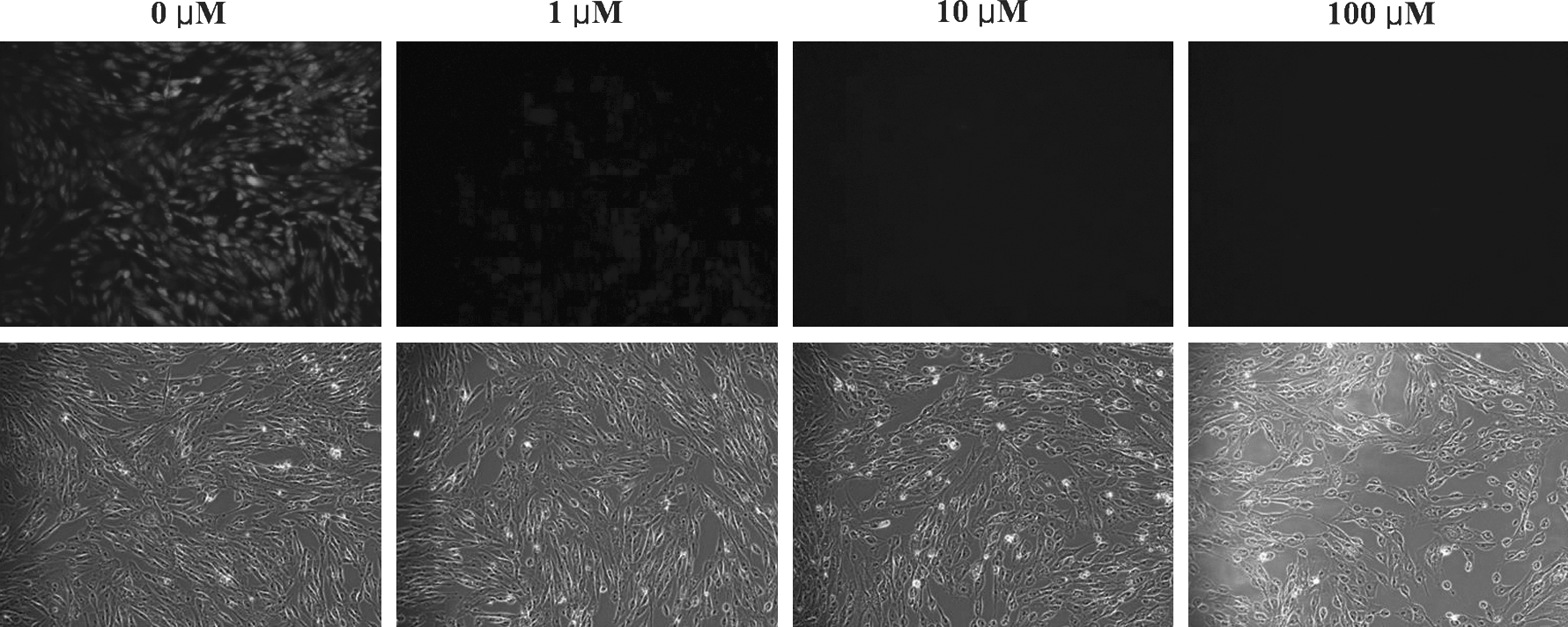
ROS level in serum-starved porcine fetal fibroblasts cultured in the absence (control) or presence of 3-hydroxyflavone (1, 10, 100 μM) for 24 h. Top panel: green fluorescence indicating ROS level; bottom panel: brightfield.
Values with different superscripts (a, b) within column denote a statistical difference (p<0.05).
Cloned embryos were produced by SCNT of donor cells that were serum-starved for 24 h to synchronize their cell cycle to G0/G1 phage.
Experiments were replicated five times.
Values in parenthesis indicate the number of embryos.
Values with different superscripts (a, b) within column denote a statistical difference (p<0.05).
Discussion
ROS are generated constitutively in mammalian cells and is thought to be an important second messenger in cell signaling. However, its excessive generation in response to external insults such as UV irradiation and serum starvation can result in extensive cellular damage (Hachiya and Akashi, 2005; Kang et al., 2011; Misawa and Takahashi, 2011; Yun et al., 2011) including developmental block during early embryonic development of mammalian embryos (Dennery, 2007; Favetta et al., 2007; Nasr-Esfahani et al., 1990; Wells et al., 2009). Here, we extended our previous study (Uhm et al., 2011) to show that 3-hydroxyflavone inhibits UV- and serum starvation-induced ROS level in oocytes and donor cells, respectively, and thereby improves the in vitro development of cloned porcine embryos.
Using porcine parthenotes as a model system, we first observed that UV irradiation for 10 sec or more was detrimental to early embryonic development in vitro. Even a shorter exposure of 5 sec inhibited the progression of the first cell cycle and reduced the rate of cleavage. Nonetheless, UV-irradiated parthenotes could develop to the blastocyst stage. Thus, in addition to DNA damages, UV irradiation possibly also induces cytoplasmic damages. These results are similar to previous studies in which UV irradiation was shown to cause cell cycle arrest in both somatic cells (Devary et al., 1992; Terada et al., 1995; Yun et al., 2011) and oocytes (Bradshaw et al., 1995; Velilla et al., 2002). Focused irradiation of both short wave UV-C (254 nm) and long wave UV-A (>330 nm) to germinal vesicle (GV) or metaphage II (MII) stage bovine oocytes interfered with MPF kinase activity, protein synthesis pattern, cell cycle progression and postfertilization pronuclear formation (Bradshaw et al., 1995). Exposure of bovine oocytes to long-wave UV-A, which is commonly used during enucleation step of SCNT, also caused the loss of oolemma integrity, altered uptake of methionine, marked difference in protein synthesis pattern, increased mitochondrial membrane potential, and damages to cytoplasmic organelles (Smith, 1993). UV irradiation also induced the activation of Src tyrosine kinases, followed by activation of Ras and Raf-1 signaling cascade that resulted in increased phosphorylation of c-Jun and activation of several transcription factors in somatic cells (Devary et al., 1992). On the contrary, short-wave (254 nm) UV irradiation induced a wide range of chromatin defects, including condensation and dispersal of DNA fragments, formation of micronuclei in porcine oocytes (Leal et al., 1999). Thus, effects of UV irradiation vary with their wave length (Dhanwada et al., 1995; Dominko et al., 2000) with long-wave UV irradiation causing alterations to both membrane and intracellular components. In our study, we used long-wave UV irradiation that is normally used during enucleation step of SCNT procedure, and thus, our data suggest that UV irradiation of oocytes for longer than 5 sec may affect their subsequent in vitro development after SCNT. Furthermore, UV-irradiated parthenotes that developed to blastocyst stage had lower rate of cellular proliferation and contained significantly less number of cells than that of control. Thus, despite successful early cleavages, UV irradiation may have long-term effect on embryos that may be evident only at a later stage of development, and hence, care must be taken when using this methodology to visualize the metaphase chromosomes during the enucleation step of SCNT (Dominko et al., 2000; Li et al., 2004; Smith, 1993).
The lowered in vitro development of UV-irradiated parthenotes could, however, be rescued by their culture in the presence of 3-hydroxyflavone. This rescuing effect of 3-hydroxyflavone was associated with their ability to reduce the ROS level in UV-irradiated parthenotes. The blastocysts generated from UV-irradiated parthenotes cultured in the presence of 3-hydroxyflavone also had increased ability to expand and contained higher number of cells than those of the control group. The improvement in the cell number of 3-hydroxyflavone-treated embryos may be due to the increased cellular proliferation and/or reduced apoptosis, as they had increased expressions of ERK 1 and ERK 2 proteins and decreased expression of Caspase 3 protein, which are involved in signaling pathways controlling the cellular proliferation and apoptosis, respectively (Johnson and Lapadat, 2002; Uhm et al., 2007b). These data are similar to previous studies that showed modulation of ROS level and ERK signaling pathway by 3-hydroxyflavone in somatic cells (Kim et al., 2008; Lee et al., 2008) and in vitro aged oocytes (Uhm et al., 2011). Thus, 3-hydroxyflavone possibly rescued the in vitro development of UV-irradiated parthenotes by reducing the ROS production and activating the ERK signaling pathway. Importantly, however, reduced in vitro development of UV-irradiated parthenotes could not be completely rescued by 3-hydroxyflavone when the duration of UV-irradiation was increased to 30 sec. Thus, in addition to increased ROS production, UV irradiation also induces other cellular damages that cannot be rescued by 3-hydroxyflavone (Bradshaw et al., 1995; Dominko et al., 2000; Leal et al., 1999; Smith, 1993; Villa et al., 1992; Yang et al., 1990). Nonetheless, when the duration of UV irradiation during the enucleation step of SCNT procedure was kept to a minimum (∼5 sec/oocyte), culture of resultant cloned embryos in the presence of 3-hydroxyflavone resulted in significantly improved rates of blastocyst formation. These blastocysts were also of improved quality as they had significantly increased ability to expand and contained significantly higher number of cells than those of controls.
Previous studies have also demonstrated that serum starvation during the culture of mammalian cells can induce cell death, especially through apoptosis, and therefore, may contribute to the low efficiency of nuclear transplantation (Kues et al., 2002; Lee and Piedrahita, 2002). Consequently, culture of donor cells in the presence of the apoptosis inhibitor was suggested to improve the cloning efficiency (Lee and Piedrahita, 2002). However, antioxidants were found to give better result than apoptosis inhibitors in reducing apoptosis in serum-starved cells (Lee and Piedrahita, 2002), possibly because ROS are involved in the initiation of apoptosis of serum-starved cells (Kang et al., 2011; Terada et al., 1995; Zhang et al., 1997). Indeed, we observed that serum starvation of donor cells resulted in massive accumulation of ROS that could be inhibited by their culture in the presence of 3-hydroxyflavone. SCNT of these 3-hyroxyflavone-treated, serum-starved donor cells also resulted in improved rate of in vitro development compared to nontreated control. The blastocysts generated from 3-hyroxyflavone-treated donor cells also had higher ability expand and contained higher number of cells than nontreated controls. Thus, the increased development competence of serum-starved cells cultured in the presence of 3-hydroxyflavone is likely due to decrease in ROS level, and hence, decreased apoptosis and/or increased cellular proliferation in the resultant embryos (Kues et al., 2002; Lee and Piedrahita, 2002). We also noticed that, in the presence of 3-hydroxyflavone in the embryo culture medium, treatment of serum-starved donor cells with the 3-hydroxyflanone did not had any additional benefit in improving the in vitro embryonic development (data not shown). Thus, beneficial effects of 3-hydroxyflavone on SCNT embryos was possibly due to scavenging of free radicals received from donor cells as well as from those generated during UV-irradiation of oocytes during enucleation and/or IVC-induced oxidative stress. Taken together, our data suggest that in vitro culture of cloned embryos in the presence of 3-hydroxyflavone can improve their in vitro development by negating the adverse effects of ROS generated upon UV irradiation of oocytes during enucleation and/or serum starvation of donor cells. It is also likely that inclusion of other antioxidants into the culture medium may have similar benefits in improving the in vitro development competence of cloned embryos produced by SCNT of serum-starved donor cells into recipient cytoplasts exposed to UV-irradiation during the enucleation step.
In conclusion, our data confirms the ROS-inhibiting effect of 3-hydroxyflavone and reinforces the possible involvement of ROS in the low development competence of cloned embryos produced by SCNT of serum-starved donor cells into recipient cytoplasts exposed to UV irradiation during the enucleation step. Culture of serum-starved donor cells and/or cloned embryos in the presence of 3-hydroxyflavone can improve their in vitro embryonic development and quality in terms of total number of cells per blastocyst and their ability to expand.
Footnotes
Acknowledgments
This work was supported by a grant (Code #20070401034017) from BioGreen 21 Program, RDA, Republic of Korea.
Author Disclosure Statement
The authors declare that no conflicting financial interests exist.