
Abstract
Abstract
Aberrant epigenetic nuclear reprogramming of somatic nuclei is a major cause of low success in cloning. It has been demonstrated that treatment of histone deacetylase inhibitors (HDACi) enhances developmental potential of somatic cell nuclear transfer (SCNT) embryos by alteration of epigenetic status. The aim of the present study was to investigate the effect of oxamflatin, a novel HDACi, on the developmental competence of porcine SCNT embryos. Treatment with 1 μM oxamflatin for 9 h after activation of SCNT embryos increased both in vitro and in vivo developmental competence. Treatment of SCNT embryos with 1 μM oxamflatin significantly increased blastocyst rate and total cell number in blastocysts (33.3±6.0 and 73.1±1.6, respectively) than that of controls (10.3±3.7 and 54.1±3.5, respectively) or scriptaid (16.4±4.6 and 64.4±2.1, respectively). Moreover, oxamflatin showed significant higher overall cloning efficiency from 0.9% to 3.2%, whereas scriptaid demonstrated 0% to 1.8%. In conclusion, these results indicate that oxamflatin treatment improves the developmental competence of porcine SCNT embryos.
Introduction
Several epigenetic remodeling drugs, such as the histone deacetylase inhibitors (HDACi) trichostatin A (TSA) (Beebe et al., 2009; Ding et al., 2008; Kishigami et al., 2006; Li et al., 2008), valporic acid (Costa-Borges et al., 2010; Miyoshi et al., 2010), sodium butyrate (Das et al., 2010), suberoylanilide hydoxamic acid (SAHA) (Ono et al., 2010), or DNA methyltransferase inhibitor 5-azacytidine (Ding et al., 2008; Enright et al., 2003), have been studied to improve the developmental potential of SCNT embryos. There are reports that scriptaid enhances developmental competence, especially in porcine embryos (Zhao et al., 2009;Zhao et al., 2010). However, low efficiency and cellular toxicity require further studies. Recently, Wakayama et al. found that oxamflatin, which can inhibit classes I and IIa/b HDACs (Blackwell et al., 2008; Chuang et al., 2009), is significantly effective in improving the full-term development of cloned mice and increasing establishment of nuclear transfer-generated embryonic stem cell (ESC) lines without obvious abnormalities (Ono et al., 2010).
Hence, it is necessary to explore whether the oxamflatin treatment has beneficial effects on somatic cloning in species other than the mouse and cow, especially its effect on the full-term development. The objective of this study was to investigate, optimize, and compare the application of scriptaid and oxamflatin on the reprogramming of somatic nuclei after SCNT on in vitro and in vivo development.
Materials and Methods
Collection of oocytes and in vitro maturation
Porcine ovaries were collected at a local abattoir and transported to the laboratory (within 2 h after collection) in sterile physical saline at 30–35°C. Cumulus–oocyte complexes (COCs) were aspirated from antral follicles (3–6 mm) with an 18-gauge needle attached to a 10-mL disposable syringe. COCs with several layers of cumulus cells and uniform cytoplasm were chosen and cultured in tissue culture medium (TCM)-199 (Invitrogen, Carlsbad, CA, USA) supplemented with 10 ng/mL epidermal growth factor (EGF), 0.57 mM cysteine, 0.91 mM sodium pyruvate, 5 μg/mL insulin, 1% (vol/vol) penicillin-streptomycin (Pen-Strep; Invitrogen), 0.5 μg/mL follicle-stimulating hormone, 0.5 μg/mL luteinizing hormone, and 10% porcine follicular fluid at 39°C in a humidified atmosphere of 5% CO2, first, with gonadotropin-releasing hormone (GnRH) for 22 h and then without them for a further 22 h in four-well dishes (Nunc, Roskilde, Denmark). COCs were washed with same in vitro maturation (IVM) media at each step. After a total of 44 h maturation culture, oocytes were denuded by pipetting with 0.1% hyaluronidase in Dulbecco's phosphate-buffered saline (DPBS) (Invitrogen) supplemented with 0.1% polyvinyl alcohol. Denuded oocytes with evenly granulated and homogeneous cytoplasm were selected and then used for parthenogenesis and SCNT.
Oxamflatin was prepared according to the method of Tetsuo et al. (Ono et al., 2010). Briefly, oxamflatin was dissolved in dimethyl sulfoxide (DMSO) as 200× stock solutions and stored at −20°C. These reagents were added to the activation medium to 10× of experimental concentration (0.1, 1, or 10 μM).
Parthenogenesis
Selected oocytes were equilibrated in pulsing medium and transferred to a chamber containing two electrodes overlaid with the pulsing medium. The pulsing medium was 0.26 M mannitol solution containing 0.5 mM HEPES, 0.1 mM CaCl2, and 0.1 mM MgSO4. Oocytes were activated with a single DC pulse of 1.5 kV/cm for 60 μsec using a BTX electro-cell Manipulator 2001 (BTX, Inc., San Diego, CA, USA).
Primary cell establishment and donor cell preparation
At day 35, the female was euthanized, and fibroblasts were derived from fetuses and established as previously described (Cho et al., 2011). Briefly, the fetuses were recovered and rinsed three times with Ca2+- and Mg2+-free DPBS (Invitrogen, CA, USA). After removal of head, intestine, liver, limbs, and heart, the remaining tissues were finely minced into pieces (1 mm3) using scissors in DPBS. Minced tissue pieces were digested with 200 IU/mL collagenase (Invitrogen, CA, USA) and in Dulbecco's modified Eagle's/Nutrient Mixture F-12 medium (DMEM/F12; Invitrogen, CA, USA) for 9 h at 38.5°C and 5% CO2 in air. The digestion was rinsed in cell culture medium by centrifugation at 450×g for 2 min to remove the residual enzyme. The cell pellet was resuspended in DMEM/F12 supplemented with 10% fetal bovine serum (FBS; Invitrogen, CA, USA) and 1% Antibiotic-Antimycotic Solution (Invitrogen, CA, USA), seeded in a 75-cm2 culture dishes (Falcon, CA, USA), and left to culture until confluent.
For SCNT, the cells were thawed, cultured, and subsequently used between passages four and seven.
Somatic cell nuclear transfer
For SCNT, a micromanipulator (NT-88-V3, Nikon-Narishige, Tokyo, Japan) attached to an inverted microscope (TE2000, Nikon Instrument, Tokyo, Japan) was used. A cumulus-free oocyte was held with a holding micropipette, and the zona pellucida was partially dissected with a fine glass needle to make a slit near the adjacent cytoplasm, presumably containing the metaphase II chromosomes, which were extruded by aspiration with the same needle. Enucleation was confirmed by staining the cytoplasm with 0.5 μg/mL bisbenzimide (Hoechst 33342) during manipulation. A single fibroblast cell with a smooth surface was selected under a microscope and transferred into the perivitelline space of enucleated oocytes. Membrane fusion was performed according to Cho et al. (2011). Briefly, COCs were placed in a 280 mM mannitol solution (pH 7.2) containing 0.15 mM MgSO4, 0.01% (wt/vol) polyvinyl alcohol (PVA), and 0.5 mM HEPES and held between two electrode needles. Membrane fusion was induced with an electro cell fusion generator (LF101, Nepagene, Ichikawa, Japan) by applying a single direct current pulse (200 V/mm, 20 μsec) and a pre- and postpulse alternating current field of 5 V, 1 MHz, for 5 sec, respectively. The reconstructed embryos were cultured in porcine zygote medium-5 (PZM-5) (IFP0410P, Funakoshi, Tokyo, Japan) for 1–1.5 h and then subjected to electrical activation. Electrical activation was performed the same as for parthenogenetic embryos.
Postactivation treatment and embryo culture
Electrically activated embryos were washed and chemically activated using 2 mM 6-dimethylaminopurine (6-DMAP) and 7.5 μg/mL cytochalasin B supplemented with various concentrations of oxamflatin (0, 0.1, 1, 10 μM) or scriptaid for 3 h in PZM-5 (IFP0410P, Funakoshi, Tokyo, Japan). Embryos were then cultured in PZM-5 at the same concentrations of oxamflantin or 500 nM scriptaid as above for a further 6, 11–13 h, respectively. After three washes in PZM-5, embryos were cultured in 450 μL of PZM-5 covered with mineral oil. The embryos were cultured in a humidified atmosphere of 5% CO2, 5% O2, and 90% nitrogen at 38.5°C for 7 days. Cleavage rates were evaluated on day 2. On day 4, 10% FBS was added to the culture medium. Blastocyst formation and the number of nuclei were determined in day 7. Briefly, blastocysts were fixed in absolute alcohol and nuclei were stained with 25 μg/mL bisbenzamide (Hoechst 33342) overnight at 4°C. Fixed and stained blastocysts were mounted on a glass slide in a drop of glycerol, gently flattened with a cover glass, and visualized for cell counting with a fluorescence microscope using a 346-nm excitation filter.
Total RNA extraction, RT-PCR, and real-time PCR
Total RNA was isolated from fresh or previously frozen (−80°C) blastocysts using the easy-spin™ (DNA free) Total RNA Extraction Kit (iNtRON Biotechnology, Seoul, Korea) according to the manufacturer's instructions, quantified by a NanoDrop 2000 Spectrophotometer (Thermo Fisher Scientific, Wilmington, DE, USA), and immediately stored at −80°C until used for reverse transcriptase PCR (RT-PCR) and quantitative real-time PCR (qRT-PC)R. cDNA was produced from 1 μg of total RNA from blastocysts samples using a SuperScript™ III First-Strand cDNA Synthesis Kit (Invitrogen) primed with oligonucleotide-dT (18-mer) and followed by RNase H digestion of RNA in a total volume of 20 μL as per the manufacturer's instructions. qRT-PCR was performed in a 7300 Real Time PCR System (Applied Biosystems, Foster, CA, USA) using the SYBR premix ExTaq perfect Real Time (Takara Bio Inc., Shiga, Japan) with little modification. In brief, all the primers were standardized by standard curve. The PCR plate (MicroAmp optical 96-well reaction plate, Singapore) was made by adding 2 μL of cDNA, 1 μL (10 pM) of forward primer, 1 μL (10 pM) of reverse primer, 10 μL of SYBR Premix Ex Tag (Takara Bio Inc), 0.4 μL of ROX Reference Dye (Takara Bio Inc.), and 5.6 μL of nuclease-free water (Ambion Inc., Austin, TX, USA). Four replications were made for each sample. Primer sequences and the approximate sizes of the amplified fragments of all transcripts are shown in Table S1 (Supplementary Data are available at www.liebertonline/cell/) containing the specific primer to amplify POU5F1, NANOG, CDX2, REX01, HDAC1, HDAC2, BCL2L1, and BAX.
Embryo transfer and pregnancy diagnosis
Embryo transfer was performed according to the report of Koo et al. (2009). Reconstructed embryos were cultured in vitro for 1–2 days, and after cleavage check, 70–100 embryos were loaded into a sterilized 0.25-mL straw (Minitüb, Tiefenbach, Germany) and kept in a portable incubator (Minitüb) during transportation to the embryo transfer facility. An estrus-synchronized recipient was anesthetized by a combination of ketamine (1.13 mg/mL; Yuhan ketamine®, Yuhan Corp., Seoul, Korea) and xylazine (0.3 mg/kg; Celactal®, Bayer Animal Health Corp., Tokyo, Japan) intravenously for induction and 3% isoflurane (Ifran®, Hana Pharm Co., Ltd, Hwasung, Korea) for maintenance. One oviduct was exposed by laparotomy. The straw containing the embryos was inserted directly into the oviduct of the recipient, and embryos were expelled from the straw using a 1-mL syringe (Becton Dickinson). We transferred embryos to 11 recipients. When the SNU1 cell line was used as the nuclear donor, SCNT embryos were transferred to seven surrogates. When the SNU2 cell line was used, SCNT embryos were transferred to four. Recipients were checked for pregnancy by transabdominal ultrasound examination at day 30 after embryo transfer.
Statistical analysis
All data were subjected to one-way analysis of variance (ANOVA) followed by Tukey's test using Prism version 4.0 (Graphpad Software, San Diego, CA, USA) to determine differences among experimental groups. Statistical significance was determined when the p value was less than 0.05.
Results
Effect of different concentrations of oxamflatin on development of porcine parthenogenetic embryos in vitro
Developmental competence was assessed in terms of cleavage rate, blastocyst rate, and total cell number in the blastocyst. No significant difference was observed in the cleavage rate, blastocyst rate, and total cell number in blastocysts between parthenogenetic embryos treated with 0.1 μM or 1 μM oxamflatin. However, cleavage rate, blastocyst rate, and total cell number in blastocysts of the 10 μM oxamflatin-treated group were significantly lower (p<0.05) than those of control (Table 1).
Values are mean±standard error of the mean (SEM). Values with different superscripts in the same column differ significantly (p<0.05). Experiments were repeated at least five times.
Effect of different concentration of oxamflatin on development of porcine SCNT embryos in vitro
After treatment of oxamflatin with 0.1 μM or 1 μM to SCNT embryos, cleavage rate was similar with that of control; however, blastocyst rate and total cell number in blastocyst of 1 μM oxamflatin-treated group (35.7±6.5 and 72.3±2.3, respectively) were increased significantly compared to those of control or 0.1 μM (13.2±4.7 and 50.6±3.5, 15.4±5.1, and 57.6±3.8, respectively) (Table 2).
Values are mean±SEM. Values with different superscripts in the same column differ significantly (p<0.05). Experiments were repeated at least five times.
Comparison of the effect between oxamflatin and scriptaid on development of porcine SCNT embryos in vitro
One micromolar oxamflatin significantly increased blastocyst rate and total cell number in blastocysts (33.3±6.0 and 73.1±1.6, respectively) than those of control (10.3±3.7 and 54.1±3.5), respectively, or scriptaid (16.4±4.6 and 64.4±2.1, respectively) (Table 3).
Values are mean±SEM. Values with different superscripts in the same column differ significantly (p<0.05). Experiments were repeated at least five times.
Gene expression profile among control, oxamflatin-, or scriptaid-treated SCNT embryos during in vitro development
To test whether the effects of oxamflatin on gene expression during preimplantation and development, the expression levels of pluripotency-related genes (POU5F1, NANOG, CDX2, REX01), histone deacetylase genes (HDAC1, HDAC2), and apoptosis-related genes (BCL2L1, BAX) were studied in control, oxamflatin-, or scriptaid-treated blastocysts using real-time PCR (Fig. 1). The expression levels of POU5F1, NANOG, REX01, and BCL2L1 were significantly higher in the oxamflatin-treated group than control or scriptiad-treated groups.
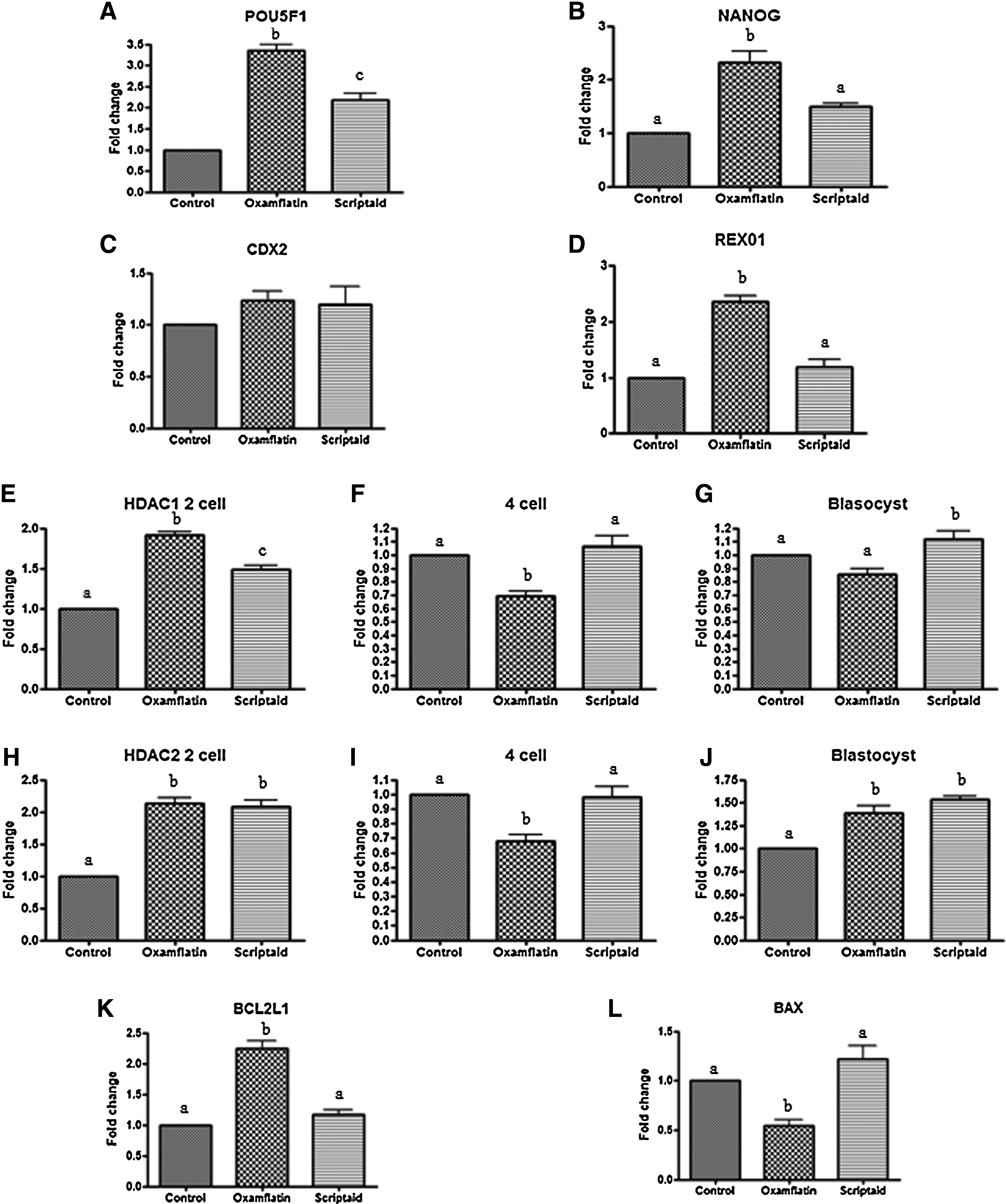
Expression profiles of POU5F1, NANOG, CDX2, REX01, HDAC1, HDAC2, BCL2L1, and BAX in SCNT embryos during in vitro development. Relative expression patterns of POU5F1 (
At the two-cell stage, the oxamflatin- or scriptaid-treated embryos showed significantly higher expression levels of HDAC1 and HDAC2 than control embryos. However, expression patterns of HDAC1 and HDAC2 in the oxamflatin-treated group was significantly decreased compared to those of the control or scriptaid-treated groups at the four-cell stages. HDAC1 expression in scriptaid-treated blastocysts was significantly higher than those in control or oxamflatin-treated blastocysts, whereas both scriptaid- and oxamflatin-treated blastocysts demonstrated significantly higher expression than the control. Oxamflatin-treated blastocysts showed a higher level of BCL2L1 and a lower level of BAX than counterparts (p<0.05).
Full-term development of SCNT embryos following oxamflatin or scriptaid
The SCNT embryos that were reconstructed with two white Yucatan miniature transgenic cell lines (SNU1 and SNU2) were transferred to 11 surrogates to investigate the effect of oxamflatin treatment on full-term development. Oxamflatin treatment resulted in a higher percentage of full-term development compared to the scriptaid-treated group. When the SNU1 cell line was used as the nuclear donor, all three surrogates became pregnant in the oxamflatin treatment group and one went to term with a cloning efficiency of 0.9% (Table 4); however, whereas 2 of 4 recipients became pregnant in the scriptaid treatment group, no live piglet was obtained. When SNU2 was the donor cell, 1 of 2 surrogates became pregnant and 3 piglets were born in the oxamflatin-treated group with a cloning efficiency of 3.2%, whereas 2 piglets were obtained from one litter in the scriptaid treatment group with a cloning efficiency of 1.8%.
Values with different superscripts in the same column differ significantly (P<0.05).
(+) Pregnant; (−) not pregnant.
Cloning efficiency=No. of piglets/No. of embryos transferred.
Discussion
Reprogramming following the transfer of somatic nuclei into oocyte cytoplasm occurs at the epigenetic level (Rybouchkin et al., 2006). Considering previous reports, insufficient and abnormal epigenetic modifications, including DNA methylation and histone acetylation, occur (Kang et al., 2001; Kishigami et al., 2006) during SCNT, and these seem to be associated with low success rate of cloning (Kishigami et al., 2007).
Increased histone acetylation by scriptaid could dramatically enhance the developmental competence of SCNT embryos in pig (Whitworth et al., 2011; Zhao et al., 2009; Zhao et al., 2010); however, low efficiency and cellular toxicity when used at effective concentration (Su et al., 2000) require further studies. A recent study in mice and cow showed that oxamflatin treatment (1 μM oxamflatin for 9 h after nuclear transfer) significantly improved the in vitro culture and full-term development of cloned mice (Ono et al., 2010; Su et al., 2011). In the present study, we investigated the effect of oxamflatin on the in vitro and in vivo development and compared with scrptaid-treated group. We found that oxamflatin treatment was superior to scriptaid treatment and enhanced the developmental capacity of reconstructed embryos both in vitro and in vivo.
We found that 1 μM of oxamflatin was effective and 10 μM of oxamflatin showed toxicity to the embryo; thus, a concentration of 1 μM was used for the further experiments. Compared to the control SCNT embryos, both the scriptaid- or oxamflatin-treated embryos indicated increased total cell number, although there was no difference of embryo quality as judged by blastocyst formation rate between control and scriptaid-treated groups. HDAC inhibitors, including scriptaid and oxamflatin, can induce hyperacetylation of the core histone, indicating structural alleviation in chromatin, the transcriptionally permissive state, as well as DNA methylation of donor cell–derived genome (Armstrong et al., 2006; Simonsson and Gurdon et al., 2004). Often reduced histone methylation and thus activation of genes that are crucial for embryo development are induced (Cervoni and Szyf et al., 2001).
The oxamflatin group significantly increased both blastocyst formation rate and total cell number compared to the scriptaid-treated group, whereas both HDACi-treated groups showed improvement of in vitro development than the control. The mechanism underlying how oxamflatin improves in vitro development and cloning efficiency remains unclear.
To elucidate the mechanism behind this, we measured the relative expression levels of the pluripotency-related genes (POU5F, NANOG, CDX2, REX01), histone deacetylase genes (HDAC1, HDAC2), and apoptosis-related genes (BCL2L1, BAX) in the control, oxamflatin-, and scriptaid-treated SCNT blastocysts using real-time PCR.
POU5F1, the homeodomain containing transcription factor, is a key regulator of pluripotency and cell differentiation (Nichols et al., 1998). It seems that POU5F1 plays an important role in determining early steps in embryogenesis and differentiation (Thompson and Gudas et al., 2002). POU5F1-deficient embryos lose their developmental pluripotency, and low POU5F1 levels result in differentiation into only the trophectoderm (Niwa et al., 2000). Expression of POU5F1 was lower in porcine SCNT blastocysts than in in vitro fertilized (IVF) counterparts (Kumar et al., 2007) and could be associated with low developmental competence of SCNT embryos. However, oxamflatin-treated SCNT blastocysts had higher POU5F1 expression levels than both control SCNT and scriptaid-treated blastocysts in this study. NANOG is also an important transcription factor for maintaining the pluripotent cells of the inner cell mass (ICM) (Chambers et al., 2003) and known to be regulated by POU5F1 (Hatano et al., 2005). Oxamflatin-treated SCNT blastocysts showed significantly higher expression levels than the control group, and this observation coincides with NANOG expression under POU5F1 regulation. REX01 is a developmentally regulated acidic zinc finger gene (Zfp-42) (Hosler et al., 1989) whose protein binds DNA and regulates transcription. REX01 expression is restricted to the ICM, the epiblast, and the endoderm (Talbot et al., 2007) in pig, whereas REX01 mRNA has been detected in the blastocyst stage, in trophoblast-derived tissues, and in meiotic germ cells in mice (Rogers et al., 1991). Our result shows the co-expression of POU5F1 and REX01 in the blastocyst, and considering the data that the REX01 promoter contains an octamer motif (for binding of POU family members), the level of expression of POU5F1 protein would influence on the REX01 promoter. From our results, we can conclude that oxamflatin treatment can correct abnormally reprogrammed porcine SCNT embryos through REX01 and NANOG controlled by POU5F1.
It is known that histone acetylation regulates transcription mediated throughout activities of HDACs and HATs. Dependence on expression of the embryonic genome cannot be detected until the mid two-cell stage in the mouse, the four-cell stage in the pig, and the eight-cell stage in the sheep (Braude et al., 1988). Embryonic gene activation (EGA) has been considered important evidence of reprogramming, which transforms the genome from transcriptional quiescence at fertilization to robust transcriptional activity. The same events are likely involved when the oocyte initiates the reprogramming process after SCNT. Reprogramming involves a series of molecular events that control gene expression and has a profound effect on nuclear architecture (Misteli, 2003). HDAC1 and HDAC2 were implicated as candidate genes involved in genome activation (Zeng and Schultz et al., 2005). McGraw et al. looked at the expression patterns for bovine HDAC1 from immature oocytes to blastocysts. The lowest level of mRNA of HDAC1 is at the eight-cell stage when EGA starts; however, HDAC1 is strongly expressed in the blastocyst stage (McGraw et al., 2003). Repression could be associated with structural changes in the chromatin, and the discovery that histone deacetylation can be associated with chromatin offers enlightenment regarding how histone deacetylation relates to repression of gene expression (Schultz et al., 1999). In a previous experiment, HDAC2 showed a low level of expression at the four-cell stage, with a dramatic decrease from the two-cell stage and an increase reaching a maximum at the blastocyst in IVF and in vivo porcine embryos, whereas steady changes were observed in nuclear transfer embryos (Kumar et al., 2007). In addition, there are reports that embryonic activated genes are suppressed (Suzuki et al., 2006; Vassena et al., 2007) and nucleolar reactivation is delayed in SCNT-generated embryos (Svarcova et al., 2009). We observed that oxamflatin treatment enhances in vitro development competence and cloning efficiency along with decreased HDAC1 and HDAC2 expression at the four-cell stage. Taken together, these findings suggest that HDACi treatment promotes the onset of EGA to relieve the incompetence of enucleated oocytes to induction of early embryonic stage-specific gene expression. Therefore, oxamflatin promoted normal EGA to overcome the delayed activation of nucleolar function in SCNT-generated embryos. We believe that oxamflatin, HDACi treatment had an effect on EGA in SCNT-generated cloned embryos to support a more accurate regulation of embryonic development.
The proapoptotic gene BAX regulates apoptosis, whereas the antiapoptotic gene BCL2L1 protects against apoptosis during embryo development. There was lower expression of BAX and higher expression of BCL2L1 in oxamflatin-treated blastocysts compared with those in scriptaid or control. Oxamflatin also suppresses apoptosis in bovine SCNT embryos (Su et al., 2011), indicating reduced apoptosis of cells in oxamflatin-treated blastocysts.
Embryo transfer results showed that oxamflatin treatment can produce viable cloned piglets and the development of SCNT embryos to the piglets after treatment with oxamflatin was twice than that of scriptaid-treated group.
Here, we focused on the application and optimization of the HDAC inhibitor oxamflatin in improving the cloning efficiency of pigs. The expression of development-related and histone acetylation genes subsequently improves nuclear reprogramming both the in vitro and in vivo development of porcine SCNT embryos. Further studies are necessary to elucidate the mechanism of oxamflatin treatment. In conclusion, the in vitro and in vivo developmental competence of SCNT pigs was significantly improved by oxamflatin treatment.
Footnotes
Acknowledgments
This study was supported by grants from MKE (#10033805, #10033839), BK21 for Veterinary Science and Hanwha L&C. The authors are grateful to Youn-Sang Kwon for technical assistance.
Author Disclosure Statement
The authors declare that no conflicting financial interests exist.
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.