
Abstract
Abstract
In this study, we examined whether Hanganutziu-Deicher (H-D) antigens are important as an immunogenic non-α1,3-galactose (Gal) epitope in pigs with a disrupted α1,3-galactosyltransferase gene. The targeting efficiency of the AO blood genotype was achieved (2.2%) in pig fibroblast cells. A total of 1800 somatic cell nuclear transfer (SCNT) embryos were transferred to 10 recipients. One recipient developed to term and naturally delivered two piglets. The α1,3-galactosyltransferase activity in lung, liver, spleen, and testis of heterozygote α1,3-galactosyltransferase gene knockout (GalT-KO) pigs was significantly decreased, whereas brain and heart showed very low decreasing levels of α1,3-galactosyltransferase activity when compared to those of control. Enzyme-linked lectinosorbent assay showed that the heterozygote GalT-KO pig had more sialylα2,6- and sialylα2,3-linked glycan than the control. Furthermore, the heart, liver, and kidney of the heterozygote GalT-KO pig had a higher N-glycolylneuraminic acid (Neu5Gc) content than the control, whereas the lung of the heterozygote GalT-KO pig had Neu5Gc content similar to the control. Collectively, the data strongly indicated that Neu5Gc is a more critical xenoantigen to overcoming the next acute immune rejection in pig to human xenotransplantation.
Introduction
The enzyme α1,3-galactosyltransferase catalyzes the binding of α1,3-galactose (Gal) to N-acetyllactosamine (Galβ1,4GlcNAc) to produce Galα1,3Galβ1,4GlcNAc-R (α-Gal epitopes), which is found on the cell surface of almost all mammals with the exception of humans, apes, and Old World monkeys (Galili et al., 1988). The α-Gal epitopes are the major xenoantigens that cause hyperacute rejection in pig-to-human xenotransplantation (Good et al., 1992; Cooper et al., 1993; Galili, 2001). α-Gal epitopes are also involved in acute vascular rejection of xenografts (Galili, 2001; Logan, 2000; Sandrin and McKenzie, 1999). Heterozygote α1,3-galactosyltransferase gene knockout (GalT-KO) pigs exhibit reduced α-Gal epitope expression (Ramsoondar et al., 2003; Takahagi et al., 2003). Another study reported that baboon and human serum antibodies showed a different reactivity pattern to pig glycolipid antigens, indicating that nonhuman primates have limitations as a human preclinical model for immune rejection studies (Diswall et al., 2010). Recently, Byrne et al. (2008) identified an array of anti-non-Gal antibodies in baboons following Wild Type (WT) and GalT-KO pig cardiac transplantation. Also, Diswall et al. (2010) reported that knockout of α1,3-galactosyltransferase by gene targeting resulted in the elimination of Gal determinants. In addition, structurally novel α1,2 fucose-terminated blood group H compounds were identified in the GalT-KO tissue. These compounds are not expected to be recognized by the human immune system.
Recently, Kim et al. (2008) demonstrated that Neu5Gc-Gal-GlcNAc and Galα1-3Lewisx are novel antigens, by structural analysis of N-glycans from the miniature pig kidney using matrix-assisted laser desorption/ionization time-of-flight mass spectrometry (MALDI-TOF/MS) and tandem mass spectrometry (MS/MS). However, the proposed antigens appear to be related to the α1,3-galactosyltransferase and Hanganutziu-Deicher (H-D) antigens (Neu5Gc). Despite its importance in development and the need for a basic understanding of Hanganutziu-Deicher (H-D) antigens, the underlying mechanisms have been poorly studied. To this end, we examined whether the terminal Neu5Gc (H-D), and Sia-GalNAc-R (sialyl-Tn) structures in heterozygote GalT-KO pigs could represent possible targets for natural human antibodies.
Materials and Methods
The study protocol followed was according to the Institutional Animal Care and Use Committee of the South Korea National Institute of Animal Science (IACUC approval number: 2010-006, D-grade). Minnesota miniature swine were maintained at Finch University of Health Sciences/Chicago Medical School (North Chicago, IL, USA) in barrier-sustained specific pathogen-free facilities and were donated by Dr. Yoon Berm Kim of Rosalind Franklin University of Medicine and Science (North Chicago, IL, USA) (Setcavage and Kim, 1976; Sweeney and Kim, 2004). They were designated Seoul National University (SNU) miniature pigs.
ABO blood typing
Pig ear–derived tissues or cells were incubated with a lysis buffer solution (10 mM of Tris-HCl, pH 8.0, 0.1 M of EDTA) containing 0.5% sodium dodecyl sulfate (SDS) and 20 μL of 20 mg/ml proteinase K (Promega) at 55°C for 6 h. PCR amplification was performed using genomic DNA in a 20-μL reaction volume containing 50–100 ng of nDNA, 0.5 μM of specific primers for porcine AO blood group gene (A allele forward, 5′-CGCCAGTCCTTCACCTACGAAC-3′, reverse, 5′-CGGTTCCGAATCTCTGCGTG-3′; O allele forward, 5′-GGCATATGGAGGTTCCCAGGC-3′, reverse, 5′-AGAGTTCCCGTCGTGGCACAG-3′), 200 μM deoxyribonucleotide triphosphates (dNTPs), PCR buffer (10 mM Tris-HCl, pH 8.3, 50 mM KCl, 1.5 mM MgCl2), and 0.5 U of Supertherm™ DNA polymerase (JMR Holdings, Kent, UK) using a Thermocycler 3000 (Biometra, Goettingen, Germany). PCR conditions were designed with the following steps: denaturing at 95°C for 3 min, followed by 35 cycles of a three-step process of 1 min at 94°C, 45 sec at 66°C, and 45 sec at 72°C, and a final incubation at 72°C for 10 min. Electrophoresis was used to analyze the PCR products on a 1.5% agarose gel in 1×Tris-acetate-EDTA (TAE) buffer. The gel was viewed by staining with ethidium bromide and visualized under ultraviolet light.
Transfection and selection of donor cells
We isolated porcine ear fibroblasts from a 10-day-old male SNU miniature pig, and the fibroblasts were thawed and cultured in Dulbecco's modified Eagle's medium (DMEM) containing 20% fetal bovine serum (FBS), 1×nonessential amino acids, 1×sodium pyruvate, 10−4 M β-mercaptoethanol, 100 unit/mL penicillin, and 100 μg/mL streptomycin. The cells were trypsinized and resuspended at a concentration of 1.25×107 cells/mL in F10 nutrient mixture (WelGene). The GalT gene knockout was constructed as reported previously (Ahn et al., 2011), and DNA was purified according to the Qiagen's protocol for transfection (Qiagen, Valencia, CA, USA). Four hundred microliters of the cell suspension were electroporated with 10 μg DNA in a 4-mm cuvette with four pulses of 1 msec duration using 400 V capacitive discharges and an ECM 2001 Electro Cell Manipulator (BTX, Holliston, MA, USA). Electroporated cells were mixed with 10 mL of fresh medium and plated in 100-mm-diameter culture dishes. Two days later, selection was carried out for 12 days using a culture medium containing 300 μg/mL G418 (Gibco BRL, Carlsbad, CA, USA), and G418-resistant colonies were isolated using a cloning cylinder (Sigma-Aldrich). Colonies were transferred to wells of 24-well plates and passaged in 12-well plates, six-well plates, 60 mm-diameter culture dishes, and 100 mm-diameter culture dishes for expansion.
PCR analysis of targeted cells
To screen G418-resistant colonies, 200 μL of a cell suspension from wells of a 24-well plate was recovered by centrifugation. The cells were resuspended in 50 μL of distilled water containing 0.05 mg/mL proteinase K (Roche, Basal, Switzerland). To extract genomic DNA, the cells were incubated at 55°C for 130 min and heated to 98°C for 10 min to inactive proteinase K. To amplify the targeted allele, 25 μL of genomic extract was added to a sterile 0.5-μL tube containing 5 μL of 10×PCR buffer, 4 μL of 10 mM dNTP mixture, 1 μL of forward primer (10 pmol/μL; Bioneer), 1 μL of reverse primer (10 pmol/μL; Bioneer), and 1.25 U of EX Taq DNA polymerase (5 U/μL; TaKaRa, Shiga, Japan), and the volume was adjusted to 50 μL with distilled water. The forward primer was derived from intron 8 of the α1,3-galactosyltransferase gene sequence, 5′-ACCAGTCAGGTAAGCCACTCCACCTC-3′, and the reverse primer was derived from the α1,3-galactosyltransferase gene sequence that was not included in the KO vector, 5′-GTGCTGAACATCAAGTCAGTGCAATGGCTC-3′. PCR was carried out for 35 cycles in a thermal cycler (Bioneer). Each cycle consisted of denaturation for 30 sec at 94°C, annealing for 45 sec at 60°C, extension for 4 min at 72°C, and final extension for 10 min at 72°C. A 20 μL reaction sample was run on a 0.8% agarose gel using standard procedures.
In vitro maturation of oocytes
Prepubertal gilts (Duroc×Yorkshire×Landrace) ovaries were collected at a local slaughterhouse and transported to the laboratory in phosphate-buffered saline (PBS) prewarmed at 25–35°C. The antral follicles (2–6 mm in diameter) were collected by aspiration method using an 18-gauge needle. The aspirated oocytes contained an evenly granulated cytoplasm that was surrounded by at least three uniform layers of compact cumulus cells. The selected oocytes were washed three times in tyroid lactate (TL)-HEPES with 0.1% polyvinyl alcohol (PVA) and cultured in four-well plates containing medium (500 μL/well) supplemented with 10% porcine follicular fluid, 0.6 mmol/L cysteine, 1 mmol/L dibutyryl cyclic adenosine monophosphate (dbcAMP, Sigma-Aldrich), and 0.1 IU/mL human menopausal gonadotropin (hMG; Teikokuzoki, Tokyo, Japan) for 20 h. Then, the oocytes were cultured without dbcAMP and hMG for a further 18–24 h, as reported previously (Chae et al., 2006; Chae et al., 2008; Cho et al., 2007; Hill and Plank, 2004; Hwang et al., 2009; Kim et al., 2011; Lee et al., 2007; Park et al., 2004; Park et al., 2005; Park et al., 2009; Park et al., 2011a).
SCNT and embryo transfer
Briefly, mature eggs with the first polar body were cultured in North Carolina State University Medium-23 (NCSU-23 medium) supplemented with 0.4 mg/mL demecolcine (Sigma-Aldrich) and 0.05 mol/L sucrose for 1 h. Treated eggs with a protruding membrane were transferred to medium supplemented with 5 mg/mL cytochalasin B and 0.4 mg/mL demecolcine, and the protrusion was removed using a beveled pipette. A single donor cell was injected into the perivitelline space of each egg and electrically fused using two direct current pulses of 150 V/mm for 50 msec in 0.28 mol/L mannitol supplemented with 0.1 mM MgSO4 and 0.01% PVA (Sigma-Aldrich). Fused eggs were cultured in medium containing 0.4 mg/mL colcemid for 1 h before parthenogenesis activation. Then, the cells were cultured in 5 mg/mL of cytochalasin B–supplemented medium for 4 h. The reconstructed oocytes were activated by two direct current pulses of 100 V/mm for 20 msec in 0.28 mol/L mannitol supplemented with 0.1 mmol/L MgSO4, and 0.05 mmol/L CaCl2. These activated eggs were cultured in the NCSU-23 medium for 1–2 days in an atmosphere of 5% CO2 and 95% air at 39°C. Gilts at least 8 months old were used as recipients. Estrus was synchronized in recipients, as reported previously (Betthauser et al., 2000; Chae et al., 2006; Chae et al., 2008; Cho et al., 2007; Gong et al., 2004; Hill and Plank, 2004; Hwang et al., 2009; Kim et al., 2011; Lee et al., 2007; Park et al., 2004; Park et al., 2005; Park et al., 2009; Park et al., 2011a; Watanabe et al., 2005). The embryo transfer procedure was carried out surgically by transfer of SCNT embryos into the oviducts of synchronized recipients. The pregnancy status of the recipients was analyzed by ultrasonography between day 30 and day 35 up to delivery.
Preparation of the samples and protein determination
Both control and heterozygote GalT-KO organs were homogenized using a tissue grinder and liquid nitrogen. The organs were washed twice with PBS and then centrifuged at 1500×g for 10 min. The resulted organ homogenates were resuspended in 20 mM HEPES (pH 7.4) containing 0.5% Nonidet P-40 and protease inhibitor cocktail (Roche) and lysed by sonication for the α1,3-galactosyltransferase activity assay,. For sialic acids analysis, the organ homogenates were resuspended in Mammalian Lysis Buffer (Qiagen, Valencia, CA, USA) containing protease inhibitor cocktail and 1 U of nuclease and then lysed by sonication. The homogenates were centrifuged at 14,000×g for 10 min in a microcentrifuge tube precooled to 4°C, and the supernatants were transferred to a new microcentrifuge tube. Four volumes of ice-cold acetone were added to the protein fraction prior to incubation for 30 min on ice. The mixture was centrifuged for 10 min at 12,000×g in a microcentrifuge tube precooled to 4°C. The supernatant was discarded, and the pellet was air-dried. The dried pellet was resuspended in 20 mM HEPES (pH 7.4) containing 0.5% Nonidet P-40. The whole-organ lysates were frozen at −80°C. The thawed samples were centrifuged at 12,000×g for 10 min, and the supernatants were collected. The protein concentrations were assayed using a BCA (bicinchoninic acid) Protein Assay Kit (Pierce, Rockford, IL, USA).
α1,3-Galactosyltransferase activity assay
α1,3-Galactosyltransferase activity was assayed as previously described with minor modifications (Miyagawa et al., 1999; Tanemura et al., 1998). Lacto-N-neo-tetraose (LNnT; Sigma-Aldrich) acceptor substrate was labeled using 2-aminobenzamide (2-AB). The mixture, which contained 2 mg of LNnT, 0.2 mg of 2-AB, 0.24 mg of sodium cyanoborohydride, 6 μL of acetic acid, and 14 μL of dimethyl sulfoxide (DMSO), was incubated at 65°C for 3 h and purified using a GlycoClean S cartridge (ProZyme, Hayward, CA, USA). The assay mixture contained 20 mM HEPES buffer, 0.25% Nonidet P-40, 10 mM MnCl2, 33 mM NaCl, 3 mM KCl, 20 mM UDP-galactose, 200 mM galactose, 100 μM acceptor substrate (LNnT-AB) (pH 7.2), and 6 μL of the organ homogenates in a total volume of 20 μL. The reaction was terminated by boiling for 5 min. The samples were centrifuged at 15,000×g for 10 min. The resulted supernatant was analyzed by reversed-phase high-performance liquid chromatography (HPLC) using an ODS column (4.6×150 mm, TSK-gel column ODS-80TM; Tosoh Bioscience, Tokyo, Japan). The product and substrate were separated using 20 mM ammonium acetate buffer (pH 4.0) containing 0.15% n-butanol at 55°C. Each peak was detected using a model RF-10A fluorescence detector (Shimadzu, Tokyo, Japan) with an excitation and emission wavelengths of 330 and 420 nm, respectively.
Enzyme-linked lectinosorbent assays
Control and heterozygote GalT-KO organs were analyzed by enzyme-linked lectinosorbent assays (ELLA), using Maackia amurensis agglutinin (MAA) and Sambucus nigra agglutinin (SNA) (Sigma-Aldrich) (Shibuya et al., 1987; Wang and Cummings, 1988). A total of 50 μL of the organ lysates (25 μg of protein/well) were diluted in carbonate/bicarbonate buffer (pH 9.4), dispensed into 96-well microtiter plates, and incubated at room temperature for 2 h. After the organ lysates were dispersed and washed once with PBS containing 0.1% Tween 20 (PBST), blocking was performed with PBST containing 2% BSA. One hundred microliters of biotinylated MAA and SNA solutions (both 0.1 μg/mL) were applied and incubated at room temperature for 2 h. Then, the sample was washed three times using PBST and finally incubated for 2 h with 100 μL of horseradish peroxidase–conjugated hen egg white avidin (0.1 μg/mL). The reaction was enhanced and stopped using O-phenylenediamine dihydrochloride (Pierce) and 2.5 M sulfuric acid, respectively, as recommended by the manufacturer. The absorbance was measured at a 492 nm using a Multiskan FC microplate photometer (Thermo Scientific, Pittsburgh, PA, USA), and each signal level was normalized using actin (Bystryak and Mekler, 1992; Graham et al., 1984).
Analysis of sialic acid contents
A total of 0.2 mL of the control and heterozygote GalT-KO organs lysate (1 mg of protein) was mixed with an equal volume of 4 M propionic acid (pH 2.3). Sialic acids (Sias) were released by incubation at 80°C for 4 h (Malykh et al., 2003). The samples were cooled and centrifuged for 10 min at 20,000×g. The resulting pellets were resuspended in 0.5 mL of 0.1 M HCl, incubated for 50 min at 80°C to completely release Sias from glycoconjugates, and centrifuged as described above. The supernatants recovered after hydrolyses were passed through an Amicon Ultra-0.5, Ultracel-3 Membrane filter (Millipore, Bedford, MA, USA), and the total released Sias samples were lyophilized, resuspended in 10 μL of 2 M acetic acid, and incubated at 55°C for 2 h in the dark with 50 μL of a reagent consisting of 7.0 mM 1,2-diamino-4,5-methylenedioxybenzene (DMB), 0.75 M 2-mercaptoethanol, and 18 mM NaHSO3 dissolved in water. Samples were cooled after derivatization, and 10-μL aliquots were analyzed by HPLC on a reversed-phase ODS column (4.6×150-mm, TSK-gel column ODS-80TM; Tosoh Bioscience) with a 30% methanol mobile phase. The flow rate was maintained at 0.5 mL/min, and the derivatized N-acetylneuraminic acid (Neu5Ac) and N-glycolylneuraminic acid (Neu5Gc) were detected using a model RF-10A fluorescence detector (Shimadzu, Tokyo, Japan) at an excitation and emission wavelengths of 373 nm and 448 nm, respectively (Malykh et al., 1998). Quantification of Neu5Ac and Neu5Gc was performed by measuring the chromatographic peak areas for each sample and by using a calibration curve obtained for the DMB derivative of standard Neu5Ac and Neu5Gc (Sigma-Aldrich).
Statistical analysis
Values are reported as the mean±standard deviation (SD). All experimental data on α1,3-galactosyltransferase activity, ELLA, and analysis of Sias contents were acquired using 3 control pigs and 2 heterozygote GalT-KO pigs. Each sample was evaluated in triplicate experiments. Statistical significances were confirmed by the t-test and one-way analysis of variance (one-way ANOVA).
Results
Pig AO blood typing in donor cells for SCNT
Antibodies against α-Gal antigens are a vital part of xenotransplantation and carbohydrate antigens, such as AO blood, and are possible targets of the immune response to transplanted organs or tissue (Smith et al., 2006). To overcome incompatible xenotransplantation, we developed methods for the production of ABO-compatible organs incompatible xenograft. Previously, we reported that the O allele has a large deletion between exon 7 of the AO blood group gene and the neighboring SURF 6 and that the ABO blood group antigens in human and the AO blood group antigen in pig are coded by a mutation within the orthologous glycosyltransferase gene (Nguyen et al., 2011). Eighteen SNU germ-free miniature pigs were genotyped using multiple PCR primers. Among them, the frequency of pigs with blood group A (genotyped as AO) was slightly lower (41.89%) than blood group O (58.11%), whereas no blood type AA homozygotes were detected. Presently, we designed a set of O allele–specific PCR primers, which yielded a 216-bp fragment. Because exon 8 primers produced only 340-bp products from the A allele, the multiple PCR results from the two primer sets provided clear genotyping evidence of a porcine AO blood group gene (Fig. 1). The allele frequency of O and A alleles was 0.79 and 0.21, respectively. Donor cells for SCNT were obtained from small ear tissue of a male SNU miniature pig of blood group AO.
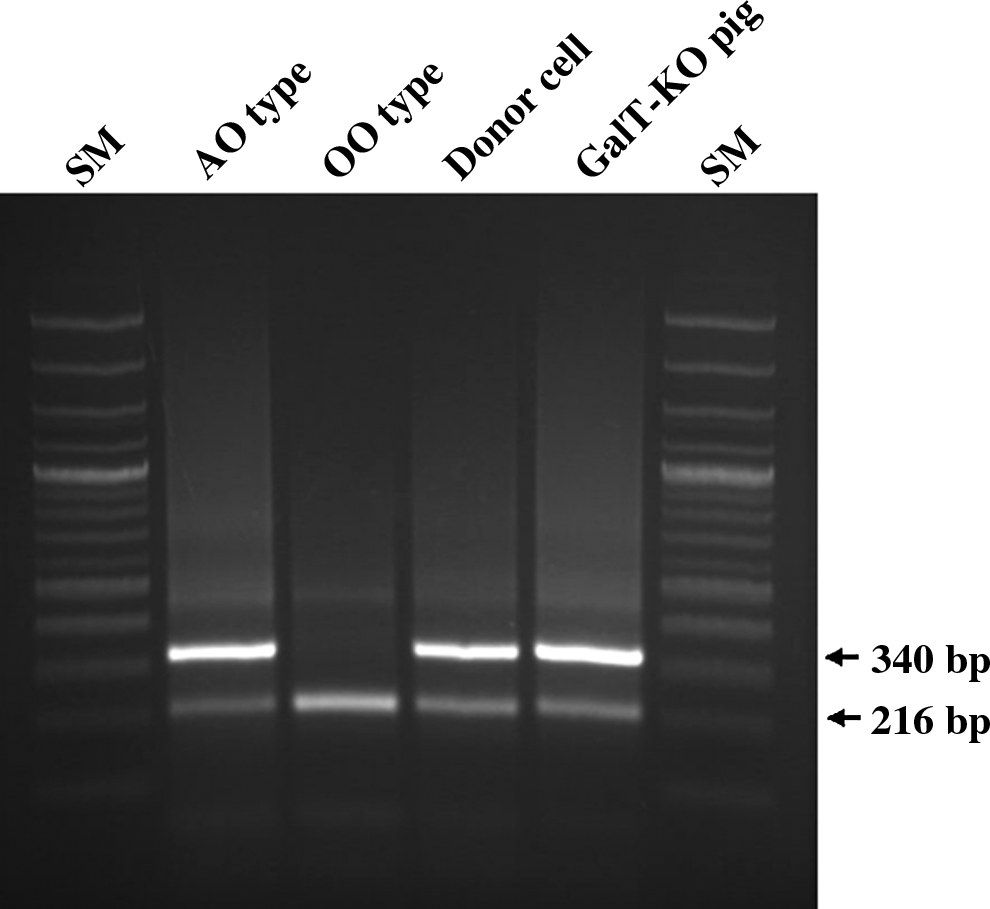
PCR-based A or O blood typing. For genotying, PCR was performed with two pairs of primers specific to alleles A and O. Lanes 1 and 6 are 100-bp ladders of DNA size marker (SM). If both 340-bp (specific to allele A) and 216-bp (specific to allele O) bands were amplified, the blood genotype was AO blood type (lane 2), whereas if the single 216-bp (specific for allele O) band was amplified, the genotype was OO (lane 3). Lanes 4 and 5 show the results amplified from donor cells and heterozygote α1,3-galactosyltransferase gene knockout (GalT-KO) pig, respectively, indicating the AO blood type.
Production of heterozygote GalT-KO gnotobiotic miniature pigs with blood type AO
The pig genomic DNA encoding the α1,3-galactosyltransferase locus on chromosome 1 consists of nine exons, with the translation start codon located within exon 4. A targeting vector was designed to disrupt the α1,3-galactosyltransferase gene via homologous recombination in pig fibroblast cells. The vector contained 4.8 kb of 5′- and 1.9 kb 3′-homologous DNA. Positive and negative selection was performed by using a PGK-neo cassette and an MC1 promoter-diphtheria toxin-A (pMC1-DTA) selection cassette, respectively. The targeted pig fibroblast cells expressed neomycin. The pMC1-DTA cassette was located at the end of the 3′ homology arm, and its incorporation was hindered by the homologous recombination. In the targeted cells, PGK-neo replaced exon 9 of the α1,3-galactosyltransferase genomic gene, resulting in a loss-of-function mutation. To screen neomycin-resistant pig fibroblast cells, PCR primers were used in the region of homologous 3′ arm. The forward 5′ PCR primers were located in intron 8 of the α1,3-galactosyltransferase gene, whereas the reverse 5′ PCR primer was located in the 3′ untranslated region of the α1,3-galactosyltransferase gene, which was not included in the KO vector. In correctly targeted cells, the size of the expected PCR product was 2.3 kb and 3.6 kb.
To remove nontransfected or spontaneous resistant cells after G418 treatment, pig-derived fibroblast cells were transfected with the GalT-KO vector. At 10 days posttransfection, 232 single cell–derived colonies were selected and transferred into 24 wells, cultured in the presence of G418, and then replated. A few cells from each passage were directly subjected to PCR to determine if the pig α1,3-galactosyltransferase gene was disrupted via homologous recombination. Among them, 10 colonies were positive for α1,3-galactosyltransferase gene disruption. The targeting efficiency in pig fibroblast cells was approximately 2.2%. The transfected cells were then used as donor cells in SCNT, and a total of 1800 SCNT embryos were transferred to 10 recipients. Among them, one recipient developed to term and naturally delivered two piglets. We designed PCR primers to amplify the sequence of the genomic DNA flanking the region of the α1,3-galactosyltransferase gene targeted for further confirmation that one of the α1,3-galactosyltransferase alleles was disrupted in pig fibroblast cells. Our results revealed that the α1,3-galactosyltransferase targeting vector replaced the endogenous α1,3-galactosyltransferase locus through homologous recombination (data not shown).
Heart, liver, and kidney of heterozygote GalT-KO pig displays higher N-glycolylneuraminic acids as the xenoantigenic determinant
We investigated the profiles of α1,3-galactosyltransferase enzyme activities of several organs, including the brain, heart, lung, liver, kidney, stomach, spleen, small intestine, muscle, and testis, in heterozygote GalT-KO and control pigs. α1,3-Galactosyltransferase activities of all heterozygote GalT-KO organs were lower compared to their control counterparts (Fig. 2). Among them, organs such as lung, kidney, spleen, small intestine, muscle, and testis of heterozygote GalT-KO were significantly decreased, whereas a slight decrease in the level of α1,3-galactosyltransferase activity was observed in the brain and heart compared to those of the control. Next, we examined the possibility of whether sialyloligosaccharide epitopes containing Sias, such as Neu5Ac and Neu5Gc, were increased in heterozygote GalT-KO organs by decreasing α-Gal. ELLA was performed using sialyloligosaccharide-specific lectins, such as MAA, that bind preferentially to sialylα2,3-linked glycan and SNA lectin, which binds to sialylα2,6-linked glycan, as previously described (Park et al., 2011b). Our study shows that the organs of heterozygote GalT-KO pigs had higher MAA and SNA signal levels when compared to the control (Fig. 3). This data indicates that heterozygote GalT-KO pigs have more sialylα2,3- and sialylα2,6-linked glycoconjugates than the control. Finally, we examined the Sias content in major xenograft organs, such as the heart, lung, liver, and kidney of heterozygote GalT-KO and control pigs (Table 1). The Sias content in all of the organs from heterozygote GalT-KO pigs was higher when compared with those of the controls. Except in the lung, all organs showed increased levels of Neu5Gc than Neu5Ac. In addition, the heart, liver, and kidney of heterozygote GalT-KO pigs showed a higher Neu5Gc content than the controls; however, the Neu5Gc content in the lung of the heterozygote GalT-KO pigs was similar to that of the controls (Fig. 4).

Assay of α1,3-galactosyltransferase activity. Enzyme activity was measured by reversed-phase high-performance liquid chromatography (HPLC) using a fluorescence detector with 2-aminobenzamide-labeled lacto-N-neo-tetraose as the substrate. α1,3-Galactosyltransferase activity from each organ lysate (15 μg of protein, 6 h at 37°C) was examined. Each value is the mean±standard deviation (SD) of independent triplicate examinations. Statistical significances were confirmed by t-test. (*) p<0.05; (***) p<0.001. The liver data were obtained from our previous data (Park et al., 2011b).

Enzyme-linked lectinosorbent assays (ELLA) of Maackia amurensis agglutinin (MAA) and Sambucus nigra agglutinin (SNA) lectins in the control and heterozygote GalT-KO pig organs. Normalized relative signal levels of the MAA (
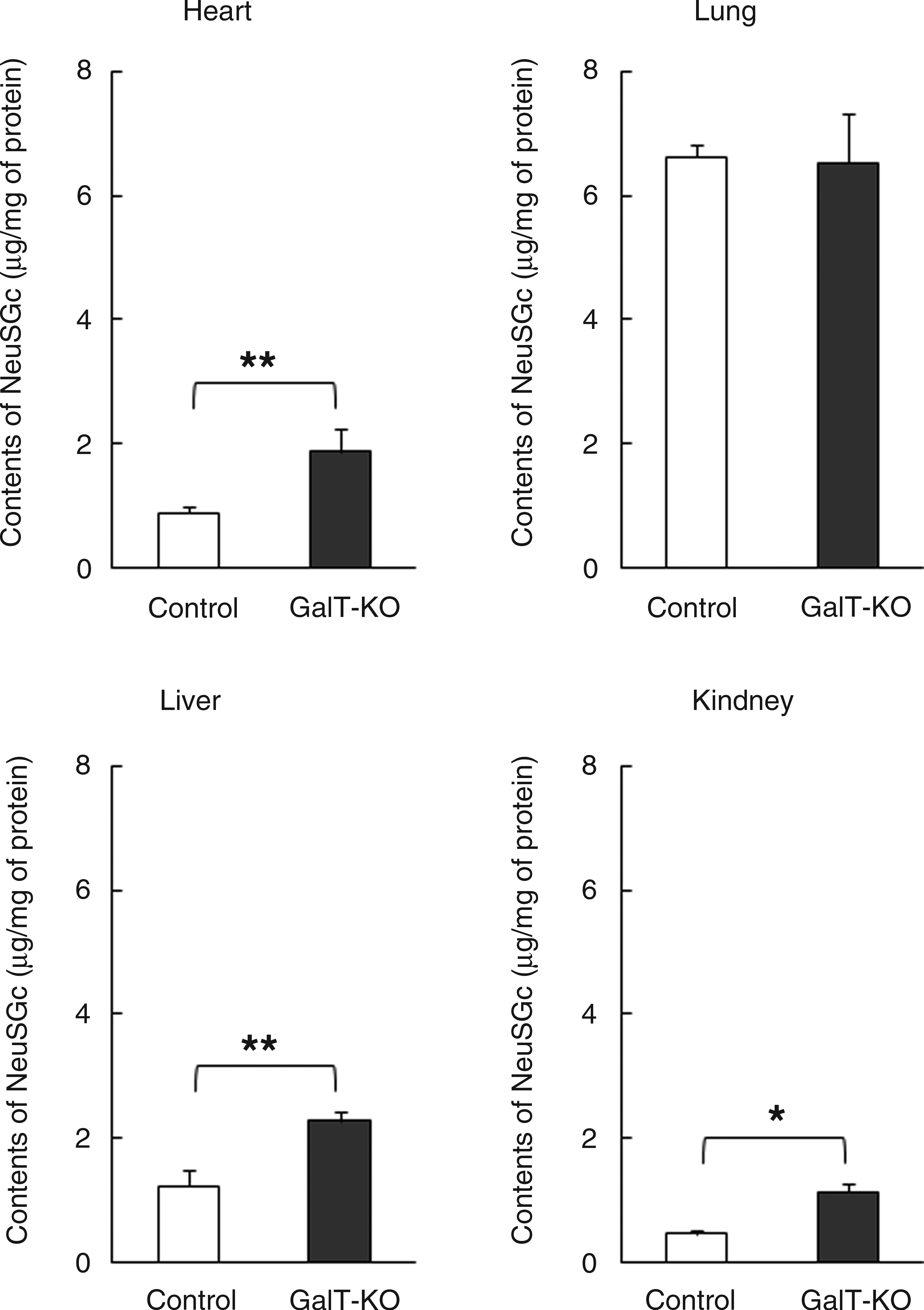
Comparison of N-glycolylneuraminic acid (Neu5Gc) contents between control and hetrozygote GalT-KO pig organs. After isolation of the glycoproteins from each of organ lysates of control and heterozygote GalT-KO heart, lung, liver, and kidney, free Neu5Gc was liberated by weak acid treatment of glycoproteins. The Neu5Gc content was determined based on the signal intensities (peak areas) of each 1,2-diamino-4,5-methylenedioxybenzene (DMB) fluorescence–labeled Neu5Gc. Each value is the mean±SD of triplicate determinations. Statistical significances were confirmed by t-test. (*) p<0.05; (**) p<0.005.
Each value is the mean±standard deviation (SD) of triplicate determinations. (*) p<0.05; (**) p<0.01; (***)p<0.001.
GalT-KO, α1,3-galactosyltransferase gene knockout; Neu5Gc, N-glycolylneuraminic acid; Neu5Ac, N-acetylneuraminic acid.
Discussion
Although in our previous study we produced SCNT knockout pigs by genetic deletion of α1,3-galactosyltransferase (Ahn et al., 2010), we ignored selection and use of animals with ABO-compatible blood types during the process of transplanting pig organ and tissues into humans. In this study, we selected donor cells with an AO blood type and then developed a heterozygote GalT-KO pig, which partially lacked α-Gal epitopes. This study provides a new avenue to reduce the risk of xenotransplantation across ABO barriers.
The pig has long been a model favored for biomedical purposes (Kim et al., 2009; Lai et al., 2002). However, xenotransplantation programs face two major barriers. First, the double-edged sword of the immune system has to be overcome, especially the problems associated with rejection of implanted cells, tissues, and/or organs (Ezzelarab et al., 2005; Houser et al., 2004; Kuwaki et al., 2005; Sandrin and McKenzie, 1994; Yamada et al., 2005). Second, prevention of cross-species infections (zoonotic infections) must be ensured (Fishman et al., 1994). The main obstacle of hyperacute rejection is being addressed using genetic modifications of pigs, such as making of transgenic pigs with human CD55 and CD59 (Menoret et al., 2004; Ramirez et al., 2005), and KO of the α1,3-galactosyltransferase genes (Dai et al., 2002; Kuwaki et al., 2005; Lai et al., 2002; van Poll et al.; Yamada et al., 2005). In humans, the α-Gal epitope is absent, but all humans produce antibodies against the α-Gal epitope, which is called anti-Gal (Sandrin and McKenzie, 1994). However, pigs naturally express ten million different types of glycolipid and glycoprotein epitopes on each cell. The binding of the anti-Gal antibody molecules of the recipient to α-Gal epitopes on the endothelial cells of the xenografts activates complement and platelet aggregation. Finally, it resulted in the collapse of the vascular bed and hyperacute rejection of the graft (Ezzelarab et al., 2005). We developed a heterozygote GalT-KO pig by disruption of the α1,3-galactosyltransferase gene by insertion of the neomycin resistance gene into the region of the gene that encodes exon 9 (Ahn et al., 2011). A previous study indicated that organs from transgenic animals lacking the major xenoepitope α-Gal by KO of the α-1,3-galactosyltransferase gene were remarkably protected from human complement-mediated injury, but that xenografted organs from pig to baboon ultimately failed due to acute humoral xenograft rejection (Houser et al., 2004). This observation warrants further development for controling acute vascular and cellular rejection and chronic rejection of xenografts (e.g., by induction of tolerance).
With the exception of humans and birds, Neu5Gc, which is a component of the H-D antigen, is widely distributed in mammalian species, including monkeys and apes (Asaoka et al., 1992; Hayakawa et al., 2006; Schauer et al., 2009; Varki, 2001). Neu5Gc is one of the most crucial epitopes in pig-to-human xenotransplantation; it might be a possible target for natural antibodies, and it can elicit antibody production in human serum (Ezzelarab et al., 2005). Therefore, we addressed whether the H-D antigens are important as an immunogenic non-Gal epitope by investigating the signal levels of sialyloligosascharide with ELLA. MAA consists of two molecules, which are a hemagglutinating hemagglutinin (MAH) and a mitogenic leukoagglutinin (MAL), and are able to interact with sialic acid–containing glycoconjugates. The MAL specifically binds to the Siaα2,3Galβ1,4GlcNAc structures of N-glycans, but the MAH has higher affinity for Siaα2,3 Galβ1,3[Siaα2,6]GalNAc present on O-glycan. In addition, the SNA lectin specifically binds to Siaα2,6Gal/GalNAc structures of N- or O-glycan chains (Haseley et al., 1999; Imberty et al., 2000; Shibuya et al., 1987). As shown in Figure 3A, the expression level of MAA in all heterozygote GalT-KO organs was higher than in the control. Moreover, the increased SNA signal levels of heterozygote GalT-KO organs represent an increase in the Siaα2,6GalNAc-R structures, which is an another xenoantigenic epitope (Fig. 3B). Taken together, these results might indicate that the sialyltransferases catalyze the transfer of Sias to a terminal uncapped β1,4Gal of N-acetyllactosamine caused by decreased α1,3-galactosyltransferase activity and produce more sialylα2,3- and sialylα2,6-linked glycan chains in heterozygote GalT-KO organs than in controls.
Most mammals express two common Sias, Neu5Ac and Neu5Gc. Because the contents of Sias were likely altered by the decrease in α1,3-galactosyltransferase activity in the heterozygote GalT-KO organs, we measured the Neu5Gc content in major xenograft organs, such as the heart, lung, liver, and kidney between heterozygote GalT-KO and control pigs. As expected, the data clearly showed that the Neu5Gc contents were increased in GalT-KO–derived organs relative to the control (Fig. 4). However, interestingly, no difference in Neu5Gc content was observed in the lungs between heterozygote GalT-KO and control pigs. Even though α1,3-galactosyltransferase activity of the heterozygote GalT-KO lung was significantly decreased relative to the control, the α1,3-galactosyltransferase activity was 3- to 10-fold higher than those of heterozygote GalT-KO heart, liver, and kidney (Fig. 2). It is conceivable that the α1,3-glactosyltransferase in the heterozygote GalT-KO lung reacted as a more dominant enzyme than sialyltransferases so that the nonreducing N-acetyllactosamine residue could be more preferentially bound to α-Gal than Sias. This is demonstrated by the signal levels of ELLA by MAA and SNA in the control, and the heterozygote GalT-KO lung cells were not changed when compared with those of heart, liver, and kidney (Fig. 3). It is estimated that the sialyltransferases in heterozygote GalT-KO lung might act as a minor enzyme that results in no change of Sias- bound glycoconjugates.
In the present study, a decrease in the α1,3-galactosyltransferase activity was evident, although the heterozygote GalT-KO lung retained α1,3-galactosyltransferase as the dominant enzyme, clearly increasing the Neu5Gc as a component of the H-D antigen in pig organs, such as the heart and kidney. These data correlate with our previous data; α2,3- and α2,6-sialyltransferase in heterozygote GalT-KO liver showed higher activities in comparison to control (Park et al., 2011b). The increased α2,6-sialyltransferase may specifically transfer Neu5Gc to the nonreducing galactose of the N-acetyllactosamine residue (Hamamoto et al., 1995). This might have a relationship to the link between pig α1,3-galactosyltransferase and Neu5Gc in the intracellular enzyme environment and indicates that the deficiency of GalT increased the content of α2,6-linked Neu5Gc glycoconjugates. Our results suggest that Neu5Gc, which is a prime non-Gal antigen candidate for the prevention of the hyperacute rejection, is a more critical xenoantigen for overcoming the next acute immune rejection problem in pig against human xenotransplantation.
Finally, the selection and use of animals with ABO-compatible blood types during the process of transplanting pig organ and tissues into humans can contribute positively to acute immune rejection due to the difference in the blood group. Several researchers have reported successful cloning of miniature pigs for xenotransplantation due to their anatomical and physiological similarities to humans and short gestation period and production of multiple offspring (Hao et al., 2006; Kurome et al., 2008; Li et al., 2010; Miyoshi et al., 2007; Wakai et al., 2008). However, ABO typing of these miniature pigs is still unknown. Therefore, in this study, PCR blood genotyping of donor cells was performed using two pairs of primers specific to porcine blood group gene alleles A and O. Both a 216-bp fragment specific to allele O and a 340-bp fragment specific to allele A were amplified from donor cells and KO pigs, indicating that the blood genotype of donor cells and KO pigs was AO. These findings imply that blood type A patients who cannot produce anti-A antibodies will have a sharply reduced xenotransplantation risk across ABO barriers if heterozygote GalT-KO pigs are used to derive the donor cell with blood type A. Experimental data of Galili et al. (2006) showed that the anti-Gal specificity in humans comprises a very large proportion of anti-A and anti-B activities. For example, from blood type A donor, blood type O recipients transplanted with a kidney allograft produce anti-Gal B antibodies as well as anti-Gal A/B, which bind to the epitope of α-Gal A in both blood groups A and B (Galili et al., 2002). Therefore, a heterozygote GalT-KO pig with blood type AO can foster homozygote piglets with blood types AA or OO by breeding heterozygote female and male pigs with blood type AO. This is the reason we used an AO cell line for production of a SCNT-derived KO pig. On the basis of the immune response to the ABO-incompatible antigen on the xenograft, it would help to minimize rejection.
In conclusion, we have developed a heterozygote GalT-KO pig that partially lacks α-Gal epitopes, where the α1,3-galactosyltransferase gene was disrupted by insertion of the neomycin resistance gene into the region of the gene that encodes exon 9. The pig, which had blood type AO, may be a possible clinical application for pig–human xenotransplantation. Although our gnotobiotic facility system has limitations, we successfully produced a sperm with high motility from the heterozygote GalT-KO pig. Therefore, these findings would give some solution to the puzzle of why GalT-KO–derived pig organs transplanted to the animals like baboon lead to acute rejection 179 days after transplantation (Kuwaki et al., 2005).
Footnotes
Acknowledgments
The author dedicates this study to Dr. Kyung-Kwang Lee, who died on March 1, 2009, in Daejeon, Republic of Korea, and who made exceptional contributions to research on animal biotechnology and cloning. He was only 58 years old when he died, finally succumbing to cancer of the lung during performance of this project. We will memorize his unlimited affection and contributions for this project forever. This work was supported by Woo Jang-Choon project (PJ007849) from the Rural Development Administration (RDA) and IPET (111047051CG000), Republic of Korea.
Author Disclosure Statement
The authors declare that there is no conflict of interest.